
_Kazoglou.png)
Military Activity Impact on Vegetation in Pannonian Dry Sandy Grasslands

 , ,
, ,  , , ,
, , ,
Abstract
1. Introduction
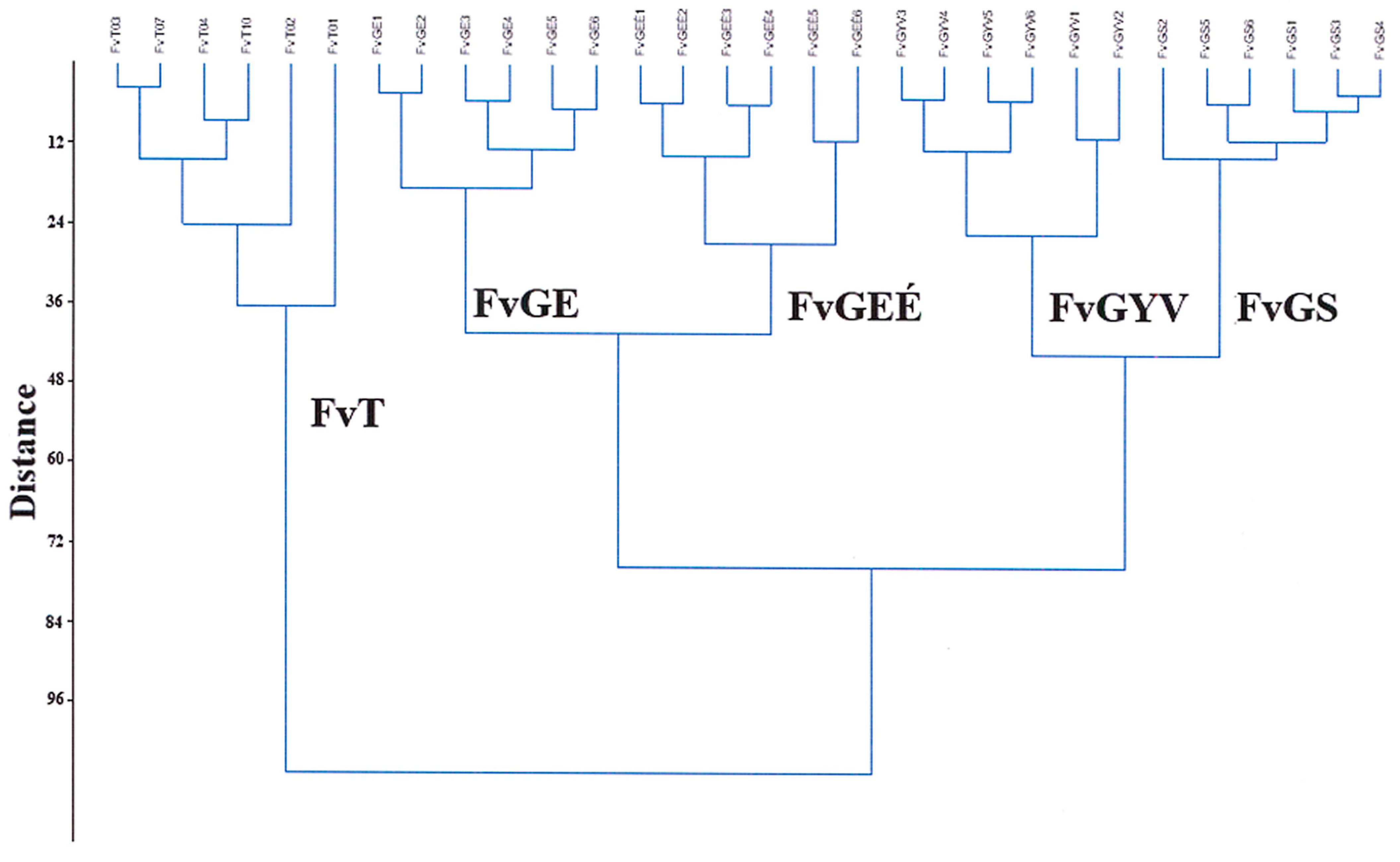
2. Materials and Methods
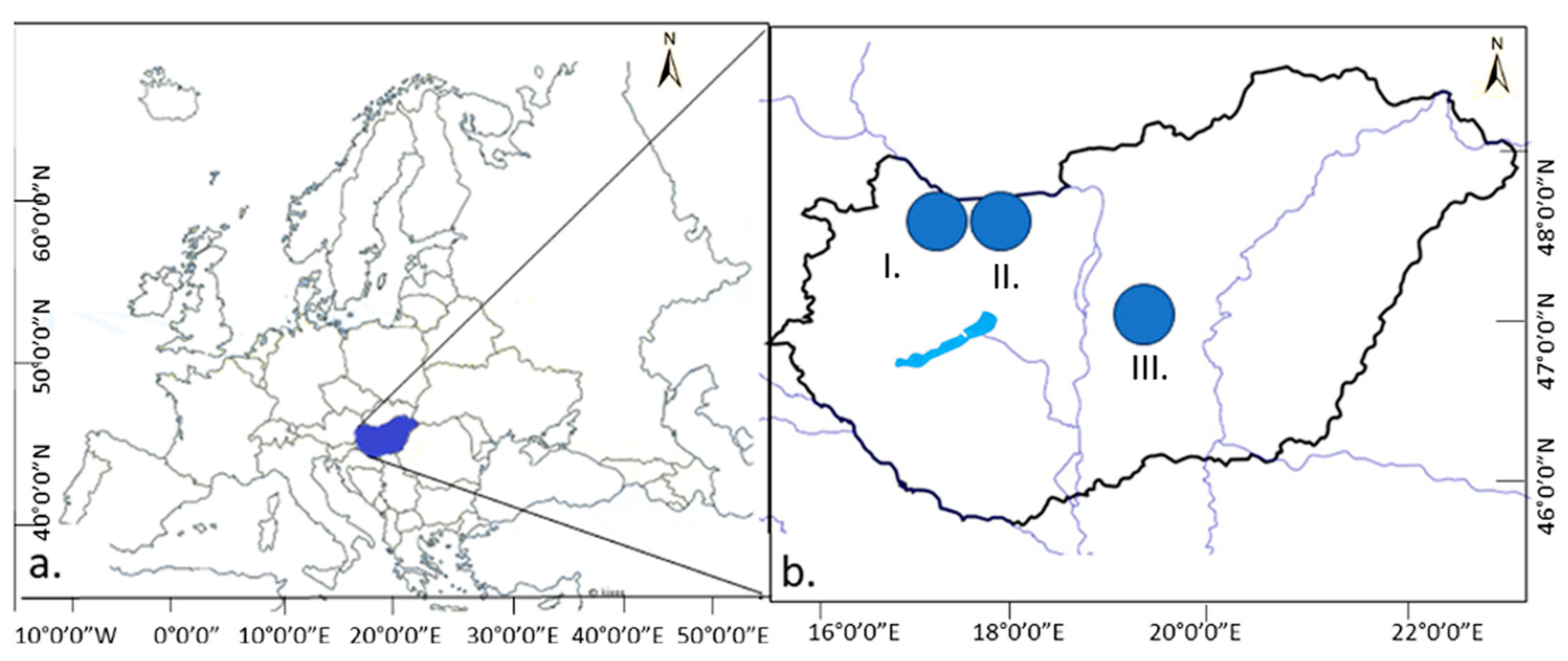
2.1. Description of the Study Areas
- I.
- Military Shooting Range of Gönyű. Its main purpose was the habitat restoration and establishment of Festucetum vaginatae.
- I.1.
- FvGE: Festucetum vaginatae, an original, natural, calcareous, open sandy grassland, unmanaged.
- I.2.
- FvGEÉ: Festucetum vaginatae, an original, natural, calcareous, open sandy grassland, but burnt for 3 years and abandoned 5 years before this study, unmanaged.
- I.3.
- FvGS: Festucetum vaginatae, a manually sown semi-natural grassland, where an artificial sandy ridge, rampart, was established.
- II.
- Military Exercise Area of Győrszentiván. Its main purpose was to establish a habitat through grassland establishment for economic and nature conservation values.
- II.1.
- FvGYV: Festucetum vaginatae, a sown semi-natural grassland.
- II.2.
- FrGYV: Festucetum rupicolae, a sown semi-natural grassland.
- II.3.
- FrGYI: Festucetum rupicolae, a natural grassland.
- III.
- Military Exercise Area of Tatárszentgyörgy. There is still active military activity (military exercises) here.
- III.1.
- FpT: Festucetum pseudovaginatae, natural or semi-natural grassland.
- III.2.
- FvT: Festucetum vaginatae, natural or semi-natural grassland.
2.2. Field Methodology
2.3. Statistical Analysis
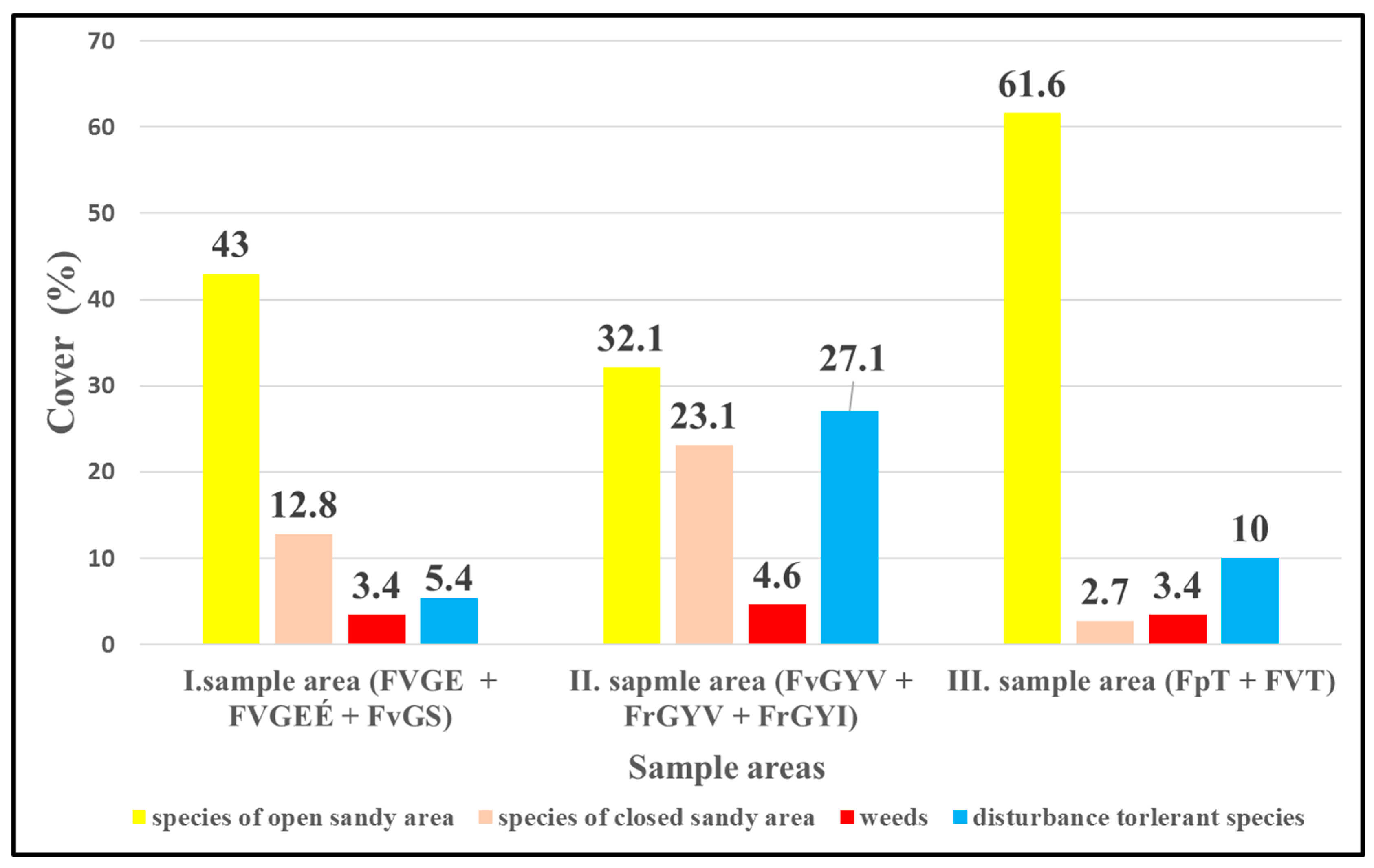
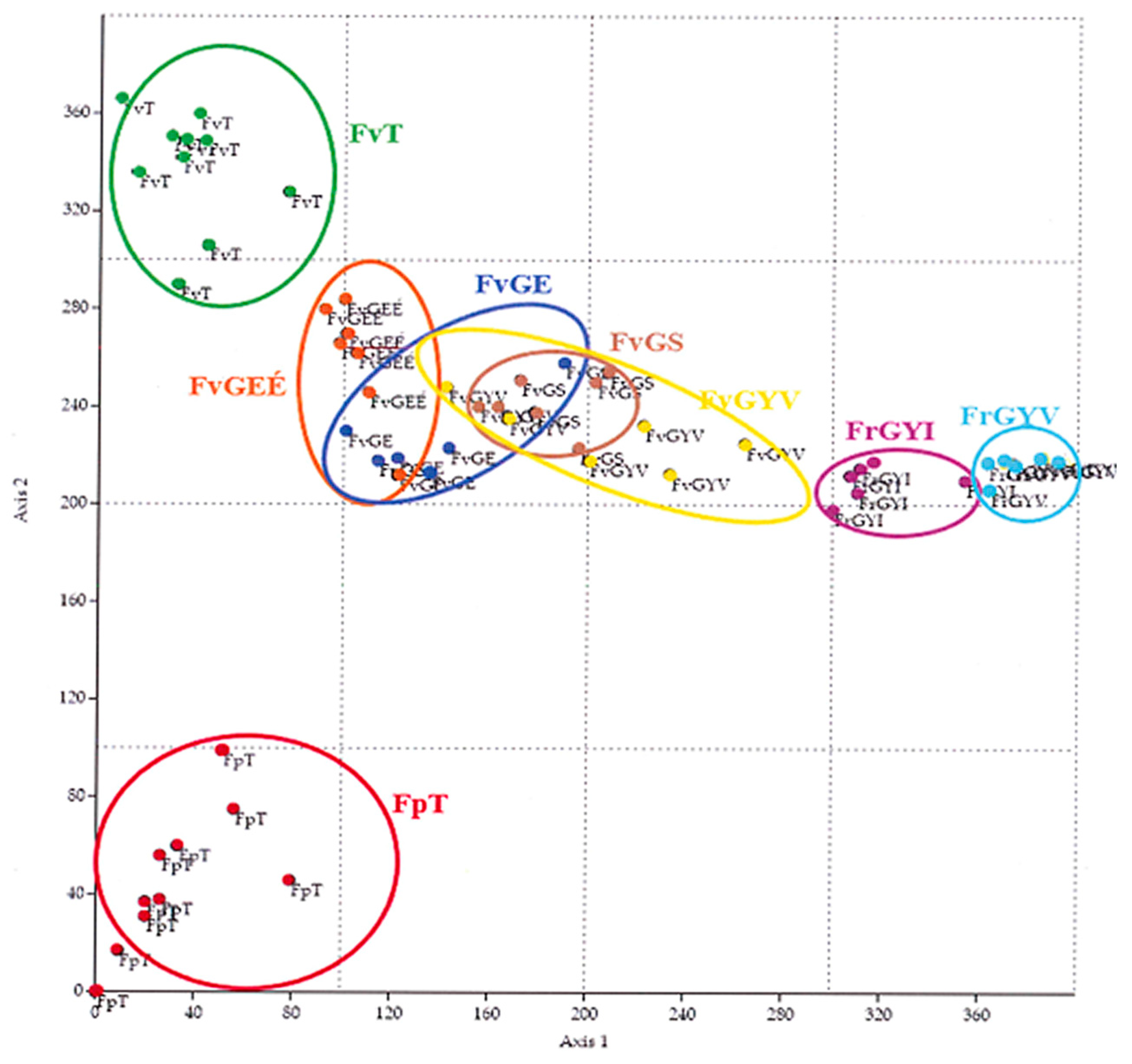
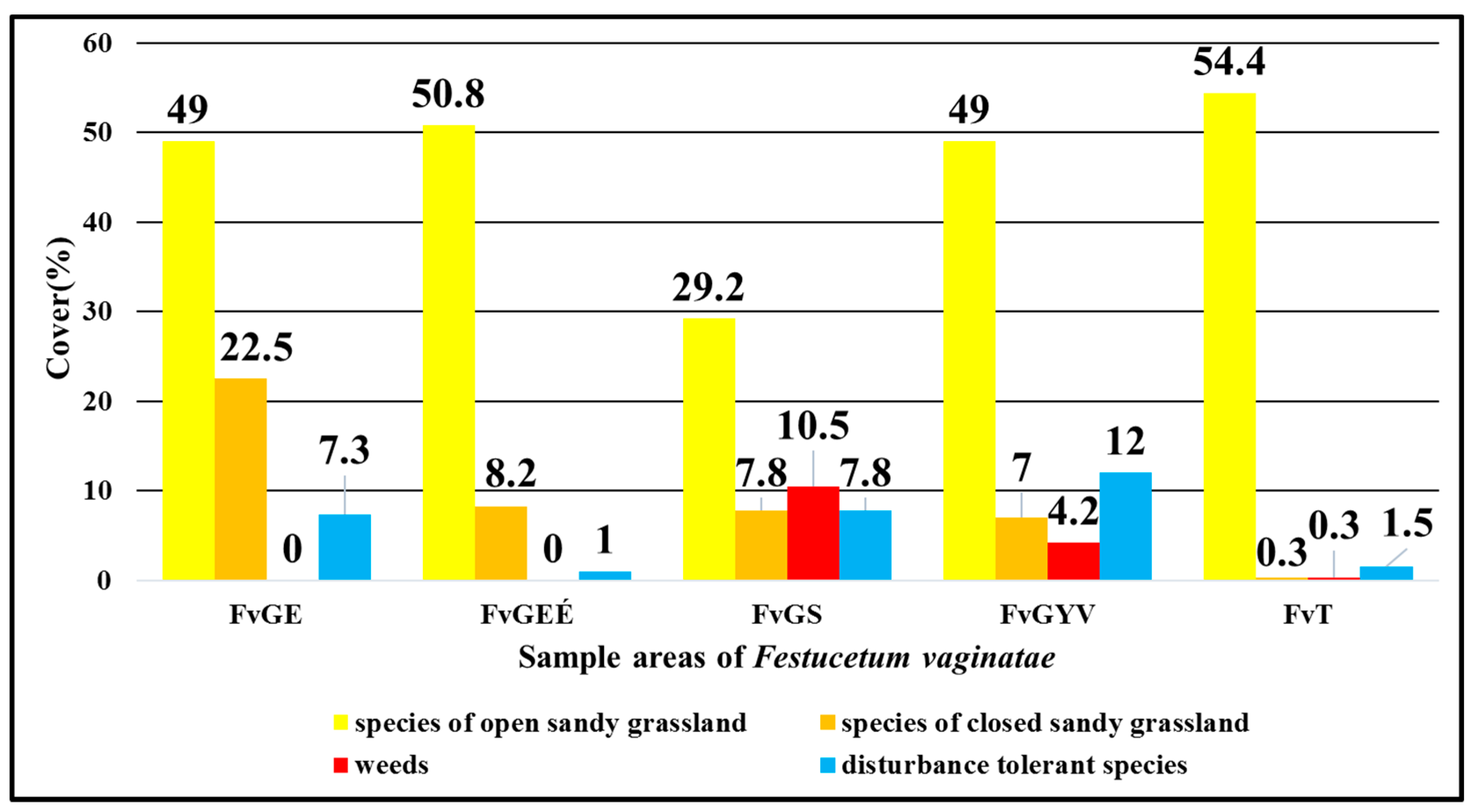
3. Results
- Characteristic species in open sandy grassland (Festucion vaginatae, Corynephoretalia, Festucetalia vaginatae & rupicolae, and Festucetalia vaginatae & valesiacae).
- Characteristic species in closed natural sandy grassland (Brometea, Festuco-Brometea, Festucetalia valesiacae) refer to the natural condition of the grasslands.
- 3.
- Weeds.
- 4.
- Disturbance-tolerant species (Figure 2).
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bischoff, A.; Auge, H.; Mahn, E.G. Seasonal changes in the relationship between plant species richness and community biomass in early succession. Basic Appl. Ecol. 2005, 6, 385–394. [Google Scholar] [CrossRef]
