
Integrated Land-Use Systems Contribute to Restoring Water Cycles in the Brazilian Cerrado Biome

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
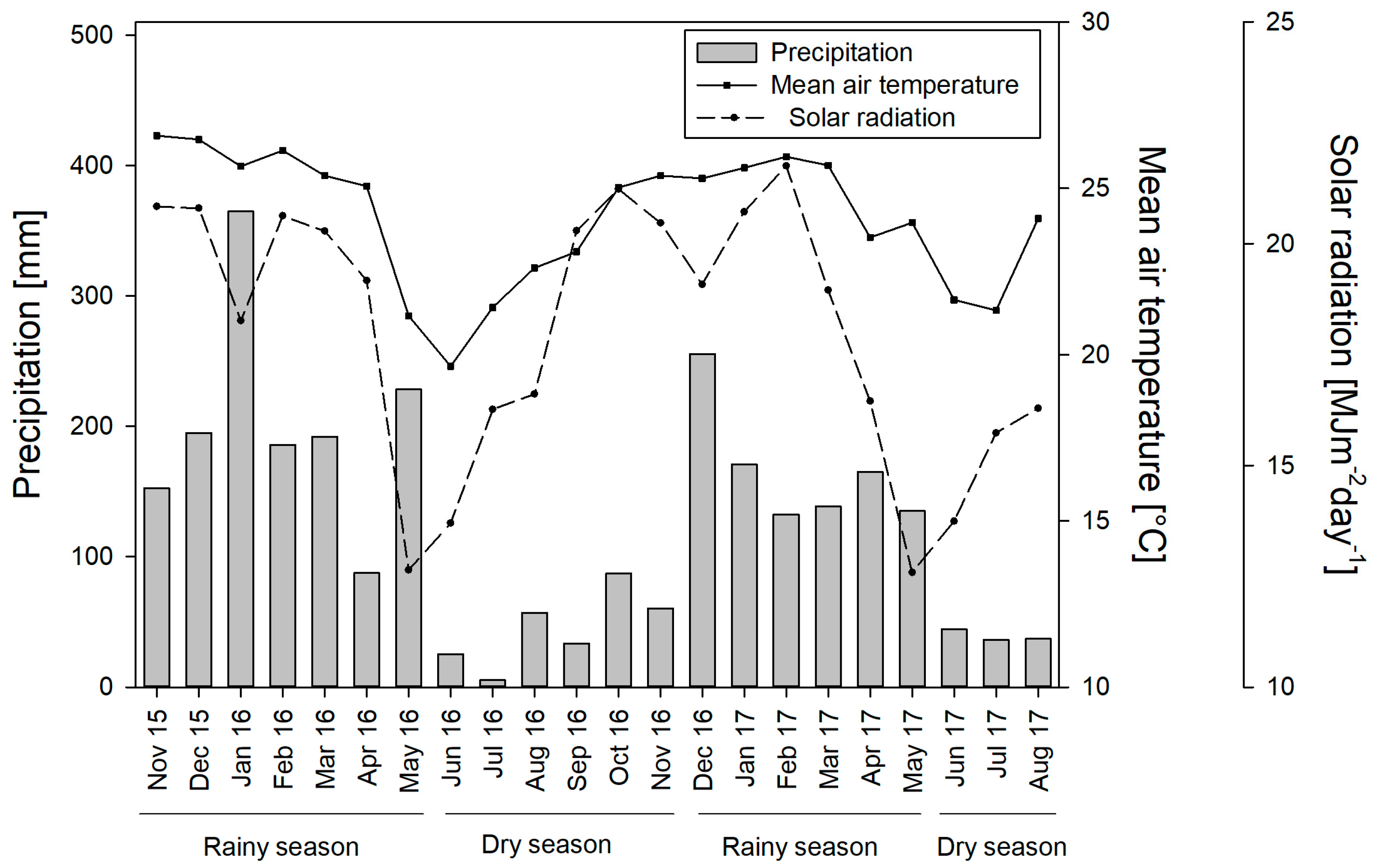
2.1. Experimental Area
2.2. Treatments
2.3. Soil Characteristics
2.4. Soil Moisture and Aboveground Biomass Measurements
2.5. Data Analysis
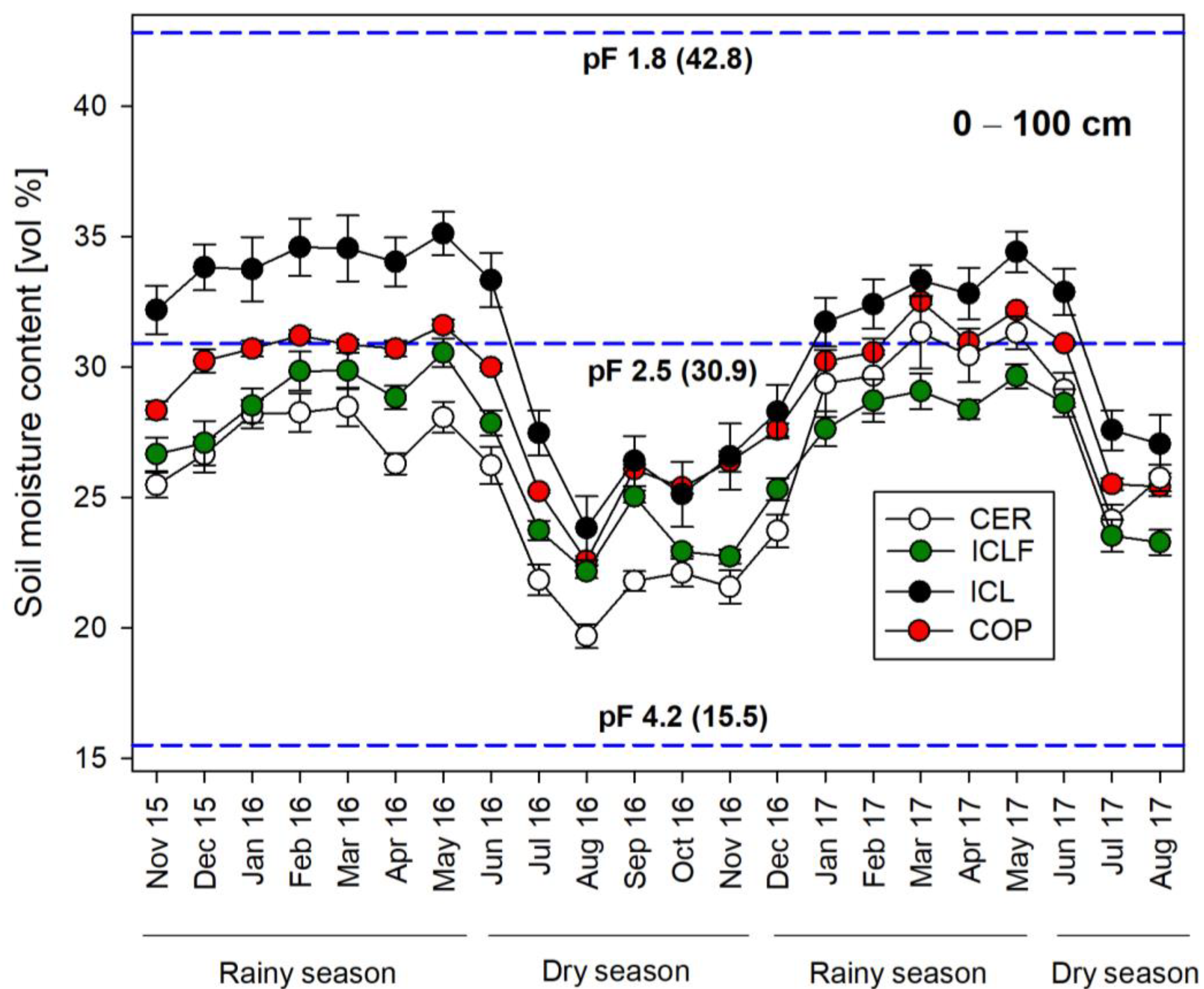
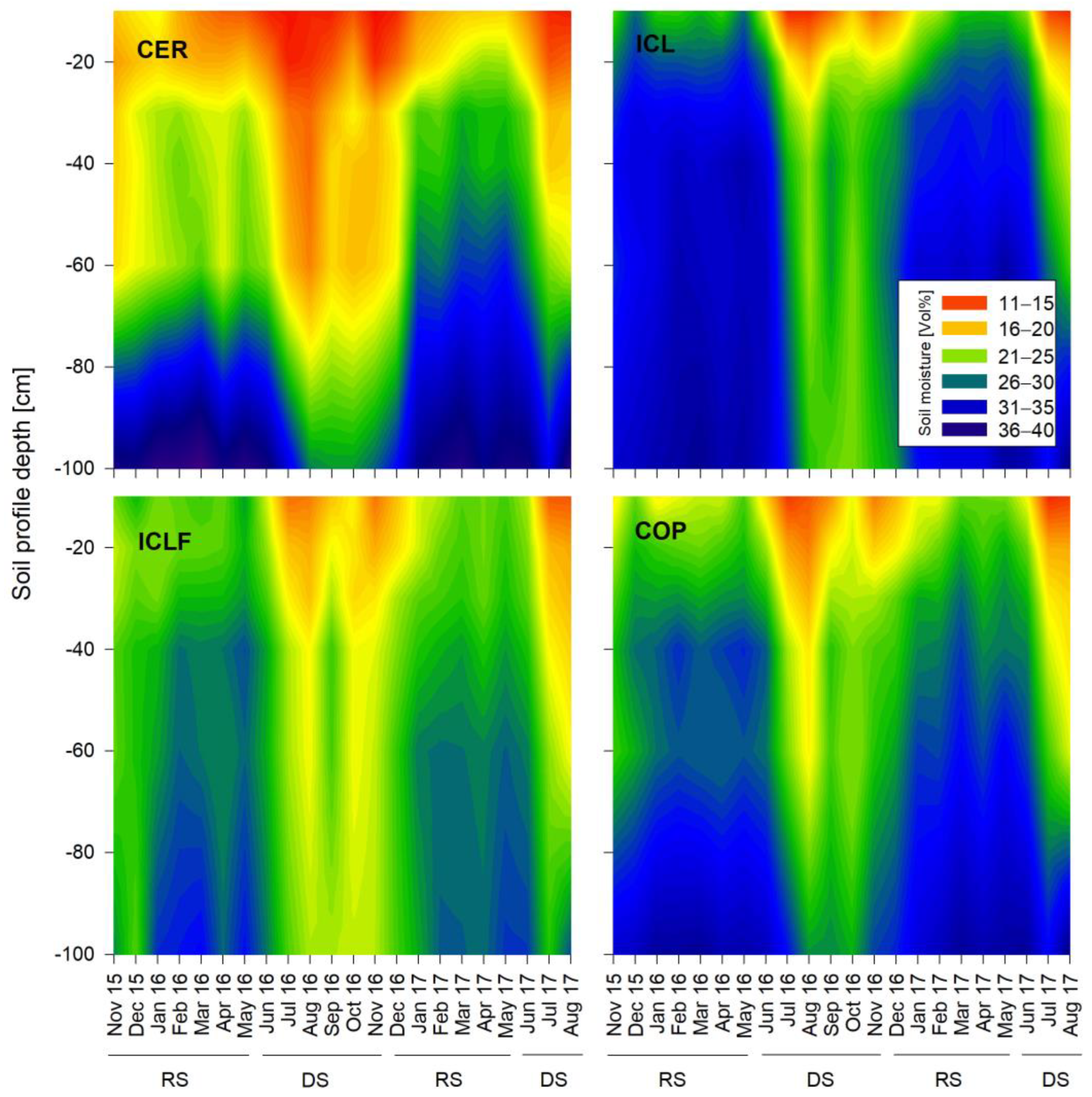
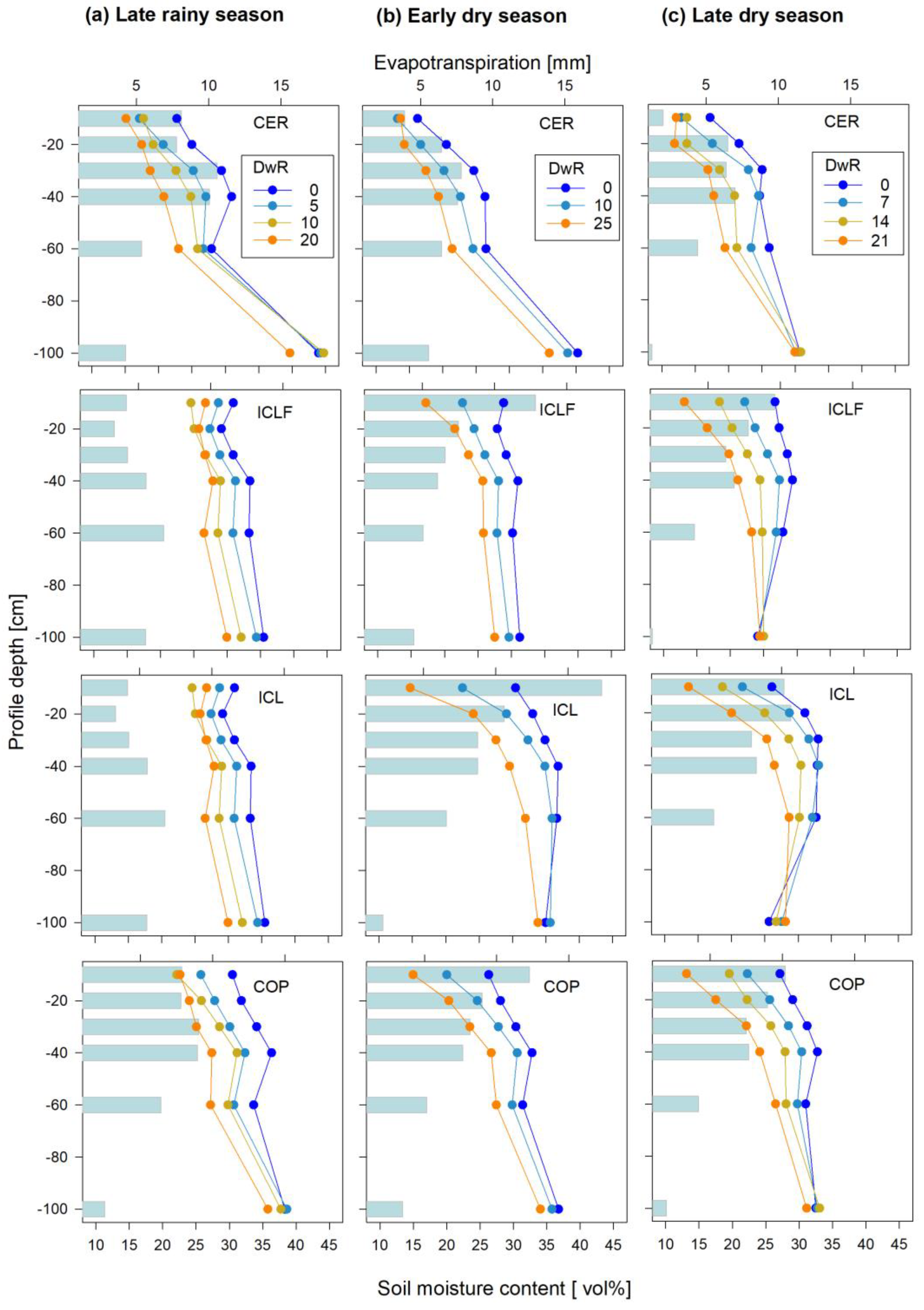
3. Results
3.1. Seasonal Dynamics of Soil Moisture Content
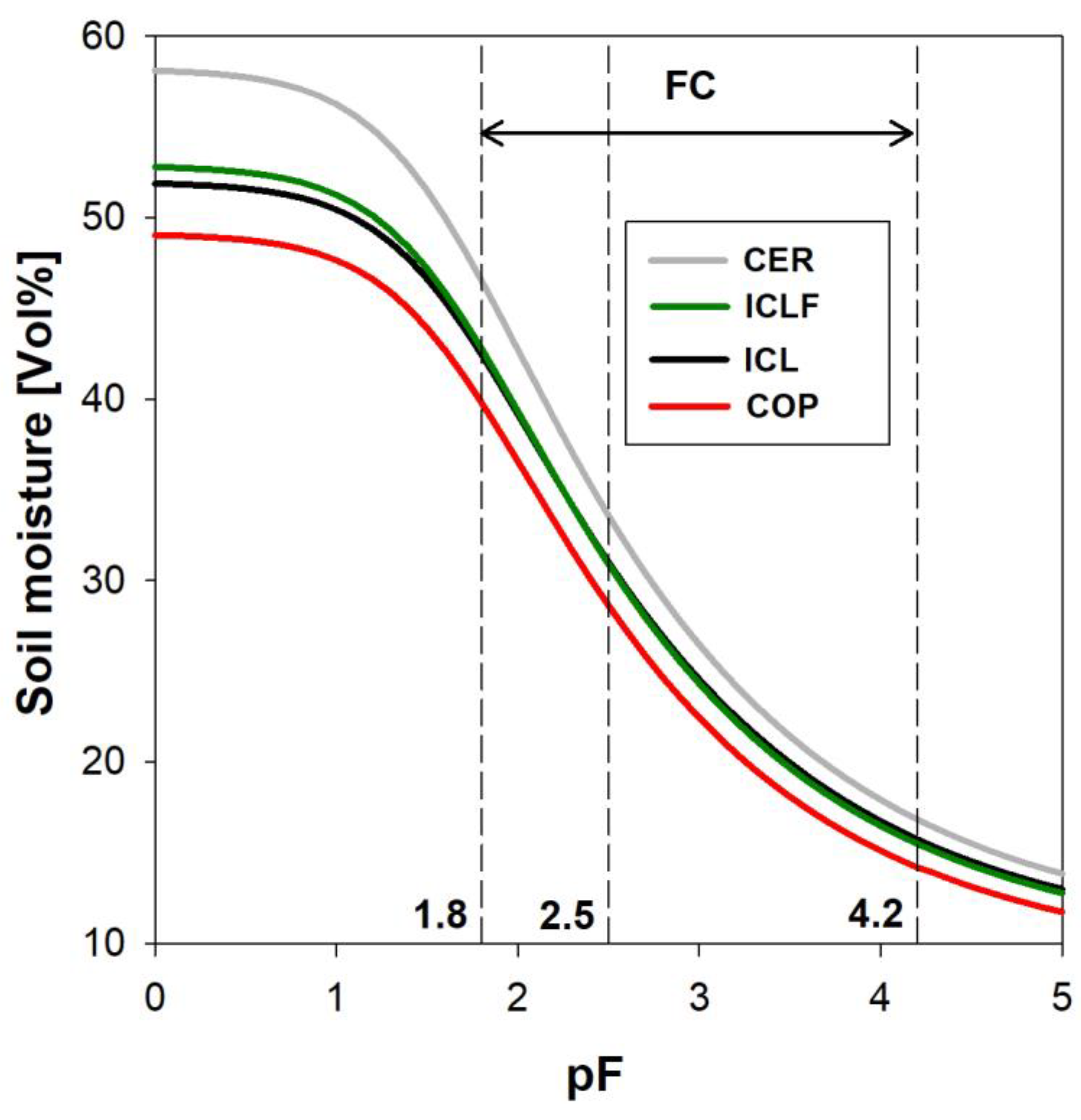
3.2. Soil Moisture Kinetics during Drying Events
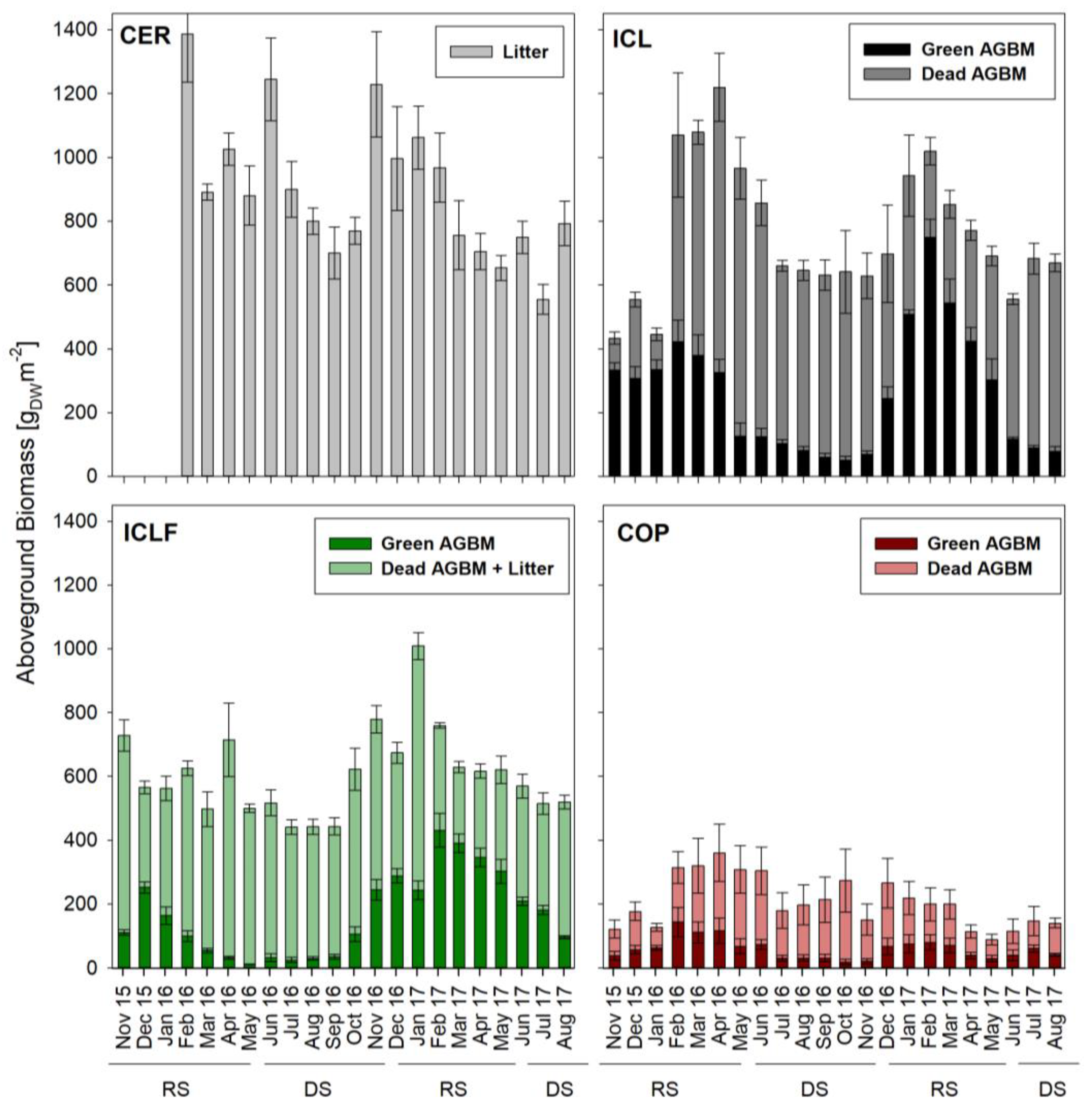
3.3. Aboveground Biomass
4. Discussion
4.1. Soil Moisture Dynamics in the Different Land Use Systems
4.2. Seasonal and Vertical-Spatial Soil Moisture Variations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunke, P.; Mueller, E.N.; Schröder, B.; Zeilhofer, P. The Brazilian Cerrado: Assessment of water and soil degradation in catchments under intensive agricultural use: Ecohydrological assessment of the Brazilian Cerrado. Ecohydrology 2015, 8, 1154–1180. [Google Scholar] [CrossRef]
- Sano, E.E.; Rosa, R.; Brito, J.L.S.; Ferreira, L.G. Land cover mapping of the tropical savanna region in Brazil. Environ. Monit. Assess. 2010, 166, 113–124. [Google Scholar] [CrossRef]
- Klink, C.A.; Machado, R.B. Conservation of the Brazilian Cerrado. Conserv. Biol. 2005, 19, 707–713. [Google Scholar] [CrossRef]
- Marris, E. The forgotten ecosystem. Nature 2005, 437, 944–945. [Google Scholar] [CrossRef]
- Spera, S.A.; Galford, G.L.; Coe, M.T.; Macedo, M.N.; Mustard, J.F. Land-use change affects water recycling in Brazil’s last agricultural frontier. Glob. Chang. Biol. 2016, 22, 3405–3413. [Google Scholar] [CrossRef]
