
Effects of Population Declines on Habitat Segregation and Activity Patterns of Rabbits and Hares in Doñana National Park, Spain

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
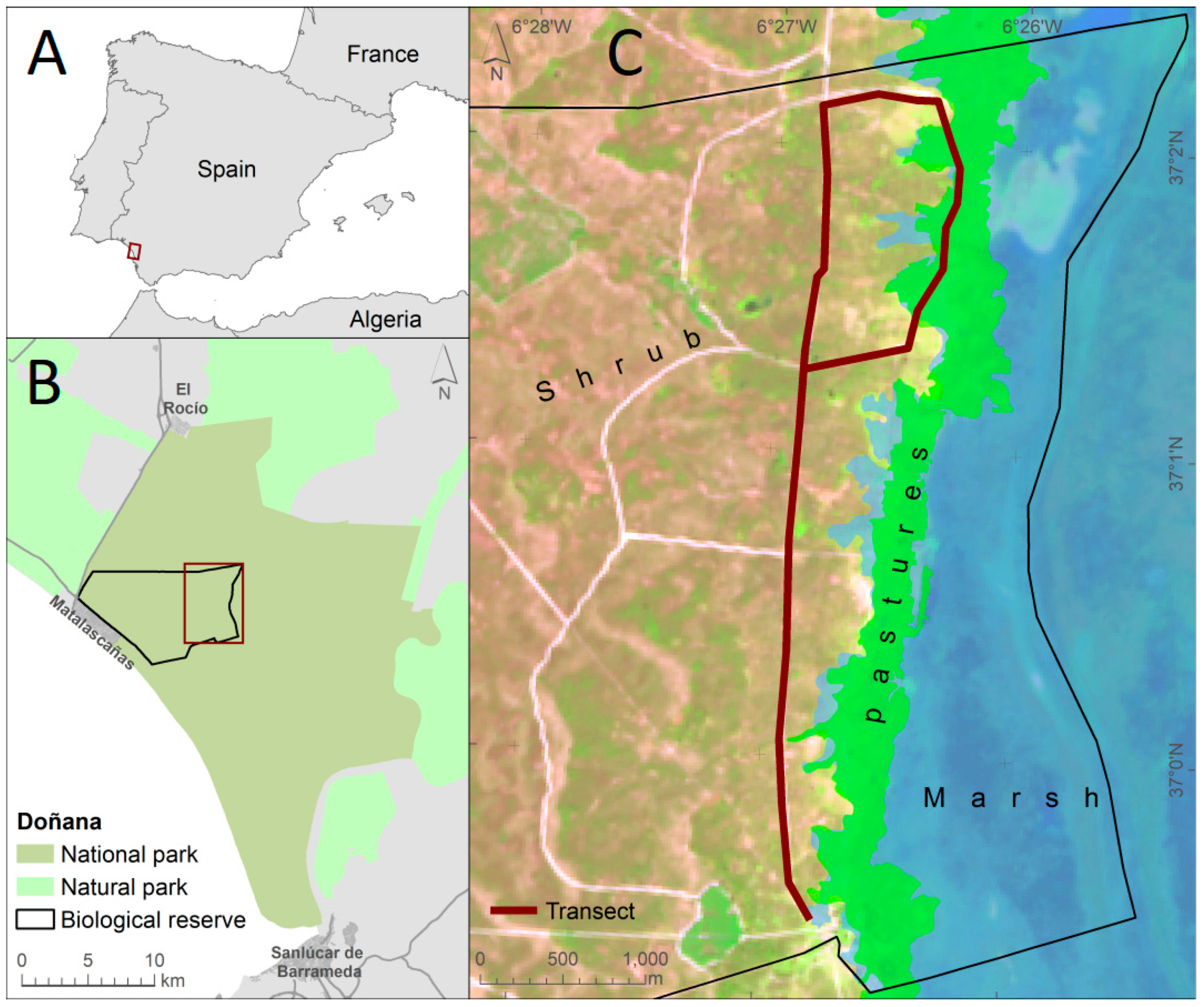
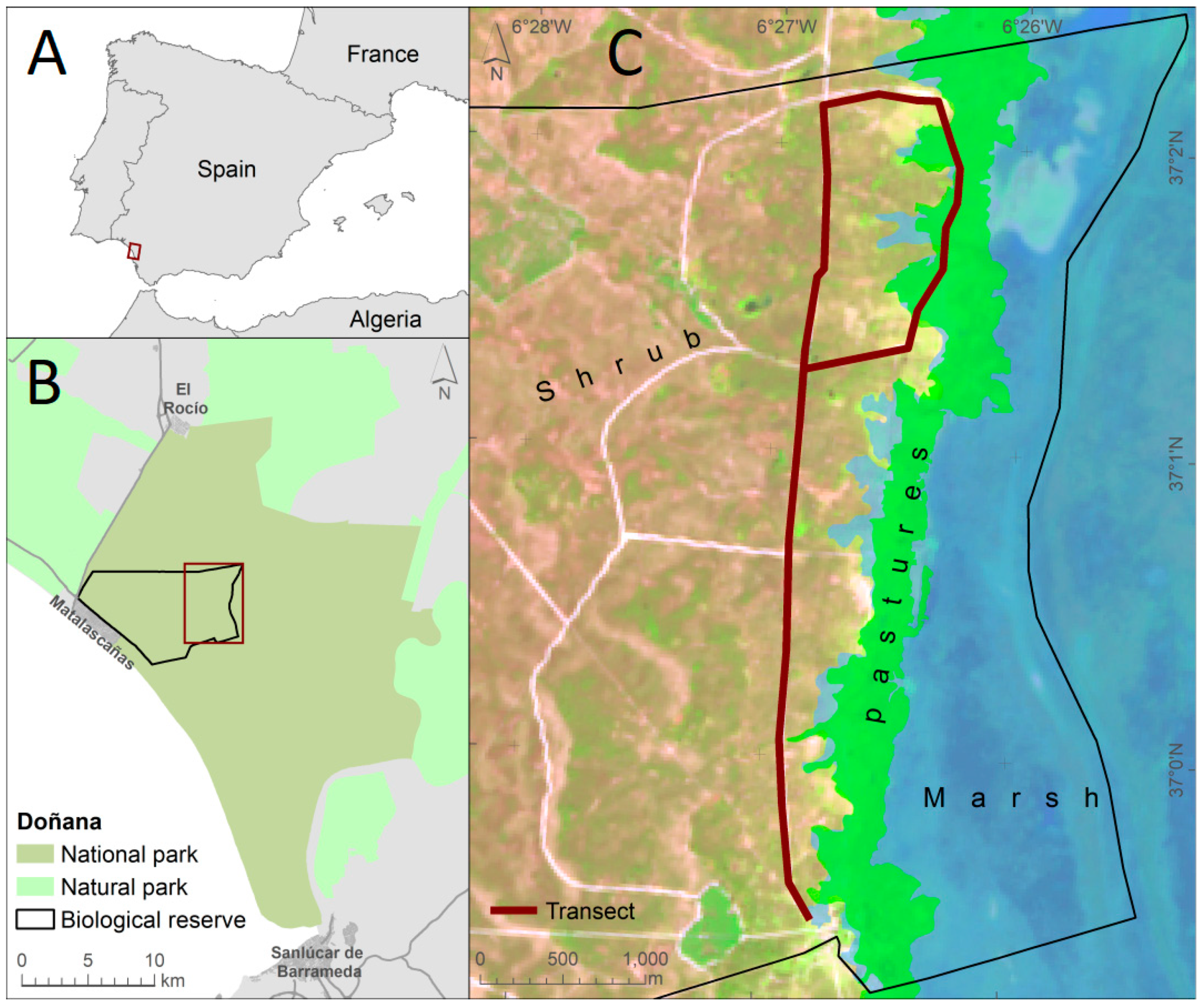
2.1. Study Area
2.2. First Period of the Study: 1983–1985
2.3. Second Period of Study: 2005–2016
3. Results
3.1. First Period: 1983–1985
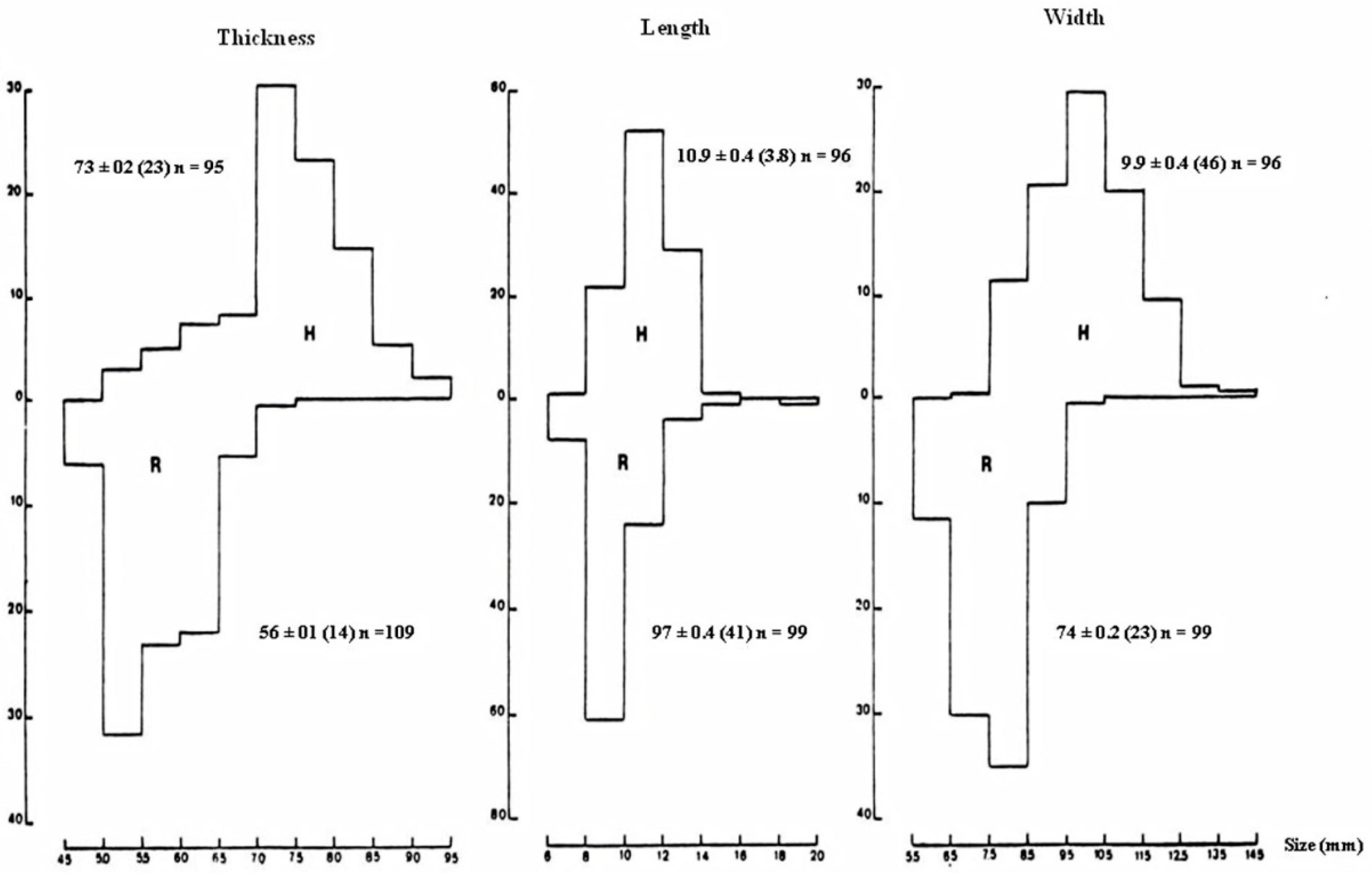
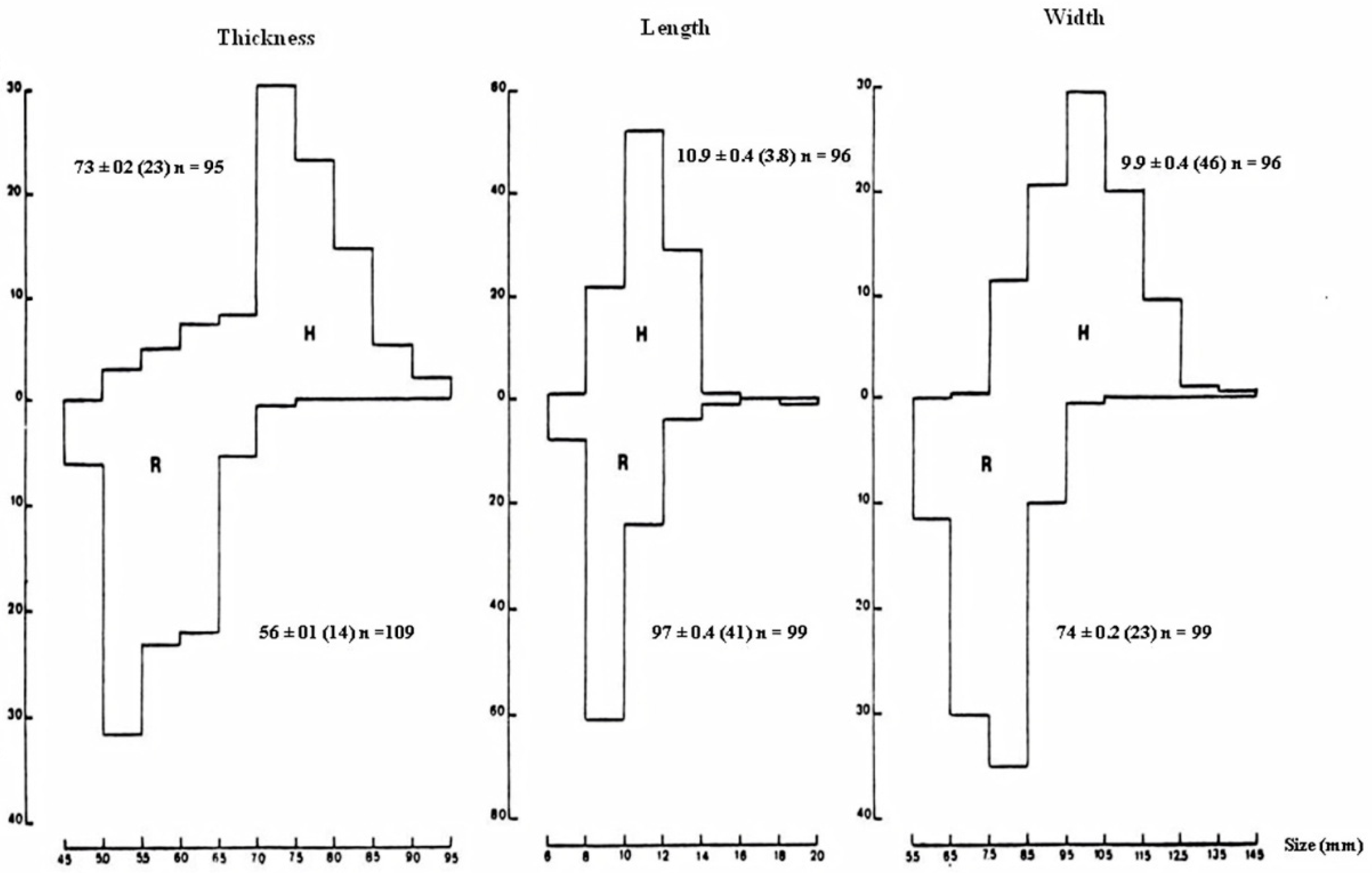
3.1.1. Pellet Characteristics
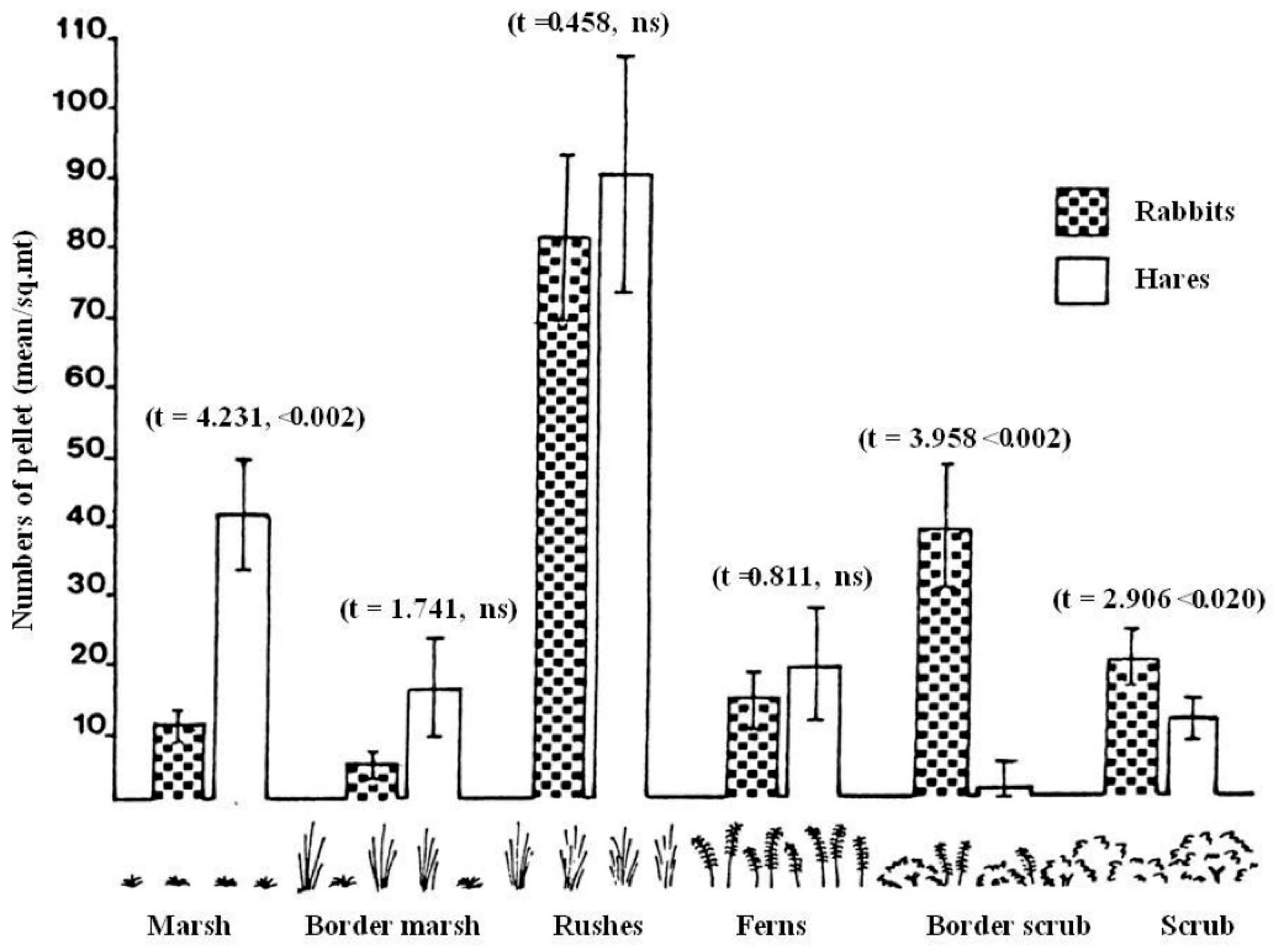
3.1.2. Habitat Use
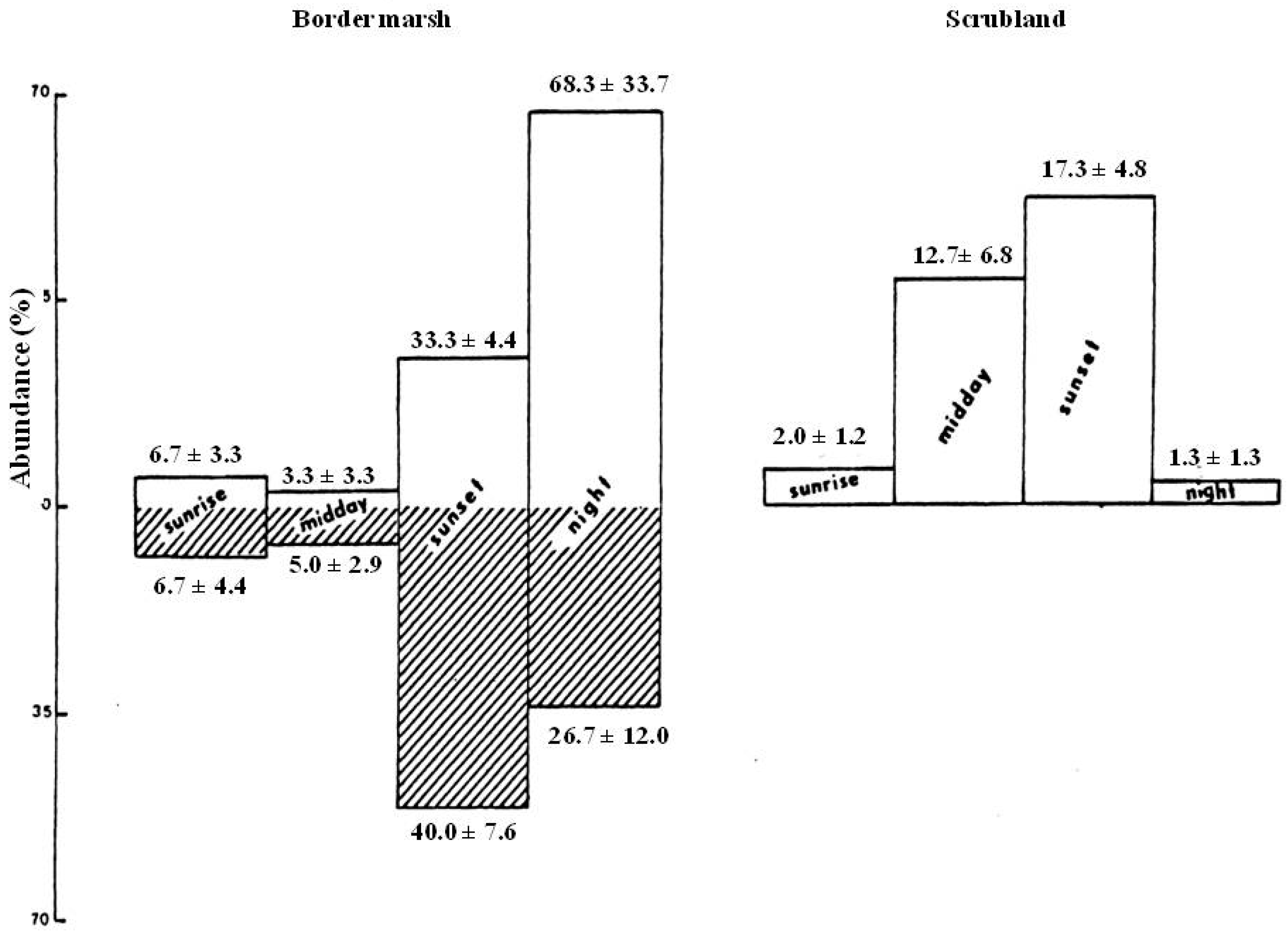
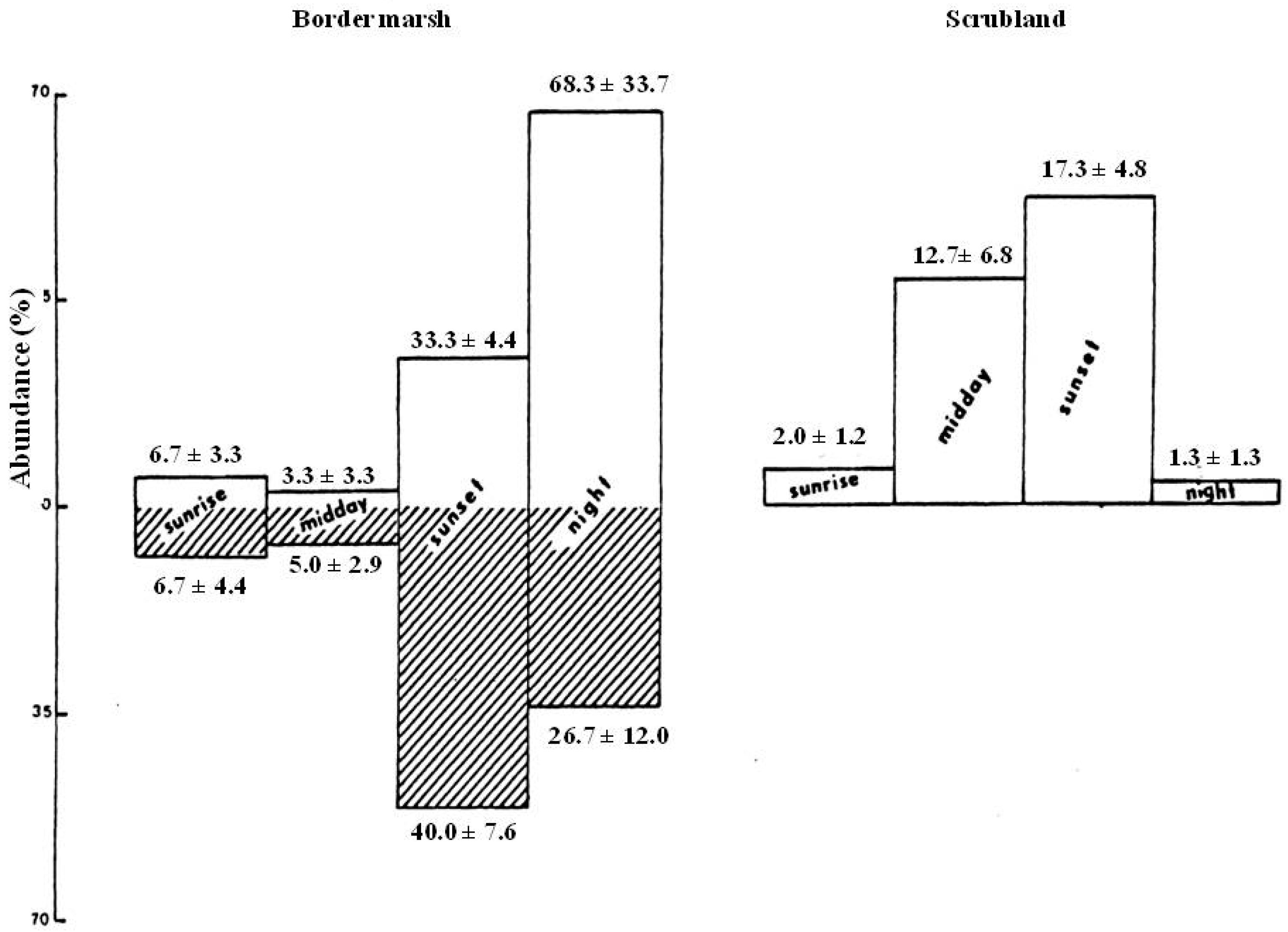
3.1.3. Circadian Activity
3.2. Second Period: 2005–2016
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
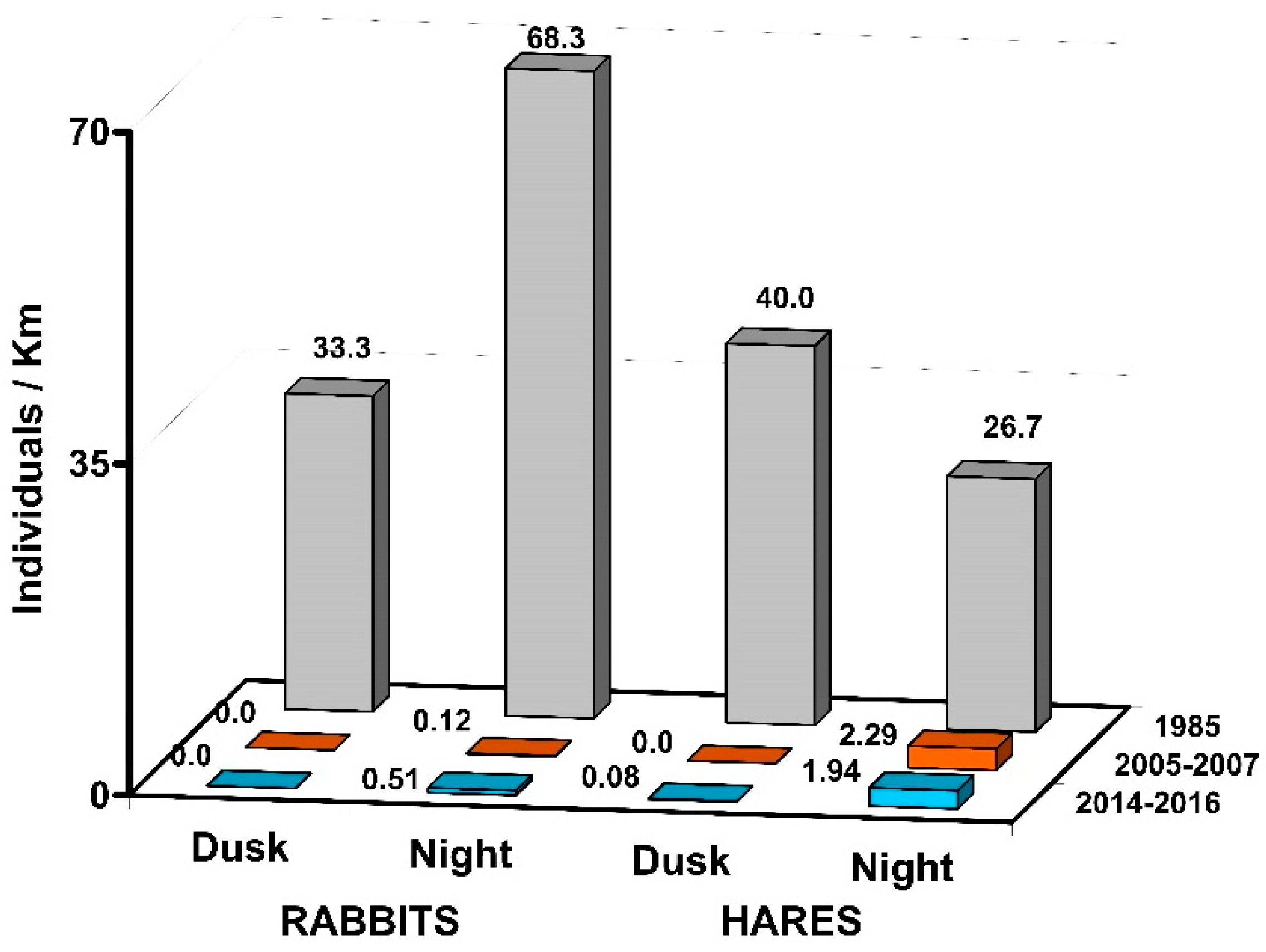
| Year | Month | Rabbits | Hares | ||
|---|---|---|---|---|---|
| Sunset | Night | Sunset | Night | ||
| 2005 | April | 0.00 | 0.70 | 0.35 | 0.35 |
| June | 0.70 | 0.70 | 0.00 | 1.76 | |
| September | 0.00 | 0.00 | 0.00 | 0.00 | |
| Mean ± SE | 0.23 | 0.47 ± 0.02 | 0.12 | 0.70 ± 0.54 | |
| 2006 | March | 0.35 | 0.00 | 0.00 | 0.35 |
| June | 0.00 | 1.60 | 0.00 | 1.40 | |
| September | 0.35 | 0.00 | 0.00 | 0.00 | |
| Mean ± SE | 0.23 ± 0.12 | 0.35 | 0.00 | 0.58 ± 0.42 | |
| 2007 | March | 0.35 | 1.06 | 0.00 | 6.33 |
| June | 0.35 | 0.00 | 0.32 | 3.52 | |
| September | 0.00 | 1.06 | 0.00 | 2.81 | |
| Mean ± SE | 0.23 ± 0.12 | 0.71 ± 0.35 | 0.12 | 4.22 ± 1.10 | |
| 3-year mean ± SE | 0.23 | 0.51 ± 0.11 | 0.08 ± 0.04 | 1.84 ± 1.19 | |
| 2014 | March | 0.00 | 0.00 | 0.00 | 2.11 |
| September | 0.00 | 0.35 | 0.00 | 0.35 | |
| Mean ± SE | 0.00 | 0.18 | 0.00 | 1.23 ± 0.88 | |
| 2015 | March | 0.00 | 0.00 | 0.00 | 5.27 |
| September | 0.00 | 0.35 | 0.00 | 0.00 | |
| Mean ± SE | 0.00 | 0.18 | 0.00 | 2.64 | |
| 2016 | March | 0.00 | 0.00 | 0.00 | 3.87 |
| September | 0.00 | 0.00 | 0.00 | 2.11 | |
| Mean ± SE | 0.00 | 0.00 | 0.00 | 2.99 ± 0.88 | |
| 3-year mean ± SE | 0.00 | 0.00 | 0.00 | 2.29 ± 0.54 | |
| Year | Month | Rabbits | Hares | ||
|---|---|---|---|---|---|
| Sunset | Night | Sunset | Night | ||
| 2005 | April | 0.00 | 0.57 | 0.00 | 0.00 |
| June | 0.19 | 0.19 | 0.00 | 0.00 | |
| September | 0.00 | 0.00 | 0.00 | 0.00 | |
| Mean ± SE | 0.06 | 0.25 ± 0.17 | 0.00 | 0.00 | |
| 2006 | March | 0.00 | 0.00 | 0.00 | 0.00 |
| June | 0.38 | 0.00 | 0.00 | 0.38 | |
| September | 0.38 | 0.00 | 0.00 | 0.00 | |
| Mean ± SE | 0.25 ± 0.13 | 0.00 | 0.00 | 0.13 | |