- Guo, Q. The diversity–biomass–productivity relationships in grassland management and restoration. Basic Appl. Ecol. 2007, 8, 199–208. [Google Scholar] [CrossRef]
- Penksza, K.; Saláta, D.; Pápay, G.; Péter, N.; Bajor, Z.; Lisztes-Szabó, Z.; Fűrész, A.; Fuchs, M.; Michéli, E. Do Sandy Grasslands along the Danube in the Carpathian Basin Preserve the Memory of Forest-Steppes? Forests 2021, 12, 114. [Google Scholar] [CrossRef]
- Albert, Á.J.; Kelemen, A.; Valkó, O.; Miglécz, T.; Csecserits, A.; Rédei, T.; Deák, B.; Tóthmérész, B.; Török, P. Secondary succession in sandy old-fields: A promising example of spontaneous grassland recovery. Appl. Veg. Sci. 2014, 17, 214–224. [Google Scholar] [CrossRef]
- Bajor, Z.; Zimmermann, Z.; Szabó, G.; Fehér, Z.; Járdi, I.; Lampert, R.; Kerény-Nagy, V.; Penksza, P.; Lisztes-Szabó, Z.; Székely, Z.; et al. Effect of conservation management practices on sand grassland vegetation in Budapest, Hungary. Appl. Ecol. Environ. Res. 2016, 14, 233–247. [Google Scholar] [CrossRef]
- Catorci, A.; Piermarteri, K.; Penksza, K.; Házi, J.; Tardella, F.M. Filtering effect of temporal niche fluctuation and amplitude of environmental variations on the trait-related flowering patterns: Lesson from sub-Mediterranean grasslands. Sci. Rep. 2017, 7, 12034. [Google Scholar] [CrossRef] [PubMed]
- Clevell, A.F. Restoring for natural authenticity. Ecol. Restor. 2000, 18, 216–217. [Google Scholar] [CrossRef][Green Version]
- Cornwell, W.K.; Grubb, P.J. Regional and local patterns in plant species richness with respect to resource availability. Oikos 2003, 100, 417–428. [Google Scholar] [CrossRef]
- Coulson, T.; Mace, G.M.; Hudson, E.; Possingham, H. The use and abuse of population viability analysis. Trends Ecol. Evol. 2001, 16, 219–221. [Google Scholar] [CrossRef]
- Critchley, C.N.R.; Burke, M.J.W.; Stevens, D.P. Conservation of lowland semi-natural grasslands in the UK: A review of botanical monitoring results from agri-environment schemes. Biol. Conserv. 2004, 115, 263–278. [Google Scholar] [CrossRef]
- Csecserits, A.; Czúcz, B.; Halassy, M.; Kröel-Dulay, G.; Rédei, T.; Szabó, K.; Szitár, R.; Török, K. Regeneration of sandy old-fields in the forest steppe region of Hungary. Plant Biosyst. 2011, 145, 715–729. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Kelemen, A.; Török, P.; Miglécz, T.; Ölvedi, T.; Lengyel, S.; Tóthmérész, B. Litter and graminoid biomass accumulation suppresses weedy forbs in grassland restoration. Plant Biosyst. 2011, 145, 730–737. [Google Scholar] [CrossRef]
