
Biomass Characteristics of Tropical Montane Rain Forest in National Park of Hainan Tropical Rainforest

Abstract
1. Introduction
2. Study Area and Methodology
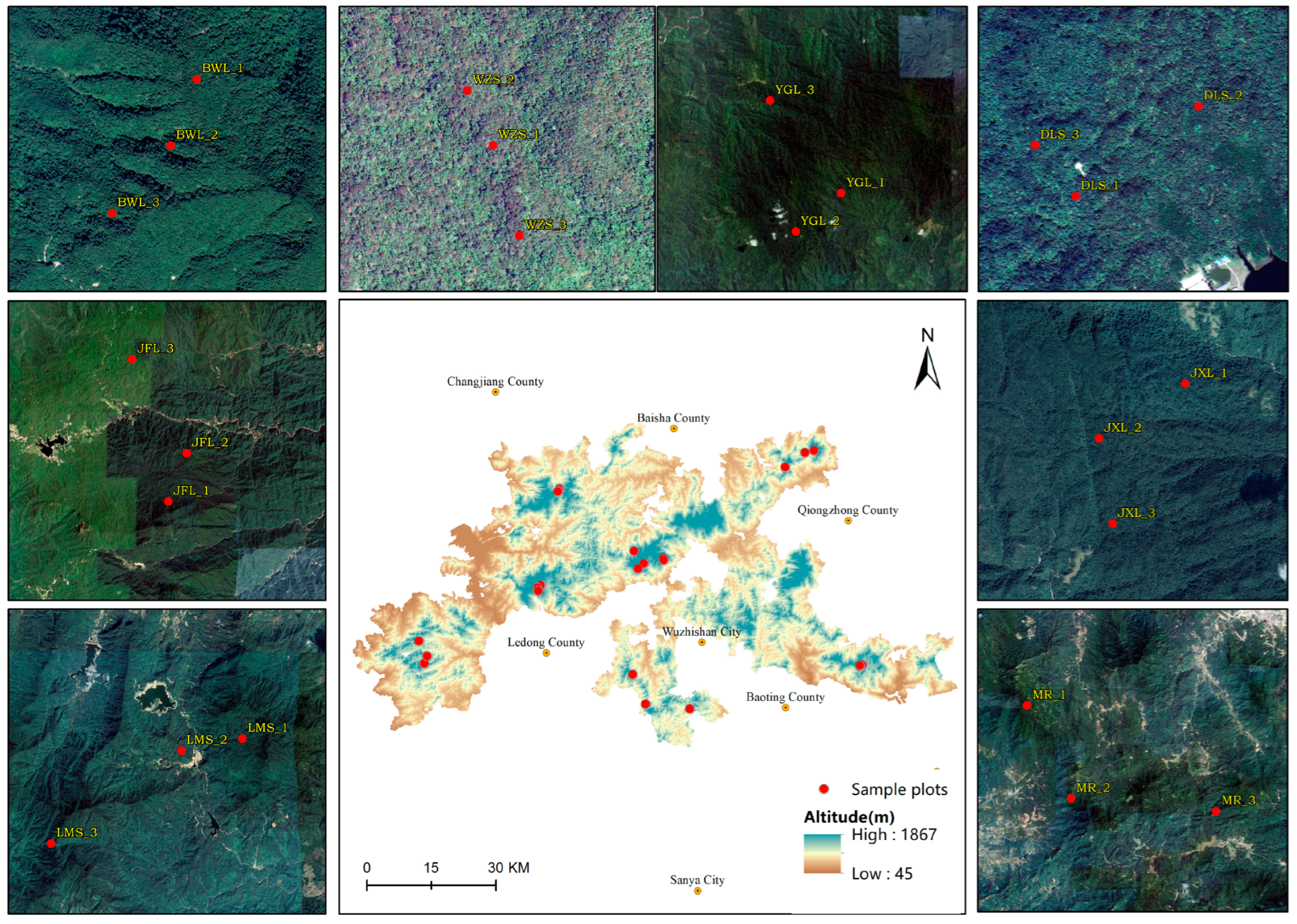
2.1. Overview of the Study Area
2.2. Data Source
2.3. Diameter Classification
2.4. Biomass Calculation
2.5. Spatial Distribution Simulation
3. Results and Analysis
3.1. Characteristics of Aboveground Biomass in Different Areas
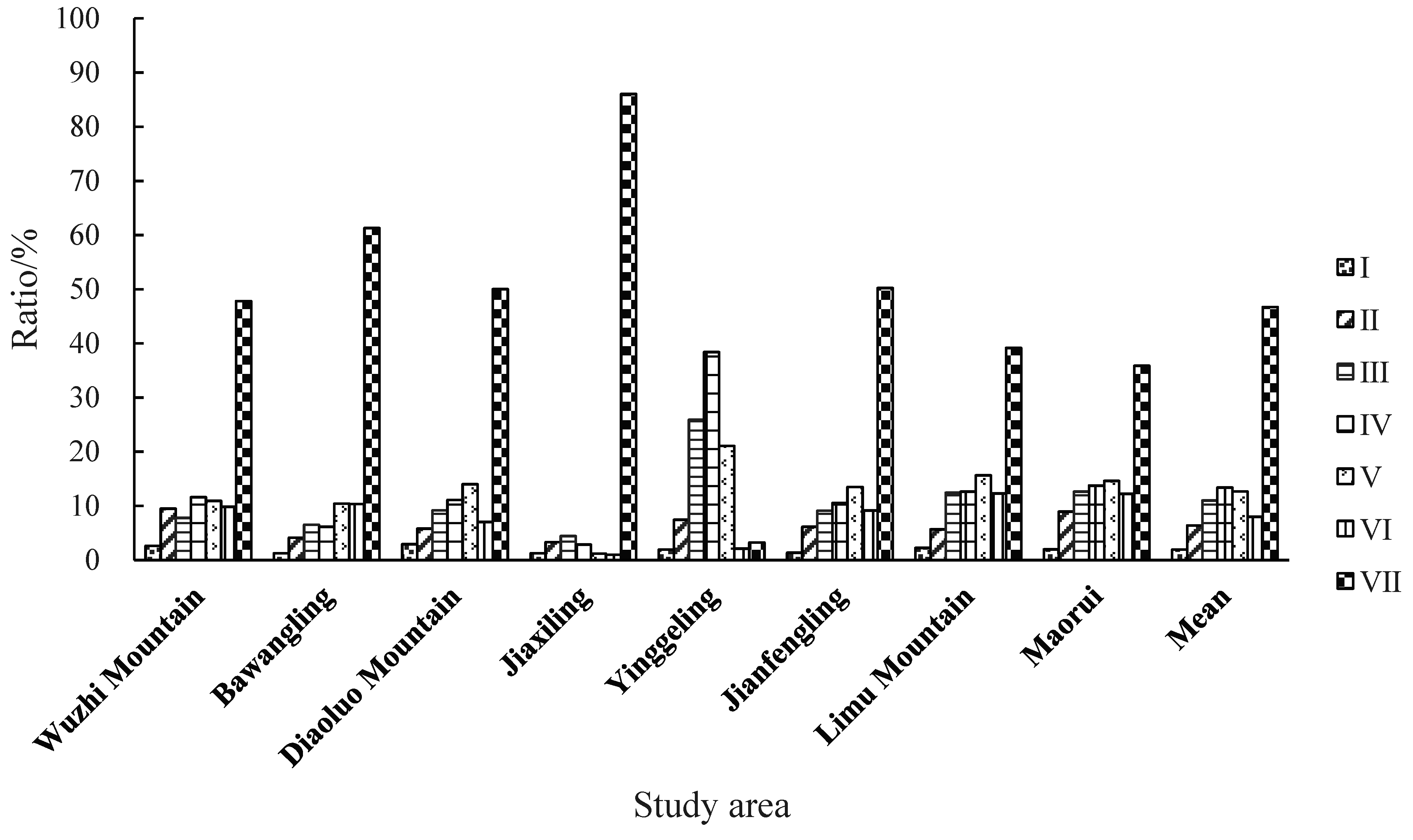
3.2. Distribution Pattern of Aboveground Biomass by Diameter Class
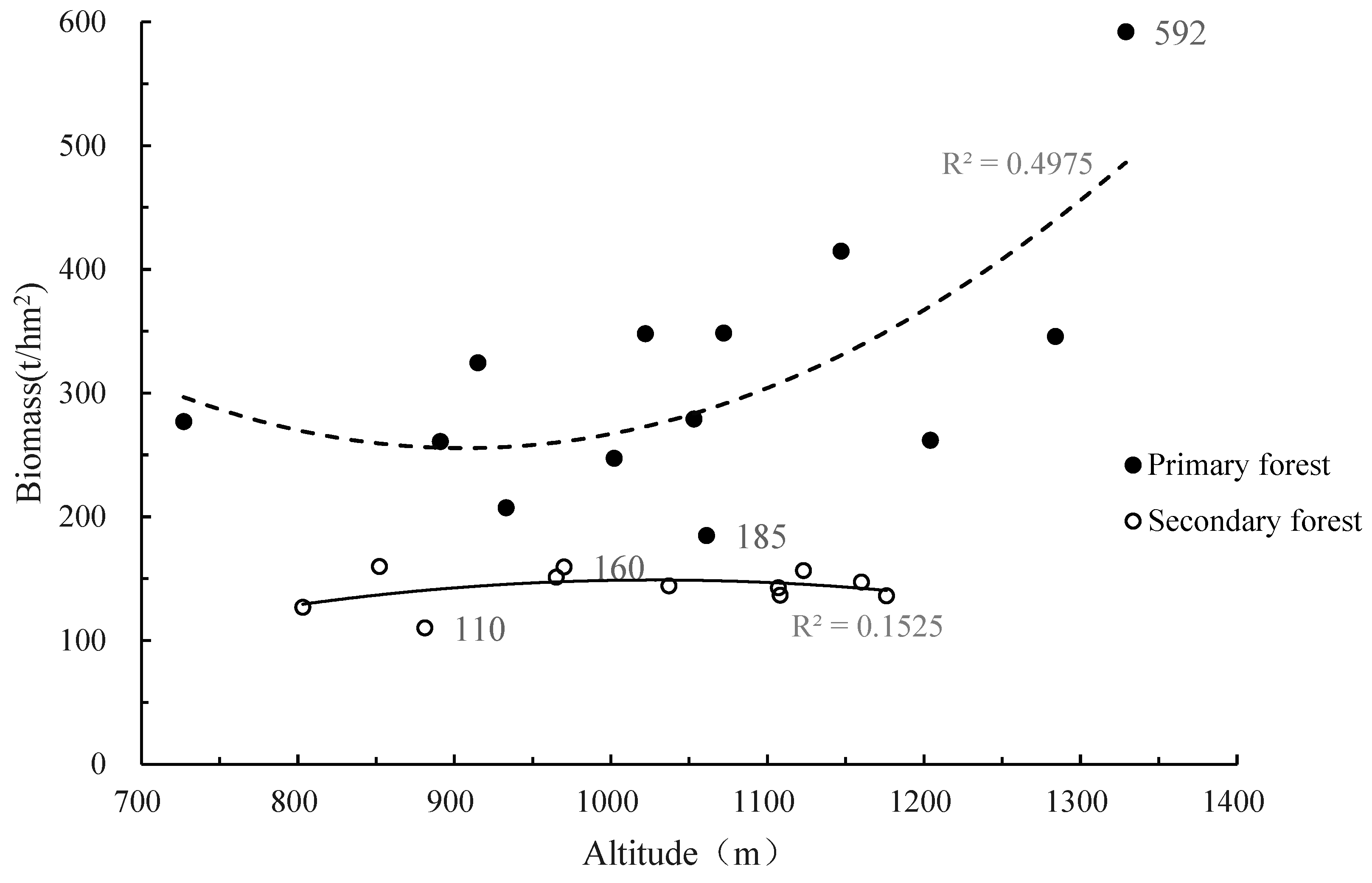
3.3. Distribution Pattern of Aboveground Biomass with Altitude
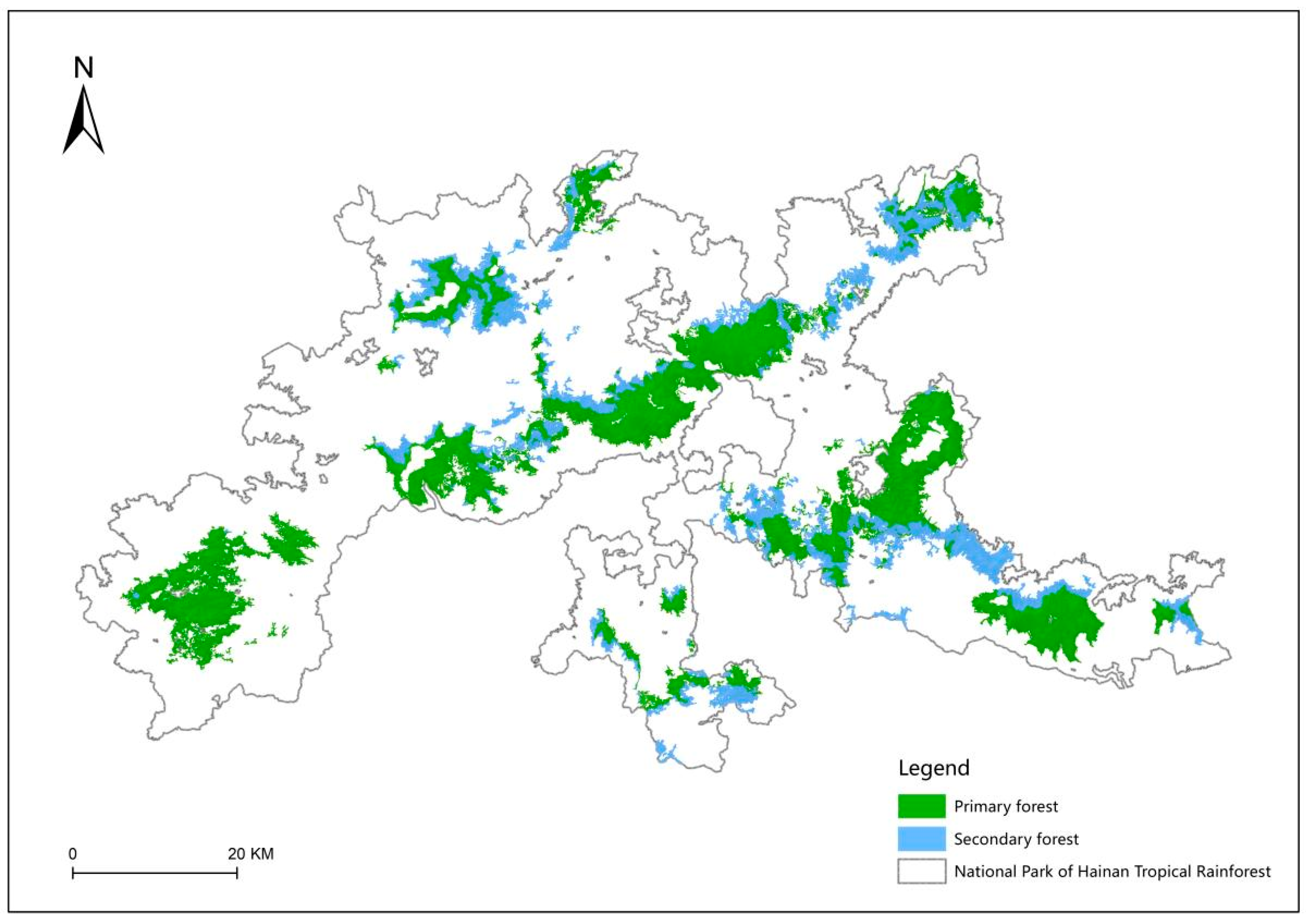
3.4. Characterization of the Spatial Distribution of Tropical Montane Rainforest
4. Discussion
4.1. Controlling Factors of Tropical Montane Rainforest Biomass in Hainan
4.2. Relationship Between Biomass in Tropical Montane Rainforests and in Other Tropical Forests
4.3. Uncertainty Analysis of Biomass Estimation
4.4. Prospect
