


Potential Interactions between Climate Change and Land Use for Forest Issues in the Eastern United States



Highlights
- We examined climate change and land use and disturbance changes in the eastern U.S.
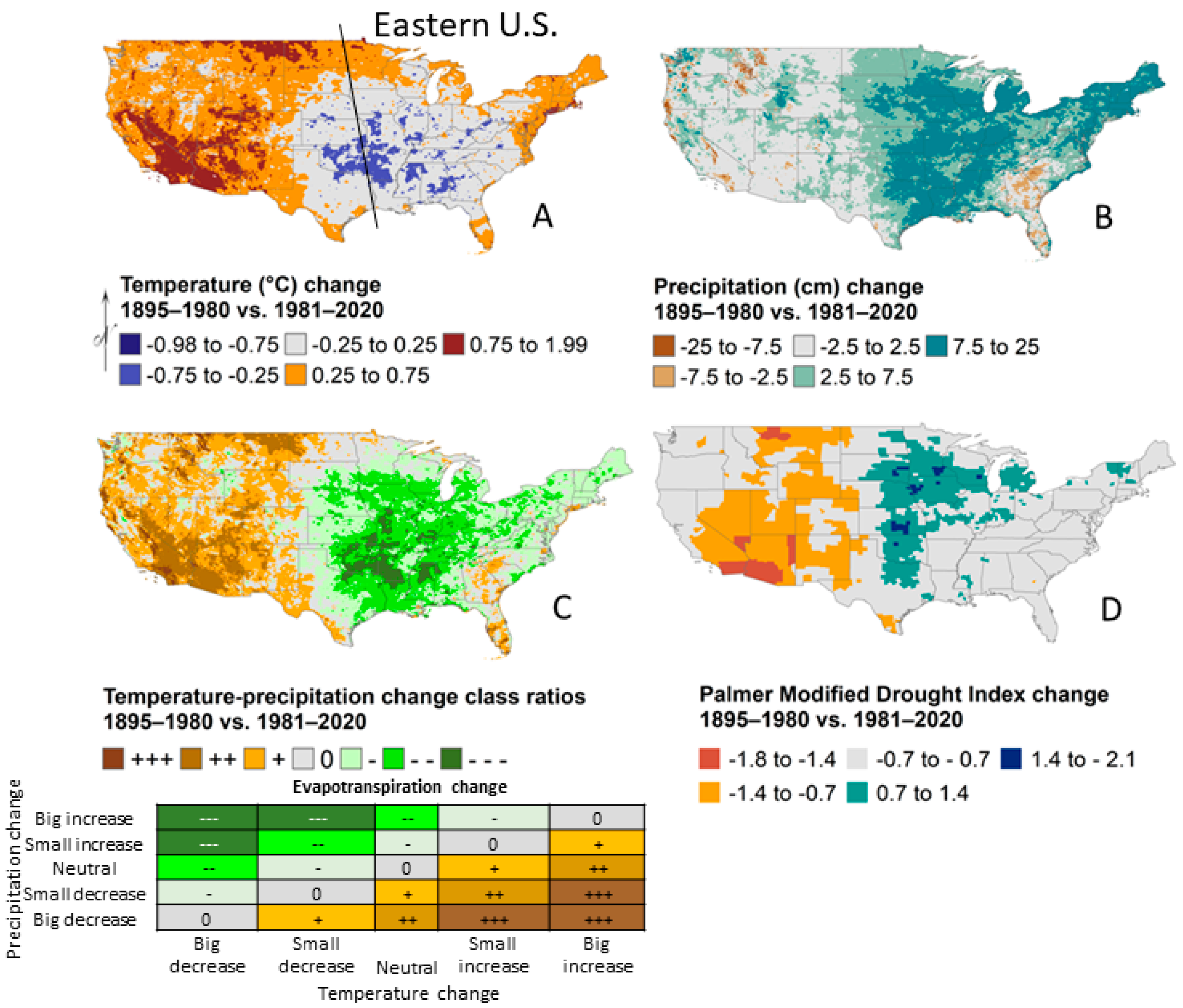
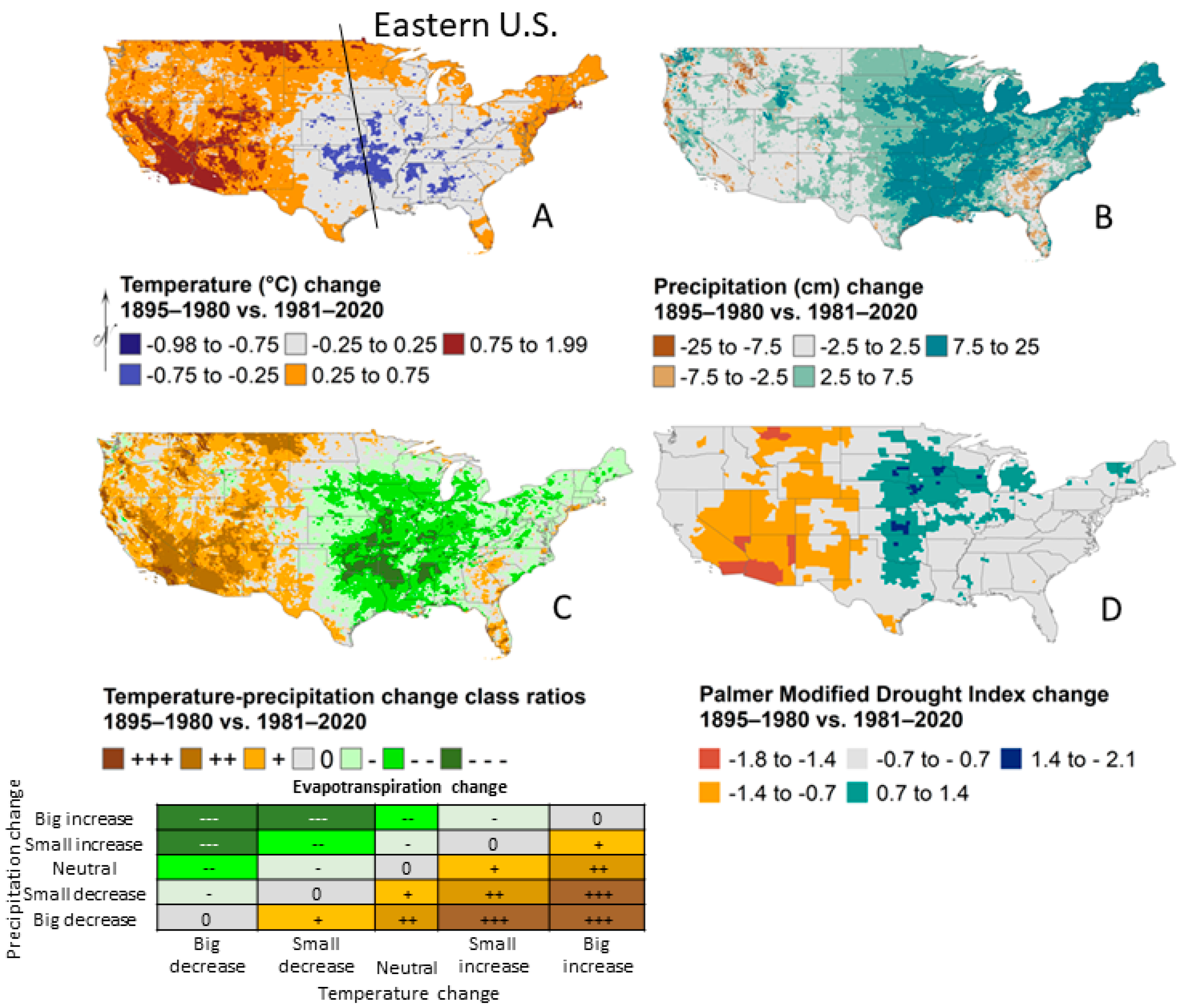
- Climate has not warmed but precipitation has increased.