- Dolt, C.; Goverde, M.; Baur, B. Effects of experimental small-scale habitat fragmentation on above- and below-ground plant biomass in calcareous grasslands. Acta Oecologica 2005, 27, 49–56. [Google Scholar] [CrossRef]
- Gillman, L.N.; Wright, S.D. The influence of productivity on the species richness of plants. a critical assessment. Ecology 2006, 87, 234–1243. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Steiner, C.F.; Scheiner, S.M.; Gross, K.L.; Reynolds, H.L.; Waide, R.B.; Willig, M.R.; Dodson, S.I.; Gough, L. What is the observed relationship between species richness and productivity? Ecology 2001, 82, 2381–2396. [Google Scholar] [CrossRef]
- Rehounková, K.; Prach, K. Spontaneous vegetation succession in disused gravel-sand pits: Role of local site and landscape factors. J. Veg. Sci. 2006, 17, 583–590. [Google Scholar] [CrossRef]
- Schaffers, A.P. Soil, biomass, and management of semi-natural vegetation—Part II. Factors controlling species diversity. Plant Ecol. 2002, 158, 247–268. [Google Scholar] [CrossRef]
- Valkó, O.; Deák, B.; Török, P.; Kirmer, A.; Tischew, S.; Kelemen, A.; Tóth, K.; Miglécz, T.; Radócz, S.; Sonkoly, J.; et al. High-diversity sowing in establishment gaps: A promising new tool for enhancing grassland biodiversity. Tuexenia 2016, 36, 359–378. [Google Scholar] [CrossRef]
- Hölzel, N.; Otte, A. Restoration of a species-rich flood meadow by topsoil removal and diaspore transfer with plant material. Appl. Veg. Sci. 2003, 6, 131–140. [Google Scholar] [CrossRef]
- Jongepierová, I.; Mitchley, J.; Tzanopoulos, J. A field experiment to recreate species rich hay meadows using regional seed mixtures. Biol. Conserv. 2007, 139, 297–305. [Google Scholar] [CrossRef]
- Lepŝ, J.; Doleẑal, J.; Bezemer, T.M.; Brown, V.K.; Hedlund, K.; Igual Arroyo, M.; Jörgensen, H.B.; Lawson, C.S.; Mortimer, S.R.; Peix Geldart, A.; et al. Long-term effectiveness of sowing high and low diversity seed mixtures to enhance plant community development on ex-arable fields. Appl. Veg. Sci. 2007, 10, 97–110. [Google Scholar] [CrossRef]