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gatti, C.R.; Castaldi, S.; Lindsell, A.J.; Coomes, D.A.; Marchetti, M.; Maesano, M.; Di Paola, A.; Paparella, F.; Valentini, R. The impact of selective logging and clearcutting on forest structure, tree diversity and above-ground biomass of African tropical forests. Ecol. Res. 2015, 30, 119–132. [Google Scholar] [CrossRef]
- Blundo, C.; Carilla, J.; Grau, R.; Malizia, A.; Malizia, L.; Osinaga, A.O.; Bird, M.; Bradford, M.; Catchpole, D.; Ford, A.; et al. Taking the pulse of Earth’s tropical forests using networks of highly distributed plots. Biol. Conserv. 2021, 260, 108849. [Google Scholar]
- Schleussner, C.; Rogelj, J.; Schaeffer, M.; Lissner, T.; Licker, R.; Fischer, E.M.; Knutti, R.; Levermann, A.; Frieler, K.; Hare, W. Science and policy characteristics of the Paris Agreement temperature goal. Nat. Clim. Change 2016, 6, 827–835. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, A.R.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Avitabile, V.; Herold, M.; Heuvelink, G.B.; Lewis, S.L.; Phillips, O.L.; Asner, G.P.; Armston, J.; Ashton, P.S.; Banin, L.; Bayol, N.; et