
Effects of the Invasive Tree Species Ailanthus altissima on the Floral Diversity and Soil Properties in the Pannonian Region

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
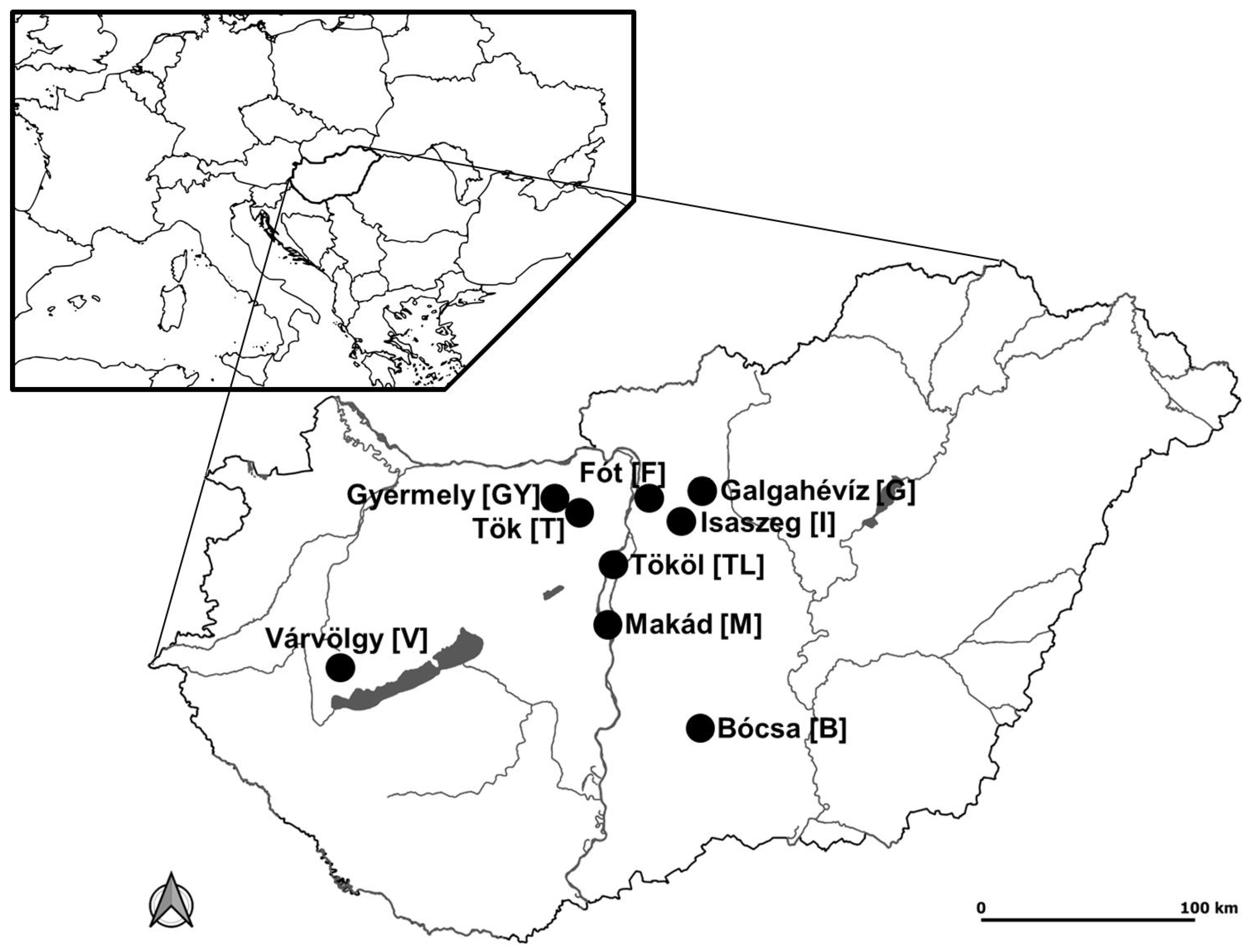
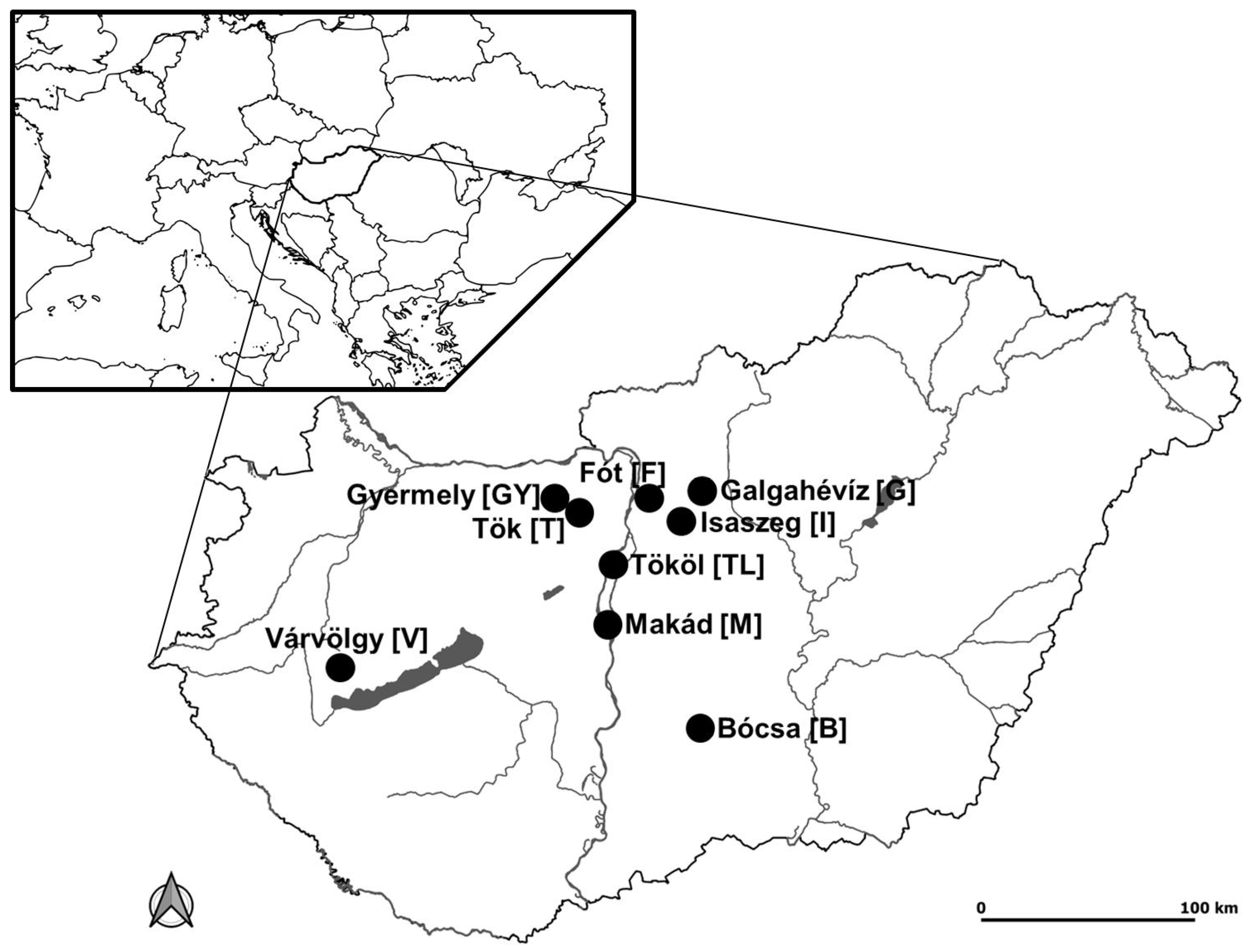
2.1. Study Sites