- Climate and land use have interacted to reduce wildfire frequency and increase tree growth.
- Human activities facilitated the expansion of native tree species distribution, non-native species invasion, and damaging native species outbreaks.
- Recent climate change and land use have not influenced deer herbivory levels.
- A warmer and drier climate may reverse interactions with land use, varying by species.
- Management can correct non-climate stressors and support ecosystems against climate change.
Abstract

1. Introduction to Climate Change and Land Use
- What are the dynamics of wildfire in response to climate and land use?
- What are the dynamics of tree growth and distributions in response to climate and land use?
- What are the dynamics of non-native species, damaging native insects and disease, and white-tailed deer (Odocoileus virginianus, one of the last remaining widespread large herbivores) in response to climate and land use?
- How can ecological and adaptive management mitigate negative impacts of climate change and land use to maintain forest resiliency?
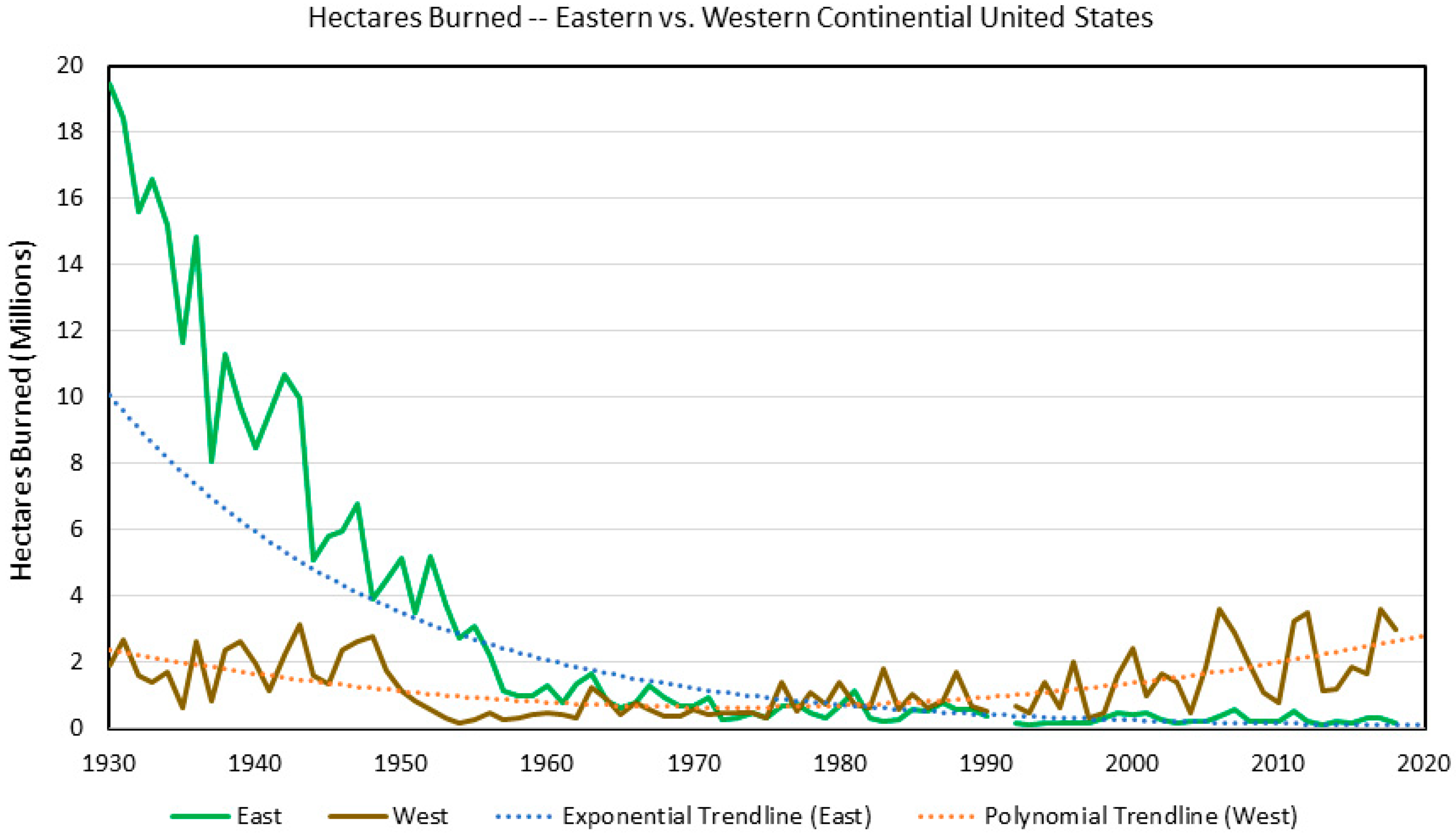
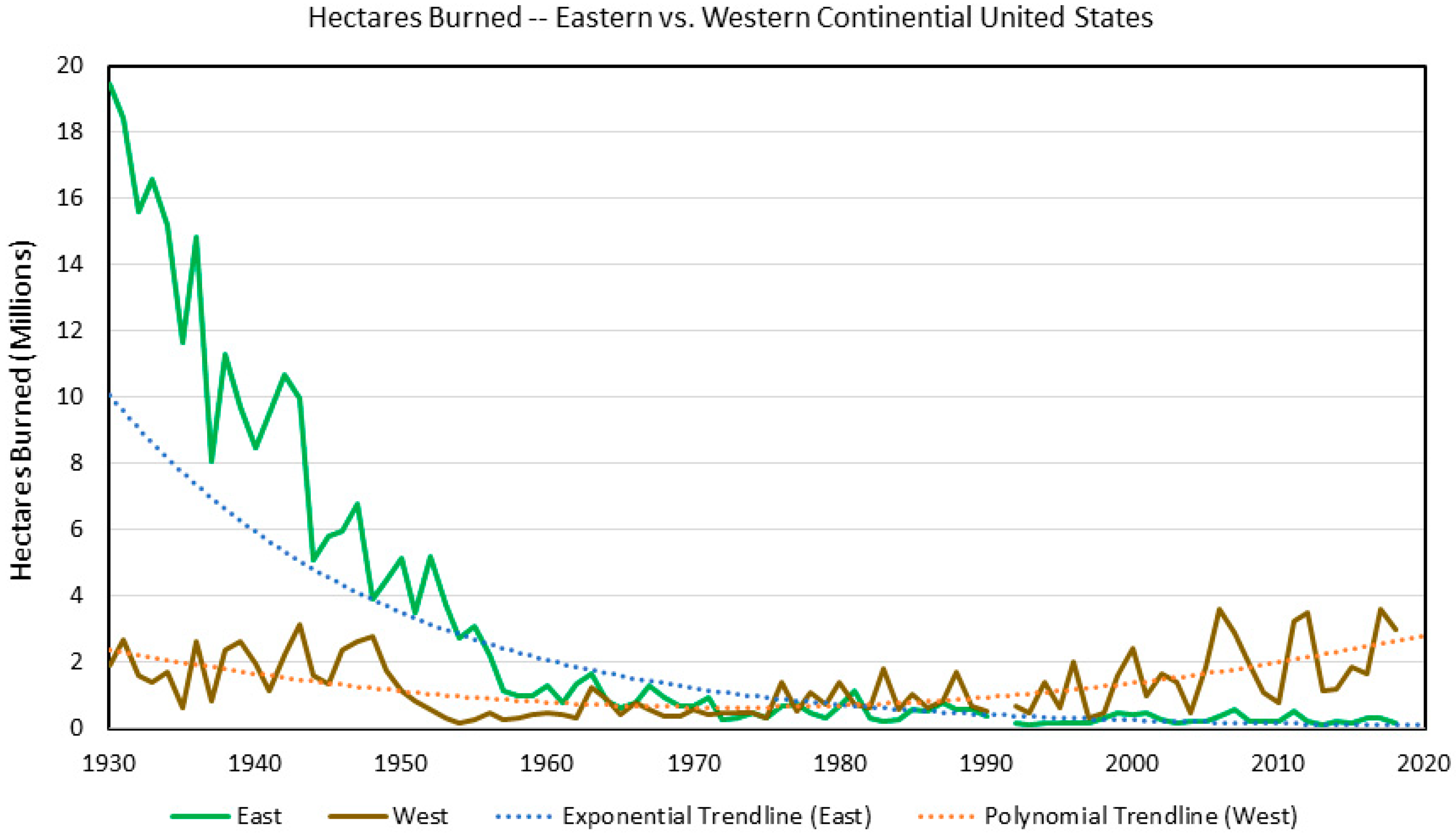
2. Fire
2.1. Climate Change Effects
2.2. Land Use Effects
2.3. Interactions and Relative Factor Strength
2.4. Summary and Future Prediction
3. Tree Growth
3.1. Climate Change Effects
3.2. Land Use Effects
3.3. Interactions and Relative Factor Strength
3.4. Summary and Future Prediction
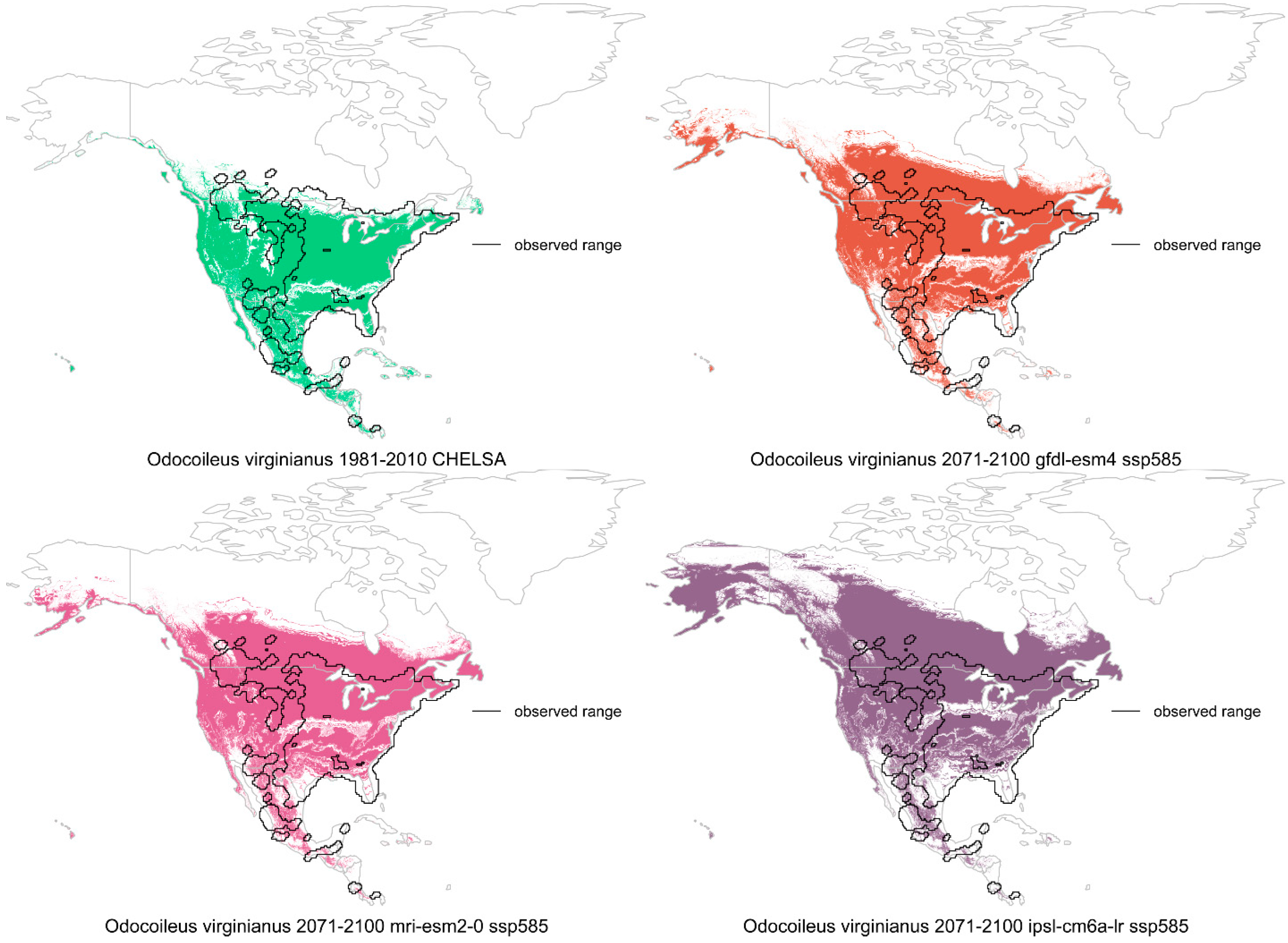
4. Tree Species Distributions
4.1. Climate Change Effects
4.2. Land Use Effects
4.3. Interactions and Relative Factor Strength
4.4. Summary and Future Prediction
5. Invasions and Outbreaks: Non-Native Plants and Non-Native and Native Insects and Diseases
5.1. Non-Native Species
5.2. Outbreaks of Native Insects and Fungi
5.3. Summary and Future Prediction
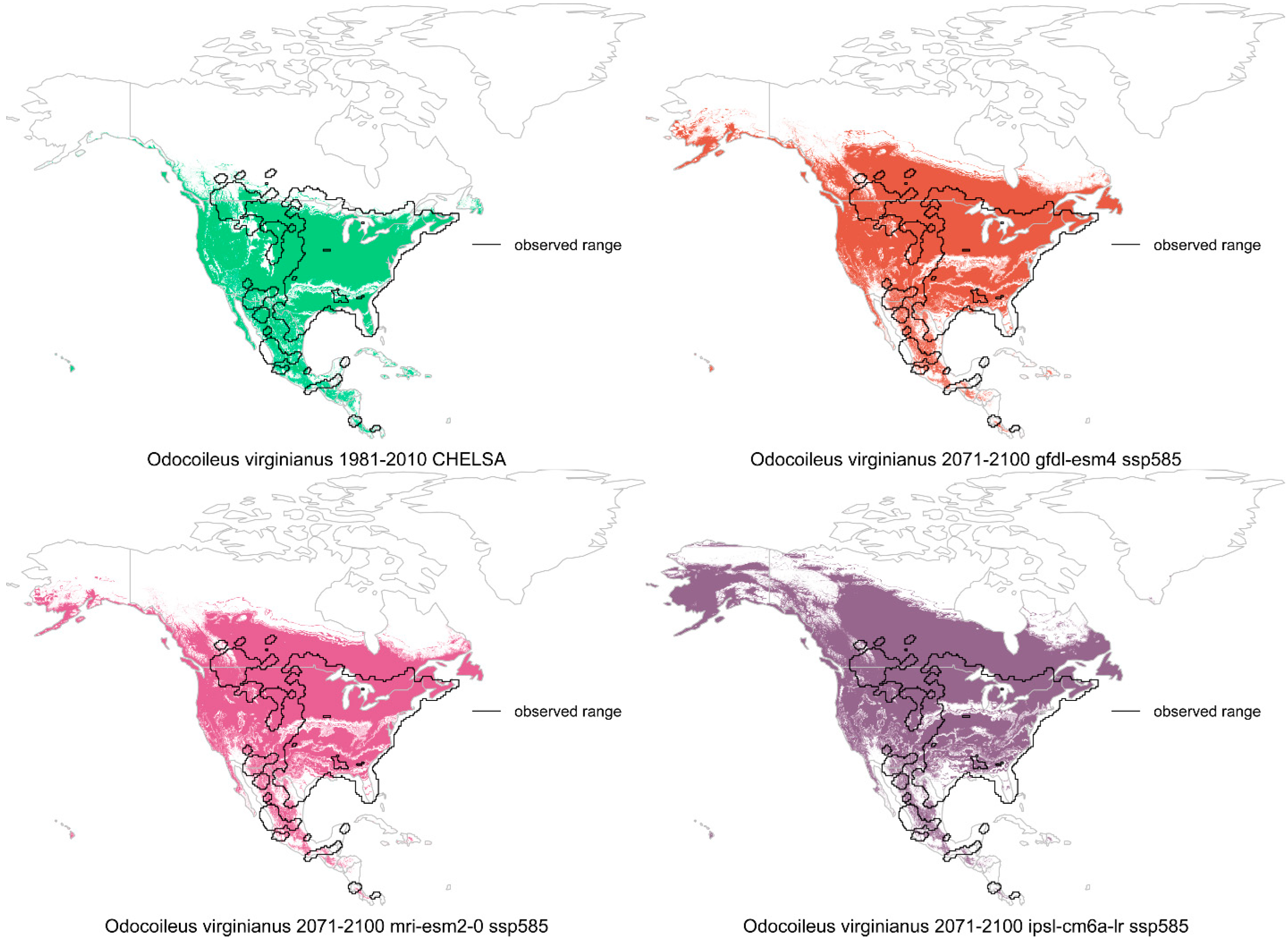
6. Deer, a Remaining Large Herbivore
6.1. Deer Herbivory as a Forest Health Issue
6.2. Climate Change Effects
6.3. Land Use Effects
6.4. Interactions, Relative Factor Strength, and Summary
7. Ecological Forestry to Mitigate Climate Change
8. Summary of Interactions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics: Updated Edition; John Wiley and Sons: Hoboken, NJ, USA, 1996. [Google Scholar]
- Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services [IPBES]. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019; Available online: https://www.ipbes.net/global-assessment (accessed on 10 January 2022).
- Intergovernmental Panel on Climate Change [IPCC]. Summary for Policymakers. In Climate Change 2021, The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, S., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Monteiro, S., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 3–32. [Google Scholar]
- PRISM Climate Group. Oregon State University. 2021. Available online: https://prism.oregonstate.edu (accessed on 29 October 2021).