| 2007 | March | 0.00 | 0.19 | 0.00 | 0.19 |
| June | 0.19 | 0.00 | 0.00 | 0.57 | |
| September | 0.00 | 0.38 | 0.00 | 0.76 | |
| Mean ± SE | 0.06 | 0.19 ± 0.11 | 0.00 | 0.51 ± 0.17 | |
| 3-year mean ± SE | 0.13 ± 0.06 | 0.15 ± 0.08 | 0.00 | 0.21 ± 0.15 | |
| 2014 | March | 0.00 | 0.00 | 0.00 | 0.00 |
| September | 0.00 | 0.77 | 0.00 | 0.77 | |
| Mean ± SE | 0.00 | 0.39 | 0.00 | 0.39 | |
| 2015 | March | 0.00 | 0.19 | 0.00 | 0.19 |
| September | 0.19 | 0.19 | 0.00 | 0.00 | |
| Mean ± SE | 0.10 | 0.19 | 0.00 | 0.10 | |
| 2016 | March | 0.00 | 0.00 | 0.00 | 0.19 |
| September | 0.00 | 0.19 | 0.00 | 0.19 | |
| Mean ± SE | 0.00 | 0.10 | 0.00 | 0.19 | |
| 3-year mean ± SE | 0.03 | 0.22 ± 0.09 | 0.00 | 0.22 ± 0.09 | |
References
- Amarasekare, P. Competitive coexistence in spatially structured environments: A synthesis. Ecol. Lett. 2003, 6, 1109–1122. [Google Scholar] [CrossRef]
- Cheeseman, A.E.; Ryan, S.J.; Whipps, C.M.; Cohen, J.B. Competition alter seasonal resource selection and promotes use of invasive shrubs by an inperiled native cottontail. Ecol. Evol. 2018, 8, 11122–11133. [Google Scholar] [CrossRef] [Green Version]
- Woodgate, Z.; Distiller, G.; O´Riain, M.J. Hare today gone tomorrow: The role of interspecific competition in shaping riverine rabbit occurrence. Endanger. Species Res. 2021, 44, 351–361. [Google Scholar] [CrossRef]
- Lombardi, L.; Fernández, N.; Moreno, S.; Villafuerte, R. Habitat-related differences in rabbit (Oryctolagus cuniculus) abundance, distribution, and activity. J. Mammal. 2003, 84, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Terborg, J.; Estes, J.A. Trophic Cascades: Predators, Prey, and the Changing Dynamics of Nature; Island Press: Washington, DC, USA, 2010. [Google Scholar]
- Viviano, A.; Mori, E.; Fattorini, N.; Mazza, G.; Lazzeri, L.; Panichi, A.; Strianese, L.; Mohamed, W. Spatiotemporal overlap between the European Brown Hare and its potential predators and competitors. Animals 2021, 11, 562. [Google Scholar] [CrossRef]
- Moreno, S.; Beltrán, J.F.; Cotilla, I.; Kuffner, B.; Laffitte, R.; Jordán, G.; Ayala, J.; Quintero, M.C.; Jiménez, A.; Castro, F.; et al. Long-term decline of the European wild rabbit (Oryctolagus cuniculus) in south-western. Wildl. Res. 2007, 34, 652–658. [Google Scholar] [CrossRef] [Green Version]
