
Testing the Effect of Relative Pollen Productivity on the REVEALS Model: A Validated Reconstruction of Europe-Wide Holocene Vegetation

 ,
,  , ,
, ,  , , , , ,
, , , , ,  , , , , , , , , , , , , , , ,
, , , , , , , , , , , , , , ,  , , , ,
, , , ,  , , , , , , , , , , , , , , , , , , , , , , ,
, , , , , , , , , , , , , , , , , , , , , , ,  , , , , , , , , , , , ,
, , , , , , , , , , , ,  , , , , ,
, , , , ,  ,
,  , , , , , , , , , , , ,
, , , , , , , , , , , ,  , ,
, ,  , , ,
, , ,  , , , , , , , , , , , , , and
, , , , , , , , , , , , , and
Abstract
1. Introduction
2. Methods
2.1. The REVEALS Model
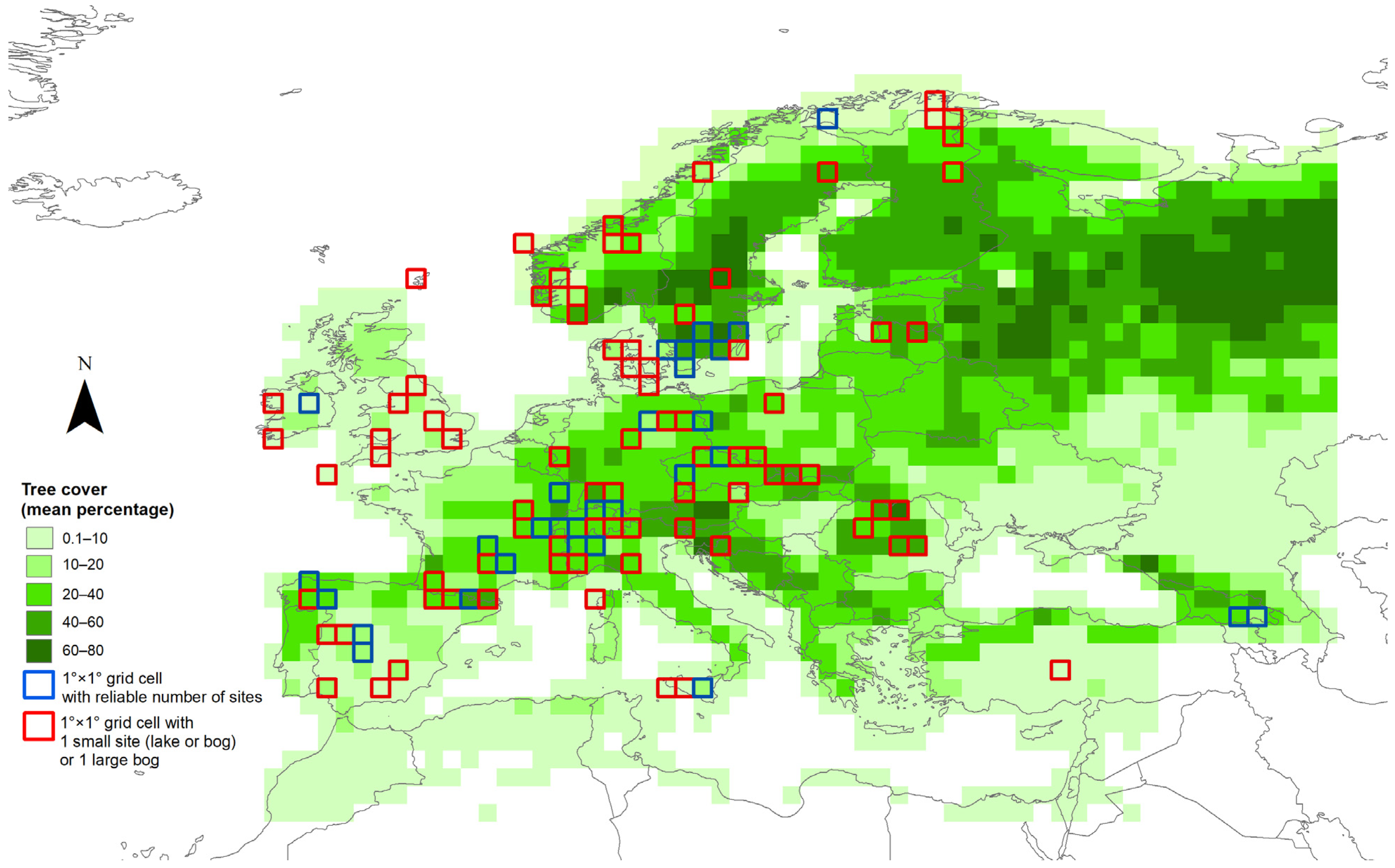
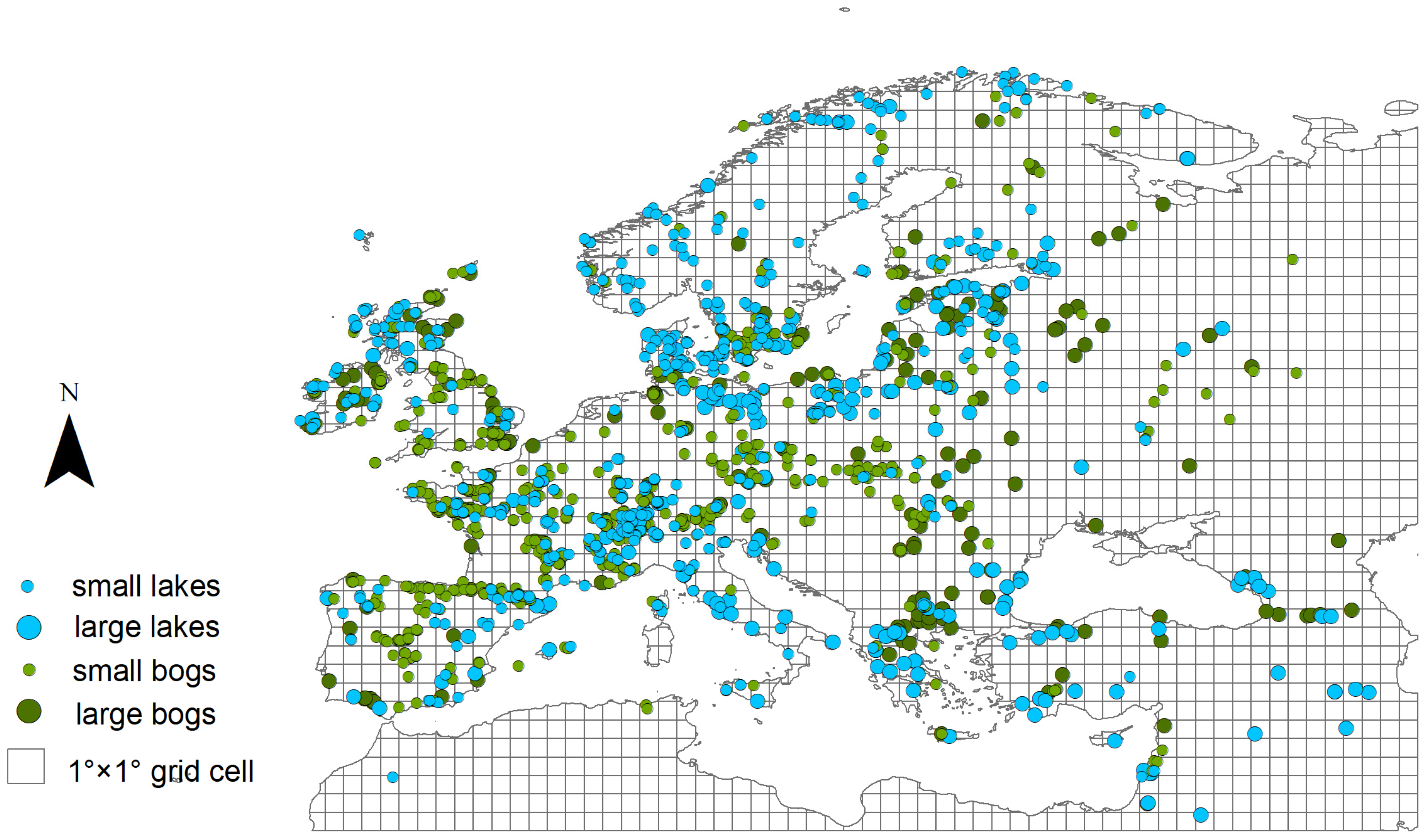
2.2. Fossil Pollen Records: Data Compilation and Preparation
2.3. Modern Vegetation and Pollen Datasets
2.4. REVEALS Run and Data Analysis
3. Results
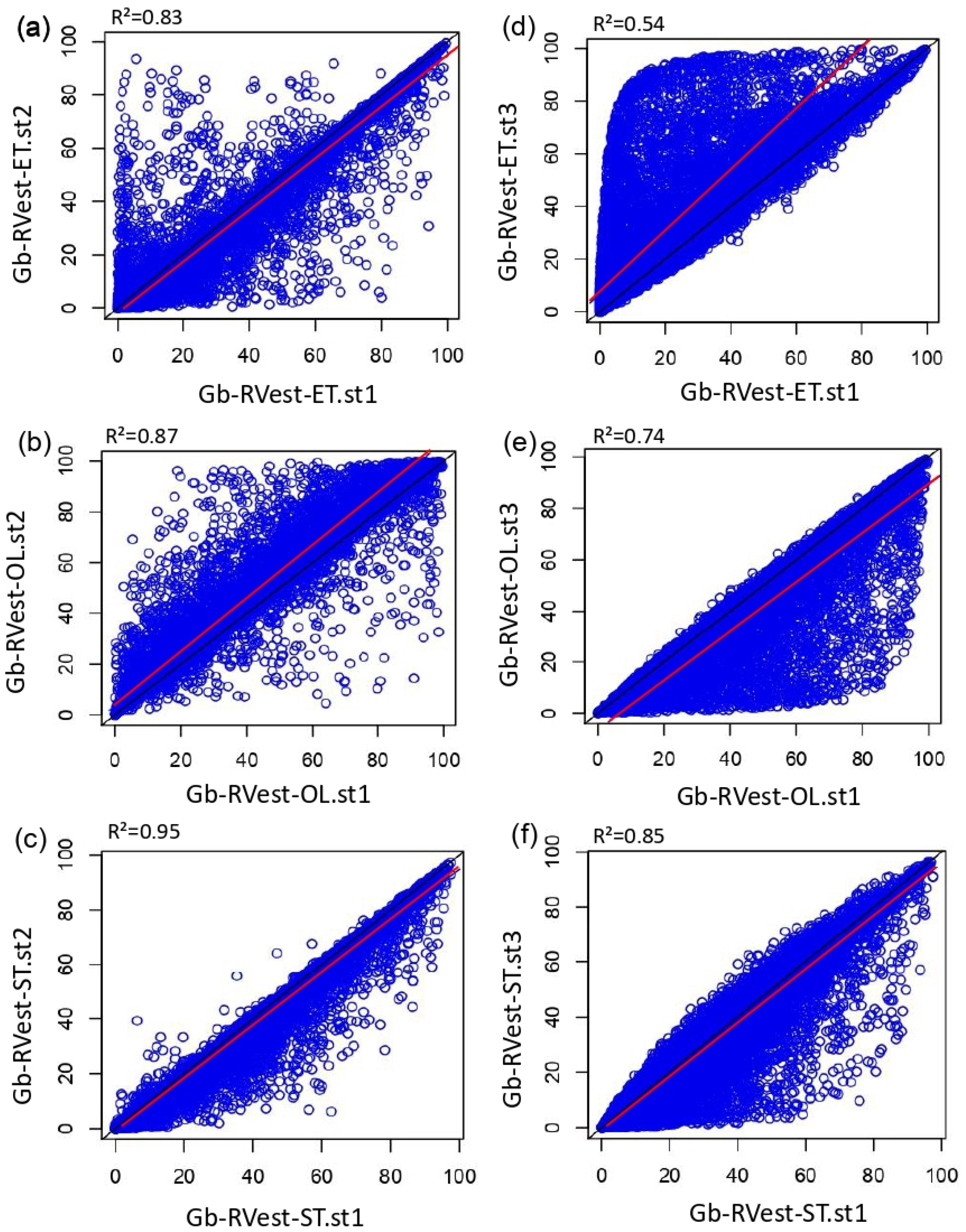
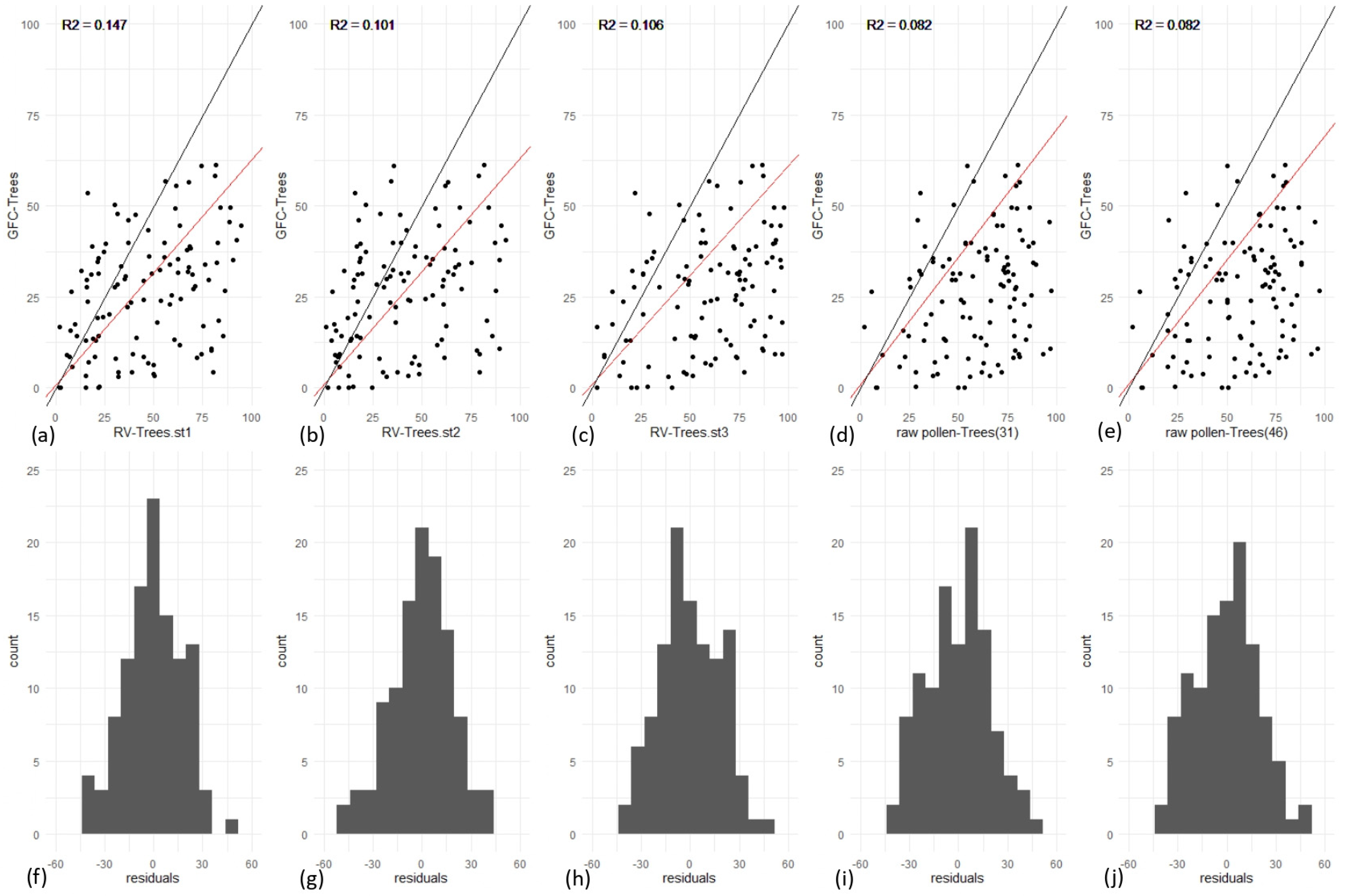
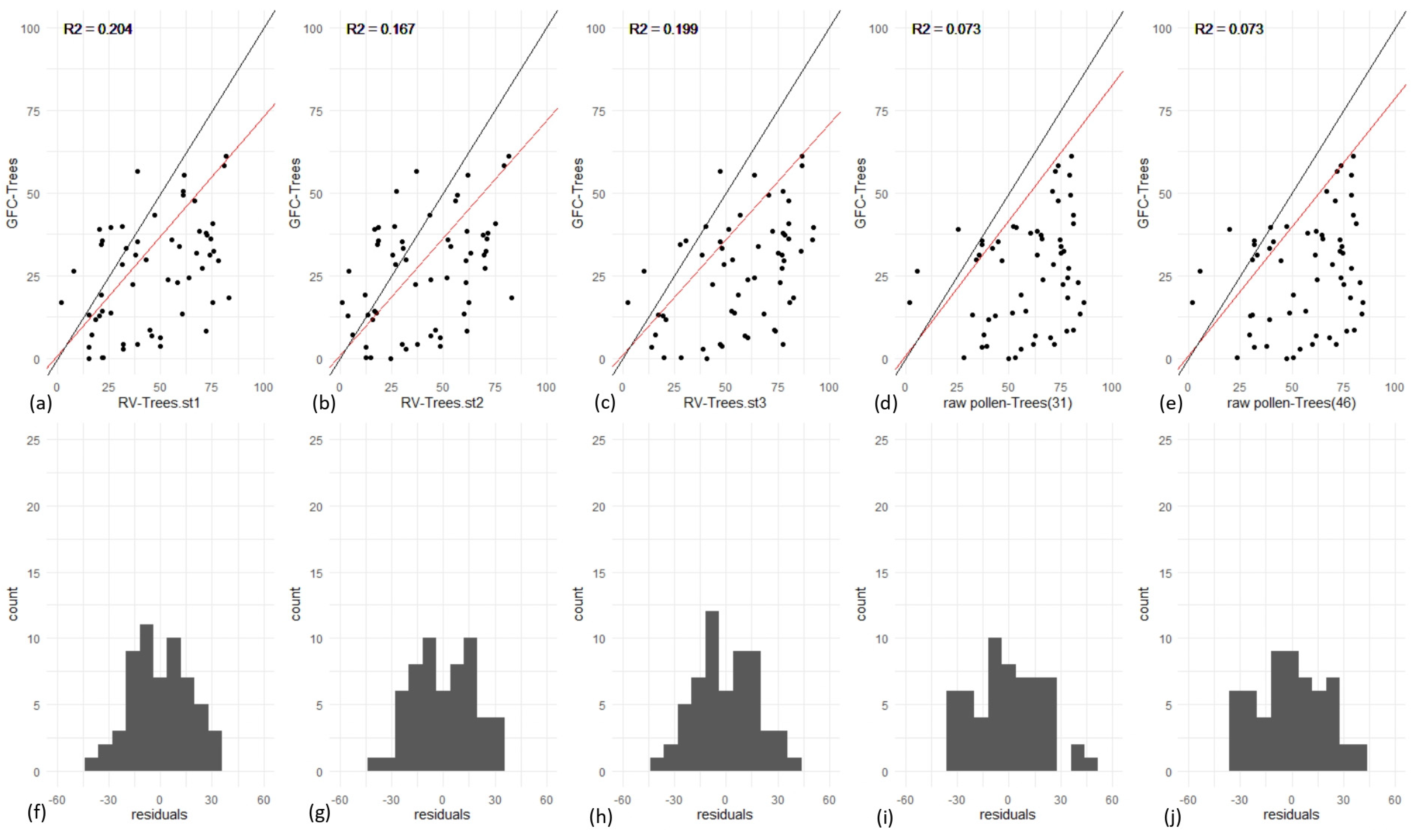
3.1. Effect of Type and Number of Pollen Taxa in RPP-Means Datasets on Gb-RVest-LCTs
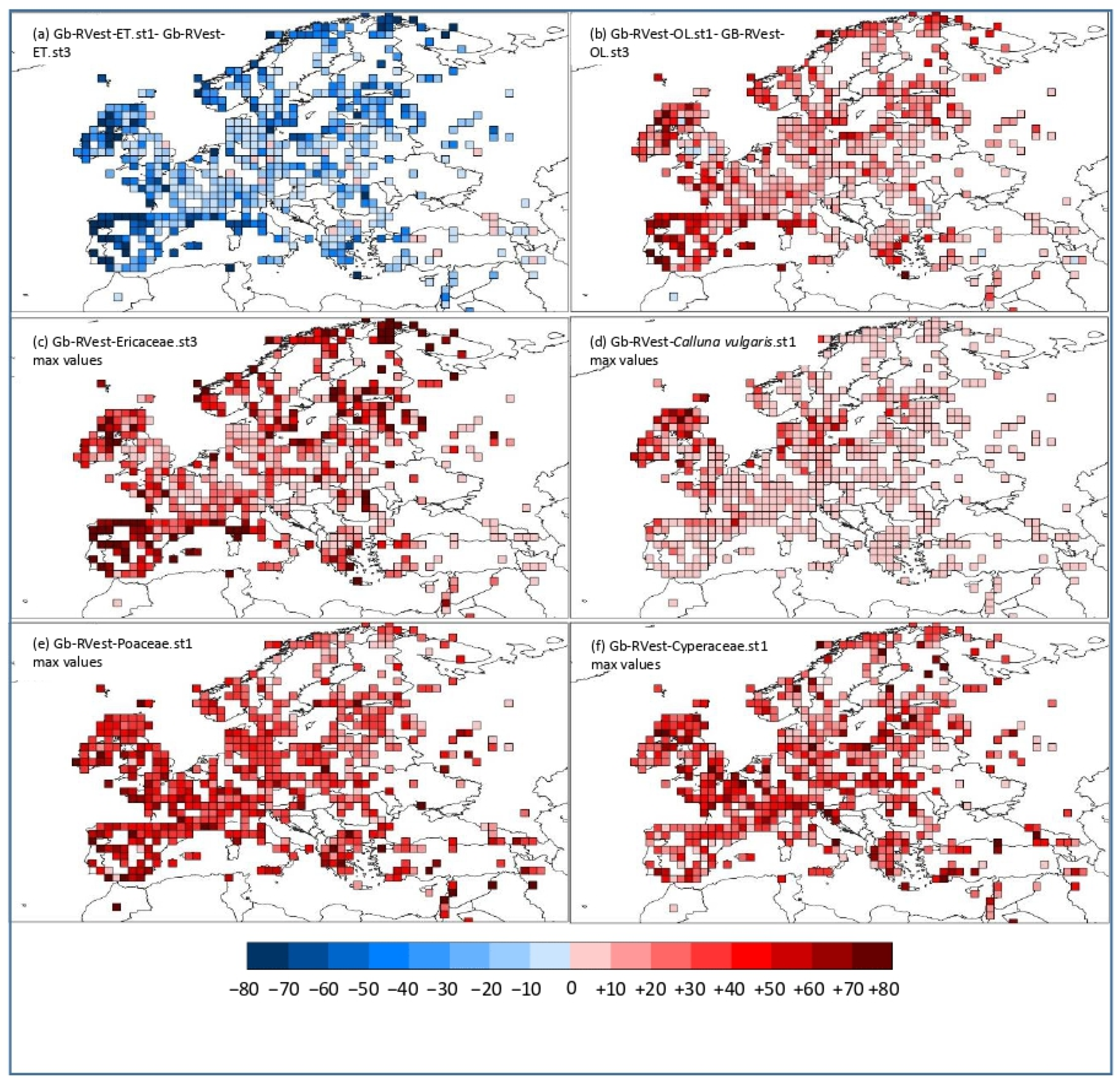
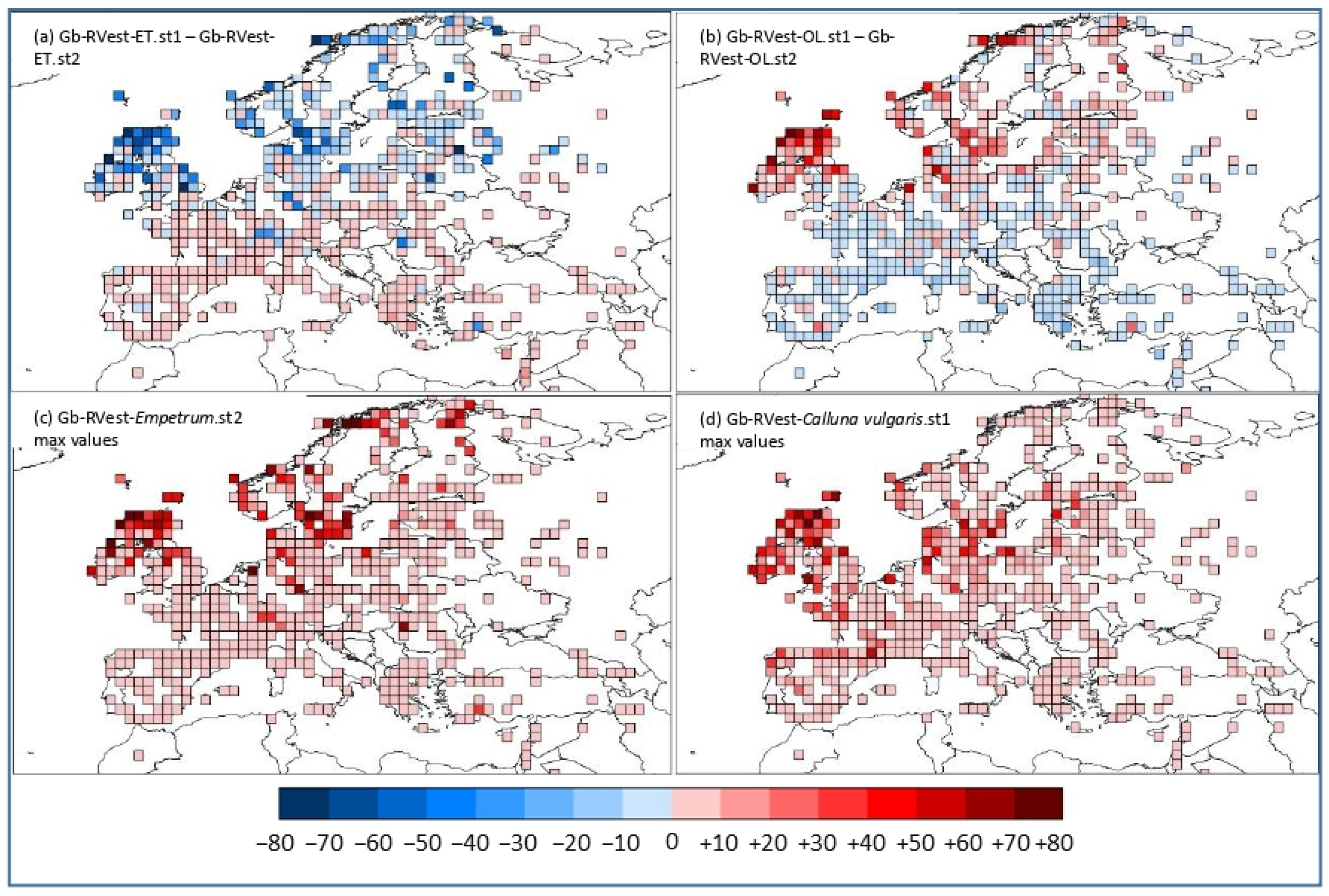
3.2. Geographical Pattern of the Gb-RVest-LCTs Differences between RPP-Means Datasets
3.3. REVEALS Validation for All Europe
4. Discussion
4.1. New Insight after Validation
4.2. Influence of RPPs.sts on REVEALS Model Sensitivity and Pattern of Difference at the Spatial Level
4.3. Challenging Taxa: Ericaceae and Empetrum
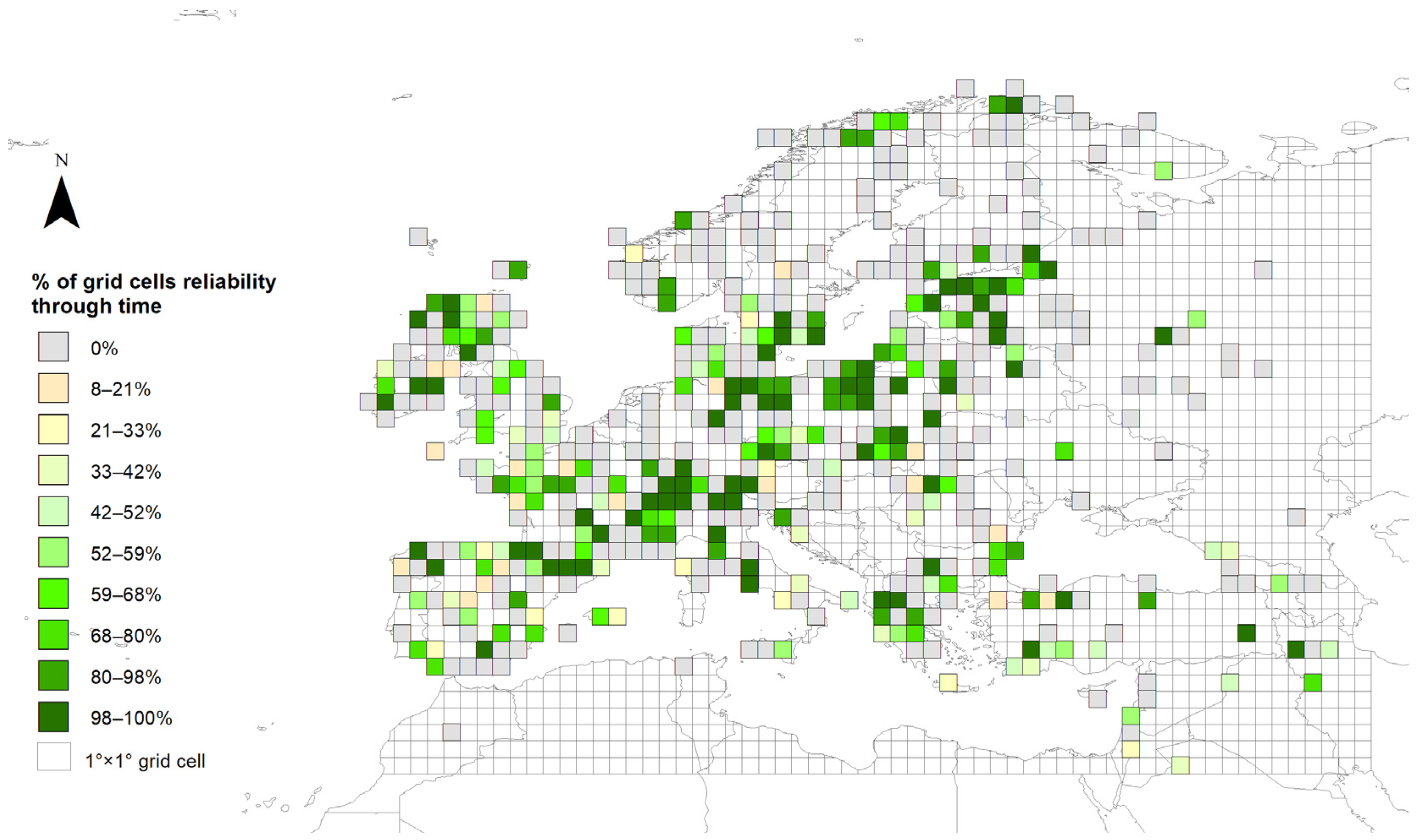
4.4. Importance of the Number of Pollen Records in Europe: Data Reliability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moss Polster Sites Used to Calculate RPPs | Lake Sites Used to Calculate RPPs | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Region | Finland [89] | C Sweden [75] | S Sweden [98,99] | Norway [96] | England [91] | Swiss Jura [100] | C Bohemia (Czech Rep.) [92] | Bialowieza Forest (Poland) [97] | Estonia [94] | Denmark [93] | Swiss Plateau [101] | Germany * [68] | Germany ** [95] | France Mediterranean [31] | Romania [87] |
| ERV 3 | ERV 3 | ERV 3 | ERV 1 | ERV 1 | ERV 1 | ERV 1 | ERV 3 | ERV 3 | ERV 1 | ERV 3 | ERV 3 | ERV | ERV | |
| 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) |
| Herb taxa | |||||||||||||||
| 4.28 (0.27) | ||||||||||||||
| 0.26 (0.009) 1,3 | 5.91 (1.23) 1,3 | |||||||||||||
| 2.77 (0.39) 3 | 3.48 (0.20) 1,2 | 5.56 (0.020) 3 | 5.89 (3.16) 1,2,3 | |||||||||||
| 0.30 (0.03) 1,2 | 4.70 (0.69) 1,2,3 | 1.07 (0.03) | 1.10 (0.05) | |||||||||||
| 3.20 (1.14) | 0.0462 (0.0018) 1,2,3 | 1.60 (0.07) | 0.75 (0.04) | 0.00076 (0.0019) 1,2,3 | 9.00 (1.92) 1,2,3 | 0.08 (0.001) 1,2,3 | 0.22 (0.12) 1,2,3 | |||||||
| 0.24 (0.06) 1,3 | 0.06 (0.004) 1,3 | 0.17 (0.03) 1,3 | 1.161 (0.675) 1,3 | 0.16 (0.1) 1,3 | ||||||||||
| 0.10 (0.008) 1,3 | ||||||||||||||
| 0.002 (0.0022) 1,2,3 | 0.89 (0.03) | 1.00 (0.16) | 0.29 (0.01) 1,2,3 | 0.73 (0.08) | 1.23 (0.09) 3 | |||||||||
| 0.07 (0.06) 1,2,3 | 0.11 (0.03) 1,3 | |||||||||||||
| 0.07 (0.04) 1,2 | 4.265 (0.094) 3 | |||||||||||||
| 2.48 (0.82) | 3.39 (missing, 0.00) 3 | 3.13 (0.24) | ||||||||||||
| 12.76 (1.83) 1,2,3 | 1.99 (0.04) | 3.70 (0.77) 3 | 0.90 (0.23) | 0.24 (0.15) 1,2 | 2.73 (0.043) 3 | 0.58 (0.32) 1,2,3 | ||||||||
| 1.27 (0.18) 1,3 | ||||||||||||||
| 0.74 (0.13) 1,3 | ||||||||||||||
| 2.47 (0.38) 1,3 | 0.14 (0.005) 1,3 | 0.96 (0.13) 1,3 | ||||||||||||