- Oliveira, P.T.S.; Nearing, M.A.; Moran, M.S.; Goodrich, D.C.; Wendland, E.; Gupta, H.V. Trends in water balance components across the Brazilian Cerrado. Water Resour. Res. 2014, 50, 7100–7114. [Google Scholar] [CrossRef]
- Butt, N.; de Oliveira, P.A.; Costa, M.H. Evidence that deforestation affects the onset of the rainy season in Rondonia, Brazil. J. Geophys. Res. Atmos. 2011, 116, D11120. [Google Scholar] [CrossRef]
- Oliveira, R.S.; Bezerra, L.; Davidson, E.A.; Pinto, F.; Klink, C.A.; Nepstad, D.C.; Moreira, A. Deep root function in soil water dynamics in cerrado savannas of central Brazil. Funct. Ecol. 2005, 19, 574–581. [Google Scholar] [CrossRef]
- Quesada, C.A.; Hodnett, M.G.; Breyer, L.M.; Santos, A.J.B.; Andrade, S.; Miranda, H.S.; Miranda, C.; Lloyd, J. Seasonal variations in soil water in two woodland savannas of central Brazil with different fire history. Tree Phys. 2008, 28, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Balbino, L.C.; Cordeiro, L.A.M.; Porfírio-da-Silva, V.; Moraes, A.de; Martínez, G.B.; Alvarenga, R.C.; Kichel, A.N.; Fontaneli, R.S.; dos Santos, H.P.; Franchini, J.C.; et al. Evolução tecnológica e arranjos produtivos de sistemas de integração lavoura-pecuária-floresta no Brasil. Pesqui. Agropecuária Bras. 2011, 46, 1–12. [Google Scholar] [CrossRef]
- Bono, J.A.M.; Macedo, M.C.M.; Tormena, C.A.; Nanni, M.R.; Gomes, E.P.; Müller, M.M.L. Infiltração de água no solo em um latossolo vermelho da região sudoeste dos cerrados com diferentes sistemas de uso e manejo. Rev. Bras. Ciência Solo 2012, 36, 1845–1853. [Google Scholar] [CrossRef][Green Version]
- Nair, P.K.R. An Introduction to Agroforestry; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1993; p. 499. [Google Scholar]
- Bosi, C.; Pezzopane, J.R.M.; Sentelhas, P.C. Soil water availability in a full sun pasture and in a silvopastoral system with eucalyptus. Agrofor. Syst. 2020, 94, 429–440. [Google Scholar] [CrossRef]