al. An integrated pan-tropical biomass map using multiple reference datasets. Glob. Change Biol. 2016, 22, 1406–1420. [Google Scholar] [CrossRef]
- Anja, R. The declining tropical carbon sink. Nat. Clim. Change 2021, 11, 727–728. [Google Scholar]
- Fan, L.; Wigneron, J.P.; Ciais, P.; Chave, J.; Brandt, M.; Fensholt, R.; Saatchi, S.S.; Bastos, A.; Al-Yaari, A.; Hufkens, K.; et al. Satellite-observed pantropical carbon dynamics. Nat. Plants. 2019, 5, 944–951. [Google Scholar] [CrossRef]
- Labrière, N.; Locatelli, B.; Vieilledent, G.; Kharisma, S.; Basuki, I.; Gond, V.; Laumonier, Y. Spatial congruence between carbon and biodiversity across forest landscapes of northern Borneo. Glob. Ecol. Conserv. 2016, 6, 105–120. [Google Scholar] [CrossRef]
- Chen, C.; Xiao, W.; Chen, H.Y.H. Meta-analysis reveals global variations in plant diversity effects on productivity. Nature 2025, 638, 1–6. [Google Scholar] [CrossRef]
- Patrick, L.T.; Sonia, K.; Yuval, R.Z.; Dee, L.E.; Wang, S.; De Mazancourt, C.; Loreau, M.; Gonzalez, A. Scaling up biodiversity–ecosystem functioning relationships: The role of environmental heterogeneity in space and time. Proc. R. Soc. B Biol. Sci. 2021, 288, 20202779. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, R.; Zhu, Z.; Xue, Y.; Ding, H.; Wang, X.; Na, J.; Xia, W. SCARF: A new algorithm for continuous prediction of biomass dynamics using machine learning and Landsat time series. Remote Sens. Environ. 2024, 314, 114348. [Google Scholar] [CrossRef]
- Lin, M.; Ling, Q.; Pei, H.; Song, Y.; Qiu, Z.; Wang, C.; Liu, T.; Gong, W. Remote Sensing of Tropical Rainforest Biomass Changes in Hainan Island, China from 2003 to 2018. Remote Sens. 2021, 13, 1696. [Google Scholar] [CrossRef]
- Abbas, S.; Wong, M.S.; Wu, J.; Shahzad, N.; Muhammad Irteza, S. Approaches of Satellite Remote Sensing for the Assessment of Above-Ground Biomass across Tropical Forests: Pan-tropical to National Scales. Remote Sens. 2020, 12, 3351. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Spaecies. Version 2013.1. Available online: http://www.iucnredlist.org (accessed on 2 July 2013).