| 3.85 (0.72) 1,3 | 0.07 (0.004) 1,3 | 2.037 (0.335) 1,3 | ||||||||||||
| 3.95 (0.59) 1,3 | 0.42 (0.01) 1,3 | 3.47 (0.35) 1,3 | 7.97 (1.08) 1,3 | |||||||||||
| 4.74 (0.83) | 0.13 (0.004) 1,2 | 1.56 (0.09) | 2.76 (0.022) 3 | |||||||||||
| 3.02 (0.05) | 4.08 (0.96) 3 | 4.87 (0.006) 3 | ||||||||||||
| 2.29 (0.36) 1,3 | ||||||||||||||
| 10.52 (0.31) 1,3 | ||||||||||||||
| 0.01 (0.01) 1,2,3 | ||||||||||||||
| Tree taxa | |||||||||||||||
| 3.83 (0.37) | 9.92 (2.86) | |||||||||||||
| 1.27 (0.45) 1,3 | 0.32 (0.10) 1,3 | 0.3 (0.09) 1,3 | ||||||||||||
| 4.20 (0.14) 1,2 | 8.74 (0.35) | 2.56 (0.32) 1,2,3 | 15.95 (0.6622) 3 | 13.93 (0.15) | 15.51 (1.25) 3 | 13.68 (0.049) 3 | ||||||||
| 4.6 (0.70) | 2.24 (0.20) | 8.87 (0.13) 3 | 6.18 (0.35) | 13.94 (0.2293) 1,2,3 | 1.81 (0.02) 3 | 2.42 (0.39) | 9.62 (1.92) 3 | 19.70 (0.117) 1,2,3 | ||||||
| 1.89 (0.068) | ||||||||||||||
| 2.53 (0.07) 1,2 | 4.48 (0.0301) 3 | 4.56 (0.85) | 9.45 (0.51) 1,2,3 | |||||||||||
| 0.24 (0.07) | ||||||||||||||
| 3.258 (0.059) | ||||||||||||||
| 1.40 (0.04) | 1.51 (0.06) | 1.35 (0.0512) 3 | 2.58 (0.39) | 3.44 (0.89) 1,2,3 | ||||||||||
| 1.618 (0.16) 1,2,3 | ||||||||||||||
| 7.53 (0.08) | 5.83 (0.00) | 1.76 (0.20) 3 | 18.47 (0.1032) 1,2,3 | 7.39 (0.20) | 2.56 (0.39) | 2.15 (0.17) 3 | 17.85 (0.049) 1,2,3 | 1.10 (0.35) 1,2,3 | ||||||
| 11.043 (0.261) | ||||||||||||||
| 0.4 (0.07) 1,3 | ||||||||||||||
| 6.67 (0.17) 3 | 1.20 (0.16) 1,2 | 5.09 (0.22) | 0.76 (0.17) 1,2 | 5.83 (0.45) 3 | 9.63 (0.008) 1,2,3 | |||||||||
| 0.67 (0.03) | 0.70 (0.06) 3 | 1.11 (0.09) 3 | 1.39 (0.21) | 6.74 (0.68) 1,2,3 | 1.35 (0.012) 3 | 2.99 (0.88) 1,2,3 | ||||||||
| 0.11 (0.45) 1,2,3 | 2.07 (0.04) | |||||||||||||
| 8.77 (1.81) 1,2,3 | ||||||||||||||
| 0.512 (0.075) | ||||||||||||||
| 2.78 (0.21) | 1.76 (missing, 0.00) 1,2 | 8.43 (0.30) 3 | 4.73 (0.13) | 1.19 (0.42) 1,2 | 0.57 (0.16) 1,2,3 | 1.58 (0.28) 1,2,3 | 5.81 (0.007) 3 | |||||||
| 8.40 (1.34) | 21.58 (2.87) 1,2,3 | 5.66 (missing, 0.00) | 6.17 (0.41) 3 | 23.12 (0.2388) 1,2,3 | 5.07 (0.06) | 1.35 (0.45) 1,2,3 | 5.66 (0.00) 3 | 5.39 (0.222) 3 | ||||||
| 0.755 (0.201) | ||||||||||||||
| 2.66 (1.25) 1,3 | ||||||||||||||
| 0.09 (0.03) | 1.27 (0.31) | 1.05 (0.17) | 1.19 (0.12) 3 | 2.31 (0.08) | ||||||||||
| 1.30 (0.12) 1,3 | ||||||||||||||
| 0.80 (0.03) | 1.36 (0.26) 3 | 0.98 (0.0263) 1,2,3 | 1.47 (0.23) 3 | 12.38 (0.101) 1,2,3 | ||||||||||
| 1.27 (0.05) | 11.51 (0.101) 1,2,3 | |||||||||||||
| 5 | 9 | 25 | 11 | 6 | 10 | 11 | 7 | 10 | 6 | 12 | 13 | 14 | 10 | 11 |
Appendix C
References
- Díaz, S.M.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S.; et al. The Global Assessment Report on Biodiversity and Ecosystem Services: Summary for Policy Makers; IPBES Secretariat: Bonn, Germany, 2019; 1148p. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Scholes, R.J.; Agard, J.; Archer, E.; Arneth, A.; Bai, X.; Barnes, D.; Burrows, M.; Chan, L.; Cheung, W.L.; et al. Scientific Outcome of the IPBES-IPCC Co-Sponsored Workshop on Biodiversity and Climate Change; Secretariat of the Intergovernmental Science-Policy Platform for Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2021. [Google Scholar] [CrossRef]
- Marquer, L.; Gaillard, M.J.; Sugita, S.; Poska, A.; Trondman, A.K.; Mazier, F.; Nielsen, A.B.; Fyfe, R.M.; Jönsson, A.M.; Smith, B.; et al. Quantifying the Effects of Land Use and Climate on Holocene Vegetation in Europe. Quat. Sci. Rev. 2017, 171, 20–37. [Google Scholar] [CrossRef]
