Abstract
Although renewable energy holds great promise in mitigating climate change, there are socioeconomic and ecological tradeoffs related to each form of renewable energy. Forest-related bioenergy is especially controversial, because tree plantations often replace land that could be used to grow food crops and can have negative impacts on biodiversity. In this study, we examined public perceptions and ecosystem service tradeoffs between the provisioning services associated with cover types associated with bioenergy crop (feedstock) production and forest habitat-related supporting services for birds, which themselves provide cultural and regulating services. We combined a social survey-based assessment of local values and perceptions with measures of bioenergy feedstock production impacts on bird habitat in four countries: Argentina, Brazil, Mexico, and the USA. Respondents in all countries rated birds as important or very important (83–99% of respondents) and showed lower enthusiasm for, but still supported, the expansion of bioenergy feedstocks (48–60% of respondents). Bioenergy feedstock cover types in Brazil and Argentina had the greatest negative impact on birds but had a positive impact on birds in the USA. In Brazil and Mexico, public perceptions aligned fairly well with the realities of the impacts of potential bioenergy feedstocks on bird communities. However, in Argentina and the USA, perceptions of bioenergy impacts on birds did not match well with the data. Understanding people’s values and perceptions can help inform better policy and management decisions regarding land use changes.
Keywords:
aspen; biodiversity; Elaeis guineensis; eucalyptus; land use change; oil palm; Populus; public perceptions 1. Introduction
Climate change is arguably the greatest environmental and sociopolitical threat of our time. Addressing it will require all available mitigation strategies, including increased development of renewable energy sources [1]. Such development must be tailored to regional energy source availability [2]. Some regions are well-suited to wind or solar development; others may have an abundance of forest and agricultural crop and residue, making bioenergy a viable option. Biogeographic and economic suitability is an essential part of building a portfolio of renewable energy sources (heat, power, or liquid transportation fuels). Today, forest-related bioenergy crops and utilization technologies are contributing to all three energy needs [3].
Despite the promise of renewable energy to mitigate climate change impacts, there are socioeconomic and ecological tradeoffs related to each form of renewable energy. Hydroelectric power from dams can negatively impact aquatic communities including commercial and recreational fisheries [4]. Wind turbines can kill birds, bats, and flying insects and negatively impact property values [5]. Similarly, forest-related bioenergy, i.e., energy produced from forest plantation products such as fruit oil or wood pulp, often involves social–ecological tradeoffs. Forests provide diverse supporting, regulating, provisioning, and cultural ecosystem services [6]. However, there are sometimes trade-offs among these services [7,8]. For example, in some parts of Argentina, converting grazing land to eucalyptus (Eucalyptus spp.) tree plantations for bioenergy decreases ranching jobs while increasing employment in the forestry sector [9]. Bioenergy from eucalyptus may provide a lower net carbon source of heat or electricity compared with natural gas or coal over its life cycle but it may negatively impact rangeland-adapted species that are ecologically important and/or provide key ecosystem services to local people [10].
Mitigating the impact of forest-related bioenergy on biodiversity and associated ecosystem services is essential, because approximately 40% of the Earth’s land surface is already cultivated or grazed by domestic animals [11], and more than 25% of the net primary production of potential global terrestrial vegetation has been appropriated for human use [12]. In fact, one of the greatest challenges facing biodiversity conservation is understanding how land-use change related to agriculture and forestry will influence species composition and persistence [13,14]. Current nature reserves are not sufficient to protect the majority of biodiversity [15], and so it is essential to protect and retain biodiversity and associated ecosystem services in sustainable production landscapes of the future [10,16,17,18,19].
Birds are a component of biodiversity that are relatively easy for people to see, hear, and appreciate [20], and birds are known to provide many ecosystem services in the form of seed dispersal, pest control, scavenging, and pollination [21,22,23,24]. Beyond the material or tangible benefits that birds provide to people, they provide many cultural services as well, which are related to human perception and experience [24]. These types of benefits, which include emotional attachment, spirituality and sense of place, may be just as important to people as the direct economic benefits [25,26]. Some recent studies, primarily in urban areas, have suggested a link between bird species richness (either real or perceived) and psychological well-being [25,27,28].
Birds are highly mobile and often able to quickly evaluate and respond to subtle changes in land cover, making them one of the most cost-effective bioindicators for measuring the effects of land-use change on a broader suite of biodiversity, including whole ecosystems that support many additional ecosystem services [10,29,30]. The value of tree plantations for bird communities has been debated in the literature, e.g., [15,31]. However, the consensus seems to be that plantation impacts on birds depend on several factors; it is necessary to consider the wider context of a plantation forest and to take at least the following into account: (1) previous land use, (2) alternative land uses, (3) plantation tree species, and (4) type of plantation management. These factors as well as the habitat specialization of bird species determine the way that tree plantations influence birds and other biodiversity [10,31].
Here, we focused on ecosystem service tradeoffs between forest habitat-related supporting services for birds (who in turn can provide cultural, regulating, and provisioning services) versus provisioning services associated with bioenergy feedstock production [32]. We conducted social surveys in four bioenergy plantation/cover type landscapes across the Americas to examine residents’ perceptions of the value of birds and the impacts of bioenergy feedstocks on birds. We also conducted bird surveys in each landscape to determine and the relationship of those perceptions with measured characteristics of the bird communities. Our research questions were: (1) do survey respondents value birds; (2) do survey respondents favor bioenergy feedstock expansion; (3) are survey respondent’s perceptions of bioenergy feedstock impacts on birds consistent with the actual measured bird diversity in each type of feedstock? Our study sites included three Latin American nations and the USA: (1) northern Argentina with local wood pellet production from eucalyptus (Eucalyptus spp.) and pine (Pinus elliottii and P. taeda) plantations [33]; (2) northern Brazil, where multiple industrial and smallholders have established oil palm (Elaeis guineensis) plantations for use in biodiesel manufacturing and other food and industrial applications [34]; (3) southern Mexico, where, similar to Brazil, extensive oil palm plantations have been established in the last 30 years on ejido (cooperative), industrial and smallholder lands for a mix of food, fuel, and industrial uses [35,36]. Finally, in the USA, we studied an area in the Midwest where a regional bioelectricity plant was using woody feedstocks from regional aspen (Populus spp.) and northern hardwood forests [37,38,39,40,41,42,43].
This paper makes a unique contribution by integrating ecosystem service measurement of provisioning, supporting and cultural services that some have argued should be more frequently incorporated in assessments [44]. We combine a social survey-based assessment of local values with measures of bioenergy energy feedstock production impacts on bird habitat. We also present potential mitigation strategies for reducing negative habitat impacts on birds and other biodiversity. Our results highlight the importance of understanding local values and how they are being impacted by changes in land use. Understanding these connections can inform better policy and management to protect the diverse values associated with land use changes brought about by expansion of forest plantations or even food production systems. Finally, the paper fills a gap in social–ecological studies that integrate social and ecological data with the goal of making policy and management recommendations [45].
1.1. Background to Study Area
1.1.1. Feedstocks and Human Communities
Argentina: In Argentina, favorable policies have led to local and foreign investment in tree plantations across the country, particularly in the northeastern regions of the country [46,47] where land traditionally used for cattle ranching and agriculture has been converted to plantations [33]. From 1990 to 2015, the area of land dedicated to tree plantations of mostly non-native eucalyptus and pine increased by 40%, and now covers over one million ha in Corrientes, Entre Ríos and Misiones provinces [48]. These fast-growing trees native to Australia (eucalyptus) and the southern United States (pine) were traditionally used for timber and wood pulp for paper, and the woody residues were burned on site to produce power at sawmills [33]. Recently, new renewable energy goals in Argentina provide a different possible end use for these woody residues—pellets for bioenergy production [33]. The first large-scale renewable energy project using woody residues to make wood pellets opened in the city of Concordia, Entre Ríos, in 2015 [49]. In a study of public perceptions of eucalyptus plantations in Entre Ríos, our group [33] found more positive perceptions about tree plantations in a community near large eucalyptus plantations and with a well-developed forestry industry, and more negative perceptions in a community near small eucalyptus plantations and lacking a forestry industry. Both communities shared concerns about labor conditions on tree plantations and loss of cultural values [33].
Brazil: In 2010, the Brazilian federal government launched the Sustainable Palm Oil Production Program (SPOPP) in the Amazonian state of Pará, which attracted large investments from national and international agribusiness and contributed to doubling the area of oil palm in just four years (2010–2014) [50,51]. Oil palm production in the state was marketed as being simultaneously a profitable business, a method of poverty reduction, and a key resource for addressing climate change through bioenergy production and the recovery of degraded pasture lands [52,53]. Policies were designed to restrict oil palm expansion to degraded and marginal lands to avoid further deforestation [54]. Farming via contracts was implemented to increase farmers’ control of their own labor, and the Social Fuel Seal, a government certification, was provided to favor the inclusion of small farmers in the oil palm agrochain [51,55,56]. In this region, oil palm plantations have mostly replaced pasture, traditional crops such as maize, beans, and cassava, and forest fragments [54], and this has led to a rapid process of urbanization in the communities surrounding plantations due to the increase in salaried jobs [51,57]. Communities believed that conversion to oil palm plantations impacted ecosystem services far more negatively than conversion to traditional annual crops, and that water availability, air and water quality were the most heavily impacted ecosystem services [51]. However, the majority of participants supported expansion of oil palm in the region, since they recognized the associated socioeconomic benefits in terms of job opportunities [51].
