
Spatiotemporal Dynamics in Bird Species Assembly in the Coastal Wetlands of Sicily (Italy): A Multilevel Analytical Approach to Promote More Satisfactory Conservation Planning


Abstract
1. Introduction
2. Materials and Methods
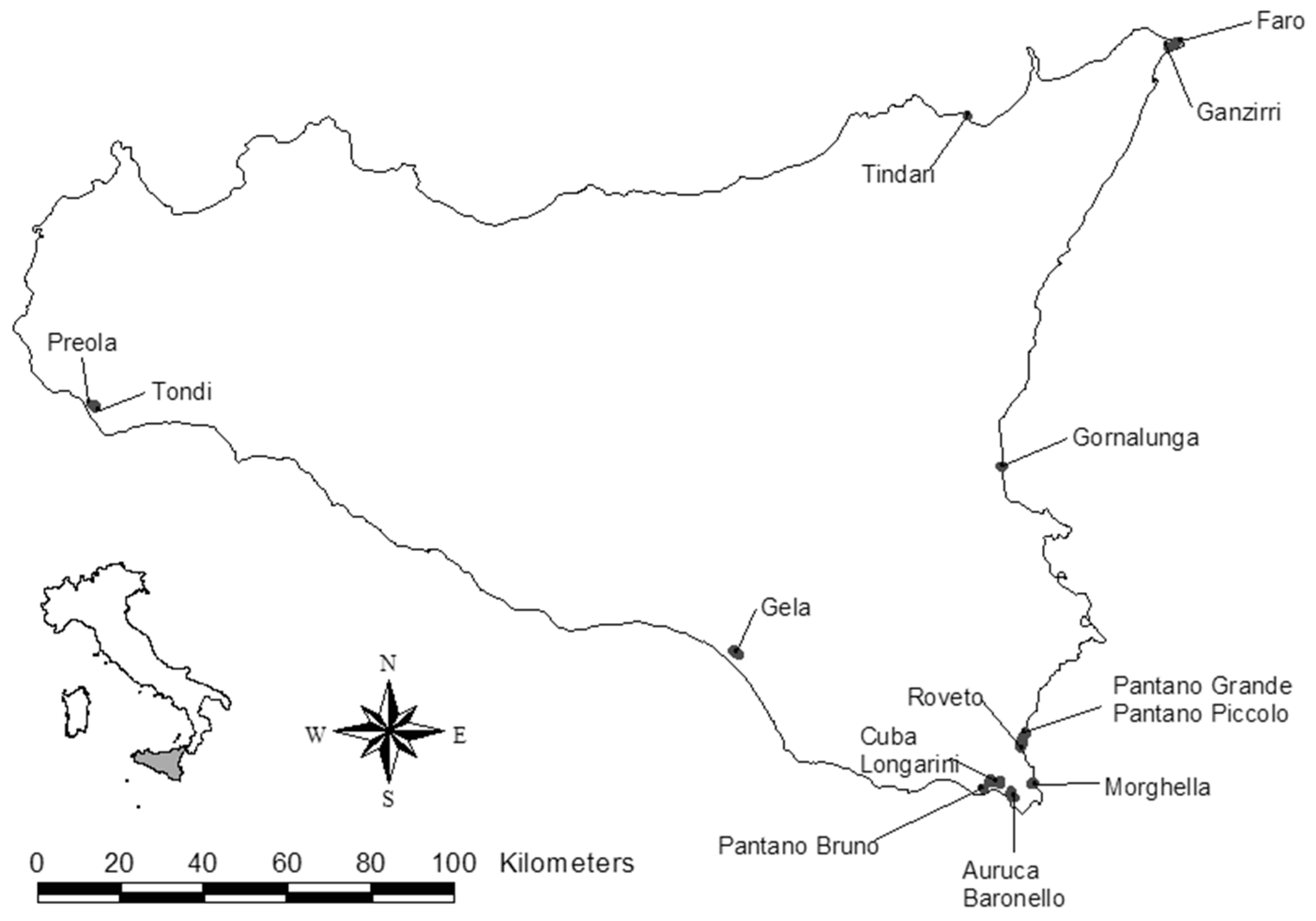
2.1. Study Area and Field Surveys
2.2. Data Analyses
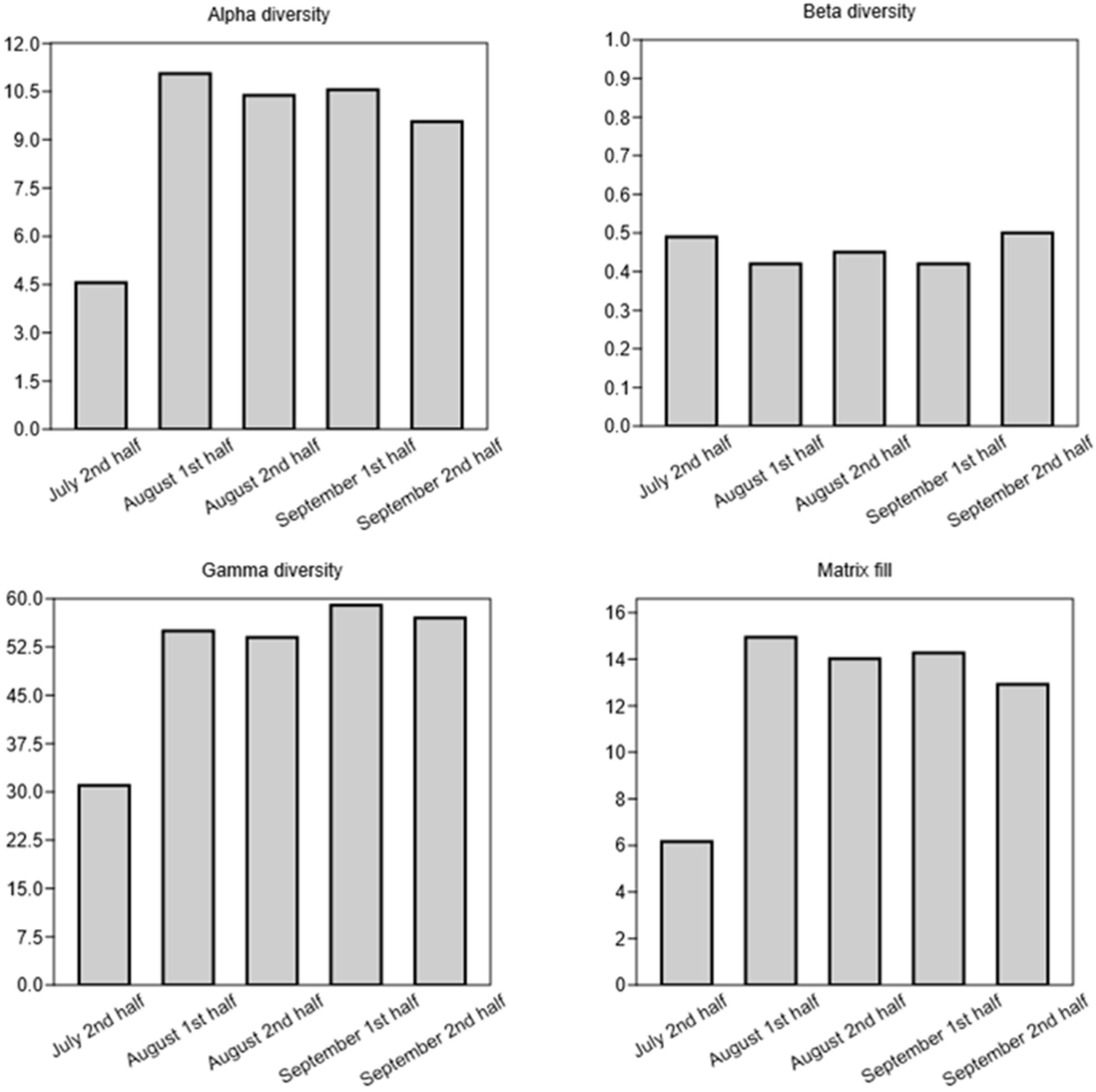
2.2.1. Metacommunity Level
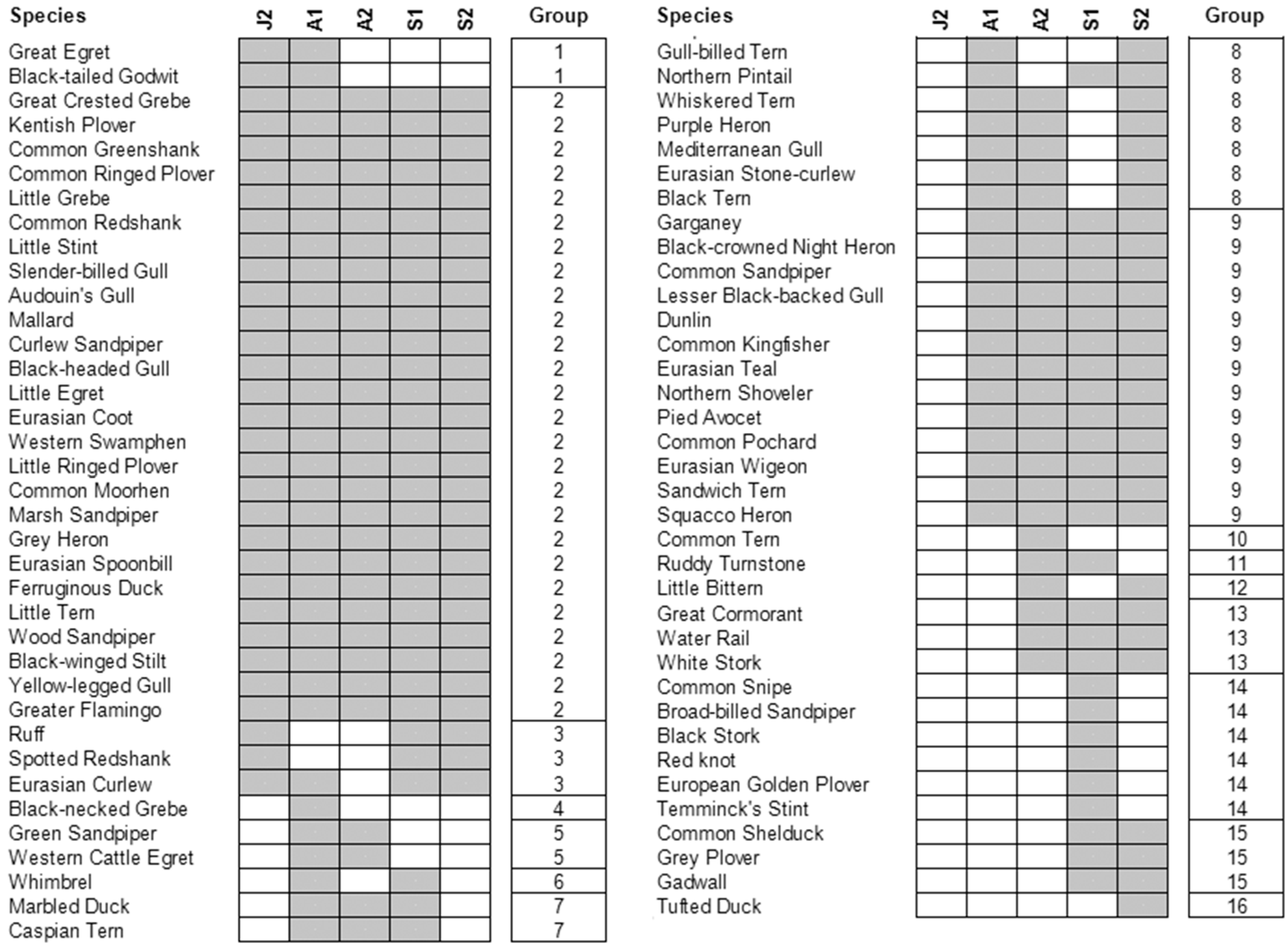
2.2.2. Community Level
2.2.3. Guild Level
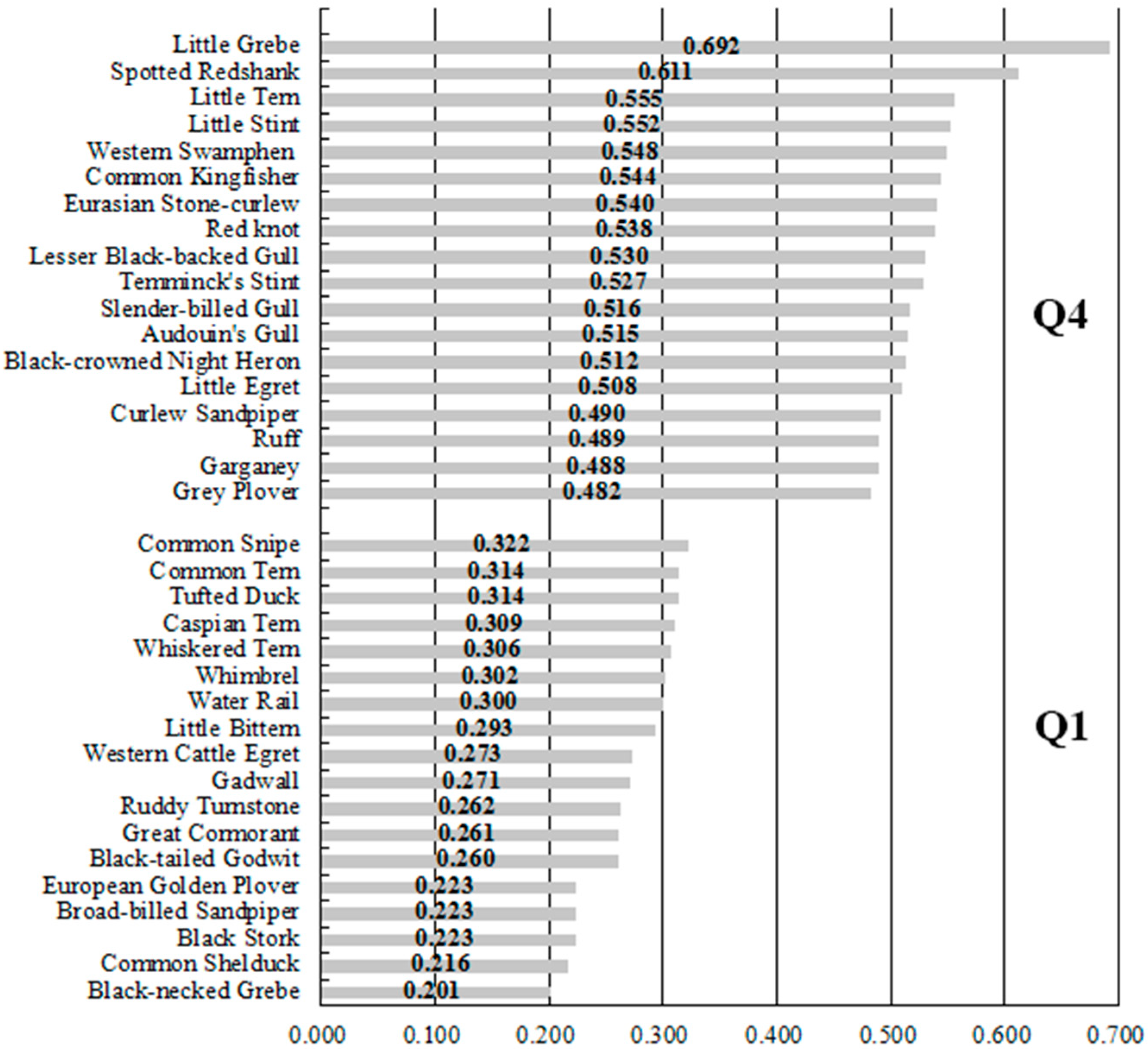
2.2.4. Species Level
3. Results
4. Discussion
Implications for the Conservation of the Waterbird Species of the Sicilian Wetlands
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gardner, R.C.; Barchiesi, S.; Beltrame, C.; Finlayson, C.M.; Galewski, T.; Harrison, I.J.; Paganini, M.; Perennou, C.; Rosenqvist, A.; Walpole, M.; et al. State of the World’s Wetlands and Their Services to People: A Compilation of Recent Analyses; Social Science Electronic Publishing: Gland, Switzerland, 2015. [Google Scholar]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; John Wiley & Sons: New York, NY, USA, 2015. [Google Scholar]
- Gardner, R.C.; Finlayson, C.M. Global Wetland Outlook: State of the World’s Wetlands and Their Services to People; Ramsar Convention: Gland, Switzerland, 2018. [Google Scholar]