2.2. Botanical Sampling
2.3. Measurements of Biotic and Abiotic (Soil and Other Environmental Parameters) Data
2.4. Statistical Analysis
3. Results
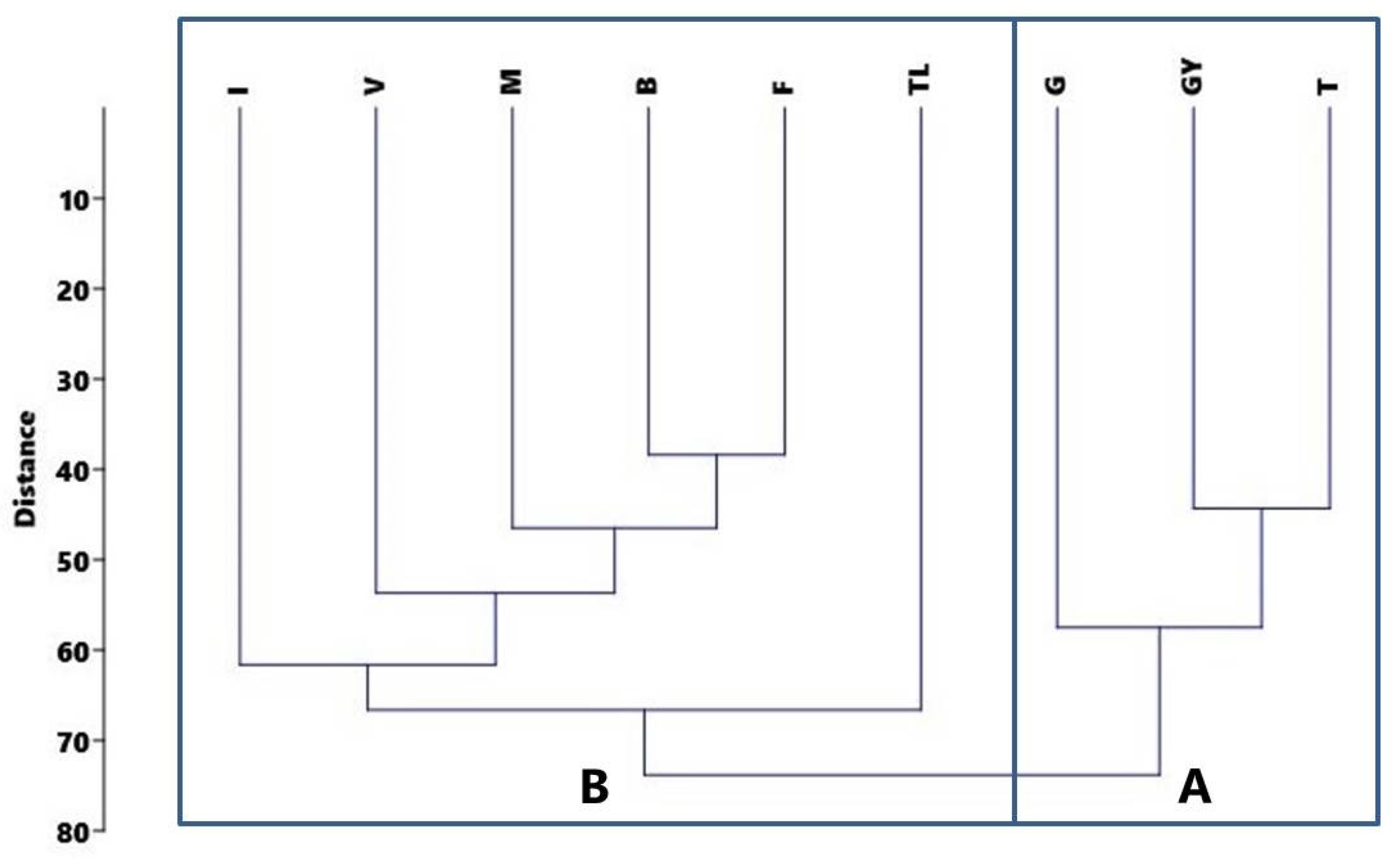
3.1. Composition of Vegetation and Similarity of Study Sites