Mexico: Mexico has experienced heavy local investments in oil palm plantations since the 1980s, with government subsidies and promotion programs favoring oil palm for the purported purposes of generating rural employment, reducing imports of vegetable oil, and producing biodiesel [58,59]. Current oil palm production occurs on both smallholder plots and large plantations, and is concentrated in the states of Campeche, Chiapas, Tabasco, and Veracruz [60]. Several types of government subsidies, totaling more than 10 million USD, were made available for small producers that included cost-share and credit guarantee programs for palm growers, financial support for palm oil processing plants, support for producer cooperatives, technology transfer, and road improvements to facilitate the movement of palm fruit shipments [58]. Local communities and small producers of oil palm in the state of Tabasco tended to benefit more from production if they were organized into a cooperative, based on the traditional ejidal system of shared ownership of land [58]. This likely stemmed from the cooperative providing both a framework for community-level production and a way for them to interact with state and national governments [59,61]. However, some of the community members that had depended on local food cultivation to supplement their diet reported that the transition to oil palm had reduced their access to locally grown food [58]. Furthermore, in communities where workers traveled to work on large oil palm plantations, benefits were uneven and lack of enforcement of labor laws left workers vulnerable to unsafe working conditions [58]. Overall, however, we found that community support for oil palm was relatively high in Tabasco [35]. Most community members believed the crop provided a moderate number of ecosystem services and that the impact of future oil palm expansion would be positive [35].
USA: In the USA, most renewable energy policy is at the state level. More than half of the states have a renewable portfolio standard or similar policy that requires utilities to generate a percentage of their energy from renewable resources [37,62]. For states with extensive forest cover, bioenergy or bioelectricity generated from woody feedstocks is a natural choice to help meet these renewable energy mandates [63]. In Wisconsin, forest covers about half of the state, and for this reason, several utilities are focusing on bioenergy [41]. In the USA, there is broad public support for renewable energy development [64,65]. In Wisconsin, communities near bioelectricity plants generally supported local bioelectricity production, although there were some concerns about potential impacts on timber prices and supplies and local environmental quality [41]. The communities interviewed were also fairly knowledgeable about bioenergy, likely because of the presence of the local bioelectricity plant [37,41]. Finally, the interviews showed that local landowners valued resource-extractive ecosystem services more than other types of ecosystem services generated from forests, such as cultural or regulating ecosystem services [41].
1.1.2. Feedstocks and Bird Communities
Aspen: Aspen (Populus tremuloides and grandidentata in North America and P. tremula in northern Europe) is a group of fast growing, relatively short-lived trees that in many parts of its range regenerates clonally via root sprouts, and with proper management, can be grown in relatively monotypic stands over many cutting cycles [66]. In the northern Great Lake states of the USA, these characteristics have made aspen popular for pulp and saw logs [67]. Stands are managed using a relatively short rotation of cutting at between 30–50 years [66]. Compared to pre-Euro-American settlement forests in this region, aspen (especially stands less than 50 years old) now covers a far larger proportion of the landscape [68,69], which has favored species associated with early successional aspen forests compared to other forests with larger proportions of native trees such as eastern hemlock (Tsuga canadensis) and white pine (Pinus strobus), which have declined [70]. Although both species of aspen are native to eastern North America, their dramatically expanded spatial extent and long history of short-rotation management has led to the widespread persistence of relatively young monotypic stands with a plantation-like age class and tree community structure.
Studies of breeding bird use of aspen forests in the Great Lakes region have shown that these forests support a rich diversity of native species [71,72], including some imperiled species such as the golden-winged warbler (Vermivora chrysoptera) [43]. In the Great Lakes region, aspen is usually managed by clear cutting and allowing root suckers to replace the stand with varying amounts of retained conifer and hardwood species. Aspen breeding bird communities differ based, in part, on the degree of retained legacy trees as well as the age of the dominant aspen [73]. Although there is some overlap between bird communities associated with different age classes of aspen, many bird species found in the youngest ages will abandon stands as the stem density increases and average tree height and diameter expands [74]. Unharvested aspen stands eventually break up as larger trees die and fall and the canopy opens; bird community shifts accompany these changes [72]. In some parts of the range of aspen such as Saskatchewan, Canada, 80–110-year-old stands supported the greatest bird species richness [75], while in northern Wisconsin, it is often the youngest stands (<15 years) that had the highest number of breeding species [42]. It thus appears that there are regional differences in total bird species richness found in different age classes of aspen. Importantly, the widespread finding that breeding bird assemblages change as aspen ages suggests that land and forest managers should strive to maintain large blocks of varying-age aspen stands at the landscape scale [72].
Eucalyptus: Eucalyptus plantations have rapidly increased in extent throughout much of South America, due to their fast growth and utility for wood products and bioenergy [10]. The impacts of these novel, non-native monocultures on native bird communities of the region appear to be very negative. For instance, in the Atlantic forest of Brazil, only eight bird species were recorded in <30-year-old eucalyptus plantations, while more than 100 were recorded in a nearby forest reserve [76]. Similarly, only six bird species were captured in eucalyptus plantations without understory in the Cerrado region of Brazil [77]; understory plants are often controlled in eucalyptus plantations using herbicides or cutting. Avian species richness was also extremely low in eucalyptus plantations in the Brazilian Amazon [78]. When eucalyptus plantations are established in areas where shrub and grassland ecosystems are the native plant communities, the tall tree monocultures contrast greatly with the natural structure and composition of the vegetation, and negatively impact native openland bird communities [79,80]. Nevertheless, these exotic tree plantations can actually be more preferable than many other intensive land uses, such as annual crops or pastoral activities, especially if managed in ways that provide habitat for native biodiversity [31,81]. For instance, eucalyptus plantations managed to include scattered native trees and early successional understory plants were associated with an increase in the proportion of bird species from the regional pool able to occupy the plantations [82].
Oil palm: A recent analysis of land-cover change associated with oil palm plantation expansion in Latin America found that roughly 79% of plantations replaced previously farmed lands, mostly cattle pastures (56%), while the remaining 21% replaced forested areas, especially in the Amazon and in northern Guatemala [83]. This result is in stark contrast to Southeast Asia, where the majority (>50%) of oil palm plantations replaced primary or secondary forest [84]. As the majority of oil palm plantations in Latin America replaced previously farmed land rather than primary or secondary forest [83], the impact of oil palm on biodiversity in this region is expected to be lower than in Southeast Asia. Still, oil palm plantations are a relatively monotypic cover with management for a single species, suggesting that they would support fewer native species compared to native plant communities. In Brazil and Colombia, the functional diversity, in terms of the roles that species play in the ecosystem, as well as the richness of birds, was lower in oil palm plantations than in either forest fragments, riparian corridors or pasture [85,86,87]. In Brazil, those functional traits most affected by oil palm were related to birds’ diet and foraging stratum [85]. Brazilian oil palm also had far fewer birds that are range-restricted or sensitive to environmental perturbation than forest fragments or riparian corridors, and a similar species composition to pastures and agrarian land-uses [86]. However, forest fragments within oil palm plantations contained a relatively species-rich bird community, including several globally threatened species [86].
Using an experimental translocation of individuals, Knowlton et al. [88] found that forest birds living within the forest fragments behaved as if oil palm were a barrier to their movements and preferred to move between fragments via riparian corridors. Similarly, in Venezuela, oil palm and plantain (Musa spp.) plantations contained only birds typical of very disturbed ecosystems; cacao (Theobroma cacao) plantations had greater bird species richness due to the greater structural complexity and lower use of agrochemicals and inorganic fertilizers [89]. In an experimental manipulation of understory vegetation in oil palm plantations in Guatemala, Nájera and Simonetti [15] found that oil palm plantations where understory plants were allowed to persist had higher bird species richness than those where the understory was actively controlled, and concluded that enhancing the understory vegetation in oil palm plantations furnished significant benefits for birds and other animals by providing food resources, refuge and breeding sites [15]. In addition, in Guatemala, bird species richness was highest in forest, followed by guamiles (regeneration sites) and finally oil palm plantations; only 11% of species and 5% of the total captured birds were recorded in oil palm plantations [90].
2. Materials and Methods
2.1. Survey Methods
Our social survey was designed with parallel sets of questions about local bioenergy impacts and expansion as well as socioeconomic and environmental values across communities next to bioenergy projects in all four countries (Argentina, Brazil, Mexico, and the USA, Figure 1). These sites were paired with field studies of bird communities (see Ecological Methods below). The survey was administered in Spanish, English, or Portuguese—the dominant language of each country in which it was conducted.
Figure 1.
Location of the four study sites where ecological and social data were gathered for this paper, and type of bioenergy feedstock examined at each site: Aspen in Wisconsin, USA; oil palm in Tabasco, Mexico and Pará, Brazil, and eucalyptus in Entre Ríos, Argentina.
We randomized household selection within each community and the survey respondent within each household was randomly chosen based on person with the next birthday from each household’s adults over 18 years old. Due to low literacy rates and low familiarity with survey completion, the survey was administered orally in person in Argentina, Brazil, and Mexico. In the USA, it was administered using the drop off/pick up method [91]. This method involves sending households a pre-survey letter, visiting households and meeting the residents to drop the survey off, and returning at a pre-arranged date to collect the completed survey.
In Brazil, we attempted to survey 354 households, with 280 completing the survey for a response rate of 79% percent. Mexico’s response rate was 83% (130 households surveyed out of 156 attempted) and Argentina’s was 75% (215 households surveyed out of 286 attempted). The response rate in the USA was 61% (292 households surveyed out of 478 attempted).
Women were well-represented among respondents. Fifty-one percent of Argentinian respondents were female (Brazil 53%; Mexico 56%; USA 45%). The percentage of Argentinian survey respondents with at least a high school degree was 11% (Brazil 12%; Mexico 19%; USA 90%). Only the minority of respondents’ work depended on the forest or bioenergy sector for most of their income. Argentinian survey respondents included 10% whose household received most of its income from work in the forest or bioenergy sector (Brazil 22%; Mexico 15%; USA 23%).