- Fraser, L.H.; Keddy, P.A. The World’s Largest Wetlands: Ecology and Conservation; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Costanza, R.; De Groot, R.; Sutton, P.; Van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Davidson, N.C.; Dam, A.V.; Finlayson, C.M.; Mcinnes, R.J. Worth of wetlands: Revised global monetary values of coastal and inland wetland ecosystem services. Mar. Freshw. Res. 2019, 70, 1189. [Google Scholar] [CrossRef]
- Davidson, N.C. How Much Wetland Has the World Lost? Long-Term and Recent Trends in Global Wetland Area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Erwin, K.L. Wetlands and global climate change: The role of wetland restoration in a changing world. Wetl. Ecol. Manag. 2009, 17, 71–84. [Google Scholar] [CrossRef]
- Ferrarini, A.; Celada, C.; Gustin, M. Preserving the Mediterranean bird flyways: Assessment and prioritization of 38 main wetlands under human and climate threats in Sardinia and Sicily (Italy). Sci. Total Environ. 2020, 751, 141556. [Google Scholar] [CrossRef]
- Barlow, P.M.; Reichard, E.G. Saltwater intrusion in coastal regions of North America. Hydrogeol. J. 2010, 18, 247–260. [Google Scholar] [CrossRef]
- Rodríguez-Santalla, I.; Navarro, N. Main Threats in Mediterranean Coastal Wetlands. The Ebro Delta Case. J. Mar. Sci. Eng. 2021, 9, 1190. [Google Scholar] [CrossRef]
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef]
- Spina, F.; Volponi, S. Atlante della Migrazione Degli Uccelli in Italia; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale: Rome, Italy, 2008. [Google Scholar]
