
The Importance of Low-Intensive Agricultural Landscape for Birds of Prey


Abstract
1. Introduction
2. Materials and Methods
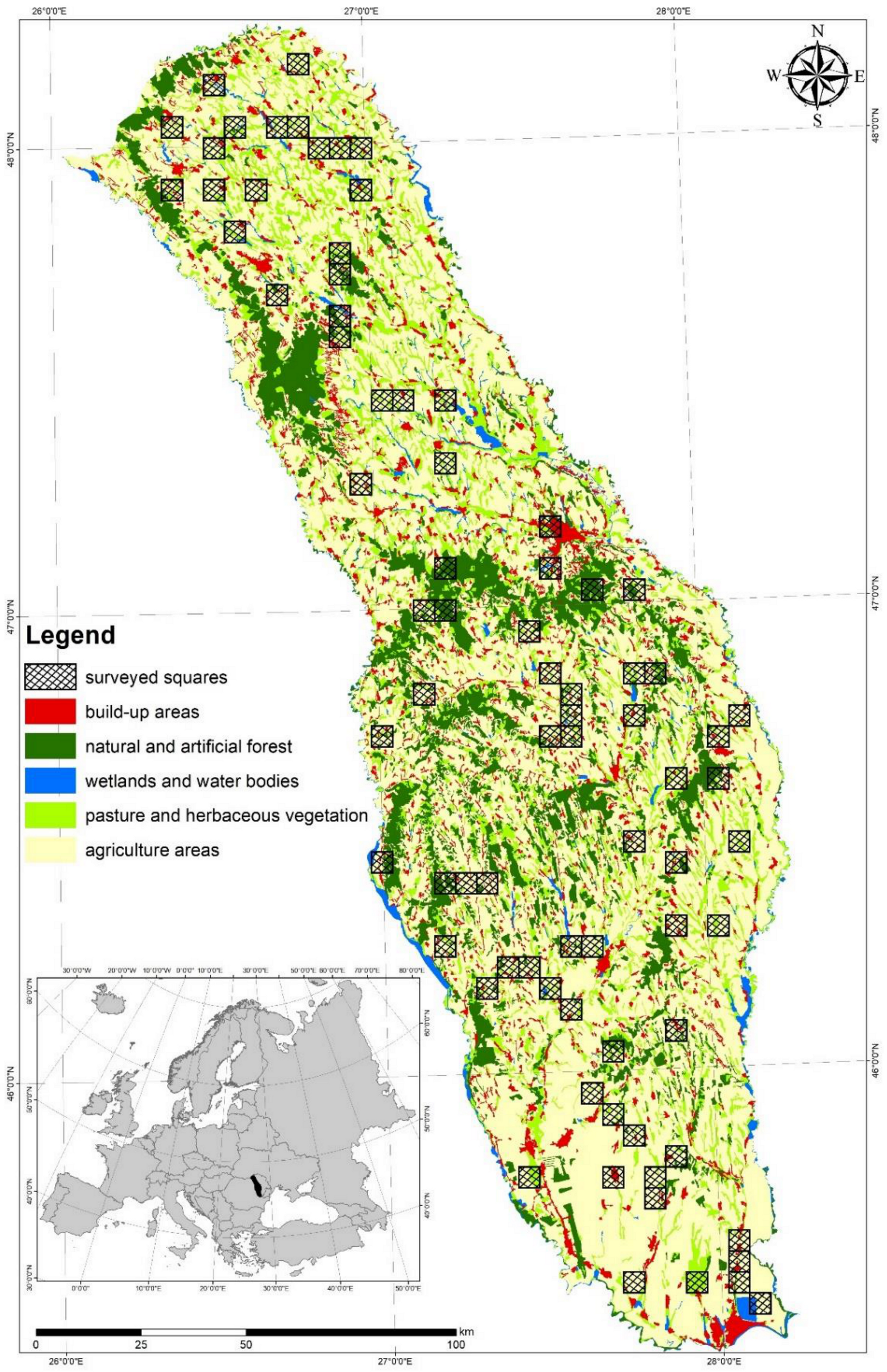
2.1. Study Area
2.2. Study Design and Buzzard Data Collection
2.3. Environmental and Climatic Predictor Variables
2.4. Data Analyses
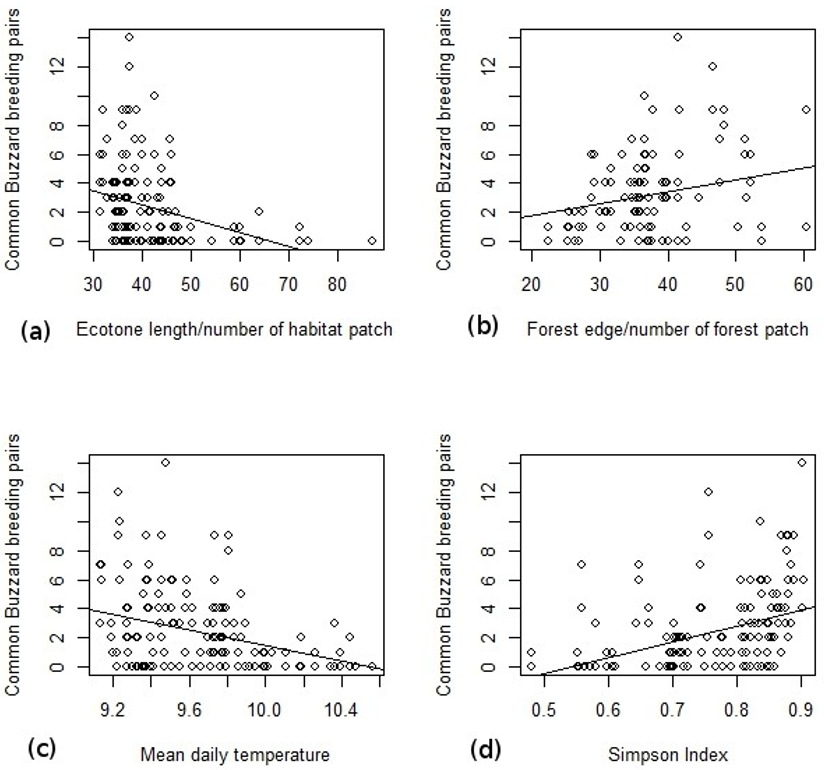
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| No. crt. | English Name | Scientific Name | Individuals/Breeding Season | |
|---|---|---|---|---|
| 2011 | 2012 | |||
| Breeding bird of prey species | ||||
| 1 | Northern Goshawk | Accipiter gentilis | 27 | 19 |
| 2 | Sparrowhawk | Accipiter nisus | 1 | 1 |
| 3 | Booted Eagle | Aquila pennata | 2 | 1 |
| 4 | Lesser Spotted Eagle | Aquila pomarine | 19 | 8 |
| 5 | Common Buzzard | Buteo buteo * | 207 | 171 |
| 6 | Long-legged Buzzard | Buteo rufinus * | 8 | 5 |
| 7 | Short-toed Eagle | Circaetus gallicus | 0 | 1 |
| 8 | Marsh Harrier | Circus aeruginosus | 29 | 31 |
| 9 | Hobby | Falco subbuteo | 5 | 6 |
| 10 | Kestrel | Falco tinnunculus | 80 | 38 |
| 11 | Red-footed Falcon | Falco vespertinus | 12 | 36 |
| 12 | Black Kite | Milvus migrans | 1 | 0 |
| 13 | Honey Buzzard | Pernis apivorus | 32 | 31 |
| Non-breeding bird of prey species | ||||
| 14 | Steppe Eagle | Aquila heliaca | 0 | 2 |
| 15 | Hen Harrier | Circus cyaneus | 4 | 0 |
| 16 | Montagu’s Harrier | Circus pygargus | 1 | 1 |
| 17 | Saker Falcon | Falco cherrug | 0 | 1 |
| Owl species | ||||
| 18 | Little Owl | Athene noctua | 1 | 0 |
| 19 | Tawny Owl | Strix aluco | 1 | 0 |
References
- Begon, M.; Harper, J.L.; Townsend, C.R. Ecology: Individuals, Populations and Communities; Blackwell Science Ltd.: Oxford, UK, 1996. [Google Scholar]
- Watling, J.I.; Nowakowski, A.J.; Donnelly, M.A.; Orrock, J.L. Meta-analysis reveals the importance of matrix composition for animals in fragmented habitat. Glob. Ecol. Biogeogr. 2010, 20, 209–217. [Google Scholar] [CrossRef]
- Ehrlich, P.R.; Pringle, R.M. Where does biodiversity go from here? A grim business-as-usual forecast and a hopeful portfolio of partial solutions. Proc. Natl. Acad. Sci. USA 2008, 105, 11579–11586. [Google Scholar] [CrossRef]
- Gilroy, J.J.; Anderson, G.Q.; Grice, P.V.; Vickery, J.A.; Bray, I.; Watts, P.N.; Sutherland, W.J. Could soil degradation contribute to farmland bird declines? Links between soil penetrability and the abundance of yellow wagtails Motacilla flava in arable fields. Biol. Conserv. 2008, 141, 3116–3126. [Google Scholar] [CrossRef]