- Poska, A.; Väli, V.; Vassiljev, J.; Alliksaar, T.; Saarse, L. Timing and Drivers of Local to Regional Scale Land-Cover Changes in the Hemiboreal Forest Zone during the Holocene: A Pollen-Based Study from South Estonia. Quat. Sci. Rev. 2022, 277, 107351. [Google Scholar] [CrossRef]
- Tinner, W.; Hofstetter, S.; Zeugin, F.; Conedera, M.; Wohlgemuth, T.; Zimmermann, L.; Zweifel, R. Long-Distance Transport of Macroscopic Charcoal by an Intensive Crown Fire in the Swiss Alps—Implications for Fire History Reconstruction. Holocene 2006, 16, 287–292. [Google Scholar] [CrossRef]
- Kalis, A.J.; Merkt, J.; Wunderlich, J. Environmental Changes during the Holocene Climatic Optimum in Central Europe—Human Impact and Natural Causes. Quat. Sci. Rev. 2003, 22, 33–79. [Google Scholar] [CrossRef]
- Roberts, N.; Fyfe, R.M.; Woodbridge, J.; Gaillard, M.-J.; Davis, B.A.S.; Kaplan, J.O.; Marquer, L.; Mazier, F.; Nielsen, A.B.; Sugita, S.; et al. Europe’s Lost Forests: A Pollen-Based Synthesis for the Last 11,000 Years. Sci. Rep. 2018, 8, 716. [Google Scholar] [CrossRef]
- Ellis, E.C.; Gauthier, N.; Goldewijk, K.K.; Bird, R.B.; Boivin, N.; Díaz, S.; Fuller, D.Q.; Gill, J.L.; Kaplan, J.O.; Kingston, N.; et al. People Have Shaped Most of Terrestrial Nature for at Least 12,000 Years. Proc. Natl. Acad. Sci. USA 2021, 118, e2023483118. [Google Scholar] [CrossRef]
- Mottl, O.; Flantua, S.G.A.; Bhatta, K.P.; Felde, V.A.; Giesecke, T.; Goring, S.; Grimm, E.C.; Haberle, S.; Hooghiemstra, H.; Ivory, S.; et al. Erratum: Global Acceleration in Rates of Vegetation Change over the Past 18,000 Years. Science 2021, 373, 860–864. [Google Scholar] [CrossRef]
- Williams, N.S.G.; Hahs, A.K.; Vesk, P.A. Urbanisation, Plant Traits and the Composition of Urban Floras. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 78–86. [Google Scholar] [CrossRef]
- Fyfe, R.M.; Woodbridge, J.; Roberts, C.N. Trajectories of Change in Mediterranean Holocene Vegetation through Classification of Pollen Data. Veg. Hist. Archaeobot. 2018, 27, 351–364. [Google Scholar] [CrossRef]
- Ellis, E.C.; Goldewijk, K.K.; Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic Transformation of the Biomes, 1700 to 2000. Glob. Ecol. Biogeogr. 2010, 19, 589–606. [Google Scholar] [CrossRef]
- Froyd, C.A.; Willis, K.J. Emerging Issues in Biodiversity & Conservation Management: The Need for a Palaeoecological Perspective. Quat. Sci. Rev. 2008, 27, 1723–1732. [Google Scholar] [CrossRef]
- Willis, K.J.; Bailey, R.M.; Bhagwat, S.A.; Birks, H.J.B. Biodiversity Baselines, Thresholds and Resilience: Testing Predictions and Assumptions Using Palaeoecological Data. Trends Ecol. Evol. 2010, 25, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Fredh, D.; Broström, A.; Rundgren, M.; Lagerås, P.; Mazier, F.; Zillén, L. The Impact of Land-Use Change on Floristic Diversity at Regional Scale in Southern Sweden 600 BC-AD 2008. Biogeosciences 2013, 10, 3159–3173. [Google Scholar] [CrossRef]
- Andersen, T.; Carstensen, J.; Hernández-García, E.; Duarte, C.M. Ecological Thresholds and Regime Shifts: Approaches to Identification. Trends Ecol. Evol. 2009, 24, 49–57. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Felde, V.A.; Seddon, A.W.R. Biodiversity Trends within the Holocene. Holocene 2016, 26, 994–1001. [Google Scholar] [CrossRef]