- Bijlsma, R.G. Bottleneck Areas for Migratory Birds in the Mediterranean Region: An Assessment of the Problems and Recommendations for Action; International Council for Bird Preservation: Cambridge, UK, 1990. [Google Scholar]
- Heath, M.F.; Evans, M.I. (Eds.) Important Bird Areas in Europe. Priority Sites for Conservation; BirdLife International: Cambridge, UK, 2000; Volume 2. [Google Scholar]
- Ferrarini, A.; Celada, C.; Gustin, M. Anthropogenic Pressure and Climate Change Could Severely Hamper the Avian Metacommunity of the Sicilian Wetlands. Diversity 2022, 14, 696. [Google Scholar] [CrossRef]
- Ferrarini, A.; Celada, C.; Gustin, M. Waterbird Species are Highly Sensitive to Wetland Traits: Simulation-Based Conservation Strategies for the Birds of the Sicilian Wetlands (Italy). Biology 2024, 13, 242. [Google Scholar] [CrossRef]
- Ferrarini, A.; Gustin, M.; Celada, C. Twenty-three years of land-use changes induced considerable threats to the main wetlands of Sardinia and Sicily (Italy) along the Mediterranean bird flyways. Diversity 2021, 13, 240. [Google Scholar] [CrossRef]
- Griffith, D.A. Spatial Autocorrelation: A Primer; Association of American Geographers: Washington, DC, USA, 1987. [Google Scholar]
- Hutto, R.L.; Pletschet, S.M.; Hendricks, P. A fixed-radius point count method for nonbreeding and breeding season use. Auk 1986, 103, 593–602. [Google Scholar] [CrossRef]
- Cramp, S.E.; Simmons, K.E.L.; Brooks, D.J.; Perrins, C.M. Handbook of the Birds of Europe, the Middle East and North Africa: The Birds of the Western Palearctic; Oxford University Press: Oxford, UK, 1977–1994; Volume 1–9. [Google Scholar]