2.2. Ecological Methods
We assessed avian species richness and abundance within a gradient of biofuel cropping systems in replicated fields/stands in all four case study countries (Argentina, Brazil, Mexico and USA, Figure 1). In each case, we examined bird species richness and abundance across a range of land use intensities, from monocultures that are intensively managed for biofuel feedstock production, to less intensive agroecosystems and natural land cover. Given the intensive nature of the field measurements, we collected data in one country at a time, rotating across the four case studies. The sampling methods differed slightly across countries due to variation in local conditions and land uses.
2.3. Site Description
Argentina: The study region was in the Entre Ríos province of Argentina, near the city of Concordia (31°240 S, 58°20 W). This area is within the pampas, once one of Earth’s greatest native grasslands extending into Uruguay and Brazil [92]. Most of the pampas has now been converted to pasture for livestock grazing and row crop agriculture. Climate in this region ranges from a mean of 12.6 °C in the austral winter to 25.3 °C in the summer, with an average of 1300 mm of rainfall annually [93]. We censused the avian communities in 16 plots, four within each of four common land uses in Entre Ríos: (1) pastures used for livestock grazing and rotational annual crops; (2) large-scale eucalyptus plantations (100 ha); (3) small mixed-use farms with citrus, blueberry (Vaccinium spp.), pastures and small-scale eucalyptus stands (<10 ha); (4) native espinal savanna woodland (see [10]).
Brazil: In Brazil, the eastern Amazonian state of Pará hosts the great majority (>80%) of existing oil palm plantations, within the Belém Area of Endemism (BAE) biodiversity hotspot [94,95]. The study area in Pará state within the Agropalma agroindustrial complex (02°36′18″ S, 48°47′06″ W) consists of 64,000 ha of forest fragments (legal reserves) and 39,000 ha of oil palm plantations. The forest fragments are dominated by terra firme forest and together account for between 5000 and 17,000 ha. Bird surveys were conducted in 24 plots; eight in mature oil palm plantations (>15 years), eight in riparian corridors, and eight in adjacent fragments of native forest (see [85]).
Mexico: The study region in the state of Tabasco is characterized by a hot and humid climate with the highest rainfall of any Mexican state [96]. The vegetation consists of tropical lowland evergreen and semi-deciduous forests with extensive wetlands. The native vegetation has now been almost completely replaced by pasture, banana (Musa spp.) and tree plantations. The mean annual temperature is >25 °C and mean annual rainfall varies from 1500 mm near the coast to 5000 mm in the highlands [97]. Three distinct seasons include a dry period from April to June, a wet period from June to November, and a period of northern winds and heavy rains from December to April [98]. Bird surveys were conducted in 24 plots; six in mature oil palm plantations (18–20 years), six in young oil palm plantations (4–6 years), six in cattle pasture, and six in forest fragments [99].
USA: The study region was within Vilas and Oneida Counties in Wisconsin. Twelve coppiced, even-aged, no-retention aspen forest stands ranging from 8 to 45 years post-harvest were selected and divided into 3 age classes: young (8–15 years), middle (20–44 years), and mature (45 years), for a total of 4 study sites per age class (12 study sites total). Selected sites were ≥16.2 ha in size and ≥5 km from other sites. The study area has a temperate continental climate and mean monthly precipitation of 74 mm and a mean temperature of 19 °C during the birds’ breeding season [100]. All study stands had been allowed to naturally regenerate following harvest (see [72]).
2.4. Point Counts
In each of our study countries, we conducted at least 30 ten-minute 25 m or 50 m radius point counts in each land-use type. Each point was ≥200 m from any other point and ≥50 m from roads and habitat edges. At each point, individual birds were tallied by sight or sound for 8 min [101]. All point counts took place within four hours of sunrise and were conducted at least twice at each site. Aquatic species (e.g., herons and ducks) and species flying over were not considered.
Argentina: We conducted two rounds of point counts in each of the 16 study plots, for a total of 459 points. In each 300 m2 plot, we randomly identified 10-point count locations that were >200 m from one another and >50 m from a cover type edge. For each land use type and transect, one of three observers (who trained together for one month prior to beginning sampling) completed 8 min 25 m fixed-radius point counts at each point and noted all birds seen or heard within 25 m, excluding those flying overhead [102]. Counts were conducted from 1–19 September and 3–27 November 2014 during the breeding season for most local birds. The point counts began near sunrise and were completed within 4 h, allowing two observers to complete two replicates in one day. All counts were conducted in clear, mild weather without rain, fog or excessive wind [10].
Brazil: We censused the bird community twice, once in May and again in December 2012. We used 10 sampling points in each plot, separated by a distance of 200 m from one another, at which 10 min counts were conducted. In total, we sampled 128 points in oil palm plantations and 160 in forested habitats. We recorded the number of individuals of each bird species observed and/or heard within a radius of 50 m of the observer between 05:00 and 10:30 a.m. [85].
Mexico: We demarcated 203 points distributed within the 24 study plots (47 points in secondary forest fragments, 47 in mature oil palm, 53 in young oil palm, and 56 in pasture), and these were sampled three times during the year, covering the migratory and non-migratory periods (June–July 2016, October–December 2016, February–March 2017). The minimum distance between each point was 200 m, and all points were at least 100 m from a habitat edge.
USA: We randomly located 3–12 points within each of the 12 study plots, based on site area; each point was ≥200 m from others to minimize the probability of double-counting individual birds. We also placed points 200 m from roads and stand edges to minimize the impacts of edge effects. There was a cumulative total of 30 points per age class. Only one observer conducted all surveys. Point counts consisted of a 5 min survey within a 25 m fixed radius to reduce differences in bird detectability among stand ages that varied greatly in vegetation density. Each point was surveyed twice: once between 21 May and 3 July 2015 and again between 3 July and 16 July 2015. Except in cases of inclement weather (strong wind and heavy rain), we conducted point counts from sunrise through 10:00 a.m. [72].
2.5. Statistical Analyses
We used ANOVA or Kruskal–Wallis (if data did not meet assumptions of normality) to test for differences in bird species richness and abundance between land-use types within each country. Next, we compared differences in species richness of birds across plantations and alternative land uses across countries using Cohen’s d effect size statistics. Cohen’s d is calculated as the difference in mean species richness between two sites, divided by the pooled standard deviation [103].
3. Results
3.1. Community Survey Results
This section presents descriptive survey results from communities in the four countries regarding the value of birds and bird habitat, support for bioenergy expansion, and beliefs about bioenergy feedstock and harvesting impacts on bird habitat. As the Argentine communities, unlike communities in the other countries, were near significantly different types of bioenergy feedstocks or processing conditions, results for those communities are presented separately. When survey questions were asked somewhat differently across the countries, those differences are indicated.
As Table 1 shows, the vast majority of survey respondents valued birds and/or that forests provide habitat for them. The lowest percentages stating that these things were either important or very important to them were in Brazil (86%) and the USA (83%) but these percentages were still quite high. Their mean scores on this item were similarly high, with respondent scores from most countries/communities averaging 4.5 or higher on a Likert scale set of answers where 5 indicated “very important”. None averaged below 4.3 (still solidly in the “important” category). The standard deviations were similarly low, indicating generally high agreement in responses. Birds and bird habitat were important to these respondents.

Table 1.
How important are birds to you and your household (Argentina, Brazil, Mexico) OR is it important to you that forests provide bird habitat (USA)? † Likert Scale: 1 = Not at all important, 5 = Very important.
Table 2 illustrates how respondents had more mixed views about bioenergy feedstock or harvesting (USA) expansion. The mean scores clustered in the “slightly agree” area with a range of 3.3 to 3.5. Percentages of respondents supporting expansion were also fairly similar with lower levels of support in the USA (48%) and Argentina (55%) and slightly higher levels in Brazil and Mexico (both had 60% support). It is notable, however, that the separate majorities supported expansion in most countries. The exception was the USA where support was expressed by just short of a majority. Overall, a sizable number of respondents supported expansion across the four countries, but a sizeable number was also neutral about or did not support expansion.
Table 2.
Support for bioenergy feedstock or harvesting expansion (“Considering everything, I support the expansion.”). † Likert Scale: 1 = Strongly disagree, 5 = Strongly agree.
Survey respondents were asked whether their local bioenergy feedstock (eucalypts/pine in Argentina, oil palm in Brazil and Mexico) provided habitat for birds (see Table 3). This was asked as a “yes/no” question. Results here varied greatly with about one-third of respondents in Brazil responding that it did provide habitat and significant majorities of those in Argentina (71%) and Mexico (74%) saying that oil palm provided habitat. As the USA case was a change in “end use” (shifting from harvesting for paper pulp or lumber to harvesting for bioenergy) for harvested forest materials, as opposed to a change in “land use” with entirely new, non-native species plantations, this question was not asked there.
Table 3.
Responses to the statement “Bioenergy feedstock cultivation provides bird habitat.” † Likert Scale: 0 = No, 1 = Yes.
Table 4 presents our findings regarding to perceptions of bioenergy feedstock/harvesting expansion and how it would impact birds. This was measured as a five-point Likert scale (in contrast to the question presented in Table 3) and we present our findings focusing more upon respondents who perceived negative effects. Results here varied widely across the countries and feedstocks. Few respondents in Mexico (19%, oil palm) and Argentina (17%, eucalyptus/pine) viewed expansion as negatively impacting bird habitats. In contrast, a slight majority of respondents in Brazil (52%, oil palm) and a near majority of those in the USA (48%, native forest harvesting) believed that expansion would negatively impact birds. Mean response scores varied accordingly, with respondents in Brazil and the USA seeing expansion as just slightly negative and those in Mexico Argentina viewing it as somewhat positive.