- Delibes-Mateos, M.; Delibes, M.; Ferreras, P.; Villafuerte, R. Key role of the European rabbit in the conservation of the Western Mediterranean basin hotspot. Conserv. Biol. 2008, 22, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Monterroso, P.; Garrote, G.; Serronha, A.; Santos, E.; Delibes-Mateos, M.; Abrantes, J.; Pérez de Ayala, R.; Silvestre, F.; Carvalho, J.; Vasco, I.; et al. Disease-mediated bottom-up regulation: An emergent virus affects a keystone species, and alters the dynamics of trophic webs. Sci. Rep. 2016, 6, 36072. [Google Scholar] [CrossRef] [Green Version]
- Ferreras, P.; Travaini, A.; Zapata, S.C.; Delibes, M. Short responses of mammalian carnivores to a sudden collapse of rabbits in Mediterranean Spain. Basic Appl. Ecol. 2011, 12, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Calvete, C.; Estrada, R.; Villafuerte, R.; Osácar, J.J.; Lucientes, J. Epidemiology of viral haemorrhagic disease and myxomatosis in a free-living population of wild rabbits. Vet. Rec. 2002, 150, 776–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothschild, M.A.; Marsh, H. Increase of hares (Lepus europaeus Pallas) at Ashton Wold with a note on the reduction in numbers of the brown rat (Rattus norvegicus Berkenhout). Proc. Zool. Soc. Lond. 1956, 127, 441–445. [Google Scholar] [CrossRef]
- Rothschild, M. A further note in the increase of hares (Lepus europaeus) in France. Proc. Zool. Soc. Lond. 1958, 131, 328–329. [Google Scholar]
- Rothschild, M.A. Increase of hares at Ashton Wold. Proc. Zool. Soc. Lond. 1961, 137, 634–635. [Google Scholar]
- Flux, J.E.C. Hare reproduction in New Zealand. N. Z. J. Agric. 1964, 109, 483–486. [Google Scholar]
- Broekhuizen, S. The position of the wild rabbit in the life system of the European hare. In Proceedings of the XII Congress of the International Union of Game Biologists; Direcção-Geral dos Recursos Florestais Serviço de Inspecção da Caça e Pesca: Lisboa, Portugal, 1975; pp. 75–79. [Google Scholar]
- Broekhuizen, S.; Kemmers, R. The stomach worm, Graphidium strigosum (Dujardin) Railliet and Henry in the European hare, Lepus europaeus Pallas. In Ecology and Management of European Hare Populations; Pielowski, Z., Pucek, Z., Eds.; Polish Hunt. Ass.: Warsaw, Poland, 1976; pp. 157–171. [Google Scholar]
- Gibb, J.A.; Flux, J.E.C. Why New Zealand should not use myxomatosis in rabbit control operations. Search 1983, 14, 41–43. [Google Scholar]
- Fraguglione, D. Compétition interspécifique entre le lièvre commun (Lepus europaeus Pallas 1778) et le lapin de garenne (Oryctolagus cuniculus Linné 1758). Diana 1960, 10, 211–212. [Google Scholar]
- Fraguglione, D. Les enemis du lièvre commun (Lepus europaeus P.). Diana 1960, 3, 54–57. [Google Scholar]