- Routledge, R.D. On Whittaker’s components of diversity. Ecology 1977, 38, 1120–1127. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods, 4th ed.; Charles Griffin: London, UK, 1975. [Google Scholar]
- Mann, H.B. Non-parametric tests against trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Sokal, R.R.; Sneath, P.H.A. Principles of Numerical Taxonomy; W. H. Freeman: San Francisco, CA, USA, 1963. [Google Scholar]
- Brower, J.C.; Kile, K.M. Seriation of an original data matrix as applied to palaeoecology. Lethaia 1988, 21, 79–93. [Google Scholar] [CrossRef]
- Gower, J.C. Statistical methods of comparing different multivariate analyses of the same data. In Mathematics in the Archaeological and Historical Sciences; Hodson, F.R., Kendall, D.G., Tautu, P., Eds.; Edinburgh University Press: Edinburgh, UK, 1971; pp. 138–149. [Google Scholar]
- Jackson, D.A. PROTEST: A procrustean randomization test of community environment concordance. Ecoscience 1995, 2, 297–303. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.; Yang, R.; Li, D.; Qiu, Y.; Lu, K.; Cao, X.; Chen, Q. Spatiotemporal Dynamics of Constructed Wetland Landscape Patterns during Rapid Urbanization in Chengdu, China. Land 2024, 13, 806. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, S.; Tian, G.; Dong, N.; Lei, Y. Coupling Biodiversity and Human Pressures to Indicate Conservation Priorities for Threatened Waterfowl Species: A Case in the Henan Yellow River Wetland National Nature Reserve. Land 2023, 12, 1250. [Google Scholar] [CrossRef]