Table 4.
Responses to the statement “The expansion of bioenergy feedstock cultivation or harvesting in your area would impact birds [very negatively … very positively].” † Likert Scale: 1 = Very negatively, 5 = Very positively.
3.2. Ecological Results
Argentina: In total, we detected 107 bird species and 2134 individuals. Mean species richness was 6.8 ± 0.85 SE in mature eucalyptus, 11.7 ± 1.20 SE in young eucalyptus, 22.8 ± 2.56 SE in the mixed-use farms, 20.5 ± 2.60 SE in the pasture/annual crops, and 36.3 ± 3.25 SE in the espinal savanna. These differences were significant for eucalyptus compared to all other land uses, and espinal compared to all other land uses (p < 0.001). Compared to the native vegetation, mature eucalyptus plantations had a mean of 81% fewer bird species (Figure 2). Cohen’s d effect size difference between native vegetation and eucalyptus was 6.209. The most frequently detected bird species in the mature eucalyptus plantations were the Rufous-collared Sparrow (Zonotrichia capensis), White-crested Tyrannulet (Serpophaga subcristata), Picazuro Pigeon (Patagioenas picazuro) and Roadside Hawk (Rupornis magnirostris), together accounting for more than 75% of all birds recorded within the plantations [10]. In contrast, the native espinal savanna contained more than 40% of all the bird species detected in the study, including the endangered Yellow Cardinal (Gubernatrix cristata). Furthermore, 27 species were detected only in this habitat.
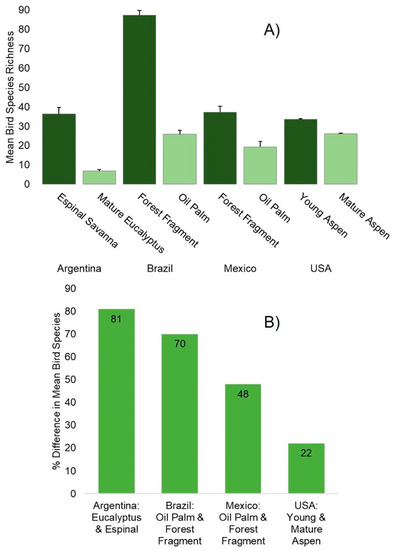
Figure 2.
(A) Differences in mean bird species richness between feedstock/harvested landscapes and native/unharvested landscapes in each of the four study countries. Error bars are +1 standard error of the mean. (B) Percent differences in mean bird species richness between feedstock/harvested landscapes and native/unharvested landscapes in each of the four study countries.
Brazil: We recorded 248 bird species, including 185 species in upland forest, 116 in riparian forest and 58 in oil palm plantations, and a total of 4799 individual birds. The mean species richness was 87.2 ± 7.0 SD in upland forest, 40.2 ± 8.7 in riparian forest, and 25.8 ± 5.7 in oil palm plantations; these differences were significant between all treatments (p < 0.001). Compared to the native vegetation, mature oil palm plantations had a mean of 70% fewer bird species (Figure 2). Cohen’s d effect size difference between native vegetation and oil palm was 9.619. The most frequently detected species in the oil palm plantations were the Southern White-fringed Antwren (Formicivora grisea), Silver-beaked Tanager (Ramphocelus carbo), Palm Tanager (Thraupis palmarum), and Blue-black Grassquit (Volatinia jacarina), which accounted for 30% of all birds seen in the plantations. The upland and riparian forest contained 134 bird species that were never seen in the oil palm plantations, while the oil palm plantations had 71 unique species.
Mexico: We detected a total of 123 bird species and 4333 individuals. Mean species richness was 37.2 ± 7.6 SD in secondary forest fragments, 40.2 ± 6.6 in pasture, 19.3 ± 7.1 in mature oil palm plantations, and 27.3 ± 8.3 in young oil palm plantations. These differences were significant between oil palm and forest fragments and pasture (p < 0.05). Compared to the native vegetation, mature oil palm plantations had a mean of 48% fewer bird species (Figure 2). Cohen’s d effect size difference between native vegetation and oil palm was 2.434. The most frequently detected bird species in the mature oil palm plantations were the Brown Jay (Psilorhinus morio), Great-tailed Grackle (Quiscalus mexicanus), American Redstart (Setophaga ruticilla), and Gray Catbird (Dumetella carolinensis), accounting for 57% of the birds seen in the mature plantations. The forest fragments contained 40 species not detected in mature oil palm, while mature oil palm had 10 bird species not seen in forest.
USA: A total of 972 individuals from 53 bird species were detected from 180 point-counts. Young, middle, and mature stands had an average species richness of 33.5 (SE ± 0.33), 28.0 (± 0.22) and 26.0 (± 0.49) species, respectively. Species richness was not significantly different among age classes (p = 0.58). Compared to the young stands, mature aspen stands had a mean of 22% fewer bird species (Figure 2). Cohen’s d effect size difference between young and mature aspen was 1.616. The most frequently detected species in both mature and middle-age stands were Red-eyed Vireo (Vireo olivaceus), Ovenbird (Seiurus aurocapilla), and Black-capped Chickadee (Poecile atricapillus), accounting for 40% of birds sighted in both age classes stands. In young stands, Chestnut-sided Warbler (Setophaga pensylvanica), White-throated Sparrow (Zonotrichia albicollis), Ovenbird, and Nashville Warbler (Leiothlypis ruficapilla) were the most abundant, accounting for 36% of all birds in this habitat. Golden-winged Warbler (Vermivora chrysoptera), a near-threatened species, was only recorded in young aspen stands. Five unique species were recorded in young stands, four in middle-age stands, and three in mature stands.
3.3. Integrated Social–Ecological Results
Here, we integrate the social survey and ecological findings by comparing respondent perceptions to actual impacts of bioenergy related land use/end use change (Table 5).
Table 5.
Combined and ranked percentage survey respondents saying bioenergy expansion would negatively impact birds and ecological data effect size for bioenergy feedstock versus native forest bird habitat. Bold numbers indicate a match between the public perception of feedstock impacts and the actual ecological impacts.
The country with the highest percentage of respondents rating bioenergy feedstock expansion as negatively impacting birds was Brazil, followed closely by the USA, then Mexico and Argentina; respondents from the last two countries, on average, expected that bioenergy expansion would have a positive impact on birds. Based on the effect size comparison, the country with the feedstock that had the largest negative impact on birds was Brazil, followed closely by Argentina and then Mexico and finally the USA. It is important to remember that in the USA, the young, recently harvested stands actually had more bird species than older ones. Brazil, the region with the most negative bioenergy-related bird habitat impacts, was also the region where the respondents viewed bioenergy expansion most negatively with regard to impacts on bird habitats. However, the site with the next most negative respondent perceptions was the USA, and this was actually the one place where bioenergy had the smallest, and arguably positive, impact on birds. Mexico had the third-place ranking in terms of percentage of people who thought bioenergy had a negative impact on birds, and this social–ecological ranking matched up with Mexico having the third most negative impact on birds. Negative impacts on birds from eucalyptus plantations were significant in Argentina, coming in at second place, but few respondents saw them as such.
4. Discussion
Local environmental perceptions are rarely studied or considered, and yet these perceptions can play a large role in ecosystem conservation [104,105]. We found that respondents in all countries placed a high value on birds, rating them as important or very important (83–99% of respondents). We also found that these respondents showed a lower but still moderate enthusiasm for the expansion of bioenergy feedstocks (48–60% of respondents). In terms of perceived impacts of bioenergy feedstocks on birds, the country with communities near bioenergy projects that had the most negative impact on bird habitats (Brazil) also had the fewest respondents seeing the bioenergy feedstock as providing bird habitat. Interestingly, in Brazil, respondents were also more likely to see bioenergy expansion as negatively impacting birds, although this number was still only roughly half of the respondents. In Mexico and Argentina, few participants surveyed recognized the negative impacts of bioenergy feedstocks on birds. In the USA, respondents were nearly as likely to view expansion as negative for birds as in Brazil (almost half of respondents). However, in the USA, aspen management for bioenergy may actually have a somewhat positive impact on bird habitat.
The fact that all country communities: (1) valued birds highly and (2) generally supported bioenergy expansion, shows the challenges related to social choice and ecosystem service trade-offs. People may either lack an understanding of the impacts of changing land uses on birds or have the desire to benefit from bioenergy while minimizing impacts to birds and other wildlife. Although ecosystems can simultaneously generate multiple services, it is often not possible to manage them to maximize all services, resulting in trade-offs [106,107,108]. Evidence from our study and others suggests that, although communities often correctly perceive negative impacts of changing land uses to ecosystem services such as those provided by biodiversity, they often still support the change in land use if it means higher wages [51,109].
In this study, the ecosystem service trade-offs were between the provisioning service of fuel generation and the cultural, regulating, and supporting services that birds provide. The information we gathered presents an opportunity to communicate to landowners, managers, and policy makers the importance of understanding where land use and commodity production can support multiple values related to biodiversity. Moreover, our results demonstrate the need to craft policy to provide outreach to communities and landowners about the actual bird impacts and about strategies to mitigate them if bioenergy feedstock expansion continues. In the case of the USA, it does appear that the multiple ecosystem services of fuel generation and quality bird habitat can be met simultaneously, which should also be communicated to community members and policy makers. That said, it may be that respondents viewed bioenergy expansion as a change in cover type rather than an intensification of an existing management practice or alternate end point for the harvested material.