- Flux, J.E.C. Field observations of behaviour in the genus Lepus. In Proceedings of the World Lagomorph Conference, Guelph, ON, Canada, 12–16 August 1979; pp. 377–394. [Google Scholar]
- Keith, L.B. Population dynamics of hares. In Proceedings of the World Lagomorph Conference, Guelph, ON, Canada, 12–16 August 1979; pp. 395–440. [Google Scholar]
- Flux, J.E.C. Prospects for hare farming in New Zealand. N. Z. Agric. Sci. 1981, 15, 24–29. [Google Scholar]
- Katona, K.; Bíró, Z.; Hahn, I.; Kertész, M.; Altbäcker, V. Competition between European hare and European rabbit in a lowland area, Hungary: A long-term ecological study in the period of rabbit extinction. Folia Zool. 2004, 53, 255–268. [Google Scholar]
- Le Gall-Reculé, G.; Zwingelstein, F.; Fages, M.P.; Bertagnoli, S.; Gelfi, J.; Aubineau, J.; Roobrouk, A.; Botti, G.; Lavazza, A.; Marchandeu, S. Characterisation of a non-pathogenic and non-protective infectious rabbit lagovirus related to RHDV. Virology 2011, 410, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Le Gall-Reculé, G.; Lavazza, A.; Marchandeu, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guérin, J.L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Reculé, G.; Lopes, A.M.; Marchandeau, S.; Alma, F.; Almeida, A.M.; Alves, P.C.; et al. Proposal for a new unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Delibes-Mateos, M.; Ferreira, C.; Carro, F.; Escudero, M.A.; Gortázar, C. Ecosystem Effects of Variant Rabbit Hemorrhagic Disease Virus, Iberian Peninsula. Emerg. Infect. Dis. 2014, 20, 2166–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carro, F.; Soriguer, R. Long-term patterns in Iberian hare population dynamics in a protected area (Doñana National Park) in southwestern Iberian Peninsula: Effects of weather conditions and plant cover. Integr. Zool. 2016, 12, 49–60. [Google Scholar] [CrossRef] [PubMed]
- García-Bocanegra, I.; Camacho-Sillero, L.; Risalde, M.A.; Dalton, K.P.; Caballero-Gómez, J.; Agüero, M.; Gómez-Guillamón, F. First outbreak of myxomatosis in Iberian hares (Lepus granatensis). Transbound. Emerg. Dis. 2019, 66, 2204–2208. [Google Scholar] [CrossRef]
- Grant, P.R. Experimental studies of competitive interaction in a two-species system. III. Microtus and Peromyscus species in enclosures. J. Anim. Ecol. 1971, 40, 323–350. [Google Scholar] [CrossRef]