- Zang, R.; Ding, Y. Forest recovery on abandoned logging roads in a tropical montane rain forest of Hainan Island, China. Acta Oecol. 2009, 35, 462–470. [Google Scholar] [CrossRef]
- Yang, X.B.; Chen, Z.Z.; Li, D.H. Classification and distribution of vegetation in Hainan, China. Sci. Sin. 2021, 51, 321–333. [Google Scholar] [CrossRef]
- Yang, X.B.; Chen, Z.Z.; Li, D.H. The Colored Illustrated Flora of Hainan Province; Science Press: Beijing, China, 2019. [Google Scholar]
- Zhu, Q.; Guo, H.; Zhang, L.; Liang, D.; Liu, X.; Wan, X.; Liu, J. Tropical Forests Classification Based on Weighted Separation Index from Multi-Temporal Sentinel-2 Images in Hainan Island. Sustainability 2021, 13, 13348. [Google Scholar] [CrossRef]
- Chiaka, C.J.; Yang, Q.; Zhao, Y.; Agostinho, F.; Almeida, C.M.; Giannetti, B.F.; Li, H.; Wu, M.; Liu, G. Assessment of Water-Related Ecosystem Services and Beneficiaries in the National Park of Hainan Tropical Rainforest. Land 2024, 13, 1804. [Google Scholar] [CrossRef]
- Wei, L.; Li, M.; Ma, Y.; Wang, Y.; Wu, G.; Liu, T.; Gong, W.; Mao, M.; Zhao, Y.; Wei, Y.; et al. Construction of an Ecological Security Pattern for the National Park of Hainan Tropical Rainforest on the Basis of the Importance of the Function and Sensitivity of Its Ecosystem Services. Land 2024, 13, 1618. [Google Scholar] [CrossRef]
- Li, Y.D. Comparative analysis for biomass measuerment of tropical mountain rain forest in Hainan Island, China. Acta Ecol. Sin. 1993, 4, 313–320. [Google Scholar]
- Li, Z.H.; He, W.; Cheng, M.F.; Hu, J.X.; An, X.; Huang, Y.; Yang, G.Y.; Zhang, H.Y. SinoLC-1: The first 1-meter resolution national-scale land-cover map of China created with the deep learning framework and open-access data. Zenodo 2023. [Google Scholar] [CrossRef]
- Cotillas, M.; Espelta, M.J.; Sánchez-Costa, E.; Sabaté, S. Aboveground and belowground biomass allocation patterns in two Mediterranean oaks with contrasting leaf habit: An insight into carbon stock in young oak coppices. Eur. J. For. Res. 2016, 135, 243–252. [Google Scholar] [CrossRef]
- Mcgarrigle, E.; Kershaw, J.A.; Lavigne, M.B.; Weiskittel, A.R.; Ducey, M. Predicting the number of trees in small diameter classes using predictions from a two-parameter Weibull distribution. Forestry 2011, 84, 431–439. [Google Scholar] [CrossRef]
- Kilak, H.S.; Alavi, J.S.; Esmailzadeh, O. Spatial resolution matters: Unveiling the role of environmental predictors in English yew (Taxus bacata L.) distribution using MaxEnt modeling. Earth Sci. Inform. 2025, 18, 214. [Google Scholar] [CrossRef]
- Wu, Y.L.; Li, J.Z.; Yang, Y.P.; Zhou, Z.X. Research advances in assessment of forest naturalness. Chin. J. Ecol. 2010, 29, 2065–2071. [Google Scholar]
- Brown, I.F.; Martinelli, L.A.; Thomas, W.W.; Moreira, M.Z.; Ferreira, C.C.; Victoria, R.A. Uncertainty in the biomass of Amazonian forests: An example from Rondônia Brazil. For. Ecol. Manag. 1995, 75, 175–189. [Google Scholar] [CrossRef]
- Tian, H.; Melillo, M.J.; Kicklighter, W.D.; McGuire, A.D.; Helfrich Iii, J.; Moore Iii, B.; Vörösmarty, C.J. Climatic and Biotic Controls on Annual Carbon Storage in Amazonian Ecosystems. Glob. Ecol. Biogeogr. 2000, 9, 315–335. [Google Scholar] [CrossRef]
- Zheng, Z.; Liu, H.M.; Feng, Z.L. Biomass of tropical montane rain forest in Xishuangbanna of Southwest China. Chin. J. Ecol. 2006, 4, 347–353. [Google Scholar]
- Hao, Q.Y.; Liu, Q.; Wang, S.Q.; Zhong, Q.; Wang, Y.; Ruan, C.; Yan, T.; Du, S.; Huang, Y. Biomass of Forest Communities at Different Altitude Regions. J. Trop. Subtrop. Bot. 2013, 21, 529–537. [Google Scholar]
- Piao, S.L. The Relationships Between Biodiversity and Ecosystem Functioning in Natural Tropical Forests of Hainan Island, China; Chinese Academy of Forestry: Beijing, China, 2013. [Google Scholar]
- Kitayama, K.; Aiba, S. Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu Borneo. J. Ecol. Eng. 2002, 90, 37–51. [Google Scholar] [CrossRef]