- Török, P.; Miglécz, T.; Valkó, O.; Kelemen, A.; Deák, B.; Lengyel, S.; Tóthmérész, B. Recovery of native grass biodiversity by sowing on former croplands: Is weed suppression a feasible goal for grassland restoration? J. Nat. Conserv. 2012, 20, 41–48. [Google Scholar] [CrossRef]
- Mijnsbrugge, V.K.; Bischoff, A.; Smith, B. A question of origin: Where and how to collect seed for ecological restoration. Basic Appl. Ecol. 2010, 11, 300–311. [Google Scholar] [CrossRef]
- Pywell, R.F.; Bullock, J.M.; Hopkins, A.; Walker, K.J.; Sparks, T.H.; Burke, M.J.W.; Peel, S. Restoration of species, rich grassland on arable land: Assessing the limiting processes using a multi, site experiment. J. Appl. Ecol. 2002, 39, 294–309. [Google Scholar] [CrossRef]
- Rasran, l.; Vogt, K.; Jensen, K. Seed content and conservation evaluation of hay material of fen grasslands. J. Nat. Conserv. 2006, 14, 34–45. [Google Scholar] [CrossRef]
- Omidi, M.; Heydari, M.; Abedi, M.; Kohzadean, M.; Valkó, O.; Prévosto, B. Evaluating the restoration potential of soil seed banks in degraded semi-arid oak forests: Influence of canopy cover types and fire-related cues on seed germination. Forest Ecol. Manag. 2022, 524, 120534. [Google Scholar] [CrossRef]
- Valkó, O.; Kelemen, A.; Miglécz, T.; Török, P.; Deák, B.; Tóth, K.; Tóth, J.P.; Tóthmérész, B. Litter removal does not compensate detrimental fire effects on biodiversity in regularly burned semi-natural grasslands. Sci. Total Environ. 2018, 622–623, 783–789. [Google Scholar] [CrossRef]
- Centeri, C.; Herczeg, E.; Vona, M.; Balázs, K.; Penksza, K. The effects of land-use change on plant-soil-erosion relations, Nyereg Hill, Hungary. J. Plant Nutr. Soil Sci. 2009, 172, 586–592. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Végvári, Z.; Hartel, T.; Schmotzer, A.; Kapocsi, I.; Tóthmérész, B. Grassland fires in Hungary—Experiences of nature conservationists on the effects of fire on biodiversity. Appl. Ecol. Environ. Res. 2014, 12, 267–283. [Google Scholar] [CrossRef]
- Broomandi, P.; Guney, M.; Kim, J.R.; Karaca, F. Soil Contamination in Areas Impacted by Military Activities: A Critical Review. Sustainability 2020, 12, 9002. [Google Scholar] [CrossRef]
- Stadler, T.; Temesi, Á.; Lakner, Z. Soil Chemical Pollution and Military Actions: A Bibliometric Analysis. Sustainability 2022, 14, 7138. [Google Scholar] [CrossRef]