- Grant, P.R. Competition between species of small mammals. In Populations of Small Mammals under Natural Conditions; Snyder, D.P., Ed.; Pymatuning Lab. of Ecol., Univ. of Pittsburg: Linesville, PA, USA, 1978; Volume 5, pp. 38–51. [Google Scholar]
- Dickman, C.R.; Woodside, D.P. A test of competition model with reference to three species of small mammals in south-eastern Australia. Oecologia 1983, 60, 127–134. [Google Scholar] [CrossRef]
- Pettorelli, N.; Coulson, T.; Durante, S.M.; Gaillard, J.M. Predation, individual variability and vertebrate population dynmics. Oecologia 2011, 167, 305–314. [Google Scholar] [CrossRef]
- Diamond, J. Overview: Laboratory experiments, field experiments, and natural experiments. In Community Ecology, Diamond J, TJ Case; Harper & Row: New York, NY, USA, 1986; pp. 3–22. [Google Scholar]
- Valverde, J.A. An Ecological Sketch of the Coto Doñana; British Birds: London, UK, 1958; Volume 51, pp. 1–23. [Google Scholar]
- Aguilar-Amat, J.; Montes del Olmo, C.; Ramírez Díaz, L.; Torres Martínez, A. Mapa Ecológico del Parque Nacional de Doñana; ICONA: Sevilla, Spain, 1979. [Google Scholar]
- Green, A.; Bustamante, J.; Janss, G.F.E.; Fernández-Zamudio, R.; Díaz-Paniagua, C. Doñana Wetlands (Spain). In The WetlandBook; Finlayson, C., Milton, G., Prentice, R., Davidson, N., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 1126–1136. [Google Scholar]
- Allier, C.; González, F.; Ramírez, L. Mapa ecológico de la Reserva Biológica de Doñana. In CSIC, División de Ciencias; Estación Biológica de Doñana: Sevilla, Spain, 1974. [Google Scholar]
- Rogers, P.M.; Myers, K. Animal distributions, landscape classification and wildlife management, Coto Doñana. J. Appl. Ecol. 1980, 17, 545–565. [Google Scholar] [CrossRef]
- Neff, J.D. The pellet-group count technique for big game trend, census, and distribution: A review. J. Wildl. Manag. 1968, 32, 597–614. [Google Scholar] [CrossRef]
- Lawrence, M.J.; Brown, R.W. Mammals of Britain, Their Tracks, Trails and Signs; Blandford Press: London, UK, 1974. [Google Scholar]
- Bang, P.; Dahlström, P. Huellas y Señales de los Animales de Europa; Ediciones Omega, S.A.: Barcelona, Spain, 1975. [Google Scholar]
- Overton, W.S. Estimating the numbers of animals in wildlife populations. In Wildlife Management Techniques, 3rd ed.; Giles, R.H., Jr., Ed.; The Wildlife Society: Washington, DC, USA, 1971; pp. 403–456. [Google Scholar]
- Salzmann-Wandeler, I.; Salzmann, H.C. Erste Erfahrungen Bei Feldhaszen Zählungen Mit Scheinwerfern; Naturhistorisches Museum Bern: Bern, Switzerland, 1973; Volume 5, pp. 1–16. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice Hall, Inc.: Englewood Cliffs, NJ, USA, 2010. [Google Scholar]