- Woodbridge, J.; Fyfe, R.; Smith, D.; Pelling, R.; de Vareilles, A.; Batchelor, R.; Bevan, A.; Davies, A.L. What Drives Biodiversity Patterns? Using Long-Term Multidisciplinary Data to Discern Centennial-Scale Change. J. Ecol. 2021, 109, 1396–1410. [Google Scholar] [CrossRef]
- Klein Goldewijk, K.; Beusen, A.; Doelman, J.; Stehfest, E. Anthropogenic Land Use Estimates for the Holocene—HYDE 3.2. Earth Syst. Sci. Data 2017, 9, 927–953. [Google Scholar] [CrossRef]
- Kaplan, J.O.; Krumhardt, K.M.; Ellis, E.C.; Ruddiman, W.F.; Lemmen, C.; Goldewijk, K.K. Holocene Carbon Emissions as a Result of Anthropogenic Land Cover Change. Holocene 2010, 21, 775–791. [Google Scholar] [CrossRef]
- Hibbard, K.; Janetos, A.; van Vuuren, D.P.; Pongratz, J.; Rose, S.K.; Betts, R.; Herold, M.; Feddema, J.J. Research Priorities in Land Use and Land-Cover Change for the Earth System and Integrated Assessment Modelling. Int. J. Climatol. 2010, 30, 2118–2128. [Google Scholar] [CrossRef]
- Gaillard, M.-J.; Sugita, S.; Mazier, F.; Trondman, A.-K.; Broström, A.; Hickler, T.; Kaplan, J.O.; Kjellström, E.; Kokfelt, U.; Kuneš, P.; et al. Holocene Land-Cover Reconstructions for Studies on Land Cover-Climate Feedbacks. Clim. Past 2010, 6, 483–499. [Google Scholar] [CrossRef]
- Willis, K.J.; Birks, H.J.B. What Is Natural? The Need for a Long-Term Perspective in Biodiversity Conservation. Science 2006, 314, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Birks, H.J.B.; Willis, K.J. Alpines, Trees, and Refugia in Europe. Plant Ecol. Divers. 2008, 1, 147–160. [Google Scholar] [CrossRef]
- Gaillard, M.-J.; Berglund, B.E.; Birks, H.J.B.; Edwards, K.J.; Bittmann, F. “Think Horizontally, Act Vertically”: The Centenary (1916–2016) of Pollen Analysis and the Legacy of Lennart von Post. Veg. Hist. Archaeobot. 2018, 27, 267–269. [Google Scholar] [CrossRef]
- Prentice, I.C.; Prentice, I.; Webb, T. Pollen Percentages, Tree Abundances and the Fagerlind Effect. J. Quat. Sci. 1986, 1, 35–43. [Google Scholar] [CrossRef]
- Sugita, S. Theory of Quantitative Reconstruction of Vegetation I: Pollen from Large Sites REVEALS Regional Vegetation Composition. Holocene 2007, 17, 229–241. [Google Scholar] [CrossRef]
- Mazier, F.; Gaillard, M.J.; Kuneš, P.; Sugita, S.; Trondman, A.K.; Broström, A. Testing the Effect of Site Selection and Parameter Setting on REVEALS-Model Estimates of Plant Abundance Using the Czech Quaternary Palynological Database. Rev. Palaeobot. Palynol. 2012, 187, 38–49. [Google Scholar] [CrossRef]
- Fyfe, R.M.; Twiddle, C.; Sugita, S.; Gaillard, M.J.; Barratt, P.; Caseldine, C.J.; Dodson, J.; Edwards, K.J.; Farrell, M.; Froyd, C.; et al. The Holocene Vegetation Cover of Britain and Ireland: Overcoming Problems of Scale and Discerning Patterns of Openness. Quat. Sci. Rev. 2013, 73, 132–148. [Google Scholar] [CrossRef]
- Trondman, A.K.; Gaillard, M.J.; Sugita, S.; Björkman, L.; Greisman, A.; Hultberg, T.; Lagerås, P.; Lindbladh, M.; Mazier, F. Are pollen records from small sites appropriate for REVEALS model-based quantitative reconstructions of past regional vegetation? An empirical test in southern Sweden. Vegetation history and archaeobotany. Veg. Hist. Archaeobot. 2016, 25, 131–151. [Google Scholar] [CrossRef]