- Warren, S.D.; Holbrook, S.W.; Dale, D.A.; Whelan, N.L.; Elyn, M.; Grimm, W.; Jentsch, A. Biodiversity and the Heterogeneous Disturbance Regime on Military Training Lands. Restor. Ecol. 2007, 15, 606–612. [Google Scholar] [CrossRef]
- Whitecotton, R.C.A.; David, M.B.; Darmody, R.G.; Price, D.L. Impact of Foot Traffic from Military Training on Soil and Vegetation Properties. Environ. Manag. 2000, 26, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Milchunas, D.G.; Schulz, K.A.; Shaw, R.B. Plant Community Structure in Relation to Long-Term Disturbance by Mechanized Military Maneuvers in a Semiarid Region. Environ. Manag. 2000, 25, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Cizek, O.; Vrba, P.; Benes, J.; Hrazsky, Z.; Koptik, J.; Kucera, T.; Marhoul, P.; Zamecnik, J.; Konvicka, M. Conservation Potential of Abandoned Military Areas Matches That of Established Reserves: Plants and Butterflies in the Czech Republic. PLoS ONE 2013, 8, e53124. [Google Scholar] [CrossRef]
- Ellwanger, G.; Reiter, K. Nature conservation on decommissioned military training areas—German approaches and experiences. J. Nat. Conserv. 2019, 49, 1–8. [Google Scholar] [CrossRef]
- Krawczyk, R.; Zubel, R.; Komsta, Ł. Military Training Areas and Vegetation—The Effect of Explosion Craters on Species Diversity along a Moisture Gradient. Pol. J. Ecol. 2019, 67, 194–205. [Google Scholar] [CrossRef]
- French, K.; Pellow, B.; Henderson, M. Vegetation of the Holsworthy Military Area. Cunninghamia 2001, 6, 893–940. [Google Scholar]
- Douglas, P.P.; Shaw, R.B. Rediscovery of Tetramolopium arenarium Subsp. arenarium var. arenarium (Asteraceae: Astereae) on the Pohakuloa Training Area, Hawaii. Ann. Mo. Bot. Gard. 1989, 76, 1182–1185. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie; Wien: New York, NY, USA, 1964; pp. 2–865. [Google Scholar] [CrossRef]
- Király, G. (Ed.) Új Magyar Füvészkönyv. Magyarország Hajtásos Növényei. Határozókulcsok [New Hungarian Herbal. The Vascular Plants of Hungary. Identification Key]; ANP Igazgatóság: Jósvafő, Hungary, 2009; pp. 3–456.
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 20 February 2023).
- Addinsoft XLSTAT 2016: Data Analysis and Statistical Solution for Microsoft Excel. Long Island, NY, USA. Available online: https://www.xlstat.com/fr/ (accessed on 5 March 2023).