- Barnes, R.F.W.; Tapper, S.C.; Williams, J. Use of pastures by brown hares. J. Appl. Ecol. 1983, 20, 179–185. [Google Scholar] [CrossRef]
- Homolka, M. Daily activity pattern of the European hare (Lepus europaeus). Folia Zool. 1986, 35, 33–42. [Google Scholar]
- Barnes, R.F.W.; Tapper, S.C. A method for counting hares by spotlight. J. Zool. 1985, 206, 273–276. [Google Scholar] [CrossRef]
- Frylestam, B. Estimating by spotlight the population density of the European hare. Acta Theriol. 1981, 26, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Frylestam, B. European hare. In CRC Handbook of Census Methods for Terrestrial Vertebrates; Davis, E.D., Ed.; CRC Press: Boca Raton, FL, USA, 1982; pp. 142–144. [Google Scholar]
- Wywialowski, A.; Stottart, L.C. Estimation of jack rabbit density methodology makes a difference. J. Wildl. Manag. 1988, 52, 57–59. [Google Scholar] [CrossRef] [Green Version]
- Rogers, P.M.; Myers, K. Ecology of the European wild rabbit, Oryctolagus cuniculus (L.) in Mediterranean habitats. I. Distribution in the landscape of the Coto Doñana, S. Spain. J. Appl. Ecol. 1979, 16, 691–703. [Google Scholar] [CrossRef]
- Sale, P.F. Overlap in resource use, and interspecific competition. Oecologia 1974, 17, 245–256. [Google Scholar] [CrossRef]
- Rosenweig, M.L. A theory of habitat selection. Ecology 1981, 62, 327–335. [Google Scholar] [CrossRef]
- Vidus-Rosin, A.; Lizier, L.; Meriggi, A.; Serrano-Pérez, L. Habitat selection and segregation by two sympatric lagomorphs: The case of the European hare (Lepus europaeus) and Eastern cottontails (Sylvilagus floridanus) in northern Italy. Acta Theriol. 2012, 57, 295–304. [Google Scholar] [CrossRef]
- Rowland, M.M.; White, G.C.; Karlen, E.M. Use of pellet group plots to measure trends in deer and elk populations. Wildl. Soc. Bull. 1984, 12, 147–155. [Google Scholar]
- Alvarez, F.; Braza, F.; de Azcárate, T.; Aguilera, E.; Martín, R. Circadian activity rhythms in a vertebrate community of Doñana. In Actas XV Congr. Nac. de Fauna Cinegética y Silvestre; Trujillo: Cáceres, Spain, 1983; pp. 379–387. [Google Scholar]
- Villafuerte, R.; Kufner, M.B.; Delibes, M.; Moreno, S. Environmental factors influencing the seasonal daily activity of the European rabbit (Oryctolagus cuniculus) in a Mediterranean area. Mammalia 1993, 57, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Carothers, J.H.; Jaksic, F.M. Time as niche difference: The role of interference competition. Oikos 1984, 42, 403–406. [Google Scholar] [CrossRef]