- Chen, P.; Yan, G.F.; Li, X.Y. The Evaluation on Effects of Natural Forest Protective Project (NFPP) in Hainan. For. Econ. 2007, 10, 52–57. [Google Scholar]
- Li, X.Y.; Li, H.L.; Zhu, Y.J. Empirical Analysis on Effects of Natural Forest Protection Project in Hainan Province. J. Beijing For. Univ. (Soc. Sci.) 2008, 3, 57–61. [Google Scholar]
- Chen, Y.F.; Qiao, T.; Lei, Y.C.; Chen, Q.; Wang, J. Analysis on Early Stage Trees Carbon Storage Change of Tropical MontaneRain Forest in Bawangling of Hainan Island. For. Res. 2013, 26, 337–343. [Google Scholar]
- Chen, D.X.; Li, Y.D.; Liu, H.P.; Xu, H.; Xiao, W.; Luo, T.; Zhou, Z.; Lin, M. Biomass and carbon dynamics of a tropical mountain rain forest in China. Sci. China Life Sci. 2010, 53, 798–810. [Google Scholar] [CrossRef]
- Li, Y.D.; Zeng, Q.B.; Wu, Z.M.; Du, Z.; Zhou, G.; Chen, B.; Zhang, Z.; Chen, H. Study on biomass of tropical mountain rain forest in Jianfengling, Hainan province. Chin. J. Plant Ecol. 1992, 4, 293–300. [Google Scholar]
- Wang, L.Y.; Zhou, Z.; Zhang, T.; Lin, M.X.; Zhang, C.S.; Li, Y.D.; Chen, D.X. Effects of tree species composition and diameter class structure on biomass restoration of secondary tropical forest. Plant Sci. J. 2022, 40, 169–176. [Google Scholar]
- Huang, Q.; Li, Y.D.; Lai, J.Z.; Peng, G. Study on biomass of tropical mountain rain forest in Limushan, Hainan island. Chin. J. Plant Ecol. 1991, 3, 197–206. [Google Scholar]
- Schuur, E.A.; Matson, P.A. Net primary productivity and nutrient cycling across a mesic to wet precipitation gradient in Hawaiian montane forest. Oecologia 2001, 128, 431–442. [Google Scholar] [CrossRef]
- Shao, X.L.; Chen, Y.K.; Wang, X.X.; Wang, X.; Wu, Y.; Hong, X.J.; Fang, Y.S.; Lu, Y.Q.; Long, W.X. Distribution patterns of aboveground biomass of tropical cloud forests in Hainan Island. Chin. J. Ecol. 2018, 37, 2566–2572. [Google Scholar]
- Brown, S. Estimating Biomass and Biomass Change of Tropical Forests: A Primer; FAO Forestry Paper No. 134; Forest Resources Assessment Publication: Rome, Italy, 1997; pp. 1–31. [Google Scholar]
- Brown, S.; Iverson, R.L.; Prasad, A.; Liu, D. Geographical distributions of carbon in biomass and soils of tropical Asian forests. Geocarto Int. 2008, 8, 45–59. [Google Scholar] [CrossRef]
- Keller, M.; Palace, M.; Hurtt, G. Biomass estimation in the Tapajos National Forest, Brazil: Examination of sampling and allometric uncertainties. Ecol. Manag. 2001, 154, 371–382. [Google Scholar] [CrossRef]
- Nebel, G.; Kvist, P.L.; Vanclay, K.J.; Christensen, H.; Freitas, L.; Ruíz, J. Structure and floristic composition of flood plain forests in the Peruvian Amazon I. Overstorey. For. Ecol. Manag. 2001, 150, 27–57. [Google Scholar] [CrossRef]
- Saldarriaga, J.G.; West, D.C.; Tharp, M.L.; Uhl, C. Long-term chronosequence of forest succession in the upper Rio Negro of Colombia and Venezuela. J. Ecol. 1988, 76, 938–958. [Google Scholar] [CrossRef]
- Brando, P.M.; Nepstad, D.C.; Davidson, E.A.; Trumbore, S.E.; Ray, D.; Camargo, P. Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest: Results of a throughfall reduction experiment. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1839–1848. [Google Scholar] [CrossRef] [PubMed]
- Nepstad, D.C.; Moutinho, P.; Dias-Filho, M.B.; Davidson, E.; Cardinot, G.; Markewitz, D.; Figueiredo, R.; Vianna, N.; Chambers, J.; Ray, D.; et al. The effects of partial throughfall exclusion on canopy processes, aboveground production, and biogeochemistry of an Amazon forest. J. Geophys. Res. 2002, 107, D20. [Google Scholar] [CrossRef]
- Long, W.X.; Zang, R.G.; Ding, Y. Community characteristics of tropical montane evergreen forest and tropical montane dwarf forest in Bawangling National Nature Reserve on Hainan Island, South China. Biodivers. Sci. 2011, 19, 558–566. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Geographic Location (°) | Sample Plot Area (m2) | Altitude (m) | Slope (°) | Number of Plants with DBH > 1 cm (Plants) | Dominant Tree Species | Forest Type |