- Prach, K.; Pyšek, P.; Šmilauer, P. Changes in Species Traits during Succession: A Search for Pattern. Oikos 1997, 79, 201–205. [Google Scholar] [CrossRef]
- Fábián, Á.P.; Matyasovszky, I. Analysis of climate change in Hungary according to an extended Köppen classification system, 1971–2060. Időjárás 2010, 114, 251–263. [Google Scholar]
- Szentes, S.; Sutyinszki, Z.; Kiss, T.; Fűrész, A.; Saláta, D.; Harkányiné Székely, Z.; Penksza, K. Verges as Fragments of Loess Grasslands in the Carpathian Basin and Their Festuca Species. Diversity 2022, 14, 510. [Google Scholar] [CrossRef]
- Süveges, K. Data on the flora of the Danube–Tisza Interfluve (Hungary). Bot. Commun. 2023, 110, 111–154. [Google Scholar] [CrossRef]
- Latzel, V.; Klimešová, J.; Doležal, J.; Pyšek, P.; Tackenberg, O.; Prach, K. The Association of Dispersal and Persistence Traits of Plants with Different Stages of Succession in Central European Man-Made Habitats. Folia Geobot. 2011, 46, 289–302. [Google Scholar] [CrossRef]
- Penksza, K.; Pápay, G.; Csontos, P. Syntaxonomical analysis of sandy grassland vegetation dominated by Festuca vaginata and F. pseudovaginata in the Pannonian basin. Hacquetia 2021, 20, 217–224. [Google Scholar] [CrossRef]
- Nzei, J.M.; Ngarega, B.K.; Mwanzia, V.M.; Musili, P.M.; Wang, Q.F.; Chen, J.M. The past, current, and future distribution modeling of four water lilies (Nymphaea) in Africa indicates varying suitable habitats and distribution in climate change. Aquat. Bot. 2021, 173, 103416. [Google Scholar] [CrossRef]
- Mkala, E.M.; Mutinda, E.S.; Wanga, V.O.; Oulo, M.A.; Oluoch, W.A.; Nzei, J.; Wasva, E.N.; Odago, W.; Nanjala, C.; Mwachala, G.; et al. Modeling impacts of climate change on the potential distribution of three endemic Aloe species critically endangered in East Africa. Ecol. Inf. 2022, 71, 101765. [Google Scholar] [CrossRef]
- Wan, J.N.; Mbari, N.J.; Wang, S.W.; Liu, B.; Mwangi, B.N.; Rasoarahona, J.R.E.; Xin, H.P.; Zhou, Y.D.; Wang, Q.F. Modeling impacts of climate change on the potential distribution of six endemic baobab species in Madagascar. Plant Divers. 2021, 43, 117–124. [Google Scholar] [CrossRef] [PubMed]
- IPCC Sixth Assessment Report—Climate Change 2023 [EB/OL]. Available online: https://www.ipcc.ch/ (accessed on 26 May 2023).