- Soriguer, R.C. El conejo: Papel ecológico y estrategia de vida en los ecosistemas mediterráneos. In XV Congreso Internacional de Fauna Cinegética y Silvestre; Trujillo: Cáceres, Spain, 1981; pp. 517–542. [Google Scholar]
- Soriguer, R.C. Consideraciones sobre el efecto de los conejos y los grandes herbívoros en los pastizales de la Vera de Doñana. Doñana Acta Vert. 1983, 10, 155–168. [Google Scholar]
- Pimm, S.L.; Pimm, J.W. Resource use, competition and resource availability in Hawaian honeycreepers. Ecology 1982, 63, 1468–1480. [Google Scholar] [CrossRef]
- Paine, R.T. Food web complexity and species diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Delibes, M. Ecología alimenticia del águila imperial ibérica (Aquila adalberti) en el Coto Doñana durante la crianza de los pollos. Doñana. Acta Vert. 1978, 5, 35–60. [Google Scholar]
- Delibes, M. Feeding ecology of the Spanish Lynx in the Coto Doñana. Acta Theriol. 1980, 25, 309–394. [Google Scholar] [CrossRef] [Green Version]
- Carro, F.; Soriguer, R.C.; Beltrán, J.F.; Andreu, A.C. Heavy flooding effects on home range and habitat selection of free-ranging Iberian hares (Lepus granatensis) in Doñana National Park (SW Spain). Acta Theriol. 2011, 53, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Villafuerte, R. Riesgo de Predación y Estrategias Defensivas Del Conejo, Oryctolagus cuniculus, en el Parque. Nacional de Doñana. Ph.D. Thesis, Universidad de Córdoba, Córdoba, Spain, 1994. [Google Scholar]
- Moreno, S.; Villafuerte, R.; Delibes, M. Cover is safe during the the day but dangerous at night: The use of vegetation by the European wild rabbit. Can. J. Zool. 1996, 74, 1656–1660. [Google Scholar] [CrossRef]
- Carro, F. Historia natural de la liebre ibérica (Lepus granatensis Rosenhauer 1856) en el Parque Nacional de Doñana. Ph.D. Thesis, University of Santiago de Compostela, Santiago de Compostela, Spain, 2005. [Google Scholar]
- Negro, J.J.; Ferrer, M. The near extinction of two large European predators: Super Specialists pay a price. Conserv. Biol. 2004, 18, 344–349. [Google Scholar]
- Palomares, F.; Delibes, M.; Ferreras, P.; Gaona, P. Mesopredator release and prey abundance: A reply to Litvaitis and Villafuerte. Cons. Biol. 1996, 10, 678–679. [Google Scholar] [CrossRef]