- Githumbi, E.; Fyfe, R.; Gaillard, M.-J.; Trondman, A.-K.; Mazier, F.; Nielsen, A.-B.; Poska, A.; Sugita, S.; Woodbridge, J.; Azuara, J.; et al. European Pollen-Based REVEALS Land-Cover Reconstructions for the Holocene: Methodology, Mapping and Potentials. Earth Syst. Sci. Data 2022, 14, 1581–1619. [Google Scholar] [CrossRef]
- Strandberg, G.; Lindström, J.; Poska, A.; Zhang, Q.; Fyfe, R.; Githumbi, E.; Kjellström, E.; Mazier, F.; Nielsen, A.B.; Sugita, S.; et al. Mid-Holocene European Climate Revisited: New High-Resolution Regional Climate Model Simulations Using Pollen-Based Land-Cover. Quat. Sci. Rev. 2022, 281, 107431. [Google Scholar] [CrossRef]
- Trondman, A.K.; Gaillard, M.J.; Mazier, F.; Sugita, S.; Fyfe, R.; Nielsen, A.B.; Twiddle, C.; Barratt, P.; Birks, H.J.B.; Bjune, A.E.; et al. Pollen-Based Quantitative Reconstructions of Holocene Regional Vegetation Cover (Plant-Functional Types and Land-Cover Types) in Europe Suitable for Climate Modelling. Glob. Chang. Biol. 2015, 21, 676–697. [Google Scholar] [CrossRef]
- Li, F.; Gaillard, M.-J.; Cao, X.; Herzschuh, U.; Sugita, S.; Ni, J.; Zhao, Y.; An, C.; Huang, X.; Li, Y.; et al. Gridded Pollen-Based Holocene Regional Plant Cover in Temperate and Northern Subtropical China Suitable for Climate Modeling. Earth Syst. Sci. Data 2023, 15, 95–112. Available online: https://essd.copernicus.org/preprints/essd-2022-148/ (accessed on 1 December 2022). [CrossRef]
- Broström, A.; Nielsen, A.B.; Gaillard, M.-J.; Hjelle, K.; Mazier, F.; Binney, H.; Bunting, J.; Fyfe, R.; Meltsov, V.; Poska, A.; et al. Pollen Productivity Estimates of Key European Plant Taxa for Quantitative Reconstruction of Past Vegetation: A Review. Veg. Hist. Archaeobot. 2008, 17, 461–478. [Google Scholar] [CrossRef]
- Wieczorek, M.; Herzschuh, U. Compilation of Relative Pollen Productivity (RPP) Estimates and Taxonomically Harmonised RPP Datasets for Single Continents and Northern Hemisphere Extratropics. Earth Syst. Sci. Data 2020, 12, 3515–3528. [Google Scholar] [CrossRef]
- Golam, S.A. Pollen Morphology and Its Systematic Significance in the Ericaceae. Ph.D. Thesis, Hokkaido University, Sapporo, Japan, 2007. [Google Scholar] [CrossRef]
- Davis, M.B. On the Theory of Pollen Analysis. Am. J. Sci. 1963, 261, 897–912. [Google Scholar] [CrossRef]
- Sugita, S.; Parshall, T.; Calcote, R.; Walker, K. Testing the Landscape Reconstruction Algorithm for Spatially Explicit Reconstruction of Vegetation in Northern Michigan and Wisconsin. Quat. Res. 2010, 74, 289–300. [Google Scholar] [CrossRef]
- Sugita, S. A Model of Pollen Source Area for an Entire Lake Surface. Quat. Res. 1993, 39, 239–244. [Google Scholar] [CrossRef]
- Sutton, O.G. Micrometeorology; McGraw-Hill: New York, NY, USA, 1953. [Google Scholar]
- Sugita, S. Pollen Representation of Vegetation in Quaternary Sediments: Theory and Method in Patchy Vegetation. J. Ecol. 1994, 82, 881–897. [Google Scholar] [CrossRef]
- Prentice, C. Records of Vegetation in Time and Space: The Principles of Pollen Analysis. In Vegetation History; Springer: Dordrecht, The Netherlands, 1988; pp. 17–42. [Google Scholar] [CrossRef]
- Prentice, I.C. Pollen Representation, Source Area, and Basin Size: Toward a Unified Theory of Pollen Analysis. Quat. Res. 1985, 23, 76–86. [Google Scholar] [CrossRef]
- Parsons, R.W.; Prentice, I.C. Statistical approaches to R-values and the pollen—Vegetation relationship. Rev. Palaeobot. Palynol. 1981, 32, 127–152. [Google Scholar] [CrossRef]