- Glatzle, S.; Stuerz, S.; Giese, M.; Pereira, M.; de Almeida, R.G.; Bungenstab, D.J.; Macedo, M.C.M.; Asch, F. Seasonal Dynamics of Soil Moisture in an Integrated-Crop-Livestock-Forestry System in Central-West Brazil. Agriculture 2021, 11, 245. [Google Scholar] [CrossRef]
- Guenni, O.; Marín, D.; Baruch, Z. Responses to drought of five Brachiaria species. I. Biomass production, leaf growth, root distribution, water use and forage quality. Plant Soil 2002, 243, 229–241. [Google Scholar] [CrossRef]
- Jha, A.; Bonetti, S.; Smith, A.P.; Souza, R.; Calabrese, S. Linking soil structure, hydraulic properties, and organic carbon dynamics: A holistic framework to study the impact of climate change and land management. J. Geophys. Res. Biogeosci. 2023, 128, e2023JG007389. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, update 2015. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- de Sousa Neto, E.L.; Andrioli, I.; de Almeida, R.G.; Macedo, M.C.M.; Lal, R. Physical quality of an Oxisol under an integrated crop-livestock-forest system in the Brazilian Cerrado. Rev. Bras. Ciência Solo 2014, 38, 608–618. [Google Scholar] [CrossRef]
- Schaap, M.G.; Leij, F.J.; van Genuchten, M.T. Rosetta: A computer program for estimating soil hydraulic parameters with hierarchical pedotransfer functions. J. Hydrol. 2001, 251, 163–176. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (3.5.3) [Computer Software]. R Foundation for Statistical Computing. 2019. Available online: https://www.R-project.org/ (accessed on 15 February 2020).
- Herrera, A.; Urich, R.; Rengifo, E.; Ballestrini, C.; González, A.; León, W. Transpiration in a eucalypt plantation and a savanna in Venezuela. Trees 2012, 26, 1759–1769. [Google Scholar] [CrossRef]
- Minasny, B.; McBratney, A.B. Limited effect of organic matter on soil available water capacity. Eur. J. Soil Sci. 2018, 69, 39–47. [Google Scholar] [CrossRef]