- Yan, X.; Wang, S.; Duan, Y.; Han, J.; Huang, D.; Zhou, J. Current and future distribution of the deciduous shrub Hydrangea macrophylla in China estimated by MaxEnt. Ecol. Evol. 2021, 11, 16099–16112. [Google Scholar] [CrossRef] [PubMed]
- Wanga, V.O.; Ngarega, B.K.; Oulo, M.A.; Mkala, E.M.; Ngumbau, V.M.; Onjalalaina, G.E.; Odago, W.O.; Nanjala, C.; Ochieng, C.O.; Gichua, M.K.; et al. Projected impacts of climate change on the habitat of Xerophyta species in Africa. Plant Divers. 2024, 46, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Rybansky, M. Determination the ability of military vehicles to override vegetation. J. Terramech 2020, 91, 129–138. [Google Scholar] [CrossRef]
- Gilewitch, D.A.; King, W.C.; Palka, E.J.; Harmon, R.S.; McDonald, E.V.; Doe III, W.W. Characterizing the desert environment for Army operations. In Military Geosciences in the Twenty-First Century: Geological Society of America Reviews in Engineering Geology; Harmon, R.S., Baker, S.E., McDonald, E.V., Eds.; Geological Society of America: Boulder, CO, USA, 2014; Volume 22, pp. 57–68. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, B.; Zhang, M.; Jie, M.; Guo, S.; Wang, Y. Relict Plants Are Better Able to Adapt to Climate Change: Evidence from Desert Shrub Communities. Plants 2023, 12, 4065. [Google Scholar] [CrossRef] [PubMed]
- Bhat, I.A.; Fayaz, M.; Qadir, R.; Rafiq, S.; Guleria, K.; Quadir, J.; Wani, T.A.; Kaloo, Z.A. Predicting potential distribution and range dynamics of Aquilegia fragrans under climate change: Insights from ensemble species distribution modelling. Environ. Monit. Assess. 2023, 195, 623. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, S.; Feng, J.; Ge, J.; Wang, T. Free-ranging livestock changes the acoustic properties of summer soundscapes in a Northeast Asian temperate forest. Biol. Conserv. 2023, 282, 110123. [Google Scholar] [CrossRef]
- Fausett, S.R.; Sandjak, A.; Billard, B.; Braendle, C. Higher-order epistasis shapes natural variation in germ stem cell niche activity. Nat. Commun. 2023, 14, 2824. [Google Scholar] [CrossRef]
- Varol, T.; Cetin, M.; Ozel, H.B.; Sevik, H.; Zeren Cetin, I. The Effects of Climate Change Scenarios on Carpinus betulus and Carpinus orientalis in Europe. Water Air Soil. Pollut. 2022, 233, 45. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic Species in Open Sandy Grassland | Characteristic Species in Closed Natural Sandy Grassland | Weed Species | Disturbance-Tolerant Species | ||||
|---|---|---|---|---|---|---|---|
| Acinos arvensis (Lam.) Dandy | Adonis vernalis L. | Arabis hirsuta (L.) Scop. | Arabis turrita L. | Ambrosia artemisiifolia L. | Anthemis ruthenica M.Bieb. | Achillea pannonica Scheele | Anchusa officinalis L. |
| Allium sphaerocephalon L. | Alyssum alyssoides (L.) L. | Arrhenatherum elatius (L.) P. Beauv. ex J. Presl & C. Presl | Asparagus officinalis L. | Apera spica-venti (L.) P.Beauv. | Agropyron repens (L.) P. Beauv. | Arabidopsis thaliana (L.) Heynh. | Bromus tectorum L. |
| Alyssum tortuosum Willd. | Artemisia campestris L. | Astragalus onobrychis L. | Berberis vulgaris L. | Camelina microcarpa Andrz. ex DC. | Capsella bursa-pastoris (L.) Medik. | Calamagrostis epigejos (L.) Roth | Bothriochloa ischaemum (L.) Keng |
| Arenaria serpyllifolia L. (* A–D: 1–20%) | Bromus squarrosus L. | Bromus erectus Huds. (A–D: 4–10%) | Carex humilis Leyss. | Carduus acanthoides L. | Carex stenophylla Wahlenb. | Centaurea micranthos (Griseb.) Hayek | Dactylis glomerata L. (A−D: 2–10%) |
| Carex liparocarpos Gaudin | Centaurea arenaria M.Bieb. ex Willd. | Dianthus giganteiformis subsp. pontederae (A.Kern.) Soó | Dorycnium germanicum (Gremli) Rikli | Cenchrus incertus M. A. Curtis | Crepis rhoeadifolia M. Bieb. | Crataegus monogyna Jacq. | Cynodon dactylon (L.) Pers. |