In Brazil, our findings on the effect of large-scale oil palm plantations on the bird community were consistent with similar studies that show pronounced negative impacts, e.g., [86]. The majority of birds detected in the oil palm were generalist species known to do well in human-altered landscapes, while over 100 species were seen only in the forest fragments. It is important for landowners, managers, and policy makers to understand that local community members place a high value on birds, and that they mostly recognize that oil palm can negatively impact their habitat. However, despite this awareness, 60% of respondents still supported oil palm expansion, illustrating the challenge facing tropical biodiversity conservation. Given that the Brazilian government incentivizes oil palm expansion, knowing that local people value birds could affect their bioenergy policies. On the other hand, the majority of people supported expansion of oil palm in the region, emphasizing the socioeconomic benefits of this crop in terms of stable jobs and more job opportunities [51]. Similar to our results, a study in Indonesia showed that while local people recognized the negative impacts oil palm plantations have on some ecosystem services such as wildlife habitat, clean water, and cultural services, the majority were still in favor of the oil palm plantations because of the extra income they provided [109]. If oil palm expansion continues, the ecological data suggest that there are key strategies for mitigating some of the negative impacts on birds through retention of nearby riparian and native forest habitats (also see [88]), which may lessen the trade-offs between ecosystem services.
In Mexico and Argentina, respondents were less likely to recognize the negative impacts of oil palm and eucalyptus/pine expansion on birds. In Argentina, we found large negative impacts of eucalyptus expansion on birds (also see [10]), and yet only 17% of respondents thought that this feedstock expansion harmed birds. The vast majority of birds we detected in the mature eucalyptus plantations were habitat generalists that are common near human habitation [10], while the native espinal savannah contained 27 unique species including Pampas endemics and one endangered species. Our previous work in this country showed that communities near large eucalyptus plantations with a strong forestry industry had many members employed by this industry, and this likely contributed to their positive perceptions of these plantations [33]. Furthermore, afforested pasture probably appears less detrimental to wildlife than the cutting of native forest and planting of oil palm to the casual viewer. It would be informative to examine how community members’ views on the expansion of eucalyptus might change if they knew that these plantations were detrimental to bird diversity. However, as in the Brazilian case, we suggest that they would still support the expansion as long as it provided extra income.
In Mexico, most community members did not tend to see oil palm as negatively impacting birds, and the ecological data showed that the actual impacts were negative but not nearly as bad as in Brazil. We found that while most species seen using the oil palm were habitat generalists, many migratory birds were also detected. However, the secondary forest fragments contained 40 species not seen in oil palm. The oil palm plantations in Mexico (especially in the state of Tabasco where we worked) tend to be much smaller than those in Brazil, and there is evidence that bird diversity and abundance are higher in smallholder oil palm plantations than in large estates [110,111]. Thus, the public perceptions in both Brazil and Mexico are fairly realistic in terms of the actual impacts to birds and their habitats.
Finally, respondents in the USA were likely to see bioenergy harvesting as negatively impacting the birds they valued even though it was actually having a positive impact on total bird species richness in aspen-dominated northern Wisconsin forests. We found that young, middle and mature aspen stands all harbored distinct bird communities, pointing to the benefits of maintaining a heterogeneous mosaic of aspen stand ages for conserving bird diversity at the landscape scale [72,75]. Furthermore, the most recently cut stands (young age class) are the preferred habitat for the near-threatened Golden-winged Warbler, and thus should be maintained in the landscape [43]. In this case, there appears to be the rare circumstance of commodity provisioning and biodiversity support being provided simultaneously. This information provides a key opportunity to communicate these findings to policy makers, especially given that many U.S. states (including Wisconsin) have policies requiring renewable energy consumption, including from bioelectricity, a policy that has incentivized increased production. It would be valuable to communicate these positive impacts (at least with the current mature–young forest mix) to the public, landowners, and surrounding communities.
Public knowledge about the services people receive from wildlife, whether cultural or other, along with perceptions of the abundance or scarcity of local wildlife, often influences the value individuals place on wild species and can influence their motivation to support various policies or management strategies [112]. Therefore, individuals’ perceptions can play a critical role in effective resource management [112,113]. For instance, people may not be motivated to adopt a conservation strategy for birds or other wildlife if they do not perceive certain agricultural landscapes as detrimental to these species [114]. Furthermore, if people have no knowledge of the many ecosystem services provided by birds, such as pest control, pollination, and seed dispersal, there may be little incentive to ensure that these species thrive [115,116]. Taking stock of perceptions and values placed on wildlife and ecosystem services and communicating the results of scientific studies in ways that can be understood by local stakeholders are crucial elements of formulating plans for sustainable resource management and navigating trade-offs in the future [112].
5. Conclusions
This paper is a rare example of a cross-national, social–ecological system (SES) study that uses empirically based primary social and ecological data to better understand the SES system and to make policy and management recommendations [45]. It demonstrates the complexity of assessing relative ecosystem service tradeoffs within such a context and illustrates a path forward to better integrating SES data. We found that while all respondents placed a high value on birds, local communities’ perceptions of the impact of bioenergy development on birds is context-specific. In Brazil and Mexico, public perceptions aligned fairly well with the realities of the impacts of bioenergy feedstocks on bird communities. In these cases, policies should favor small-scale family or community owned oil palm plantations over large ones and emphasize the importance of heterogeneity, including polyculture food crops, to allow for landscapes that provide multiple ecosystem services and support biodiversity [34,88]. In Argentina and the USA, perceptions of bioenergy impacts on birds did not match well with the data, and education regarding land uses and their impacts on ecosystem services and biodiversity should be a priority. There are no clear patterns regarding these mismatches, as the two countries differ greatly in time since bioenergy adoption and the level of contrast of these plantations with previous land uses. Future research into how people form their perceptions of land use impacts, as well as on wildlife and ecosystem services, would help land managers and conservation biologists know how to better communicate with surrounding communities.
Author Contributions
Conceptualization, J.L.K. and K.E.H.; methodology, all authors.; validation, all authors; formal analysis, K.E.H. and J.L.K.; data collection, J.A., S.L.A.-W., C.C.P., S.M.A., B.B., M.R.C., P.V.C., D.C., M.P.D.-S., J.L.D., A.E., G.M.J., J.A.L., E.M.-Z., R.M., M.A.M.-J., L.Y.M.-C., E.C.P., T.S. (Tatiana Souza), C.A.S.; resources, S.L.A.-W., C.M., E.N., C.S., S.R.S., T.S. (Theresa Selfa), C.J.V.-N.; data curation, J.L.K., K.E.H.; writing—original draft preparation, J.L.K., K.E.H.; writing—review and editing, C.R.W., D.J.F.; project administration, K.E.H.; funding acquisition, K.E.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Science Foundation (NSF) Partnerships in International Research and Education (PIRE) grant #1243444, NSF International Research Experience for Undergraduates (IREU) grant DEB–1019928, Inter-American Institute for Global Change Research (IAI) Coordinated Research Network grant #CRN3105, and the USAID program for two grants to our Mexican and Brazilian research team members.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board of the University of Oregon (#09242012 approved 12 May 2014).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author, excluding human subjects’ data.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Owusu, P.A.; Asumadu-Sarkodie, S. A review of renewable energy sources, sustainability issues and climate change mitigation. Cogent Eng. 2016, 3, 1167990. [Google Scholar] [CrossRef]
- Santangeli, A.; Toivonen, T.; Pouzols, F.M.; Pogson, M.; Hastings, A.; Smith, P.; Moilanen, A. Global change synergies and trade-offs between renewable energy and biodiversity. GCB Bioenergy 2016, 8, 941–951. [Google Scholar] [CrossRef]
- Scarlat, N.; Dallemand, J.F.; Monforti-Ferrario, F.; Nita, V. The role of biomass and bioenergy in a future bioeconomy: Policies and facts. Environ. Dev. 2015, 15, 3–34. [Google Scholar] [CrossRef]
- Botelho, A.; Ferreira, P.; Lima, F.; Pinto, L.M.C.; Sousa, S. Assessment of the environmental impacts associated with hydropower. Renew. Sustain. Energy Rev. 2017, 70, 896–904. [Google Scholar] [CrossRef]
- May, R.; Masden, E.A.; Bennet, F.; Perron, M. Considerations for upscaling individual effects of wind energy development towards population-level impacts on wildlife. J. Environ. Manag. 2019, 230, 84–93. [Google Scholar] [CrossRef]
- Felipe-Lucia, M.R.; Soliveres, S.; Penone, C.; Manning, P.; van der Plas, F.; Boch, S.; Prati, D.; Ammer, C.; Schall, P.; Gossner, M.M.; et al. Multiple forest attributes underpin the supply of multiple ecosystem services. Nat. Commun. 2018, 9, 4839. [Google Scholar] [CrossRef]
- Martín-López, B.; Gómez-Baggethun, E.; García-Llorente, M.; Montes, C. Trade-offs across value-domains in ecosystem services assessment. Ecol. Indic. 2014, 37, 220–228. [Google Scholar] [CrossRef]
- Mora, F.; Balvanera, P.; García-Frapolli, E.; Castillo, A.; Trilleras, J.M.; Cohen-Salgado, D.; Salmerón, O. Trade-offs between ecosystem services and alternative pathways toward sustainability in a tropical dry forest region. Ecol. Soc. 2016, 21, 45. [Google Scholar] [CrossRef]
- Mwanukuzi, P.K. Impact of eucalyptus and pine growing on rural livelihood: The lesson from Bukoba area, north western Tanzania. Afr. J. Ecol. 2009, 47, 105–109. [Google Scholar] [CrossRef]
- Phifer, C.C.; Knowlton, J.L.; Webster, C.R.; Flaspohler, D.J.; Licata, J.A. Bird community responses to afforested eucalyptus plantations in the Argentine pampas. Biodivers. Conserv. 2017, 26, 3073–3101. [Google Scholar] [CrossRef]
- Froehlich, H.E.; Runge, C.A.; Gentry, R.R.; Gaines, S.D.; Halpern, B.S. Comparative terrestrial feed and land use of an aquaculture-dominant world. Proc. Natl. Acad. Sci. USA 2018, 115, 5295–5300. [Google Scholar] [CrossRef] [PubMed]
- Krausmann, F.; Erb, K.H.; Gingrich, S.; Haberl, H.; Bondeau, A.; Gaube, V.; Lauk, C.; Plutzar, C.; Searchinger, T.D. Global human appropriation of net primary production doubled in the 20th century. Proc. Natl. Acad. Sci. USA 2013, 110, 10324–10329. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Zimmer, C. Black is the new green. Conserv. Mag. 2010. Available online: https://www.conservationmagazine.org/2010/08/black-is-the-new-green/ (accessed on 12 December 2018).