- Iqbal, R.; Raza, M.A.S.; Valipour, M.; Saleem, M.F.; Zaheer, M.S.; Ahmad, S.; Toleikiene, M.; Haider, I.; Aslam, M.U.; Nazar, M.A. Potential agricultural and environmental benefits of mulches—A review. Bull. Natl. Res. Cent. 2020, 44, 75. [Google Scholar] [CrossRef]
- Anache, J.A.A.; Wendland, E.; Rosalem, L.M.P.; Youlton, C.; Oliveira, P.T.S. Hydrological trade-offs due to different land covers and land uses in the Brazilian Cerrado. Hydrol. Earth Syst. Sci. 2019, 23, 1263–1279. [Google Scholar] [CrossRef]
- de Oliveira, O.C.; de Oliveira, I.P.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M. Chemical and biological indicators of decline/degradation of Brachiaria pastures in the Brazilian Cerrado. Agric. Ecosys. Environ. 2004, 103, 289–300. [Google Scholar] [CrossRef]
- Drewry, J.J.; Cameron, K.C.; Buchan, G.D. Pasture yield and soil physical property responses to soil compaction from treading and grazing—A review. Soil Res. 2008, 46, 237–256. [Google Scholar] [CrossRef]
- Greenwood, K.L.; McKenzie, B.M. Grazing effects on soil physical properties and the consequences for pastures: A review. Aus. J. Exp. Agric. 2001, 41, 1231–1250. [Google Scholar] [CrossRef]
- Nóbrega, R.L.B.; Guzha, A.C.; Torres, G.N.; Kovacs, K.; Lamparter, G.; Amorim, R.S.S.; Couto, E.; Gerold, G. Effects of conversion of native Cerrado vegetation to pasture on soil hydro-physical properties, evapotranspiration and streamflow on the Amazonian agricultural frontier. PLoS ONE 2017, 12, e0179414. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.N.; Tanveer, M.; Shahzad, B.; Yang, G.; Fahad, S.; Ali, S.; Bukhari, M.A.; Tung, S.A.; Hafeez, A.; Souliyanonh, B. Soil compaction effects on soil health and crop productivity: An overview. Environ. Sci. Pollut. Res. 2017, 24, 10056–10067. [Google Scholar] [CrossRef]