- Billeter, R.; Liira, J.; Bailey, D.; Bugter, R.; Arens, P.; Augenstein, I.; Aviron, S.; Baudry, J.; Bukacek, R.; Burel, F.; et al. Indica-tors for biodiversity in agricultural landscapes: A pan-European study. J. Appl. Ecol. 2008, 45, 141–150. [Google Scholar] [CrossRef]
- Sergio, F.; Caro, T.; Brown, D.; Clucas, B.; Hunter, J.; Ketchum, J.; McHugh, K.; Hiraldo, F. Top Predators as Conservation Tools: Ecological Rationale, Assumptions, and Efficacy. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 1–19. [Google Scholar] [CrossRef]
- Sutcliffe, L.M.E.; Batáry, P.; Kormann, U.; Báldi, A.; Dicks, L.V.; Herzon, I.; Kleijn, D.; Tryjanowski, P.; Apostolova, I.; Arlettaz, R.; et al. Harnessing the biodiversity value of Central and Eastern European farmland. Divers. Distrib. 2015, 21, 722–730. [Google Scholar] [CrossRef]
- Larsson, T.B.; Pinborg, U.; Dominique, R. Biological Diversity, in: Europe’s Environment: The Third Assessment, Environmental Assessment Report; European Environment Agency: Copenhagen, Denmark, 2003; pp. 230–249. [Google Scholar]
- Kleijn, D.; Kohler, F.; Báldi, A.; Batáry, P.; Concepción, E.; Clough, Y.; Díaz, M.; Gabriel, D.; Holzschuh, A.; Knop, E.; et al. On the relationship between farmland biodiversity and land-use intensity in Europe. Proc. R. Soc. B Boil. Sci. 2008, 276, 903–909. [Google Scholar] [CrossRef]
- Loos, J.; Dorresteijn, I.; Hanspach, J.; Fust, P.; Rakosy, L.; Fischer, J. Low-Intensity Agricultural Landscapes in Transylvania Support High Butterfly Diversity: Implications for Conservation. PLoS ONE 2014, 9, e103256. [Google Scholar] [CrossRef] [PubMed]
- Zechmeister, H.G.; Schmitzberger, I.; Steurer, B.; Peterseil, J.; Wrbka, T. The influence of land-use practices and econom-ics on plant species richness in meadows. Biol. Conserv. 2003, 114, 165–177. [Google Scholar] [CrossRef]
- Karp, D.S.; Rominger, A.J.; Zook, J.; Ranganathan, J.; Ehrlich, P.R.; Daily, G.C. Intensive agriculture erodes b-diversity at large scale. Ecol. Lett. 2012, 15, 963–970. [Google Scholar] [CrossRef]
- Southern, H.N.; Cramp, S. Handbook of the Birds of Europe, the Middle East and North Africa; the Birds of the Western Palearctic. J. Anim. Ecol. 1978, 47, 1022. [Google Scholar] [CrossRef]