- Kuparinen, A.; Markkanen, T.; Riikonen, H.; Vesala, T. Modeling Air-Mediated Dispersal of Spores, Pollen and Seeds in Forested Areas. Ecol. Modell. 2007, 208, 177–188. [Google Scholar] [CrossRef]
- Theuerkauf, M.; Joosten, H. Younger Dryas Cold Stage Vegetation Patterns of Central Europe-Climate, Soil and Relief Controls. Boreas 2012, 41, 391–407. [Google Scholar] [CrossRef]
- Theuerkauf, M.; Couwenberg, J.; Kuparinen, A.; Liebscher, V. A Matter of Dispersal: REVEALSinR Introduces State-of-the-Art Dispersal Models to Quantitative Vegetation Reconstruction. Veg. Hist. Archaeobot. 2016, 25, 541–553. [Google Scholar] [CrossRef]
- Marquer, L.; Mazier, F.; Sugita, S.; Galop, D.; Houet, T.; Faure, E.; Gaillard, M.J.; Haunold, S.; de Munnik, N.; Simonneau, A.; et al. Pollen-Based Reconstruction of Holocene Land-Cover in Mountain Regions: Evaluation of the Landscape Reconstruction Algorithm in the Vicdessos Valley, Northern Pyrenees, France. Quat. Sci. Rev. 2020, 228, 106049. [Google Scholar] [CrossRef]
- Fyfe, R.M.; de Beaulieu, J.-L.; Binney, H.; Bradshaw, R.H.W.; Brewer, S.; Le Flao, A.; Finsinger, W.; Gaillard, M.-J.; Giesecke, T.; Gil-Romera, G.; et al. The European Pollen Database: Past Efforts and Current Activities. Veg. Hist. Archaeobot. 2009, 18, 417–424. [Google Scholar] [CrossRef]
- Giesecke, T.; Davis, B.; Brewer, S.; Finsinger, W.; Wolters, S.; Blaauw, M.; De Beaulieu, J.-L.; Binney, H.; Fyfe, R.M.; Gaillard, M.-J.; et al. Towards Mapping the Late Quaternary Vegetation Change of Europe. Veg. Hist. Archaeobot. 2014, 23, 75–86. [Google Scholar] [CrossRef]
- Kuneš, P.; Abrahám, V.; Kovářík, O.; Kopecký, M.; Břízová, E.; Dudová, L.; Jankovská, V.; Knipping, M.; Kozáková, R.; Nováková, K.; et al. Czech Quaternary Palynological Database-PALYCZ: Review and Basic Statistics of the Data. Preslia Česká Kvartérní Pylová Databáze-PALYCZ: Přehled a Základní Statistika. Preslia 2009, 81, 209–238. [Google Scholar]
- Lerigoleur, E.; Mazier, F.; Jégou, L.; Perret, M.; Galop, D. PALEOPYR: Un Système d’information Pour Les Données Paléoenvironnementales Nord-Pyrénéennes. Ingéniérie Des Systèmes D’information 2015, 20, 63–87. [Google Scholar] [CrossRef]
- Marinova, E.; Harrison, S.P.; Bragg, F.; Connor, S.; De Laet, V.; Leroy, S.A.; Mudie, P.; Atanassova, J.; Bozilova, E.; Caner, H.; et al. Pollen-derived biomes in the eastern Mediterranean–Black Sea–Caspian-corridor. J. Biogeogr. 2018, 45, 484–499. [Google Scholar] [CrossRef]
- Goring, S.; Dawson, A.; Simpson, G.L.; Ram, K.; Graham, R.W.; Grimm, E.C.; Williams, J.W. Neotoma: A Programmatic Interface to the Neotoma Paleoecological Database. Open Quat. 2015, 1, 2. [Google Scholar] [CrossRef]
- Blaauw, M. Methods and Code for “classical” Age-Modelling of Radiocarbon Sequences. Quat. Geochronol. 2010, 5, 512–518. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Abraham, V.; Oušková, V.; Kuneš, P. Present-Day Vegetation Helps Quantifying Past Land Cover in Selected Regions of the Czech Republic. PLoS ONE 2014, 9, e100117. [Google Scholar] [CrossRef] [PubMed]
- Stuart, A.; Ord, J.K. Kendall’s Advanced Theory of Statistics, Distrib. Theory, 1; Scientific Research Publishing: Irvine, CA, USA, 1994; Available online: https://ci.nii.ac.jp/naid/10004597057 (accessed on 2 July 2021).