|---|---|---|---|---|---|---|---|
| Wuzhi Mountain | 109°25.5′~109°25.6′ E, 18°56.9′~18°57.1′ N | 3× (30 m × 30 m) | 1083~1101 | 9~23 | 1152 | Pinus kwangtungensis, Dacrydium pectinatum, Castanopsis fissa, Syzygium araiocladum, Dendropanax hainanensis | Secondary forest |
| Bawangling | 109°11.5′~109°11.7′ E, 19°5.4′~19°5.8′ N | 3× (30 m × 30 m) | 1044~1160 | 12~21 | 1272 | Dacrydium pectinatum, Syzygium araiocladum, Quercus patelliformis, Dendropanax hainanensis, Lithocarpus amygdalifolius | Primary forest |
| Diaoluo Mountain | 109°51.7′~109°52.0′ E, 18°43.7′~18°43.8′ N | 3× (30 m × 30 m) | 961~1061 | 8~26 | 1161 | Semiliquidambar cathayensis, Lithocarpus amygdalifolius, Quercus championii, Daphniphyllum paxianum, Schima superba | Primary forest |
| Jiaxiling | 109°8.9′~109°9.3′ E, 18°52.9′~18°58.6′ N | 3× (30 m × 30 m) | 1204~1298 | 2~19 | 1246 | Dacrydium pectinatum Lithocarpus amygdalifolius, Syzygium araiocladum, Quercus patelliformis, Castanopsis carlesii, Castanopsis tonkinensis | Primary forest |
| Yinggeling | 109°21.42′~109°23.0′ E, 18°55.48′~18°58.0′ N | 3× (30 m × 30 m) | 803~1108 | 23~30 | 1253 | Castanopsis hainanensis, Lepisanthes rubiginosa, Pterospermum lanceifolium, Diospyros strigose | Secondary forest |
| Jianfengling | 108°53.1′~108°54.2′ E, 18°43.3′~18°46.3′ N | 3× (30 m × 30 m) | 915~1022 | 19~45 | 1314 | Lithocarpus fenzelianus, Quercus patelliformis, Gironniera subaequalis, Diospyros eriantha, Lithocarpus amygdalifolius | Primary forest |
| Limu Mountain | 109°41.4′~109°45.3′ E, 19°8.5′~19°10.5′ N | 3× (30 m × 30 m) | 727~1037 | 15~25 | 1068 | Dacrydium pectinatum, Dacrycarpus imbricatus, Quercus patelliformis, Maclurodendron oligophlebium, Canarium album | Secondary forest |
| Maorui | 109°21.4′~109°29.6′ E, 18°38.1′~18°42.3′ N | 3× (30 m × 30 m) | 852~1107 | 17~31 | 1145 | Lithocarpus corneus, Nephelium topengii, Canarium album, Alseodaphnopsis hainanensis, Xanthophyllum hainanense | Secondary forest |
| Variable Type | Variable Factor Abbreviation | Description |
|---|---|---|
| Climate variables | Tem | Annual mean temperature/°C |
| Pre | Annual mean precipitation/mm | |
| Topographic variables | alt | Altitude/m |
| slo | Slope/° | |
| Vegetation variables | Landuse | Land use type |
| NDVI | Normalized difference vegetation index/% | |
| NPP | Net primary productivity/J/(m2·a) | |
| Soil variables | soil | Soil type |
| K | K value of soil erodibility/% | |
| Human-induced ecosystem stress variables | pop | Nighttime light index/% |
| Area | Class | Sample Plot Aboveground Biomass (kg) | Aboveground Biomass per Hectare (t·hm−2) | Total Aboveground Biomass per Hectare (t·hm−2) | Forest Type | Naturalness |
|---|---|---|---|---|---|---|
| Wuzhi Mountain | DBH ≥ 5 cm | 38,568 | 142.84 | 146.69 | Secondary forest | Level III |
| DBH < 5 cm | 1037 | 3.84 | ||||
| Bawangling | DBH ≥ 5 cm | 92,653 | 343.16 | 347.50 | Primary forest | Level I |
| DBH < 5 cm | 1173 | 4.34 | ||||
| Diaoluo Mountain | DBH ≥ 5 cm | 51,706 | 191.50 | 197.28 | Primary forest + Secondary forest | Levels II & III |
| DBH < 5 cm | 1560 | 5.78 | ||||
| Jiaxiling | DBH ≥ 5 cm | 106,643 | 394.97 | 399.97 | Primary forest | Level I |
| DBH < 5 cm | 1349 | 5.00 | ||||
| Yinggeling | DBH ≥ 5 cm | 33,059.67 | 122.44 | 124.6 | Secondary forest | Levels III & IV |
| DBH < 5 cm | 583.2 | 2.16 | ||||
| Jianfengling | DBH ≥ 5 cm | 78,133 | 289.38 | 293.29 | Primary forest | Level II |
| DBH < 5 cm | 1057 | 3.91 | ||||
| Limu Mountain | DBH ≥ 5 cm | 50,904 | 188.53 | 190.86 | Primary forest + Secondary forest | Levels I, II & III |
| DBH < 5 cm | 629 | 2.33 | ||||
| Maorui | DBH ≥ 5 cm | 49,829 | 184.55 | 187.8 | Primary forest + Secondary forest | Levels II & III |
| DBH < 5 cm | 877 | 3.25 |
| Area | Wuzhi Mountain | Bawangling | Diaoluo Mountain | Jiaxiling | Yinggeling | Jianfengling | Limu Mountain | Maorui | Mean | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Biomass (t·hm−2) | |||||||||||