- Bijlsma, R.G. Buteo buteo Buzzard. In The EBCC Atlas of European Breeding Birds: Their Distribution and Abundance; Hagemeijer, W.J.M., Blair, M.J., Eds.; T. and A. D. Poyser: London, UK, 1997; pp. 160–161. [Google Scholar]
- Amar, A.; Arroyo, B.; Meek, E.; Redpath, S.; Riley, H. Influence of habitat on breeding performance of Hen Harriers Circus cyaneus in Orkney. Ibis 2007, 150, 400–404. [Google Scholar] [CrossRef]
- Gamauf, A.; Tebb, G.; Nemeth, E. Honey Buzzard Pernis apivorus nest-site selection in relation to habitat and the dis-tribution of Goshawks Accipiter gentilis. Ibis 2013, 155, 258–270. [Google Scholar] [CrossRef]
- Hardey, J.; Crick, H.; Wernham, C.; Riley, H.; Etheridge, B.; Thompson, D. Raptors a Field Guide for Surveys and Monitoring, 2nd ed.; The Stationery Office Limited: Edinburgh, UK, 2009. [Google Scholar]
- Steenhof, K.; Kochert, M.N.; Carpenter, L.B.; Lehman, R.N. Long-Term Prairie Falcon Population Changes in Relation to Prey Abundance, Weather, Land Uses, and Habitat Conditions. Condor 1999, 101, 28–41. [Google Scholar] [CrossRef][Green Version]
- Byholm, P.; Nikula, A.; Kentta, J.; Taivalmäki, J.P. Interactions between habitat heterogeneity and food affect reproductive output in a top predator. J. Anim. Ecol. 2007, 76, 392–401. [Google Scholar] [CrossRef]
- Poirazidis, K.; Goutner, V.; Tsachalidis, E.; Kati, V. Comparison of nest-site selection patterns of different sympatric raptor species as a tool for their conservation. Anim. Biodivers. Conserv. 2007, 30, 131–145. [Google Scholar]
- Pereira, J.M.C.; Itami, R.M. GIS-based habitat modeling using logistic multiple regression—A study of the Mt. Graham red squirrel. Photogramm. Eng. Remote Sens. 1991, 57, 1475–1486. [Google Scholar]
- Baltag, E.S. Ecologia Șorecarilor (Aves: Buteo) din Partea de est a Moldovei (România). Ph.D. Thesis, “Alexandru Ioan Cuza” University of Iasi, Iasi, Romania, 2013. [Google Scholar]
- Tobolka, M.; Zolnierowicz, K.M.; Reeve, N.F. The effect of extreme weather events on breeding parameters of the White StorkCiconia ciconia. Bird Study 2015, 62, 377–385. [Google Scholar] [CrossRef]
- Zaharia, G.; Petrencu, L.; Baltag, E. Site selection of European ground squirrels (Spermophilus citellus) in Eastern Ro-mania and how they are influenced by climate, relief, and vegetation. Turk. J. Zool. 2016, 40, 917–924. [Google Scholar] [CrossRef]