- Eischeid, J.K.; Hoerling, M.P.; Quan, X.W.; Kumar, A.; Barsugli, J.; Labe, Z.M.; Kunkel, K.E.; Schreck, I.I.I.C.J.; Easterling, D.R.; Zhang, T.; et al. Why has the summertime central US warming hole not disappeared? J. Clim. 2023, 36, 7319–7336. [Google Scholar] [CrossRef]
- Keil, P.; Mauritsen, T.; Jungclaus, J.; Hedemann, C.; Olonscheck, D.; Ghosh, R. Multiple drivers of the North Atlantic warming hole. Nat. Clim. Chang. 2020, 10, 667–671. [Google Scholar] [CrossRef]
- Abatzoglou, J.T. Development of gridded surface meteorological data for ecological applications and modelling. Int. J. Climatol. 2013, 33, 121–131. [Google Scholar] [CrossRef]
- Cook, E.R.; Seager, R.; Heim, R.R., Jr.; Vose, R.S.; Herweijer, C.; Woodhouse, C. Megadroughts in North America: Placing IPCC projections of hydroclimatic change in a long-term paleoclimate context. J. Quat. Sci. 2010, 25, 48–61. [Google Scholar] [CrossRef]
- Abrams, M.D. Fire and the development of oak forests. BioScience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Nevle, R.J.; Bird, D.K.; Ruddiman, W.F.; Dull, R.A. Neotropical human–landscape interactions, fire, and atmospheric CO2 during European conquest. Holocene 2011, 21, 853–864. [Google Scholar] [CrossRef]
- Abrams, M.D.; Nowacki, G.J.; Hanberry, B.B. Oak forests and woodlands as Indigenous landscapes in the Eastern United States. J. Torrey Bot. Soc. 2022, 149, 101–121. [Google Scholar] [CrossRef]
- Malhi, Y.; Doughty, C.E.; Galetti, M.; Smith, F.A.; Svenning, J.C.; Terborgh, J.W. Megafauna and ecosystem function from the Pleistocene to the Anthropocene. Proc. Natl. Acad. Sci. USA 2016, 113, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Nowacki, G.J.; MacCleery, D.W.; Lake, F.K. Native Americans, ecosystem development, and historical range of variation. In Historical Environmental Variation in Conservation and Natural Resource Management; Weins, J.A., Hayward, G.D., Safford, H.D., Giffen, C.M., Eds.; Wiley-Blackwell: West Sussex, UK, 2012; pp. 76–91. [Google Scholar]
- Hanberry, B.B. Timing of tree density increases, influence of climate change, and a land use proxy for tree density increases in the eastern United States. Land 2021, 10, 1121. [Google Scholar] [CrossRef]
- Denevan, W.M. The pristine myth: The landscape of the Americas in 1492. Ann. Assoc. Am. Geogr. 1992, 82, 369–385. [Google Scholar] [CrossRef]
- Foster, D.R. Land use history (1730–1990) and vegetation dynamics in central New England, USA. J. Ecol. 1992, 80, 753–771. [Google Scholar] [CrossRef]
- Whitney, G.G. From Coastal Wilderness to Fruited Plain: A History of Environmental Change in Temperate North America from 1500 to the Present; Cambridge University Press: New York, NY, USA, 1994. [Google Scholar]
- Munoz, S.E.; Gajewski, K.; Peros, M.C. Synchronous environmental and cultural change in the prehistory of the northeastern United States. Proc. Natl. Acad. Sci. USA 2010, 107, 22008–22013. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, G.J.; Abrams, M.D. Is climate an important driver of post European vegetation change in the eastern U.S.? Glob. Chang. Biol. 2015, 21, 314–334. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, G.J.; Abrams, M.D. Demise of fire and mesophication of eastern U.S. forests. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Abrams, M.D.; Arthur, M.A.; Varner, J.M. Reviewing fire, climate, deer, and foundation species as drivers of historically open oak and pine forests and transition to closed forests. Front. For. Glob. Chang. 2020, 3, 56. [Google Scholar] [CrossRef]
- Davison, C.W.; Rahbek, C.; Morueta-Holme, N. Land-use change and biodiversity: Challenges for assembling evidence on the greatest threat to nature. Glob. Chang. Biol. 2021, 27, 5414–5429. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.M.; Varner, J.M.; Metz, M.R.; van Mantgem, P.J. Characterizing interactions between fire and other disturbances and their impacts on tree mortality in western US Forests. For. Ecol. Manag. 2017, 405, 188–199. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Gensini, V.A.; Brooks, H.E. Spatial trends in United States tornado frequency. NPJ Clim. Atmos. Sci. 2018, 1, 38. [Google Scholar] [CrossRef]
- Holl, K.D.; Brancalion, P.H. Tree planting is not a simple solution. Science 2020, 368, 580–5811. [Google Scholar] [CrossRef]
- Reilly, M.J.; Norman, S.P.; O’Brien, J.J.; Loudermilk, E.L. Drivers and ecological impacts of a wildfire outbreak in the southern Appalachian Mountains after decades of fire exclusion. For. Ecol. Manag. 2022, 524, 120500. [Google Scholar] [CrossRef]
- Shuman, B.; Henderson, A.K.; Plank, C.; Stefanova, I.; Ziegler, S.S. Woodland-to-forest transition during prolonged drought in Minnesota after ca. AD 1300. Ecology 2009, 90, 2792–2807. [Google Scholar] [CrossRef]
- Short, K.C. Spatial Wildfire Occurrence Data for the United States, 1992–2020; Forest Service Research Data Archive: Fort Collins, CO, USA, 2022. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R. Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- Adams, H.D.; Guardiola-Claramonte, M.; Barron-Gafford, G.A.; Villegas, J.C.; Breshears, D.D.; Zou, C.B.; Troch, P.A.; Huxman, T.E. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. USA 2009, 106, 7063–7066. [Google Scholar] [CrossRef] [PubMed]
- Dietze, M.C.; Moorcroft, P.R. Tree mortality in the eastern and central United States: Patterns and drivers. Glob. Chang. Biol. 2011, 17, 3312–3326. [Google Scholar] [CrossRef]
- Bazzaz, F.A. The response of natural ecosystems to the rising global CO2 levels. Annu. Rev. Ecol. Syst. 1990, 21, 167–196. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2015, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Karnosky, D.F. Impacts of elevated atmospheric CO2 on forest trees and forest ecosystems: Knowledge gaps. Environ. Int. 2003, 29, 161–169. [Google Scholar] [CrossRef]