| Cerastium semidecandrum L. | Dianthus serotinus Waldst. & Kit. | Euonymus europaeus L. | Festuca arundinacea Schreb. (A−D: 2–8%) | Crepis tectorum L. | Descurainia sophia (L.) Webb ex Prantl | Eryngium campestre L. | Erysimum diffusum Ehrh. |
| Erophila verna (L.) CHEV. | Euphorbia seguieriana Neck. | Festuca rupicola Heuff. (A−D: 2–15%) | Festuca rubra L. (A−D: 2–10%) | Echium vulgare L. | Erodium cicutarium (L.) L’Hér. | Euphorbia cyparissias L. | Festuca pseudovina Hack. ex Wiesb. (A−D: 2–15%) |
| Festuca pseudovaginata Penksza (A−D: 10–25%) | Festuca vaginata Waldst. & Kit. ex Willd. (A−D: 8–65%) | Filipendula vulgaris Moench | Helianthemum ovatum (Viv.) Dunal | Lamium amplexicaule L. | Lithospermum arvense Thunb. | Gagea pusilla (F.W.Schmidt) Sweet | Galium verum L. |
| Fumana procumbens (Dunal) Gren. & Godr. | Gypsophila fastigiata L. | Helictotrichon adsurgens (Schur ex Simonk.) Conert | Helictotrichon pubescens (Huds.) Pilg. | Melandrium album (Mill.) Garcke | Reseda lutea L. | Hypericum perforatum L. | Knautia arvensis (L.) Coult. |
| Gypsophila paniculata L. | Helichrysum arenarium (L.) Moench | Koeleria cristata (L.) Pers. em. Borbás ex Domin | Linaria genistifolia (L.) Mill. | Setaria viridis (L.) P. Beauv. | Taraxacum officinale F. H. Wigg. | Lotus corniculatus L. | Ononis spinosa L. |
| Holosteum umbellatum L. | Iris arenaria Waldst. & Kit. | Linum austriacum L. | Muscari racemosum Mill. | Tribulus terrestris L. | Ornithogalum umbellatum L. | Plantago lanceolata L. | |
| Iris pumila L. | Koeleria glauca (Spreng.) DC. | Peucedanum oreoselinum (L.) Moench (A−D: 2–15%) | Phleum phleoides (L.) H.Karst. | Poa angustifolia L. (A−D: 1–15%) | Rumex acetosella L. | ||
| Koeleria majoriflora (Borbás) Borbás ex Domin | Medicago minima (L.) Bartal. | Polygonatum odoratum (Mill.) Druce | Potentilla arenaria Borkh. | Salsola kali L. | Scabiosa ochroleuca L. | ||
| Minuartia glomerata (M.Bieb.) Degen | Minuartia verna (L.) Hiern | Stachys recta L. | Teucrium chamaedrys L. | Stellaria media (L.) Vill. | Stipa capillata L. | ||
| Myosotis stricta Link | Oxytropis pilosa (L.) DC. | Thalictrum minus L. | Thesium arvense Horv. | Trifolium campestre Schreb. | Veronica arvensis L. | ||
| Plantago indica L. | Poa bulbosa L. (A−D: 1–10%) | Verbascum lychnitis L. | Thymus glabrescens Willd. | Vicia angustifolia L. | Vicia hirsuta (L.) Gray | ||
| Polygonum arenarium W et K. | Sanguisorba minor Scop. | Veronica spicata L. | Vicia tetrasperma (L.) Schreb. | ||||
| Saxifraga tridactylites L. | Secale sylvestre Host | ||||||
| Silene conica L. | Silene otites (L.) Wibel | ||||||
| Solidago virgaurea L. | Stipa borysthenica Klokov ex Prokudin (A−D: 2–25%) | ||||||
| Veronica dillenii Crantz | Vincetoxicum hirundinaria Medik. | ||||||
| Viola kitaibeliana Schult. | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajnok, M.; Penksza, K.; Fűrész, A.; Penksza, P.; Csontos, P.; Szentes, S.; Stilling, F.; Saláta-Falusi, E.; Fuchs, M.; Melenya, C.; et al. Military Activity Impact on Vegetation in Pannonian Dry Sandy Grasslands. Land 2024, 13, 252. https://doi.org/10.3390/land13020252
Bajnok M, Penksza K, Fűrész A, Penksza P, Csontos P, Szentes S, Stilling F, Saláta-Falusi E, Fuchs M, Melenya C, et al. Military Activity Impact on Vegetation in Pannonian Dry Sandy Grasslands. Land. 2024; 13(2):252. https://doi.org/10.3390/land13020252
Chicago/Turabian StyleBajnok, Márta, Károly Penksza, Attila Fűrész, Péter Penksza, Péter Csontos, Szilárd Szentes, Ferenc Stilling, Eszter Saláta-Falusi, Márta Fuchs, Caleb Melenya, and et al. 2024. "Military Activity Impact on Vegetation in Pannonian Dry Sandy Grasslands" Land 13, no. 2: 252. https://doi.org/10.3390/land13020252
APA StyleBajnok, M., Penksza, K., Fűrész, A., Penksza, P., Csontos, P., Szentes, S., Stilling, F., Saláta-Falusi, E., Fuchs, M., Melenya, C., Házi, J., Balogh, D., & Wagenhoffer, Z. (2024). Military Activity Impact on Vegetation in Pannonian Dry Sandy Grasslands. Land, 13(2), 252. https://doi.org/10.3390/land13020252