- Hammana, C.; Pereña-Ortiz, J.F.; Meddad-Hamza, A.; Hamel, T.; Salvo-Tierra, Á.E. The Wetlands of Northeastern Algeria (Guelma and Souk Ahras): Stakes for the Conservation of Regional Biodiversity. Land 2024, 13, 210. [Google Scholar] [CrossRef]
- Sciandrello, S.; Ranno, V.; Tomaselli, V. The Role of Vegetation Monitoring in the Conservation of Coastal Habitats N2000: A Case Study of a Wetland Area in Southeast Sicily (Italy). Land 2024, 13, 62. [Google Scholar] [CrossRef]
- Zhang, J.; Qin, Y.; Zhang, Y.; Lu, X.; Cao, J. Comparative Assessment of the Spatiotemporal Dynamics and Driving Forces of Natural and Constructed Wetlands in Arid and Semiarid Areas of Northern China. Land 2023, 12, 1980. [Google Scholar] [CrossRef]
- Brittain, R.A.; Craft, C.B. Effects of sea-level rise and anthropogenic development on priority bird species habitats in coastal Georgia, USA. Environ. Manag. 2012, 49, 473–482. [Google Scholar] [CrossRef]
- Xu, Q.; Zhou, L.; Xia, S.; Zhou, J. Impact of Urbanisation Intensity on Bird Diversity in River Wetlands around Chaohu Lake, China. Animals 2022, 12, 473. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.M.; Adam, P. Climate Change and Intertidal Wetlands. Biology 2013, 2, 445–480. [Google Scholar] [CrossRef] [PubMed]