- Nájera, A.; Simonetti, J.A. Enhancing Avifauna in Commercial Plantations. Conserv. Biol. 2010, 24, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Perfecto, I.; Vandermeer, J. The agroecological matrix as alternative to the land-sparing/agriculture intensification model. Proc. Natl. Acad. Sci. USA 2010, 107, 5786–5791. [Google Scholar] [CrossRef]
- Pascual, U.; Perrings, C. Developing incentives and economic mechanisms for in situ biodiversity conservation in agricultural landscapes. Agric. Ecosyst. Environ. 2007, 121, 256–268. [Google Scholar] [CrossRef]
- Tomich, T.P.; Brodt, S.; Ferris, H.; Galt, R.; Horwath, W.R.; Kebreab, E.; Leveau, J.H.J.; Liptzin, D.; Lubell, M.; Merel, P.; et al. Agroecology: A Review from a Global-Change Perspective. Annu. Rev. Environ. Resour. 2011, 36, 193–222. [Google Scholar] [CrossRef]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef]
- Cox, D.T.C.; Gaston, K.J. Urban Bird Feeding: Connecting People with Nature. PLoS ONE 2016, 11, e0158717. [Google Scholar] [CrossRef] [PubMed]
- Şekercioğlu, C.H. Increasing awareness of avian ecological function. Trends Ecol. Evol. 2006, 21, 464–471. [Google Scholar] [CrossRef]
- Whelan, C.J.; Şekercioğlu, Ç.H.; Wenny, D.G. Why birds matter: From economic ornithology to ecosystem services. J. Ornithol. 2015, 156, 227–238. [Google Scholar] [CrossRef]
- Whelan, C.J.; Wenny, D.G.; Marquis, R.J. Ecosystem Services Provided by Birds. Ann. N. Y. Acad. Sci. 2008, 1134, 25–60. [Google Scholar] [CrossRef] [PubMed]
- Michel, N.L.; Whelan, C.J.; Verutes, G.M. Ecosystem services provided by Neotropical birds. Condor 2020, 122, duaa022. [Google Scholar] [CrossRef]
- Belaire, J.A.; Westphal, L.M.; Whelan, C.J.; Minor, E.S. Urban residents’ perceptions of birds in the neighborhood: Biodiversity, cultural ecosystem services, and disservices. Condor 2015, 117, 192–202. [Google Scholar] [CrossRef]
- Chan, K.M.A.; Guerry, A.D.; Balvanera, P.; Klain, S.; Satterfield, T.; Basurto, X.; Bostrom, A.; Chuenpagdee, R.; Gould, R.; Halpern, B.S.; et al. Where are Cultural and Social in Ecosystem Services? A Framework for Constructive Engagement. Bioscience 2012, 62, 744–756. [Google Scholar] [CrossRef]
- Dallimer, M.; Irvine, K.N.; Skinner, A.M.J.; Davies, Z.G.; Rouquette, J.R.; Maltby, L.L.; Warren, P.H.; Armsworth, P.R.; Gaston, K.J. Biodiversity and the Feel-Good Factor: Understanding Associations between Self-Reported Human Well-being and Species Richness. Bioscience 2012, 62, 47–55. [Google Scholar] [CrossRef]
- Shwartz, A.; Turbé, A.; Simon, L.; Julliard, R. Enhancing urban biodiversity and its influence on city-dwellers: An experiment. Biol. Conserv. 2014, 171, 82–90. [Google Scholar] [CrossRef]
- Gardner, T.A.; Barlow, J.; Araujo, I.S.; Ávila-Pires, T.C.; Bonaldo, A.B.; Costa, J.E.; Esposito, M.C.; Ferreira, L.V.; Hawes, J.; Hernandez, M.I.M.; et al. The cost-effectiveness of biodiversity surveys in tropical forests. Ecol. Lett. 2008, 11, 139–150. [Google Scholar] [CrossRef]
- Gardner, T.A.; Barlow, J.; Parry, L.W.; Peres, C.A. Predicting the uncertain future of tropical forest species in a data vacuum. Biotropica 2007, 39, 25–30. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Daniel, T.C.; Muhar, A.; Arnberger, A.; Aznar, O.; Boyd, J.W.; Chan, K.M.; Costanza, R.; Elmqvist, T.; Flint, C.G.; Gobster, P.H.; et al. Contributions of cultural services to the ecosystem services agenda. Proc. Natl. Acad. Sci. USA 2012, 109, 8812–8819. [Google Scholar] [CrossRef]
- Silva, C. Community Perceptions of Ecosystem Services and Human Well-Being from Tree Plantations in Argentina. Master’s Thesis, Northern Arizona University, Flagstaff, AZ, USA, 2016. [Google Scholar]
- Córdoba, D.; Selfa, T.; Abrams, J.B.; Sombra, D. Family farming, agribusiness and the state: Building consent around oil palm expansion in post-neoliberal Brazil. J. Rural Stud. 2018, 57, 147–156. [Google Scholar] [CrossRef]
- Pischke, E.C.; Rouleau, M.D.; Halvorsen, K.E. Public perceptions towards oil palm cultivation in Tabasco, Mexico. Biomass Bioenergy 2018, 112, 1–10. [Google Scholar] [CrossRef]
- Pischke, E.C.; Azahara Mesa-Jurado, M.; Eastmond, A.; Abrams, J.; Halvorsen, K.E. Community perceptions of socioecological stressors and risk-reducing strategies in Tabasco, Mexico. J. Environ. Stud. Sci. 2018, 8, 441–451. [Google Scholar] [CrossRef]
- Banerjee, A.; Schelly, C.; Halvorsen, K.E. Understanding public perceptions of wood-based electricity production in Wisconsin, United States: The place-based dynamics of social representations. Environ. Sociol. 2017, 3, 381–393. [Google Scholar] [CrossRef]
- Banerjee, A.; Schelly, C.L.; Halvorsen, K.E. Constructing a sustainable bioeconomy: Multi-scalar perceptions of sustainability. In World Sustainability Series; Springer: Berlin/Heidelberg, Germany, 2018; pp. 355–374. [Google Scholar]
- Barnett, J. Addressing Policy Challenges to Woody Biopower Production: Social Acceptance, Biomass Certification and Limited Policy Support. Ph.D. Thesis, Michigan Technological University, Houghton, MI, USA, 2018. [Google Scholar]
- Abrams, J.; Nielsen, E.; Diaz, D.; Selfa, T.; Adams, E.; Dunn, J.L.; Moseley, C. How do states benefit from nonstate governance? Evidence from forest sustainability certification. Glob. Environ. Polit. 2018, 18, 66–85. [Google Scholar] [CrossRef]
- Dunn, J. Wisconsin Non-Industrial Private Forest Landowners and Wood-Based Bioenergy. PhD Thesis, Michigan Technological University, Houghton, MI, USA, 2018. [Google Scholar]
- Roth, A.M. Retention of Canopy Trees as Biological Legacies for Balancing Woody-Biomass Production and Biodiversity in Managed Aspen Forests of the Great Lakes Region. Ph.D. Thesis, Michigan Technological University, Houghton, MI, USA, 2012. [Google Scholar]
- Roth, A.M.; Flaspohler, D.J.; Webster, C.R. Legacy tree retention in young aspen forest improves nesting habitat quality for Golden-winged Warbler (Vermivora chrysoptera). For. Ecol. Manag. 2014, 321, 61–70. [Google Scholar] [CrossRef]
- De Groot, R.S.; Alkemade, R.; Braat, L.; Hein, L.; Willemen, L. Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making. Ecol. Complex. 2010, 7, 260–272. [Google Scholar] [CrossRef]
- Guerrero, A.M.; Bennett, N.J.; Wilson, K.A.; Carter, N.; Gill, D.; Mills, M.; Ives, C.D.; Selinske, M.J.; Larrosa, C.; Bekessy, S.; et al. Achieving the promise of integration in social-ecological research: A review and prospectus. Ecol. Soc. 2018, 23, 38. [Google Scholar] [CrossRef]
- Cubbage, F.; Koesbandana, S.; Mac Donagh, P.; Rubilar, R.; Balmelli, G.; Olmos, V.M.; De La Torre, R.; Murara, M.; Hoeflich, V.A.; Kotze, H.; et al. Global timber investments, wood costs, regulation, and risk. Biomass Bioenergy 2010, 34, 1667–1678. [Google Scholar] [CrossRef]
- Cubbage, F.; Mac Donagh, P.; Sawinski, J.; Rubilar, R.; Donoso, P.; Ferreira, A.; Hoeflich, V.; Olmos, V.M.; Ferreira, G.; Balmelli, G.; et al. Timber investment returns for selected plantations and native forests in South America and the southern United States. New For. 2007, 33, 237–255. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2015; UN Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Ministerio de Cultura y Comunicación. En Entre Ríos se produce energía alternativa a partir de desechos de la madera—Noticias—Secretaría de Comunicación de la Provincia de Entre Ríos. 2015. Available online: http://www.entrerios.gob.ar/noticias/nota.php?id=41393 (accessed on 5 January 2016).