- Norby, R.J.; Warren, J.M.; Iversen, C.M.; Medlyn, B.E.; McMurtrie, R.E. CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc. Natl. Acad. Sci. 2010, 107, 19368–19373. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef]
- Mathias, J.M.; Thomas, R.B. Global tree intrinsic water use efficiency is enhanced by increased atmospheric CO2 and modulated by climate and plant functional types. Proc. Natl. Acad. Sci. USA 2021, 118, e2014286118. [Google Scholar] [CrossRef] [PubMed]
- Korner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef]
- Sperry, J.S.; Venturas, M.D.; Todd, H.N.; Trugman, A.T.; Anderegg, W.R.; Wang, Y.; Tai, X. The impact of rising CO2 and acclimation on the response of US forests to global warming. Proc. Natl. Acad. Sci. USA 2019, 116, 25734–25744. [Google Scholar] [CrossRef]
- Voelker, S.L.; Muzika, R.; Guyette, R.P.; Stambaugh, M.C. Historical CO2 growth enhancement declines with age in Quercus and Pinus. Ecol. Monogr. 2006, 76, 549–564. [Google Scholar] [CrossRef]
- McMahon, S.M.; Parker, G.G.; Miller, D.R. Evidence for a recent increase in forest growth. Proc. Natl. Acad. Sci. USA 2010, 107, 3611–3615. [Google Scholar] [CrossRef]
- Davis, E.C.; Sohngen, B.; Lewis, D.J. The effect of carbon fertilization on naturally regenerated and planted US forests. Nat. Commun. 2022, 13, 5490. [Google Scholar] [CrossRef]
- Hogan, J.A.; Domke, G.M.; Zhu, K.; Johnson, D.J.; Lichstein, J.W. Climate change determines the sign of productivity trends in US forests. Proc. Natl. Acad. Sci. USA 2024, 121, e2311132121. [Google Scholar] [CrossRef] [PubMed]
- Finzi, A.C.; Giasson, M.A.; Barker Plotkin, A.A.; Aber, J.D.; Boose, E.R.; Davidson, E.A.; Dietze, M.C.; Ellison, A.M.; Frey, S.D.; Goldman, E.; et al. Carbon budget of the Harvard Forest Long-Term Ecological Research site: Pattern, process, and response to global change. Ecol. Monogr. 2020, 90, e01423. [Google Scholar] [CrossRef]
- Caspersen, J.P.; Pacala, S.W.; Jenkins, J.C.; Hurtt, G.C.; Moorcroft, P.R.; Birdsey, R.A. Contributions of land-use history to carbon accumulation in US forests. Science 2000, 290, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Canham, C.D.; Murphy, L. The demography of tree species response to climate: Sapling and canopy tree survival. Ecosphere 2017, 8, e01701. [Google Scholar] [CrossRef]
- Bishop, D.A.; Beier, C.M.; Pederson, N.; Lawrence, G.B.; Stella, J.C.; Sullivan, T.J. Regional growth decline of sugar maple (Acer saccharum) and its potential causes. Ecosphere 2015, 6, 179. [Google Scholar] [CrossRef]
- Oswald, E.M.; Pontius, J.; Rayback, S.A.; Schaberg, P.G.; Wilmot, S.H.; Dupigny-Giroux, L.A. The complex relationship between climate and sugar maple health: Climate change implications in Vermont for a key northern hardwood species. For. Ecol. Manag. 2018, 422, 303–312. [Google Scholar] [CrossRef]
- Peters, R.L.; Groenendijk, P.; Vlam, M.; Zuidema, P.A. Detecting long-term growth trends using tree rings: A critical evaluation of methods. Glob. Chang. Biol. 2015, 21, 2040–2054. [Google Scholar] [CrossRef]
- D’Orangeville, L.; Houle, D.; Duchesne, L.; Phillips, R.P.; Bergeron, Y.; Kneeshaw, D. Beneficial effects of climate warming on boreal tree growth may be transitory. Nat. Commun. 2018, 9, 3213. [Google Scholar] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Crosby, M.K.; Fan, Z.; Fan, X.; Leininger, T.D.; Spetich, M.A. Assessing forest mortality patterns using climate and FIA data at multiple scales. In Moving from Status to Trends: Forest Inventory and Analysis (FIA) Symposium 2012; Morin, R.S., Liknes, G.C., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012; pp. 319–324. [Google Scholar]
- Johnson, S.; Abrams, M.D. Age class, longevity and growth rate relationships: Protracted growth increases in old trees in the eastern United States. Tree Physiol. 2009, 29, 1317–1328. [Google Scholar] [CrossRef]
- Abrams, M.D. Where has all the white oak gone? BioScience 2003, 53, 927–939. [Google Scholar] [CrossRef]
- Cram, M.M.; Outcalt, K.W.; Zarnoch, S.J. Growth of longleaf and loblolly pine planted on South Carolina sandhill sites. South. J. Appl. For. 2010, 34, 79–83. [Google Scholar] [CrossRef]
- Bragg, D.C. A reevaluation of superior tree performance after 48 years for a loblolly pine progeny test in southern Arkansas. Front. For. Glob. Chang. 2021, 4, 716443. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, J.M.; Birdsey, R.; McCullough, K.; He, L.; Deng, F. Age structure and disturbance legacy of North American forests. Biogeosciences 2011, 8, 715–732. [Google Scholar] [CrossRef]
- Hanberry, B.B. Confronting the issue of invasive native tree species due to land use change in the eastern United States. Land 2022, 11, 161. [Google Scholar] [CrossRef]
- Searle, E.B.; Chen, H.Y.; Paquette, A. Higher tree diversity is linked to higher tree mortality. Proc. Natl. Acad. Sci. USA 2022, 119, e2013171119. [Google Scholar] [CrossRef] [PubMed]
- Quinn Thomas, R.; Canham, C.D.; Weathers, K.C.; Goodale, C.L. Increased tree carbon storage in response to nitrogen deposition in the US. Nat. Geosci. 2010, 3, 13–17. [Google Scholar] [CrossRef]
- Clark, J.S.; Bell, D.M.; Hersh, M.H.; Nichols, L. Climate change vulnerability of forest biodiversity: Climate and competition tracking of demographic rates. Glob. Chang. Biol. 2011, 17, 1834–1849. [Google Scholar] [CrossRef]
- Duveneck, M.J.; Thompson, J.R.; Gustafson, E.J.; Liang, Y.; de Bruijn, A.M. Recovery dynamics and climate change effects to future New England forests. Landsc. Ecol. 2017, 32, 1385–1397. [Google Scholar] [CrossRef]
- Yaussy, D.A.; Iverson, L.R.; Matthews, S.N. Competition and climate affects US hardwood-forest tree mortality. For. Sci. 2013, 59, 416–430. [Google Scholar] [CrossRef]
- Lines, E.R.; Coomes, D.A.; Purves, D.W. Influences of forest structure, climate and species composition on tree mortality across the eastern US. PLoS ONE 2010, 5, e13212. [Google Scholar] [CrossRef]