- National Institute of Statistics. Romanian Statistical Yearbook; INS: Bucharest, Romania, 2018; ISSN: 1220-3246. [Google Scholar]
- Baltag, E.Ș.; Bolboacă, L.E.; Ion, C. Long-legged Buzzard (Aves:Buteo) breeding population from Moldova Region. Eur. Sci. J. 2014, 2, 346–351. [Google Scholar]
- Baltag, E.Ș.; Pocora, V.; Ion, C.; Sfîcă, L. Winter presence of Long-legged Buzzard (Buteo rufinus) in Moldova (Romania). Trav. Mus. Natl. D’Histoire Nat. Grigore Antipa 2012, 55, 285–290. [Google Scholar] [CrossRef]
- Jung, M. LecoS—A python plugin for automated landscape ecology analysis. Ecol. Inform. 2016, 31, 18–21. [Google Scholar] [CrossRef]
- Jarvis, A.; Reuter, I.H.; Nelson, A.; Guevara, E. SRTM 90 m Digital Elevation Database v4.1|CGIAR-CSI. Available online: https://cgiarcsi.community/data/srtm-90m-digital-elevation-database-v4-1/ (accessed on 1 November 2020).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Clim. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Penteriani, V.; Faivre, B. Breeding Density and Landscape-Level Habitat Selection of Common Buzzards (Buteo buteo) in a Mountain Area (Abruzzo Apennines, Italy). J. Raptor Res. 1997, 31, 208–212. [Google Scholar]
- Nemcek, V. Abundance of raptors and habitat preferences of the common buzzard Buteo buteo and the common kestrel Falco tinnunculus during the non-breeding season in an agricultural landscape (Western Slovakia). Raptor J. 2014, 7, 37–42. [Google Scholar] [CrossRef]
- Sim, I.M.W.; Cross, A.V.; Lamacraft, D.L.; Pain, D.J. Correlates of Common Buzzard Buteo buteo density and breeding success in the West Midlands. Bird Study 2001, 48, 317–329. [Google Scholar] [CrossRef]
- Baltag, E.Ș.; Pocora, V.; Petrencu, L. Nest-site preferences of Common Buzzard, Buteo buteo (Linnaeus, 1758), from Eastern Romania. Acta Zool. Bulg. 2017, 69, 55–60. [Google Scholar]
- Dare, P. The Life of Buzzards; Whittles Publishing: Caithness, UK, 2015. [Google Scholar]
- Chakarov, N.; Krüger, O. Mesopredator Release by an Emergent Superpredator: A Natural Experiment of Predation in a Three Level Guild. PLoS ONE 2010, 5, e15229. [Google Scholar] [CrossRef] [PubMed]
- Krüger, O. Analysis of nest occupancy and nest reproduction in two sympatric raptors: Common buzzard Buteo buteo and goshawk Accipiter gentilis. Ecography 2002, 25, 523–532. [Google Scholar] [CrossRef]