- He, J.; Li, H.; Rasaily, R.G.; Wang, Q.; Cai, G.; Su, Y.; Qiao, X.; Liu, L. Soil properties and crop yields after 11 years of no tillage farming in wheat–maize cropping system in North China Plain. Soil Tillage Res. 2011, 113, 48–54. [Google Scholar] [CrossRef]
- Strudley, M.W.; Green, T.R.; Ascough, J.C. Tillage effects on soil hydraulic properties in space and time: State of the science. Soil Tillage Res. 2008, 99, 4–48. [Google Scholar] [CrossRef]
- Moreira, G.M.; Neves, J.C.L.; Rocha, G.C.; Magalhães, C.A.d.S.; Farias Neto, A.L.; Meneguci, J.L.P.; Fernandes, R.B.A. Physical quality of soil under a crop-livestock-forestry system in the cerrado/amazon transition region. Rev. Árvore 2018, 42, e420213. [Google Scholar] [CrossRef]
- Assis, P.C.R.; Stone, L.F.; Medeiros, J.C.; Madari, B.E.; Oliveira, J.d.M.; Wruck, F.J. Atributos físicos do solo em sistemas de integração lavoura-pecuária-floresta. Rev. Bras. Eng. Agríc. Ambient. 2015, 19, 309–316. [Google Scholar] [CrossRef]
- Feitosa, C.E.L.; Costa, P.H.d.S.; de Meneses, K.C.; de Oliveira, U.C.; de Farias, M.F. Changes in physical quality of oxisols under different management systems in the Brazilian Cerrado. Eng. Agríc. 2020, 40, 609–616. [Google Scholar] [CrossRef]
- Eiten, G. The cerrado vegetation of Brazil. Bot. Rev. 1972, 38, 201–341. [Google Scholar] [CrossRef]
- Dias, L.C.P.; Macedo, M.N.; Costa, M.H.; Coe, M.T.; Neill, C. Effects of land cover change on evapotranspiration and streamflow of small catchments in the Upper Xingu River Basin, Central Brazil. J. Hydrol. Reg. Stud. 2015, 4, 108–122. [Google Scholar] [CrossRef]
- Magliano, P.N.; Giménez, R.; Houspanossian, J.; Páez, R.A.; Nosetto, M.D.; Fernández, R.J.; Jobbágy, E.G. Litter is more effective than forest canopy reducing soil evaporation in Dry Chaco rangelands. Ecohydrology 2017, 10, e1879. [Google Scholar] [CrossRef]