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Harper, W.V. Reduced Major Axis Regression. Wiley StatsRef Stat. Ref. Online 2016, 1–6. [Google Scholar] [CrossRef]
- Woodbridge, J.; Fyfe, R.M.; Roberts, N. A Comparison of Remotely Sensed and Pollen-Based Approaches to Mapping Europe’s Land Cover. J. Biogeogr. 2014, 41, 2080–2092. [Google Scholar] [CrossRef]
- Bossard, M. CLC2006 Technical Guidelines; European Environment Agency: Copenhagen, Denmark, 2007. [Google Scholar]
- Kinnebrew, E.; Ochoa-Brito, J.I.; French, M.; Mills-Novoa, M.; Shoffner, E.; Siegel, K. Biases and Limitations of Global Forest Change and Author-Generated Land Cover Maps in Detecting Deforestation in the Amazon. PLoS ONE 2022, 17, e0268970. [Google Scholar] [CrossRef]
- Cunningham, D.; Cunningham, P.; Fagan, M.E. Identifying Biases in Global Tree Cover Products: A Case Study in Costa Rica. Forests 2019, 10, 853. [Google Scholar] [CrossRef]
- Tropek, R.; Sedláček, O.; Beck, J.; Keil, P.; Musilová, Z.; Šímová, I.; Storch, D. Comment on “High-Resolution Global Maps of 21st-Century Forest Cover Change”. Science 2014, 344, 850–853. [Google Scholar] [CrossRef]
- Banskota, A.; Kayastha, N.; Falkowski, M.J.; Wulder, M.A.; Froese, R.E.; White, J.C. Forest Monitoring Using Landsat Time Series Data: A Review. Can. J. Remote Sens. 2014, 40, 362–384. [Google Scholar] [CrossRef]
- Matthias, I.; Nielsen, A.B.; Giesecke, T. Evaluating the Effect of Flowering Age and Forest Structure on Pollen Productivity Estimates. Veg. Hist. Archaeobot. 2012, 21, 471–484. [Google Scholar] [CrossRef]
- Coomes, D.A.; Allen, R.B. Effects of Size, Competition and Altitude on Tree Growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- Katz, D.S.W.; Morris, J.R.; Batterman, S.A. Pollen Production for 13 Urban North American Tree Species: Allometric Equations for Tree Trunk Diameter and Crown Area. Aerobiologia 2020, 36, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Hellman, S.E.V.; Gaillard, M.; Broström, A.; Sugita, S. Effects of the Sampling Design and Selection of Parameter Values on Pollen-Based Quantitative Reconstructions of Regional Vegetation: A Case Study in Southern Sweden Using the REVEALS Model. Veg. Hist. Archaeobot. 2008, 17, 445–459. [Google Scholar] [CrossRef]
- Hellman, S.; Gaillard, M.-J.; Broström, A.; Sugita, S. The REVEALS Model, a New Tool to Estimate Past Regional Plant Abundance from Pollen Data in Large Lakes: Validation in Southern Sweden. J. Quat. Sci. 2008, 23, 21–42. [Google Scholar] [CrossRef]
- Bunting, M.J.; Farrell, M.; Broström, A.; Hjelle, K.L.; Mazier, F.; Middleton, R.; Nielsen, A.B.; Rushton, E.; Shaw, H.; Twiddle, C.L. Palynological Perspectives on Vegetation Survey: A Critical Step for Model-Based Reconstruction of Quaternary Land Cover. Quat. Sci. Rev. 2013, 82, 41–55. [Google Scholar] [CrossRef]
- Bunting, M.J.; Hjelle, K.L. Effect of Vegetation Data Collection Strategies on Estimates of Relevant Source Area of Pollen (RSAP) and Relative Pollen Productivity Estimates (Relative PPE) for Non-Arboreal Taxa. Veg. Hist. Archaeobot. 2010, 19, 365–374. [Google Scholar] [CrossRef]
- Von Stedingk, H.; Fyfe, R.M.; Allard, A. Pollen Productivity Estimates from the Forest-Tundra Ecotone in West-Central Sweden: Implications for Vegetation Reconstruction at the Limits of the Boreal Forest. Holocene 2008, 18, 323–332. [Google Scholar] [CrossRef]
- Celikel, G.; Demirsoy, L.; Demirsoy, H. The Strawberry Tree (Arbutus unedo L.) Selection in Turkey. Sci. Hortic. 2008, 118, 115–119. [Google Scholar] [CrossRef]
- Mesléard, F.; Lepart, J. Germination and Seedling Dynamics of Arbutus Unedo and Erica Arbórea on Corsica. J. Veg. Sci. 1991, 2, 155–164. [Google Scholar] [CrossRef]
- Sealy, J.R. Arbutus Unedo. J. Ecol. 1949, 37, 365. [Google Scholar] [CrossRef]
- Fagúndez, J. Two Wild Hybrids of Erica L. (Ericaceae) from Northwest Spain. Bot. Complut. 2006, 30, 131–135. [Google Scholar]
- Bannister, P. Erica cinerea L. J. Ecol. 1965, 53, 527–542. [Google Scholar] [CrossRef]
- Wood, C.E. The Genera of Guttiferae (Clusiaceae) in the Southeastern United States. J. Arnlod Arbor. 1976, 57, 74–90. [Google Scholar]
- Froborg, H. Pollination and Seed Production in Five Boreal Species of Vaccinium and Andromeda (Ericaceae). Can. J. Bot. 1996, 74, 1363–1368. [Google Scholar] [CrossRef]
- Bell, J.N.B.; Tallis, J.H. Empetrum nigrum L. Source J. Ecol. 1973, 61, 289–305. [Google Scholar] [CrossRef]
- Castro-Parada, A.; Muñoz Sobrino, C. Variations in Modern Pollen Distribution in Sediments from Nearby Upland Lakes: Implications for the Interpretation of Paleoecological Data. Rev. Palaeobot. Palynol. 2022, 306, 104765. [Google Scholar] [CrossRef]
- François, L.; Ghislain, M.; Otto, D.; Micheels, A. Late Miocene Vegetation Reconstruction with the CARAIB Model. Palaeogeogr. Palaeoclim. Palaeoecol. 2006, 238, 302–320. [Google Scholar] [CrossRef]
- Romanowska, I.; Crabtree, S.A.; Harris, K.; Davies, B. Agent-Based Modeling for Archaeologists: Part 1 of 3. Adv. Archaeol. Pr. 2019, 7, 178–184. [Google Scholar] [CrossRef]
- Grindean, R.; Nielsen, A.B.; Tanţău, I.; Feurdean, A. Relative Pollen Productivity Estimates in the Forest Steppe Landscape of Southeastern Romania. Rev. Palaeobot. Palynol. 2019, 264, 54–63. [Google Scholar] [CrossRef]
- Li, F.; Gaillard, M.-J.; Xu, Q.; Bunting, M.J.; Li, Y.; Li, J.; Mu, H.; Lu, J.; Zhang, P.; Zhang, S.; et al. A Review of Relative Pollen Productivity Estimates from Temperate China for Pollen-Based Quantitative Reconstruction of Past Plant Cover. Front. Plant Sci. 2018, 9, 1214. [Google Scholar] [CrossRef]
- Räsänen, S.; Suutari, H.; Nielsen, A.B. A Step Further towards Quantitative Reconstruction of Past Vegetation in Fennoscandian Boreal Forests: Pollen Productivity Estimates for Six Dominant Taxa. Rev. Palaeobot. Palynol. 2007, 146, 208–220. [Google Scholar] [CrossRef]
- Prentice, I.C.; Parsons, R.W. Maximum Likelihood Linear Calibration of Pollen Spectra in Terms of Forest Composition. Biometrics 1983, 39, 1051. [Google Scholar] [CrossRef]
- Bunting, M.J.; Armitage, R.; Binney, H.A.; Waller, M. Estimates of ‘relative pollen productivity’ and ‘relevant source area of pollen’ for major tree taxa in two Norfolk (UK) woodlands. Holocene 2005, 15, 459–465. [Google Scholar] [CrossRef]
- Abraham, V.; Kozáková, R. Relative Pollen Productivity Estimates in the Modern Agricultural Landscape of Central Bohemia (Czech Republic). Rev. Palaeobot. Palynol. 2012, 179, 1–12. [Google Scholar] [CrossRef]
- Nielsen, A.B. Modelling Pollen Sedimentation in Danish Lakes at c. AD 1800: An Attempt to Validate the POLLSCAPE Model. J. Biogeogr. 2004, 31, 1693–1709. [Google Scholar] [CrossRef]
- Poska, A.; Meltsov, V.; Sugita, S.; Vassiljev, J. Relative Pollen Productivity Estimates of Major Anemophilous Taxa and Relevant Source Area of Pollen in a Cultural Landscape of the Hemi-Boreal Forest Zone (Estonia). Rev. Palaeobot. Palynol. 2011, 167, 30–39. [Google Scholar] [CrossRef]