| Diameter | |||||||||||
| 1 ≤ DBH < 5 (Level I) | 3.84 | 4.34 | 5.78 | 5.00 | 2.38 | 3.99 | 4.28 | 3.66 | 4.16 | ||
| 5 ≤ DBH < 10 (Level II) | 13.93 | 14.31 | 11.40 | 13.13 | 9.27 | 18.01 | 10.80 | 16.77 | 13.45 | ||
| 10 ≤ DBH < 15 (Level III) | 11.39 | 22.57 | 18.08 | 17.72 | 32.23 | 26.72 | 23.72 | 23.74 | 22.02 | ||
| 15 ≤ DBH < 20 (Level IV) | 17.02 | 21.28 | 21.89 | 11.43 | 47.86 | 30.94 | 24.07 | 25.84 | 25.04 | ||
| 20 ≤ DBH < 25 (Level V) | 16.00 | 36.12 | 27.63 | 4.71 | 26.23 | 39.54 | 29.85 | 27.46 | 25.94 | ||
| 25 ≤ DBH < 30 (Level VI) | 14.40 | 35.87 | 13.84 | 3.82 | 2.63 | 26.75 | 23.42 | 22.97 | 17.96 | ||
| DBH ≥ 30 (Level VII) | 70.11 | 213.01 | 98.67 | 344.15 | 4.00 | 147.35 | 74.72 | 67.36 | 127.42 | ||
| Vegetation Classification | Elevation Zone (m) | Area (km2) | Aboveground Biomass (10,000 t) | Proportion of Total Area (%) |
|---|---|---|---|---|
| Secondary forest only | 600–700 | 9.08 | 13.08 | 3.09 |
| 700–800 | 84.23 | 121.29 | 28.70 | |
| 800–900 | 107.06 | 154.17 | 36.47 | |
| 900–1000 | 83.78 | 120.65 | 28.54 | |
| 1000–1100 | 7.52 | 10.82 | 2.56 | |
| 1100–1200 | 0.29 | 0.41 | 0.10 | |
| 1200–1300 | 1.58 | 2.27 | 0.54 | |
| Subtotal | 293.53 | 422.70 | 100.00 | |
| Primary forest only | 600–700 | 44.03 | 133.84 | 6.38 |
| 700–800 | 100.20 | 304.61 | 14.52 | |
| 800–900 | 129.77 | 394.50 | 18.80 | |
| 900–1000 | 133.17 | 404.84 | 19.30 | |
| 1000–1100 | 147.43 | 448.17 | 21.36 | |
| 1100–1200 | 88.15 | 267.99 | 12.77 | |
| 1200–1300 | 47.42 | 144.14 | 6.87 | |
| Subtotal | 690.17 | 2098.10 | 100.00 | |
| All tropical montane rainforest | 600–700 | 53.11 | 146.92 | 5.40 |
| 700–800 | 184.43 | 425.90 | 18.75 | |
| 800–900 | 236.83 | 548.67 | 24.08 | |
| 900–1000 | 216.95 | 525.49 | 22.05 | |
| 1000–1100 | 154.94 | 459.00 | 15.75 | |
| 1100–1200 | 88.44 | 268.40 | 8.99 | |
| 1200–1300 | 48.99 | 146.42 | 4.98 | |
| Total | 983.70 | 2520.80 | 100.00 |
| Area | This Study (t·hm−2) | Historical Research (t·hm−2) |
|---|---|---|
| Yinggeling | 124.60 | 152.60 |
| Bawangling | 347.50 | 373.10 |
| Jianfengling | 293.29 | 272.9–453.13 |
| Diaoluo Mountain | 190.86 | 160.39 ± 42.13 |
| Limu Mountain | 197.28 | 507.24 |
| Mean | 236.00 | 311.27 |
| Area | Aboveground Biomass (t·hm−2) |
|---|---|
| Tropical montane rainforests in Xishuangbanna, Yunnan | 249.6 |
| Tropical montane rainforests in Borneo, Southeast Asia | 294 |
| Tropical montane rainforests in Hawaii | 217.5~250.3 |
| Tropical montane rainforests in Hainan | 236 |
| Monsoon rainforests in Hainan | 125.8 |
| Tropical cloud forests in Hainan | 164.57 |
| Lowland rainforest secondary forests in Hainan | 137.91 ± 31.02 |
| Tropical moist rainforests in Cambodia | 295 |
| Tropical moist rainforests in Malaysia | 338 |
| Tropical rainforests in Brazil | 264 |
| Tropical forests in Peru | 275.32~279.64 |
| Tropical forests in Venezuela | 234 ± 22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.; Chen, Z.; Chen, Y.; Chen, Y.; Lei, J.; Chen, X.; Li, Y.; Pan, X. Biomass Characteristics of Tropical Montane Rain Forest in National Park of Hainan Tropical Rainforest. Land 2025, 14, 608. https://doi.org/10.3390/land14030608
Wu T, Chen Z, Chen Y, Chen Y, Lei J, Chen X, Li Y, Pan X. Biomass Characteristics of Tropical Montane Rain Forest in National Park of Hainan Tropical Rainforest. Land. 2025; 14(3):608. https://doi.org/10.3390/land14030608
Chicago/Turabian StyleWu, Tingtian, Zongzhu Chen, Yiqing Chen, Yukai Chen, Jinrui Lei, Xiaohua Chen, Yuanling Li, and Xiaoyan Pan. 2025. "Biomass Characteristics of Tropical Montane Rain Forest in National Park of Hainan Tropical Rainforest" Land 14, no. 3: 608. https://doi.org/10.3390/land14030608
APA StyleWu, T., Chen, Z., Chen, Y., Chen, Y., Lei, J., Chen, X., Li, Y., & Pan, X. (2025). Biomass Characteristics of Tropical Montane Rain Forest in National Park of Hainan Tropical Rainforest. Land, 14(3), 608. https://doi.org/10.3390/land14030608