- Blake-Bradshaw, A.G.; Lancaster, J.D.; O’Connell, J.R.; Matthews, J.W.; Eichholz, M.W.; Hagy, H.M. Suitability of Wetlands for Migrating and Breeding Waterbirds in Illinois. Wetlands 2020, 40, 1993–2010. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.Y.; Li, Y.F.; Dong, B.; Qiu, C.Q.; Yang, J.L.; Zong, Y.; Chen, H.; Zhao, Y.Q.; Zhang, Y.A. Study on habitat suitability and environmental variable thresholds of rare waterbirds. Sci. Total Environ. 2021, 785, 147316. [Google Scholar] [CrossRef]
- Burfield, I.J.; Rutherford, C.A.; Fernando, E.; Grice, H.; Piggott, A.; Martin, R.W.; Balman, M.; Evans, M.I.; Staneva, A. Birds in Europe 4: The fourth assessment of Species of European Conservation Concern. Bird Conserv. Int. 2023, 33, e66. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date of Sampling | Mediterranean | Mixed | Sedentary | Trans-Saharan |
|---|---|---|---|---|
| July (2nd half) | 8 | 7 | 5 | 11 |
| August (1st half) | 13 | 18 | 5 | 19 |
| August (2nd half) | 13 | 17 | 5 | 19 |
| September (1st half) | 13 | 20 | 5 | 21 |
| September (2nd half) | 14 | 19 | 5 | 19 |
| S: | 7 | 6 | 0 | 5 |
| p (no trend): | 0.042 | 0.117 | 0.592 | 0.117 |
| Statistically significant? | yes | no | no | no |
| Date of Sampling | Divers from Flight | Divers from the Surface | Intermediate Waders | Large Waders | Small Waders | Surface Feeders |
|---|---|---|---|---|---|---|
| July (2nd half) | 1 | 3 | 6 | 7 | 6 | 8 |
| August (1st half) | 7 | 5 | 9 | 9 | 9 | 16 |
| August (2nd half) | 7 | 5 | 8 | 8 | 10 | 16 |