- Mueller, A.; Chakarov, N.; Heseker, H.; Krüger, O. Intraguild predation leads to cascading effects on habitat choice, behaviour and reproductive performance. J. Anim. Ecol. 2016, 85, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Kenward, R.E.; Walls, S.S.; Hodder, K.H. Life path analysis: Scaling indicates priming effects of social and habitat fac-tors on dispersal distances. J. Anim. Ecol. 2001, 70, 1–13. [Google Scholar] [CrossRef][Green Version]
- Krüger, O. Dissecting common buzzard lifespan and lifetime reproductive success: The relative importance of food, competition, weather, habitat and individual attributes. Oecologia 2002, 133, 474–482. [Google Scholar] [CrossRef]
- Björklund, H.; Valkama, J.; Tomppo, E.; Laaksonen, T. Habitat Effects on the Breeding Performance of Three Forest-Dwelling Hawks. PLoS ONE 2015, 10, e0137877. [Google Scholar] [CrossRef]
- Hakkarainen, H.; Mykrä, S.; Kurki, S.; Tornberg, R.; Jungell, S.; Nikula, A. Long-term change in territory occupancy pat-tern of goshawks (Accipiter gentilis). Ecoscience 2004, 11, 399–403. [Google Scholar] [CrossRef]
- Butet, A.; Michel, N.; Rantier, Y.; Comor, V.; Hubert-Moy, L.; Nabucet, J.; Delettre, Y. Responses of common buzzard (Bu-teo buteo) and Eurasian kestrel (Falco tinnunculus) to land use changes in agricultural landscapes of Western France. Agric. Ecosyst. Environ. 2010, 138, 152–159. [Google Scholar] [CrossRef]
- Krüger, O.; Lindström, J. Habitat heterogeneity affects population growth in goshawk Accipiter gentilis. J. Anim. Ecol. 2001, 70, 173–181. [Google Scholar]
- Lehikoinen, A.; Byholm, P.; Ranta, E.; Saurola, P.L.; Valkama, J.; Korpimäki, E.; Pietiäinen, H.; Henttonen, H. Reproduction of the common buzzard at its northern range margin under climatic change. Oikos 2009, 118, 829–836. [Google Scholar] [CrossRef]
- Jankowiak, Ł.; Tryjanowski, P. Cooccurrence and food niche overlap of two common predators (red fox Vulpes vul-pes and common buzzard Buteo buteo) in an agricultural landscape. Turk. J. Zool. 2013, 37, 157–162. [Google Scholar]
- Good Agricultural and Environmental Conditions. GAEC 7—Retention of Landscape Features. 2021. Available online: http://www.madr.ro/docs/dezvoltare-rurala/legislatie/OMADR_999_ecoconditionalitate_2016.pdf (accessed on 3 January 2021).