- Tilman, D. Resource competition and community structure. In Monographs Population Biology, Volume 17; Princeton University Press: New York, NY, USA, 1982. [Google Scholar]
- Taylor, R.J. Predation; Chapman and Hall: London, UK, 1984. [Google Scholar]
- Bakker, E.S.; Reiffers, R.C.; Olff, H.; Gleichman, J.M. Experimental Manipulation of Predation Risk and Food Quality: Effect on Grazing Behaviour in a Central-Place Foraging Herbivore. Oecologia 2005, 146, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Catch me if you can: Diel activity patterns of mammalian prey and predators. Ethology 2013, 119, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Beltrán, J.F.; Delibes, M. Environmental determinants of circadian activity of free ranging Iberian lynxes. J. Mammal. 1994, 75, 382–393. [Google Scholar] [CrossRef]


| Scrublands | Border Marshlands | |||
|---|---|---|---|---|
| Month | n | Rabbits | Rabbits | Hares |
| November 1983 | 3 | 10.7 ± 8.7 | 8.3 ± 3.3 | 45.0 ± 35.5 |
| December 1983 | 2 | 33.0 ± 19.1 | 0.0 | 2.5 ± 2.5 |
| April 1984 | 3 | 32.7 ± 10.5 | 11.7 ± 9.3 | 18.3 ± 8.3 |
| May 1984 | 4 | 66.5 ± 18.0 | 2.5 ± 2.5 | 5 ± 3.5 |
| July 1984 | 4 | 67.0 ± 15.7 | 27.5 ± 7.8 | 1.2 ± 1.2 |
| November 1984 | 3 | 18.7 ± 4.4 | 3.3 ± 3.3 | 1.7 ± 1.7 |
| Grand mean | 19 | 41.4 ± 7.4 | 10.0 ± 3.1 | 11.8 ± 6.1 |
| Average biomass(kg/10 km) | 37.7 | 9.1 | 23.4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beltrán, J.F.; Rau, J.R.; Soriguer, R.C.; Kufner, M.B.; Delibes, M.; Carro, F. Effects of Population Declines on Habitat Segregation and Activity Patterns of Rabbits and Hares in Doñana National Park, Spain. Land 2022, 11, 461. https://doi.org/10.3390/land11040461
Beltrán JF, Rau JR, Soriguer RC, Kufner MB, Delibes M, Carro F. Effects of Population Declines on Habitat Segregation and Activity Patterns of Rabbits and Hares in Doñana National Park, Spain. Land. 2022; 11(4):461. https://doi.org/10.3390/land11040461
Chicago/Turabian StyleBeltrán, Juan F., Jaime R. Rau, Ramón C. Soriguer, Maura B. Kufner, Miguel Delibes, and Francisco Carro. 2022. "Effects of Population Declines on Habitat Segregation and Activity Patterns of Rabbits and Hares in Doñana National Park, Spain" Land 11, no. 4: 461. https://doi.org/10.3390/land11040461
APA StyleBeltrán, J. F., Rau, J. R., Soriguer, R. C., Kufner, M. B., Delibes, M., & Carro, F. (2022). Effects of Population Declines on Habitat Segregation and Activity Patterns of Rabbits and Hares in Doñana National Park, Spain. Land, 11(4), 461. https://doi.org/10.3390/land11040461