- Brienen, R.J.; Caldwell, L.; Duchesne, L.; Voelker, S.; Barichivich, J.; Baliva, M.; Ceccantini, G.; Di Filippo, A.; Helama, S.; Locosselli, G.M.; et al. Forest carbon sink neutralized by pervasive growth-lifespan trade-offs. Nat. Commun. 2020, 11, 4241. [Google Scholar] [CrossRef]
- Perret, D.L.; Leslie, A.B.; Sax, D.F. Naturalized distributions show that climatic disequilibrium is structured by niche size in pines (Pinus L.). Glob. Ecol. Biogeogr. 2019, 28, 429–441. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Naimi, B.; Rahbek, C.; Araújo, M.B. Improvements in reports of species redistribution under climate change are required. Sci. Adv. 2021, 7, eabe1110. [Google Scholar] [CrossRef] [PubMed]
- Roe, G.H.; Baker, M.B.; Herla, F. Centennial glacier retreat as categorical evidence of regional climate change. Nat. Geosci. 2017, 10, 95–99. [Google Scholar] [CrossRef]
- Smith, W.K.; Germino, M.J.; Johnson, D.M.; Reinhardt, K. The altitude of alpine treeline: A bellwether of climate change effects. Bot. Rev. 2009, 75, 163–190. [Google Scholar] [CrossRef]
- Gehrig-Fasel, J.; Guisan, A.; Zimmermann, N.E. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 2007, 18, 571–582. [Google Scholar] [CrossRef]
- Beckage, B.; Osborne, B.; Gavin, D.G.; Pucko, C.; Siccama, T.; Perkins, T. A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc. Natl. Acad. Sci. USA 2008, 105, 4197–4202. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, X. Effects of temperature variability and extremes on spring phenology across the contiguous United States from 1982 to 2016. Sci. Rep. 2020, 10, 17952. [Google Scholar] [CrossRef]
- Bórnez, K.; Verger, A.; Descals, A.; Peñuelas, J. Monitoring the responses of deciduous forest phenology to 2000–2018 climatic anomalies in the Northern Hemisphere. Remote Sens. 2021, 13, 2806. [Google Scholar] [CrossRef]
- Day, G.M. The Indian as an ecological factor in the northeastern forest. Ecology 1953, 34, 329–346. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Noss, R.F. Locating potential historical fire-maintained grasslands of the eastern United States based on topography and wind speed. Ecosphere 2022, 13, e4098. [Google Scholar] [CrossRef]
- Hanberry, B.B. Non-native plant species richness and influence of greenhouses and human populations in the conterminous United States. Ecol. Process. 2023, 12, 27. [Google Scholar] [CrossRef]
- Bradley, B.A.; Blumenthal, D.M.; Wilcove, D.S.; Ziska, L.H. Predicting plant invasions in an era of global change. Trends Ecol. Evol. 2010, 25, 310–318. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Sobek-Swant, S.; Crosthwaite, J.C.; Lyons, D.B.; Sinclair, B.J. Could phenotypic plasticity limit an invasive species? Incomplete reversibility of mid-winter deacclimation in emerald ash borer. Biol. Invasions 2012, 14, 115–125. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cleland, E.E. Phenological niches and the future of invaded ecosystems with climate change. AoB Plants 2014, 6, plu013. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Davis, T.J.; Schaefer, H.; Cleland, E.E.; Cook, B.I.; Travers, S.E.; Willis, C.G.; Davis, C.C. Temperature-dependent shifts in phenology contribute to the success of exotic species with climate change. Am. J. Bot. 2013, 100, 1407–1421. [Google Scholar] [CrossRef]
- Robinson, E.A.; Ryan, G.D.; Newman, J.A. A meta-analytical review of the effects of elevated CO2 on plant-arthropod interactions highlights the importance of interacting environmental and biological variables. New Phytol. 2012, 194, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Meentemeyer, R.K.; Cunniffe, N.J.; Cook, A.R.; Filipe, J.A.; Hunter, R.D.; Rizzo, D.M.; Gilligan, C.A. Epidemiological modeling of invasion in heterogeneous landscapes: Spread of sudden oak death in California (1990–2030). Ecosphere 2011, 2, 1–24. [Google Scholar] [CrossRef]
- Brasier, C.M. Rapid evolution of introduced plant pathogens via interspecific hybridization. BioScience 2001, 51, 123–133. [Google Scholar] [CrossRef]
- EDDMapS. 2021. Available online: https://www.eddmaps.org/species/ (accessed on 19 November 2021).
- Liebhold, A.M.; Brockerhoff, E.G.; Garrett, L.J.; Parke, J.L.; Britton, K.O. Live plant imports: The major pathway for forest insect and pathogen invasions of the United States. Front. Ecol. Environ. 2012, 10, 135–143. [Google Scholar] [CrossRef]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1347. [Google Scholar] [CrossRef]
- McNulty, S.G.; Boggs, J.L.; Sun, G. The rise of the mediocre forest: Why chronically stressed trees may better survive extreme episodic climate variability. New For. 2014, 45, 403–415. [Google Scholar] [CrossRef]
- Asaro, C.; Koch, F.H.; Potter, K.M. Denser forests across the USA experience more damage from insects and pathogens. Sci. Rep. 2023, 13, 3666. [Google Scholar] [CrossRef] [PubMed]
- Romme, W.H.; Clement, J.; Hicke, J.; Kulakowski, D.; MacDonald, L.H.; Schoennagel, T.L.; Veblen, T.T. Recent Forest Insect Outbreaks and Fire Risk in Colorado Forests: A Brief Synthesis of Relevant Research; Colorado Forest Restoration Institute: Fort Collins, CO, USA, 2006. [Google Scholar]
- De Chazal, J.; Rounsevell, M.D. Land-use and climate change within assessments of biodiversity change: A review. Glob. Environ. Chang. 2009, 19, 306–315. [Google Scholar] [CrossRef]
- Robinson, G.S.; Burney, L.P.; Burney, D.A. Landscape paleoecology and megafaunal extinction in southeastern New York State. Ecol. Monogr. 2005, 75, 295–315. [Google Scholar] [CrossRef]
- Gill, J.L.; Williams, J.W.; Jackson, S.T.; Lininger, K.B.; Robinson, G.S. Pleistocene megafaunal collapse, novel plant communities, and enhanced fire regimes in North America. Science 2009, 326, 1100–1103. [Google Scholar] [CrossRef]
- Seton, E.Y. Lives of Game Animals; Branford: Boston, MA, USA, 1927; Volume III, Part I; Available online: https://archive.org/details/livesofgameanima0003seto/page/n15/mode/2up (accessed on 19 June 2022).