- Rosalem, L.M.P.; Anache, J.A.A.; Wendland, E. Determining forest litter interception in an area of the Cerrado sensu stricto. RBRH 2018, 23, e26. [Google Scholar] [CrossRef]
- Schunk, C.; Leuchner, M.; Wastl, C.; Ruth, B.; Menzel, A. Comparison of different methods for the in situ measurement of forest litter moisture content. Nat. Hazards Earth Syst. Sci. 2015, 3, 3733. [Google Scholar] [CrossRef]
- Ferreira, M.M.; Fernandes, B.; Curi, N. Mineralogia da fração argila e estrutura de latossolos da região sudeste do Brasil. Rev. Bras. Ciência Solo 1999, 23, 507–514. [Google Scholar] [CrossRef]
- Ferreira, M.M.; Fernandes, B.; Curi, N. Influência da mineralogia da fração argila nas propriedades físicas de latossolos da região sudeste do Brasil. Rev. Bras. Ciência Solo 1999, 23, 515–524. [Google Scholar] [CrossRef]
- Juhász, C.E.P.; Cooper, M.; Cursi, P.R.; Ketzer, A.O.; Toma, R.S. Savanna woodland soil micromorphology related to water retention. Sci. Agric. 2007, 64, 344–354. [Google Scholar] [CrossRef]
- Salako, F.K.; Kirchhof, G. Field hydraulic properties of an Alfisol under various fallow systems in southwestern Nigeria. Soil Use Manag. 2003, 19, 340–346. [Google Scholar] [CrossRef]
- Bouillet, J.-P.; Laclau, J.-P.; Arnaud, M.; M’Bou, A.T.; Saint-André, L.; Jourdan, C. Changes with age in the spatial distribution of roots of Eucalyptus clone in Congo: Impact on water and nutrient uptake. Forest Ecol. Manag. 2002, 171, 43–57. [Google Scholar] [CrossRef]
- Christina, M.; Nouvellon, Y.; Laclau, J.; Stape, J.; Bouillet, J.; Lambais, G.R.; Maire, G. Importance of deep water uptake in tropical eucalypt forest. Funct. Ecol. 2017, 31, 509–519. [Google Scholar] [CrossRef]
- Oliveira, P.T.S.; Wendland, E.; Nearing, M.A.; Perea Martins, J. Interception of rainfall and surface runoff in the Brazilian Cerrado. Geophys. Res. Abstracts 2014, 16, 4780. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | BD | SOC | Texture [%] | Ksat | ||||