| Variable Name | Variable Abbreviation | Variable Recording |
|---|---|---|
| Build-up areas | % in 5 × 5 square | |
| Arable land | Arable | % in 5 × 5 square |
| Vineyards | Vineyards | % in 5 × 5 square |
| Orchards | Orchards | % in 5 × 5 square |
| Pasture and herbaceous vegetation | Pasture | % in 5 × 5 square |
| Agriculture mixed with significant natural vegetation | Agro-environment | % in 5 × 5 square |
| Natural and artificial forest | Forest | % in 5 × 5 square |
| Wetlands and water bodies | Wetlands | % in 5 × 5 square |
| Total ecotone length | edge_t | sum in 5 × 5 square |
| Ecotone length/number of habitat patch | edge_d | - |
| Total forest edge | f_edge_t | sum in 5 × 5 square |
| Forest edge/number of forest patch | f_edge_d | - |
| Number of habitat patches | edge_c | sum in 5 × 5 square |
| Number of forest patches | f_edge_c | sum in 5 × 5 square |
| Shannon Index | DIV_SH | each surveyed square |
| Shannon Equitability | DIV_EV | each surveyed square |
| Simpson Index | DIV_SI | each surveyed square |
| Mean altitude | alt_mean | mean in 5 × 5 square |
| Altitude standard deviation | alt_std | in 5 × 5 square |
| Altitude variability | alt_var | in 5 × 5 square |
| Mean daily temperature | t_an | annual daily mean |
| Annual Precipitation | prec_an | annual total |
| Model | AIC | Log-Likelihood | Model Weight |
|---|---|---|---|
| alt_var + DIV_SI* + edge_d* + f_edge_d* + forest + t_an* | 537.8 | −255.854 | 0.32 |
| alt_var + DIV_SH + DIV_SI* + edge_d* + f_edge_d* + forest + t_an* | 538.3 | −254.920 | 0.25 |
| alt_var + DIV_SI* + edge_d* + f_edge_d* + t_an* | 539.3 | −257.744 | 0.16 |
| alt_var + DIV_SI* + edge_d* + f_edge_d* + forest + prec_an + t_an* | 539.4 | −255.430 | 0.15 |
| alt_var + DIV_SH + DIV_SI* + edge_d* + f_edge_d* + forest + prec_an + t_an* | 539.8 | −254.469 | 0.12 |
| Variable | Estimate | SE | Z Value | p |
|---|---|---|---|---|
| Intercept | 3.367 | 1.764 | 2.09 | 0.036 |
| Forest | 0.010 | 0.005 | 1.96 | 0.051 |
| edge_d | −0.0004 | 0.0001 | −2.54 | 0.011 |
| f_edge_d | 0.0003 | 0.0001 | 3.17 | 0.002 |
| alt_var | 0.002 | 0.001 | 1.62 | 0.105 |
| t_an | −0.048 | 0.015 | −3.13 | 0.002 |
| DIV_SI | 1.740 | 0.721 | 2.41 | 0.016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baltag, E.Ș.; Pocora, V.; Bolboaca, L.E.; Ion, C. The Importance of Low-Intensive Agricultural Landscape for Birds of Prey. Land 2021, 10, 252. https://doi.org/10.3390/land10030252
Baltag EȘ, Pocora V, Bolboaca LE, Ion C. The Importance of Low-Intensive Agricultural Landscape for Birds of Prey. Land. 2021; 10(3):252. https://doi.org/10.3390/land10030252
Chicago/Turabian StyleBaltag, Emanuel Ștefan, Viorel Pocora, Lucian Eugen Bolboaca, and Constantin Ion. 2021. "The Importance of Low-Intensive Agricultural Landscape for Birds of Prey" Land 10, no. 3: 252. https://doi.org/10.3390/land10030252
APA StyleBaltag, E. Ș., Pocora, V., Bolboaca, L. E., & Ion, C. (2021). The Importance of Low-Intensive Agricultural Landscape for Birds of Prey. Land, 10(3), 252. https://doi.org/10.3390/land10030252