- Hanberry, B.; Hanberry, P. Rapid digitization to reclaim thematic maps of white-tailed deer density from 1982 and 2003 in the conterminous US. PeerJ 2020, 8, e8262. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Abrams, M.D. Does white-tailed deer density affect tree stocking in forests of the eastern United States? Ecol. Process. 2019, 8, 30. [Google Scholar] [CrossRef]
- Leopold, A.; Sowls, L.K.; Spencer, D.L. A survey of over-populated deer ranges in the United States. J. Wildl. Manag. 1947, 11, 162–177. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Faison, E.K. Re-framing deer herbivory as a natural disturbance regime with ecological and socioeconomic outcomes in the eastern United States. Sci. Total Environ. 2023, 868, 161669. [Google Scholar] [CrossRef]
- Hanberry, B.B. Practical guide for retaining correlated climate variables and unthinned samples in species distribution modeling, using random forests. Ecol. Inform. 2024, 79, 102406. [Google Scholar] [CrossRef]
- Hanberry, B.B. Addressing regional relationships between white-tailed deer densities and land classes. Ecol. Evol. 2021, 11, 13570–13578. [Google Scholar] [CrossRef] [PubMed]
- Loehle, C. Height growth rate tradeoffs determine northern and southern range limits for trees. J. Biogeogr. 1998, 25, 735–742. [Google Scholar] [CrossRef]
- Abrams, M.D. Adaptations and responses to drought in Quercus species of North America. Tree Physiol. 1990, 7, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Stanturf, J.A.; Goodrick, S.L.; Outcalt, K.W. Disturbance and coastal forests: A strategic approach to forest management in hurricane impact zones. For. Ecol. Manag. 2007, 250, 119–135. [Google Scholar] [CrossRef]
- Bragg, D.C.; Hanberry, B.B.; Hutchinson, T.F.; Jack, S.B.; Kabrick, J.M. Silvicultural options for open forest management in eastern North America. For. Ecol. Manag. 2020, 474, 118383. [Google Scholar] [CrossRef]
- Holzmueller, E.J.; Jose, S.; Jenkins, M.A. The relationship between fire history and an exotic fungal disease in a deciduous forest. Oecologia 2008, 155, 347–356. [Google Scholar] [CrossRef]
- Danneyrolles, V.; Dupuis, S.; Fortin, G.; Leroyer, M.; de Römer, A.; Terrail, R.; Vellend, M.; Boucher, Y.; Laflamme, J.; Bergeron, Y.; et al. Stronger influence of anthropogenic disturbance than climate change on century-scale compositional changes in northern forests. Nat. Commun. 2019, 10, 1265. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.A.; Anderson, L.; Bunting, M.J.; Champagne, M.; Clayburn, R.M.; Crawford, J.N.; Klimaszewski-Patterson, A.; Knapp, E.E.; Lake, F.K.; Mensing, S.A.; et al. Land management explains major trends in forest structure and composition over the last millennium in California’s Klamath Mountains. Proc. Natl. Acad. Sci. USA 2022, 119, e2116264119. [Google Scholar] [CrossRef] [PubMed]
- Napier, J.D.; Chipman, M.L. Emerging palaeoecological frameworks for elucidating plant dynamics in response to fire and other disturbance. Glob. Ecol. Biogeogr. 2022, 31, 138–154. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Issue | Climate | Land Use | Realized Outcome in the Eastern U.S. | Current Interaction under Increased Precipitation | Future Interaction under Warming without Offsetting Precipitation |
|---|---|---|---|---|---|
| Fire | Fuel moisture, ignition probability, fuel quantity | Switches fire severity and frequency through fuel type (fine herbaceous or coarse fuels with mesophication of conditions) | Increased precipitation decreases fire frequency because land use (surface fire exclusion) produces forests | Synergistic to reduce fires | Antagonistic |
| Tree growth | Growth, reproduction, and mortality, and overall forest productivity | Species, age, densities | Increased precipitation and carbon fertilization promote tree growth; land use produces young, dense stands of fast-growing tree species | Synergistic to maximize annual tree growth | Antagonistic |
| Tree distribution | Shifts to stay within climate envelope | Expansion to fire-free areas | Expansion to fire-free areas | None | Varies by species |
| Non-native plants, insects, fungi | Survival, growth, reproduction, mortality, distribution shifts; water availability for trees; fire | Introduction, spread; tree densities, basal area, age; fire | Non-native species introduction and spread due to human activities | None overall | Varies by species |
| Native insects and fungi | Survival, growth, reproduction, mortality, distribution shifts; water availability for trees; fire | Tree densities, basal area, age; fire | Outbreaks in dense forests from land use | None overall | Varies by species |
| Deer | Survival, growth, reproduction, mortality, distribution shifts | Overexploitation, forest cover | Recovery after overexploitation | None | None to slight |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanberry, B.B.; Abrams, M.D.; Nowacki, G.J. Potential Interactions between Climate Change and Land Use for Forest Issues in the Eastern United States. Land 2024, 13, 398. https://doi.org/10.3390/land13030398
Hanberry BB, Abrams MD, Nowacki GJ. Potential Interactions between Climate Change and Land Use for Forest Issues in the Eastern United States. Land. 2024; 13(3):398. https://doi.org/10.3390/land13030398
Chicago/Turabian StyleHanberry, Brice B., Marc D. Abrams, and Gregory J. Nowacki. 2024. "Potential Interactions between Climate Change and Land Use for Forest Issues in the Eastern United States" Land 13, no. 3: 398. https://doi.org/10.3390/land13030398
APA StyleHanberry, B. B., Abrams, M. D., & Nowacki, G. J. (2024). Potential Interactions between Climate Change and Land Use for Forest Issues in the Eastern United States. Land, 13(3), 398. https://doi.org/10.3390/land13030398