|---|---|---|---|---|---|---|---|---|
| Depth [cm] | (CaCl2) | [g cm−3] | [%] | Sand | Silt | Clay | [cm d−1] | |
| Cerrado (CER) | ||||||||
| 10 | 4.3 | 0.91 | 3.13 | 56 | 7 | 37 | SC | 107.1 |
| 20 | 4.3 | 0.94 | 2.08 | 53 | 8 | 39 | SC | 99.8 |
| 30 | 4.4 | 1.00 | 1.68 | 52 | 7 | 41 | SC | 83.4 |
| 40 | 4.6 | 1.05 | 1.59 | 51 | 7 | 42 | SC | 71.9 |
| 60 | - | 0.99 | - | - | - | - | - | 83.6 |
| 100 | - | 1.01 | - | - | - | - | - | 79.7 |
| mean | 4.4 | 0.98 | 2.12 | 53 | 7 | 40 | SC | 87.6 |
| Integrated-crop-livestock-forestry system (ICLF) | ||||||||
| 10 | 5.4 | 1.24 | 1.83 | 56 | 6 | 38 | SC | 46.9 |
| 20 | 5.0 | 1.17 | 1.60 | 55 | 6 | 39 | SC | 55.5 |
| 30 | 4.8 | 1.11 | 1.35 | 54 | 7 | 39 | SC | 65.0 |
| 40 | 4.8 | 1.14 | 1.18 | 55 | 6 | 39 | SC | 60.3 |
| 60 | - | 1.14 | - | - | - | - | - | 60.3 |
| 100 | - | 1.10 | - | - | - | - | - | 67.2 |
| mean | 5.0 | 1.15 | 1.49 | 55 | 6 | 39 | SC | 59.2 |
| Integrated-crop-livestock system (ICL) | ||||||||
| 10 | 5.2 | 1.25 | 2.39 | 53 | 7 | 40 | SC | 41.5 |
| 20 | 5.1 | 1.18 | 1.82 | 53 | 7 | 40 | SC | 51.8 |
| 30 | 5.0 | 1.21 | 1.50 | 53 | 8 | 39 | SC | 47.4 |
| 40 | 4.8 | 1.19 | 1.24 | 52 | 7 | 41 | SC | 49.1 |
| 60 | - | 1.18 | - | - | - | - | - | 50.6 |
| 100 | - | 1.10 | - | - | - | - | - | 64.1 |
| mean | 5.0 | 1.18 | 1.74 | 61 | 5 | 34 | SC | 50.7 |
| Continuous pasture (COP) | ||||||||
| 10 | 4.9 | 1.34 | 1.75 | 64 | 5 | 31 | SCL | 49.1 |
| 20 | 4.8 | 1.24 | 1.51 | 60 | 6 | 34 | SCL | 57.5 |
| 30 | 4.7 | 1.28 | 1.29 | 61 | 4 | 35 | SCL | 52.1 |
| 40 | 4.6 | 1.24 | 1.05 | 59 | 4 | 37 | SC | 47.0 |
| 60 | - | 1.28 | - | - | - | - | - | 47.0 |
| 100 | - | 1.21 | - | - | - | - | - | 56.4 |
| mean | 4.8 | 1.27 | 1.40 | 61 | 5 | 34 | SCL | 52.4 |
| Soil Moisture Content [vol%] | SM = TxM + R | |||
|---|---|---|---|---|
| Df | SS | MS | p | |
| Treatment (T) | 3 | 888 | 296 | <0.001 |