- Theuerkauf, M.; Kuparinen, A.; Joosten, H. Pollen Productivity Estimates Strongly Depend on Assumed Pollen Dispersal. Holocene 2013, 23, 14–24. [Google Scholar] [CrossRef]
- Hjelle, K.L. Herb Pollen Representation in Surface Moss Samples from Mown Meadows and Pastures in Western Norway. Veg. Hist. Archaeobot. 1998, 7, 79–96. [Google Scholar] [CrossRef]
- Baker, A.G.; Zimny, M.; Keczyński, A.; Bhagwat, S.A.; Willis, K.J.; Latałowa, M. Pollen Productivity Estimates from Old-Growth Forest Strongly Differ from Those Obtained in Cultural Landscapes: Evidence from the Białowieża National Park, Poland. Holocene 2016, 26, 80–92. [Google Scholar] [CrossRef]
- Broström, A.; Sugita, S.; Gaillard, M.J. Pollen Productivity Estimates for the Reconstruction of Past Vegetation Cover in the Cultural Landscape of Southern Sweden. Holocene 2004, 14, 368–381. [Google Scholar] [CrossRef]
- Sugita, S.; Gaillard, M.J.; Broström, A. Landscape Openness and Pollen Records: A Simulation Approach. Holocene 1999, 9, 409–421. [Google Scholar] [CrossRef]
- Soepboer, W.; Sugita, S.; Lotter, A.F.; van Leeuwen, J.F.N.; van der Knaap, W.O. Pollen Productivity Estimates for Quantitative Reconstruction of Vegetation Cover on the Swiss Plateau. Holocene 2007, 17, 65–77. [Google Scholar] [CrossRef]
- Mazier, F.; Broström, A.; Gaillard, M.-J.; Sugita, S.; Vittoz, P.; Buttler, A. Pollen Productivity Estimates and Relevant Source Area of Pollen for Selected Plant Taxa in a Pasture Woodland Landscape of the Jura Mountains (Switzerland). Veg. Hist. Archaeobot. 2008, 17, 479–495. [Google Scholar] [CrossRef]
- Li, F.; Gaillard, M.-J.; Cao, X.; Herzschuh, U.; Sugita, S.; Tarasov, P.E.; Wagner, M.; Xu, Q.; Ni, J.; Wang, W.; et al. Towards Quantification of Holocene Anthropogenic Land-Cover Change in Temperate China: A Review in the Light of Pollen-Based REVEALS Reconstructions of Regional Plant Cover. Earth-Sci. Rev. 2020, 203, 103119. [Google Scholar] [CrossRef]
- Durbin, J.; Watson, G.S. Testing for Serial Correlation in Least Squares Regression. I. Volume II; Springer: New York, NY, USA, 1992; pp. 237–259. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology in R; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Warton, D.I.; Wright, I.J.; Falster, D.S.; Westoby, M. Bivariate line-fitting methods for allometry. Biol. Rev. Camb. Philos. Soc. 2006, 81, 259–291. [Google Scholar] [CrossRef] [PubMed]
- Warton, D.I.; Duursma, R.A.; Falster, D.S.; Taskinen, S. Smatr 3- an R Package for Estimation and Inference about Allometric Lines. Methods Ecol. Evol. 2012, 3, 257–259. [Google Scholar] [CrossRef]







| Land Cover Types (LCTs) | Plant Taxa/Pollen-Morphological Types | FSP (m/s) | RPPs.st1 (SD) | RPPs.st2 (SD) | RPPs.st3 (SD) |
|---|---|---|---|---|---|
| Evergreen Trees (ET) | Abies | 0.12 | 6.88(1.44) | 6.88(1.44) | 6.88(1.44) |
| Buxus sempervirens | 0.032 | 1.89(0.068) | 1.89(0.068) | 1.89(0.068) | |
| Empetrum | 0.038 | 0.11(0.03) | |||
| Ericaceae | 0.038 | 4.27(0.094) | 4.27(0.094) | 0.07 (0.04) | |
| Juniperus | 0.016 | 2.07(0.04) | 2.07(0.04) | 2.07(0.04) | |
| Phillyrea | 0.015 | 0.51(0.075) | 0.51(0.075) | 0.51(0.075) | |
| Picea | 0.056 | 5.44(0.10) | 5.44(0.10) | 2.62 (0.12) | |
| Pinus | 0.031 | 6.06(0.24) | 6.06(0.24) | 6.38 (0.45) | |
| Pistacia | 0.03 | 0.76(0.201) | 0.76(0.201) | 0.76(0.201) | |
| evergreen Quercus t. | 0.035 | 11.04(0.261) | 11.04(0.261) | 11.04(0.261) | |
| Summergreen Trees (ST) | Acer | 0.056 | 0.63(0.156) | ||
| Alnus | 0.021 | 13.56(0.29) | 13.56(0.29) | 9.07 (0.10) | |
| Betula | 0.024 | 5.11(0.30) | 5.11(0.30) | 3.09 (0.27) | |
| Carpinus betulus | 0.042 | 4.52(0.43) | 4.52(0.43) | 3.55 (0.43) | |
| Carpinus orientalis | 0.042 | 0.24(0.07) | 0.24(0.07) | 0.24(0.07) | |
| Castanea sativa | 0.01 | 3.26(0.059) | 3.26(0.059) | 3.26(0.059) | |
| Corylus avellana | 0.025 | 1.71(0.10) | 1.71(0.10) | 1.99 (0.20) | |
| Fagus | 0.057 | 5.86(0.18) | 5.86(0.18) | 2.35 (0.11) | |
| Fraxinus | 0.022 | 1.04(0.05) | 1.04(0.05) | 1.03 (0.11) | |
| Populus | 0.025 | 2.66(1.25) | |||
| deciduous Quercus t. | 0.035 | 4.54(0.09) | 4.54(0.09) | 5.83 (0.15) | |
| Salix | 0.022 | 1.18(0.08) | 1.18(0.08) | 1.22 (0.11) | |
| Sambucus nigra t. | 0.013 | 1.30(0.12) | |||
| Tilia | 0.032 | 1.21(0.12) | 1.21(0.12) | 0.80 (0.03) | |
| Ulmus | 0.032 | 1.27(0.05) | 1.27(0.05) | 1.27(0.05) | |
| Open Land (OL) | Amaranthaceae/Chenopodiaceae | 0.019 | 4.28(0.270) | 4.28(0.270) | 4.28(0.270) |
| Apiaceae | 0.042 | 3.09(0.615) | |||
| Artemisia | 0.025 | 3.94(0.15) | 3.94(0.15) | 3.48 (0.20) | |
| Calluna vulgaris | 0.038 | 1.09(0.03) | 1.09(0.03) | 0.82 (0.02) | |
| Cerealia t. | 0.06 | 1.85(0.38) | 1.85(0.38) | 1.85 (0.38) | |
| Comp. SF. Cichorioideae | 0.051 | 0.36(0.137) | |||
| Cyperaceae | 0.035 | 0.96(0.05) | 0.96(0.05) | 0.87 (0.06) | |
| Fabaceae | 0.021 | 0.4(0.07) | |||
| Filipendula | 0.006 | 3.00(0.28) | 3.00(0.28) | 2.81 (0.43) | |
| Comp. Leucanthemum (Anthemis) t. | 0.029 | 0.10(0.008) | |||
| Plantago lanceolata | 0.029 | 2.33(0.20) | 2.33(0.20) | 1.04 (0.09) | |
| Plantago media | 0.024 | 1.27(0.18) | |||
| Plantago montana | 0.03 | 0.74(0.13) | |||
| Poaceae | 0.035 | 1.00 (0.00) | 1.00 (0.00) | 1.00 (0.00) | |
| Potentilla t. | 0.018 | 1.19(0.133) | |||
| Ranunculus acris t. | 0.014 | 1.99(0.265) | |||
| Rubiaceae | 0.019 | 3.95(0.314) | |||
| Rumex acetosa t. | 0.018 | 3.02(0.28) | 3.02(0.28) | 2.14 (0.28) | |
| Secale cereale | 0.06 | 3.99(0.32) | 3.99(0.32) | 3.02 (0.05) | |
| Trollius | 0.013 | 2.29(0.36) | |||
| Urtica | 0.007 | 10.52(0.31) | |||
| Number of taxa | 31 | 46 | 31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serge, M.A.; Mazier, F.; Fyfe, R.; Gaillard, M.-J.; Klein, T.; Lagnoux, A.; Galop, D.; Githumbi, E.; Mindrescu, M.; Nielsen, A.B.; et al. Testing the Effect of Relative Pollen Productivity on the REVEALS Model: A Validated Reconstruction of Europe-Wide Holocene Vegetation. Land 2023, 12, 986. https://doi.org/10.3390/land12050986
Serge MA, Mazier F, Fyfe R, Gaillard M-J, Klein T, Lagnoux A, Galop D, Githumbi E, Mindrescu M, Nielsen AB, et al. Testing the Effect of Relative Pollen Productivity on the REVEALS Model: A Validated Reconstruction of Europe-Wide Holocene Vegetation. Land. 2023; 12(5):986. https://doi.org/10.3390/land12050986
Chicago/Turabian StyleSerge, M. A., F. Mazier, R. Fyfe, M.-J. Gaillard, T. Klein, A. Lagnoux, D. Galop, E. Githumbi, M. Mindrescu, A. B. Nielsen, and et al. 2023. "Testing the Effect of Relative Pollen Productivity on the REVEALS Model: A Validated Reconstruction of Europe-Wide Holocene Vegetation" Land 12, no. 5: 986. https://doi.org/10.3390/land12050986
APA StyleSerge, M. A., Mazier, F., Fyfe, R., Gaillard, M.-J., Klein, T., Lagnoux, A., Galop, D., Githumbi, E., Mindrescu, M., Nielsen, A. B., Trondman, A.-K., Poska, A., Sugita, S., Woodbridge, J., Abel-Schaad, D., Åkesson, C., Alenius, T., Ammann, B., Andersen, S. T., ... Zernitskaya, V. P. (2023). Testing the Effect of Relative Pollen Productivity on the REVEALS Model: A Validated Reconstruction of Europe-Wide Holocene Vegetation. Land, 12(5), 986. https://doi.org/10.3390/land12050986