- Benami, E.; Curran, L.M.; Cochrane, M.; Venturieri, A.; Franco, R.; Kneipp, J.; Swartos, A. Oil palm land conversion in Pará, Brazil, from 2006-2014: Evaluating the 2010 Brazilian Sustainable Palm Oil Production Program. Environ. Res. Lett. 2018, 13, 34037. [Google Scholar] [CrossRef]
- Córdoba, D.; Juen, L.; Selfa, T.; Peredo, A.M.; de Montag, L.F.A.; Sombra, D.; Santos, M.P.D. Understanding local perceptions of the impacts of large-scale oil palm plantations on ecosystem services in the Brazilian Amazon. For. Policy Econ. 2019, 109, 102007. [Google Scholar] [CrossRef]
- Córdoba, D.; Chiappe, M.; Abrams, J.; Selfa, T. Fuelling Social Inclusion? Neo-extractivism, State-Society Relations and Biofuel Policies in Latin America’s Southern Cone. Dev. Chang. 2018, 49, 63–88. [Google Scholar] [CrossRef]
- Pacheco, P.; Gnych, S.; Dermawan, A.; Komarudin, H.; Okarda, B. The Palm Oil Global Value Chain: Implications for Economic Growth and Social and Environmental Sustainability; Center for International Forestry Research (CIFOR): Bogor, Indonesia, 2017. [Google Scholar]
- Monteiro De Carvalho, C.; Silveira, S.; La Rovere, E.L.; Iwama, A.Y. Deforested and degraded land available for the expansion of palm oil for biodiesel in the state of Pará in the Brazilian Amazon. Renew. Sustain. Energy Rev. 2015, 44, 867–876. [Google Scholar] [CrossRef]
- Moreira, J.R.; Goldemberg, J. Oil palm deserves government attention in Brazil. Environ. Res. Lett. 2015, 10, 71002. [Google Scholar] [CrossRef]
- Embrapa. Zoneamento Agroecológico, Produção e Manejo da Cultura de Palma de Óleo na Amazônia (Agro-Ecological Zoning, Production and Management for Growing Oil Palm in the Amazon); Embrapa: Rio de Janeiro, Brazil, 2010. [Google Scholar]
- Nahum, J.S.; Santos, C.B. Impactos socioambientais da dendeicultura em comunidades tradicionais na Amazônia paraense. Rev. ACTA Geográfica 2013, 63–80. [Google Scholar] [CrossRef]
- Abrams, J.; Pischke, E.C.; Mesa-Jurado, M.A.; Eastmond, A.; Silva, C.A.; Moseley, C. Between Environmental Change and Neoliberalism: The Effects of Oil Palm Production on Livelihood Resilience. Soc. Nat. Resour. 2019, 32, 548–565. [Google Scholar] [CrossRef]
- Castellanos-Navarrete, A.; Jansen, K. Oil palm expansion without enclosure: Smallholders and environmental narratives. J. Peasant Stud. 2015, 42, 791–816. [Google Scholar] [CrossRef]
- Isaac-Márquez, R.; Valladares, J.L.S.; Spencer, A.E.; Arcipreste, M.E.A.; Aguilar, M.A.A.; Márquez, A.P.I.; González, M.C.S. Impactos Sociales y Ambientales de la Palma de Aceite: Perspectiva de los Campesinos en Campeche, México. J. Lat. Am. Geogr. 2016, 15, 123–146. [Google Scholar] [CrossRef]
- Castellanos-Navarrete, A.; Jansen, K. Is Oil Palm Expansion a Challenge to Agroecology? Smallholders Practising Industrial Farming in Mexico. J. Agrar. Chang. 2018, 18, 132–155. [Google Scholar] [CrossRef]
- Barbose, G.; Bird, L.; Heeter, J.; Flores-Espino, F.; Wiser, R. Costs and benefits of renewables portfolio standards in the United States. Renew. Sustain. Energy Rev. 2015, 52, 523–533. [Google Scholar] [CrossRef]
- Ebers, A.; Malmsheimer, R.W.; Volk, T.A.; Newman, D.H. Inventory and classification of United States federal and state forest biomass electricity and heat policies. Biomass Bioenergy 2016, 84, 67–75. [Google Scholar] [CrossRef]
- Devine-Wright, P. Reconsidering Public Attitudes and Public Acceptance of Renewable Energy Technologies: A Critical Review; School of Environment and Development, University of Manchester: Manchester, UK, 2007. [Google Scholar]
- Farhar, B.C. Willingness to Pay for Electricity from Renewable Resources: A Review of Utility Market Research. Renew. Energy Lab. 1999, 550, 1–20. [Google Scholar]
- Alban, D.; Perala, D.A.; Jurgensen, M.F.; Ostry, M.E.; Probst, J.R. Aspen Ecosystem Properties in the Upper Great Lakes; North Central Forest Experiment Station, Forest Service—US Department of Agriculture: St. Paul, MN, USA, 1991. [Google Scholar]
- Blyth, J.E.; Smith, W.B. Pulpwood Production in the North Central Region by County, 1987; Resource Bulletin NC-111; U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1989.
- Cole, K.L.; Davis, M.B.; Stearns, F.; Guntenspergen, G.; Walker, K. Historical landcover changes in the Great Lakes region. In Perspectives on the Land Use History of North. America; Sisk, T.D., Ed.; U.S. Geological Survey, Biological Resources Division: Reston, VA, USA, 1999; pp. 43–50. [Google Scholar]
- White, M.A.; Mladenoff, D.J. Old-growth forest landscape transitions from pre-European settlement to present. Landsc. Ecol. 1994, 9, 191–205. [Google Scholar] [CrossRef]
- Schulte, L.A.; Mladenoff, D.J.; Crow, T.R.; Merrick, L.C.; Cleland, D.T. Homogenization of northern U.S. Great Lakes forests due to land use. Landsc. Ecol. 2007, 22, 1089–1103. [Google Scholar] [CrossRef]
- Niemi, G.J.; Hanowski, J.M. Relationships of Breeding Birds to Habitat Characteristics in Logged Areas. J. Wildl. Manag. 1984, 48, 438–443. [Google Scholar] [CrossRef]
- Jarvi, G.M.; Knowlton, J.L.; Phifer, C.C.; Roth, A.M.; Webster, C.R.; Flaspohler, D.J. Avian Community Response to Short-rotation Aspen Forest Management. Northeast. Nat. 2018, 25, 308–318. [Google Scholar] [CrossRef]
- Niemi, G.J.; Howe, R.W.; Sturtevant, B.R.; Parker, L.R.; Grinde, A.R.; Danz, N.P.; Nelson, M.D.; Zlonis, E.J.; Walton, N.G.; Giese, E.E.G.; et al. Analysis of Long-Term Forest Bird Monitoring Data from National Forests of the Western Great Lakes Region; Gen. Tech. Rep. NRS-159; US Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2016.