| Month (M) | 21 | 2340 | 111 | <0.001 |
| Replication (R) | 2 | 32 | 16 | <0.001 |
| Interaction (T × M) | 63 | 199 | 3 | <0.001 |
| Residuals | 174 | 226 | 1 | |
| SM | SM = TxMxD + R | |||
|---|---|---|---|---|
| [vol%] | Df | SS | MS | p |
| Treatment (T) | 3 | 6498 | 2166 | <0.001 |
| Month (M) | 21 | 14657 | 698 | <0.001 |
| Depth (D) | 5 | 23817 | 4763 | <0.001 |
| Replication (R) | 2 | 117 | 58 | <0.001 |
| Interaction (T × M) | 63 | 1261 | 20 | <0.001 |
| Interaction (T × D) | 15 | 5623 | 375 | <0.001 |
| Interaction (M × D) | 105 | 2701 | 26 | <0.001 |
| Interaction (T × M × D) | 315 | 893 | 3 | n.s. < 0.064 |
| Residuals | 1054 | 2738 | 3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glatzle, S.; de Almeida, R.G.; Pereira Barsotti, M.; Bungenstab, D.J.; Giese, M.; Macedo, M.C.M.; Stuerz, S.; Asch, F. Integrated Land-Use Systems Contribute to Restoring Water Cycles in the Brazilian Cerrado Biome. Land 2024, 13, 221. https://doi.org/10.3390/land13020221
Glatzle S, de Almeida RG, Pereira Barsotti M, Bungenstab DJ, Giese M, Macedo MCM, Stuerz S, Asch F. Integrated Land-Use Systems Contribute to Restoring Water Cycles in the Brazilian Cerrado Biome. Land. 2024; 13(2):221. https://doi.org/10.3390/land13020221
Chicago/Turabian StyleGlatzle, Sarah, Roberto Giolo de Almeida, Mariana Pereira Barsotti, Davi José Bungenstab, Marcus Giese, Manuel Claudio M. Macedo, Sabine Stuerz, and Folkard Asch. 2024. "Integrated Land-Use Systems Contribute to Restoring Water Cycles in the Brazilian Cerrado Biome" Land 13, no. 2: 221. https://doi.org/10.3390/land13020221
APA StyleGlatzle, S., de Almeida, R. G., Pereira Barsotti, M., Bungenstab, D. J., Giese, M., Macedo, M. C. M., Stuerz, S., & Asch, F. (2024). Integrated Land-Use Systems Contribute to Restoring Water Cycles in the Brazilian Cerrado Biome. Land, 13(2), 221. https://doi.org/10.3390/land13020221