| September (1st half) | 4 | 5 | 10 | 9 | 13 | 18 |
| September (2nd half) | 6 | 6 | 10 | 9 | 8 | 18 |
| S: | 1 | 7 | 6 | 5 | 4 | 8 |
| p (no trend): | 0.408 | 0.042 | 0.117 | 0.117 | 0.242 | 0.042 |
| Statistically significant? | no | yes | no | no | no | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrarini, A.; Celada, C.; Gustin, M. Spatiotemporal Dynamics in Bird Species Assembly in the Coastal Wetlands of Sicily (Italy): A Multilevel Analytical Approach to Promote More Satisfactory Conservation Planning. Land 2024, 13, 1333. https://doi.org/10.3390/land13081333
Ferrarini A, Celada C, Gustin M. Spatiotemporal Dynamics in Bird Species Assembly in the Coastal Wetlands of Sicily (Italy): A Multilevel Analytical Approach to Promote More Satisfactory Conservation Planning. Land. 2024; 13(8):1333. https://doi.org/10.3390/land13081333
Chicago/Turabian StyleFerrarini, Alessandro, Claudio Celada, and Marco Gustin. 2024. "Spatiotemporal Dynamics in Bird Species Assembly in the Coastal Wetlands of Sicily (Italy): A Multilevel Analytical Approach to Promote More Satisfactory Conservation Planning" Land 13, no. 8: 1333. https://doi.org/10.3390/land13081333
APA StyleFerrarini, A., Celada, C., & Gustin, M. (2024). Spatiotemporal Dynamics in Bird Species Assembly in the Coastal Wetlands of Sicily (Italy): A Multilevel Analytical Approach to Promote More Satisfactory Conservation Planning. Land, 13(8), 1333. https://doi.org/10.3390/land13081333