- Moskát, C.; Szekely, T. Habitat distribution of breeding birds in relation to forest succession. Folia Zool. 1989, 38, 363–376. [Google Scholar]
- Hobson, K.A.; Bayne, E. The effects of stand age on avian communities in aspen-dominated forests of central Saskatchewan, Canada. For. Ecol. Manag. 2000, 136, 121–134. [Google Scholar] [CrossRef]
- Marsden, S.J.; Whiffin, M.; Galetti, M. Bird diversity and abundance in forest fragments and Eucalyptus plantations around an Atlantic forest reserve, Brazil. Biodivers. Conserv. 2001, 10, 737–751. [Google Scholar] [CrossRef]
- Piratelli, A.; Blake, J.G. Bird communities of the southeastern Cerrado Region, Brazil. Ornitol. Neotrop. 2006, 17, 213–225. [Google Scholar]
- Barlow, J.; Mestre, L.A.M.; Gardner, T.A.; Peres, C.A. The value of primary, secondary and plantation forests for Amazonian birds. Biol. Conserv. 2007, 136, 212–231. [Google Scholar] [CrossRef]
- Calviño-Cancela, M. Effectiveness of eucalypt plantations as a surrogate habitat for birds. For. Ecol. Manag. 2013, 310, 692–699. [Google Scholar] [CrossRef]
- Dias, R.A.; Bastazini, V.A.G.; Gonçalves, M.S.S.; Bonow, F.C.; Müller, S.C. Alterações na composição de comunidades de aves relacionadas ao florestamento de campos temperados no Sudeste da América do su. Iheringia Ser. Zool. 2013, 103, 12–19. [Google Scholar] [CrossRef]
- Simonetti, J.A.; Grez, A.A.; Estades, C.F. Providing Habitat for Native Mammals through Understory Enhancement in Forestry Plantations. Conserv. Biol. 2013, 27, 1117–1121. [Google Scholar] [CrossRef]
- Millan, C.H.; Develey, P.F.; Verdade, L.M. Stand-level management practices increase occupancy by birds in exotic Eucalyptus plantations. For. Ecol. Manag. 2015, 336, 174–182. [Google Scholar] [CrossRef]
- Furumo, P.R.; Aide, T.M. Characterizing commercial oil palm expansion in Latin America: Land use change and trade. Environ. Res. Lett. 2017, 12, 024008. [Google Scholar] [CrossRef]
- Koh, L.P.; Wilcove, D.S. Is oil palm agriculture really destroying tropical biodiversity? Conserv. Lett. 2008, 1, 60–64. [Google Scholar] [CrossRef]
- Almeida, S.M.; Silva, L.C.; Cardoso, M.R.; Cerqueira, P.V.; Juen, L.; Santos, M.P.D. The effects of oil palm plantations on the functional diversity of Amazonian birds. J. Trop. Ecol. 2016, 32, 510–525. [Google Scholar] [CrossRef]
- Lees, A.C.; Moura, N.G.; de Almeida, A.S.; Vieira, I.C.G. Poor Prospects for Avian Biodiversity in Amazonian Oil Palm. PLoS ONE 2015, 10, e0122432. [Google Scholar] [CrossRef] [PubMed]
- Prescott, G.W.; Gilroy, J.J.; Haugaasen, T.; Medina Uribe, C.A.; Foster, W.A.; Edwards, D.P. Reducing the impacts of Neotropical oil palm development on functional diversity. Biol. Conserv. 2016, 197, 139–145. [Google Scholar] [CrossRef]
- Knowlton, J.L.; Phifer, C.C.; Cerqueira, P.V.; Barro, F.D.C.; Oliveira, S.L.; Fiser, C.M.; Becker, N.M.; Cardoso, M.R.; Flaspohler, D.J.; Dantas Santos, M.P. Oil Palm Plantations Affect Movement Behavior of a Key Member of Mixed-Species Flocks of Forest Birds in Amazonia, Brazil. Trop. Conserv. Sci. 2017, 10, 194008291769280. [Google Scholar] [CrossRef]
- Molina, M.; Bohórquez, K. Bird Diversity: A Potential Indicator of Ecologic Sustainability in Agroecosystems at the South of Lake Maracaibo. Boletín Del Cent. Investig. Biológicas 2013, 47, 259–279. [Google Scholar]
- Cajas-Castillo, J.O.; Cóbar-Carranza, A.J.; Carlos, R.; Cruz, Á.-S.; Kraker-Castañeda, C.; Quiñónez-Guzmán, J.M. Diversidad de aves de sotobosque en bosques tropicales, áreas de regeneración natural y cultivos de palma africana en humedales del Lago de Izabal, Guatemala. Ornitol. Neotrop. 2015, 26, 1–12. [Google Scholar]
- Israel, G. Advances in Survey and Data Analysis Methods for Rural Social Scientists: An Introduction. J. Rural Soc. Sci. 2016, 31, 1. [Google Scholar]
- Quattrocchio, M.E.; Borromei, A.M.; Deschamps, C.M.; Grill, S.C.; Zavala, C.A. Landscape evolution and climate changes in the Late Pleistocene-Holocene, southern Pampa (Argentina): Evidence from palynology, mammals and sedimentology. Quat. Int. 2008, 181, 123–138. [Google Scholar] [CrossRef]
- INTA, Instituto Nacional de Tecnologı´a Agropecuaria. Estatsticas agroclimaticas mensuales eea Concordia INTA (Serie historica 1969–2007). 2015. Available online: http://inta.gob.ar/documentos/estadisticas-agroclimaticasde-concordia/at_multi_download/file/INTA_Concordia_Estadisticas_agroclimaticas.pdf (accessed on 8 August 2015).
- Brandão, F.; Schoneveld, G.C. The State of Oil Palm Development in the Brazilian Amazon: Trends, Value Chain Dynamics, and Business Models Governing Oil Palm Landscapes for Sustainability; Africa-China Informal Resource Trade (ACIRT): Bogor, Indonesia, 2015. [Google Scholar]
- Gascon, C.; Bierregaard, R.O., Jr.; Laurance, W.F.; Rankin-de-Merona, J. Deforestation and forest fragmentation in the Amazon. In Lessons from Amazonia: The Ecology and Conservation of a Fragmented Forest; Bierregaard, R.O., Jr., Gascon, C., Lovejoy, T.E., Mesquita, R., Eds.; Yale University Press: New Haven, CT, USA; London, UK, 2001; pp. 22–30. [Google Scholar]
- Haer, T.; Botzen, W.J.W.; Zavala-Hidalgo, J.; Cusell, C.; Ward, P.J. Economic evaluation of climate risk adaptation strategies: Cost-benefit analysis of flood protection in Tabasco, Mexico. Atmosfera 2017, 30, 101–120. [Google Scholar] [CrossRef]
- West, R.; Psuty, N. Las Tierras Bajas de Tabasco en el Sureste de México; No. 972.726 W4.; Gobierno del Estado de Tabasco: Villahermosa, Tabasco, Mexico, 1985.
- Van Der Wal, H.; Pea-Lvarez, B.; Arriaga-Weiss, S.L.; Hernndez-Daums, S. Species, functional groups, and habitat preferences of birds in five agroforestry classes in Tabasco, Mexico. Wilson J. Ornithol. 2012, 124, 558–571. [Google Scholar] [CrossRef]
- Moo Culebro, L.Y.; Knowlton, J.L.; Flaspohler, D.J.; Arriaga-Weiss, S.L.; Mata-Zayas, E.E. Avifauna associated with African palm plantations, secondary vegetation and surrounding matrix of the Sierra Tabasquena. Unpublished.
- NOAA, National Oceanic and Atmospheric Administration. Climate Data Online. National Centers for Environmental Information; NOAA: Ashville, NC, USA, 2016.
- Hutto, R.L.; Pletschet, S.M.; Hendricks, P. A Fixed-radius Point Count Method for Nonbreeding and Breeding Season Use. Auk 1986, 103, 593–602. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hill, D.A.; Hillis, D.M.; Mustoe, S. Bird Census Techniques, 2nd ed.; Academic Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 1988. [Google Scholar]
- Fernández-Llamazares, Á.; Díaz-Reviriego, I.; Guèze, M.; Cabeza, M.; Pyhälä, A.; Reyes-García, V. Local perceptions as a guide for the sustainable management of natural resources: Empirical evidence from a small-scale society in Bolivian Amazonia. Ecol. Soc. 2016, 21, 2. [Google Scholar] [CrossRef] [PubMed]
- Silva-Andrade, H.L.; de Andrade, L.P.; Muniz, L.S.; Telino-Júnior, W.R.; Albuquerque, U.P.; Lyra-Neves, R.M. Do Farmers Using Conventional and Non-Conventional Systems of Agriculture Have Different Perceptions of the Diversity of Wild Birds? Implications for Conservation. PLoS ONE 2016, 11, e0156307. [Google Scholar] [CrossRef]
- King, E.; Cavender-Bares, J.; Balvanera, P.; Mwampamba, T.H.; Polasky, S. Trade-offs in ecosystem services and varying stakeholder preferences: Evaluating conflicts, obstacles, and opportunities. Ecol. Soc. 2015, 20, 25. [Google Scholar] [CrossRef]
- Polasky, S.; Nelson, E.; Camm, J.; Csuti, B.; Fackler, P.; Lonsdorf, E.; Montgomery, C.; White, D.; Arthur, J.; Garber-Yonts, B.; et al. Where to put things? Spatial land management to sustain biodiversity and economic returns. Biol. Conserv. 2008, 141, 1505–1524. [Google Scholar] [CrossRef]
- Smith, F.P.; Gorddard, R.; House, A.P.N.; McIntyre, S.; Prober, S.M. Biodiversity and agriculture: Production frontiers as a framework for exploring trade-offs and evaluating policy. Environ. Sci. Policy 2012, 23, 85–94. [Google Scholar] [CrossRef]
- Ghazoul, J.; Hasanah, N.; Komarudin, H.; Dray, A. Beyond oil palm: Perceptions of local communities of environmental change ETH Library. Front. For. Glob. Chang. 2019, 2, 41. [Google Scholar]
- Azhar, B.; Lindenmayer, D.B.; Wood, J.; Fischer, J.; Manning, A.; McElhinny, C.; Zakaria, M. The conservation value of oil palm plantation estates, smallholdings and logged peat swamp forest for birds. For. Ecol. Manag. 2011, 262, 2306–2315. [Google Scholar] [CrossRef]
- Razak, S.A.; Saadun, N.; Azhar, B.; Lindenmayer, D.B. Smallholdings with high oil palm yield also support high bird species richness and diverse feeding guilds. Environ. Res. Lett. 2020, 15, 094031. [Google Scholar] [CrossRef]
- Kaye-Zwiebel, E.; King, E. Kenyan pastoralist societies in transition: Varying perceptions of the value of ecosystem services. Ecol. Soc. 2014, 19, 17. [Google Scholar] [CrossRef]
- Adams, W.M.; Brockington, D.; Dyson, J.; Vira, B. Managing Tragedies: Understanding Conflict over Common Pool Resources. Science 2003, 302, 1915–1916. [Google Scholar] [CrossRef]
- Santos, F.C.; Pacheco, J.M. Risk of collective failure provides an escape from the tragedy of the commons. Proc. Natl. Acad. Sci. USA 2011, 108, 10421–10425. [Google Scholar] [CrossRef] [PubMed]
- Baars, R.M.T.; Aptidon, S.M. Pastoralists’ perceptions of rangeland degradation in eastern Ethiopia. Nomadic Peoples 2002, 6, 144–157. [Google Scholar] [CrossRef]
- Baird, T.D.; Leslie, P.W.; McCabe, J.T. The effect of wildlife conservation on local perceptions of risk and behavioral response. Hum. Ecol. 2009, 37, 463–474. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).