3.2. Mass of A. altissima at Different Layers of the Studied Forest Sites
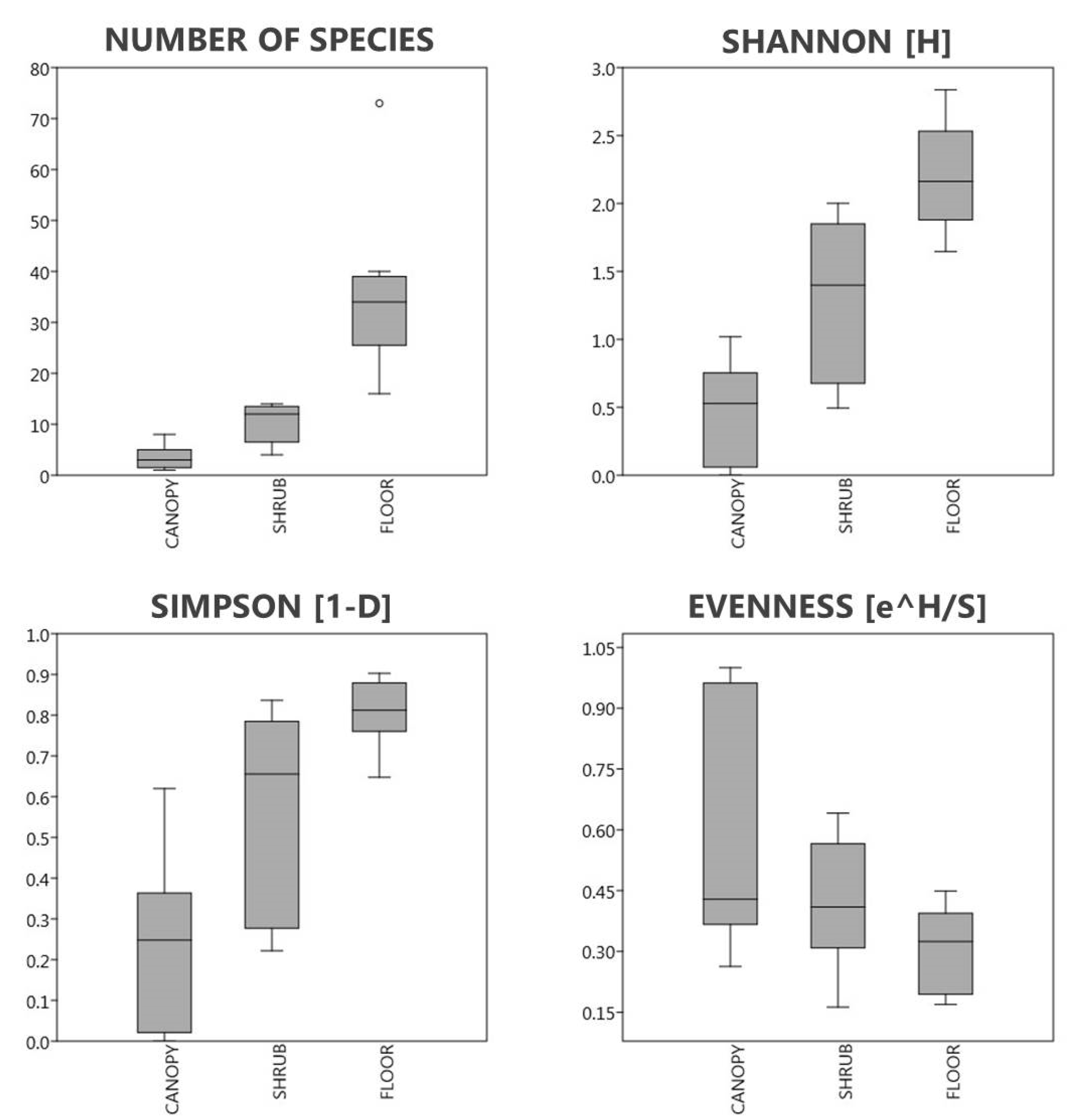
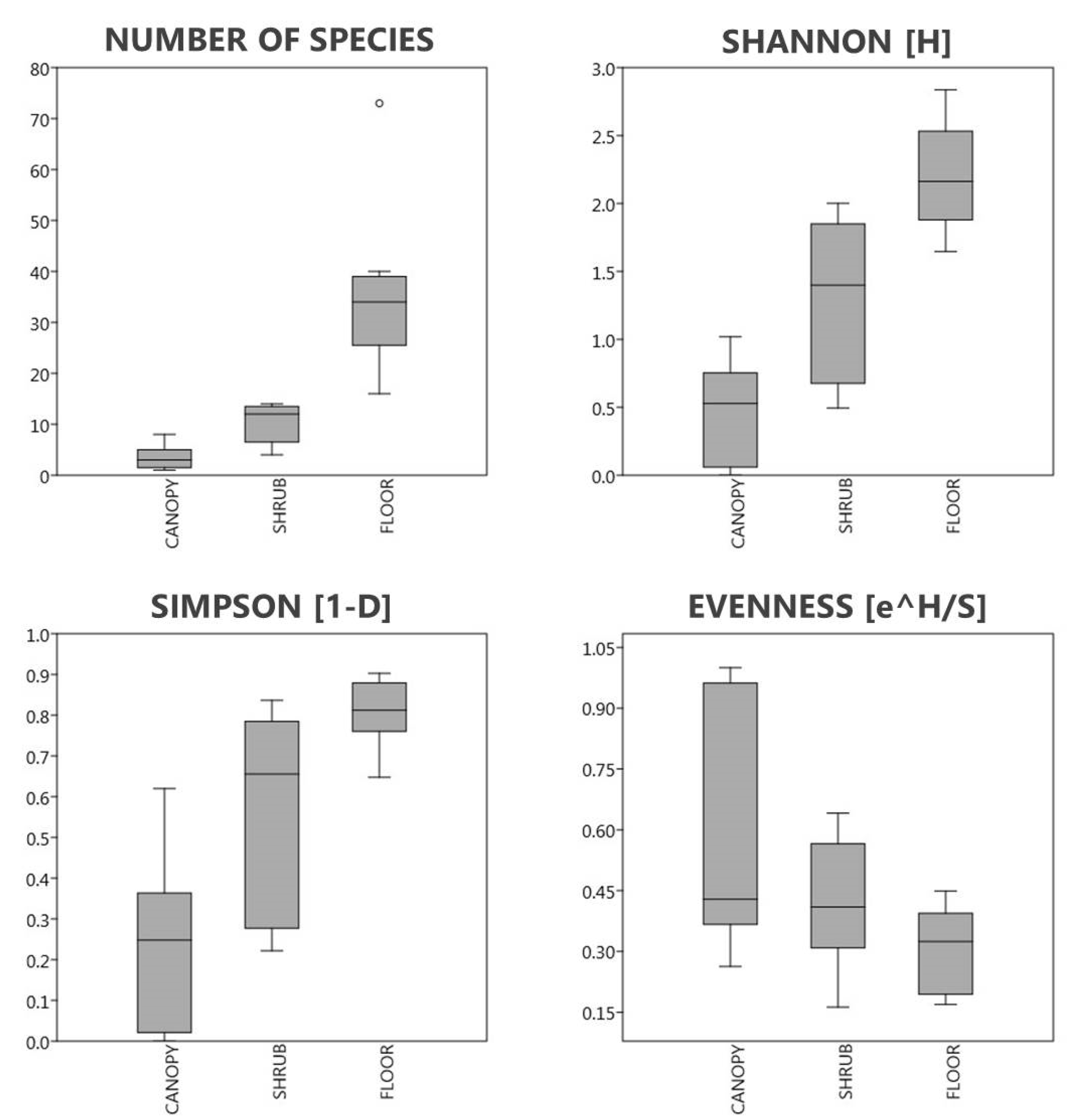
3.3. The Diversity of Each Layer Depending on the Mass of A. altissima
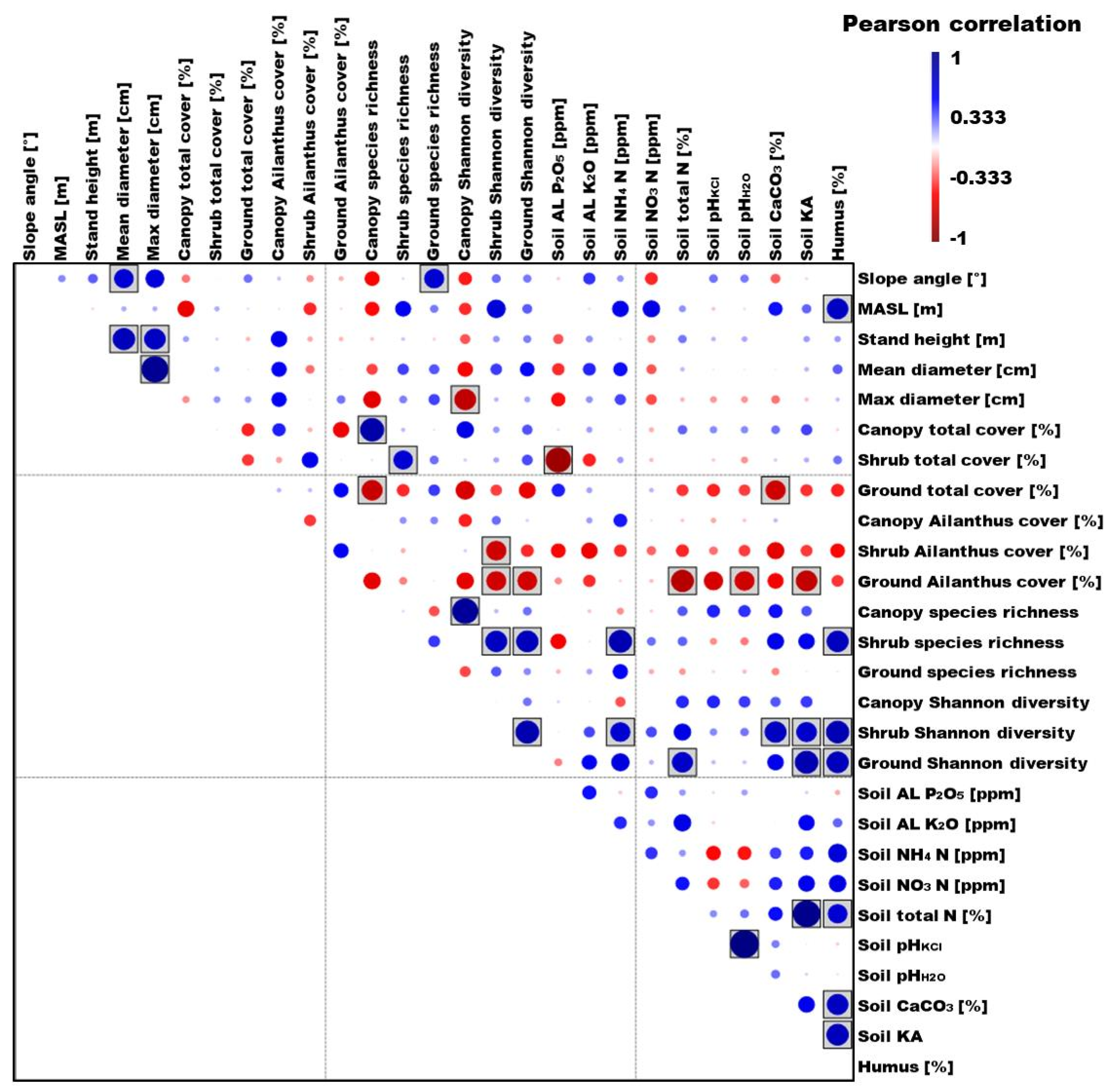
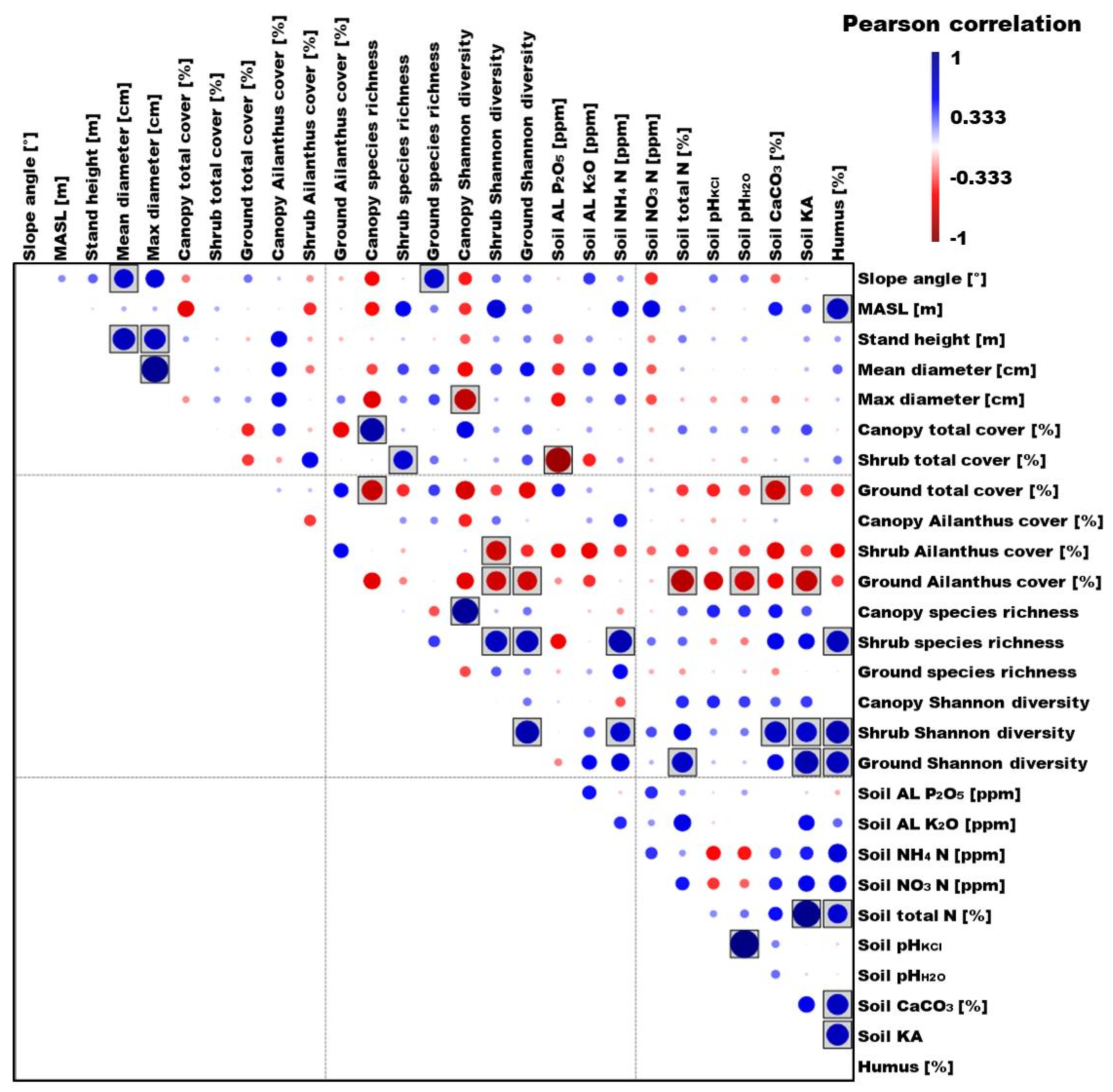
3.4. Correlation between the Examined Biotic and Abiotic Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kowarik, I.; Säumel, I. Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Perspect. Plant Ecol. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošík, V.; Pergl, J.; Winter, M.; Anastasiu, P.; et al. Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. Available online: http://www.preslia.cz/P082Lam.pdf (accessed on 24 August 2021).
- Sladonja, B.; Sušek, M.; Guillermic, J. Review on Invasive Tree of Heaven (Ailanthus altissima (Mill.) Swingle) Conflicting Values: Assessment of Its Ecosystem Services and Potential Biological Threat. Environ. Manag. 2015, 56, 1009–1034. [Google Scholar] [CrossRef] [PubMed]
- List of Invasive Alien Species of Union Concern. Available online: https://ec.europa.eu/environment/nature/invasivealien/list/index_en.htm (accessed on 9 October 2021).
- Kowarik, I.; Böcker, R. Zur Verbreitung, Vergesellschaftung und Einbürgerung des Götterbaumes (Ailanthus altissima (Mill.) SWINGLE) in Mitteleuropa. Tuexenia 1984, 4, 9–29. [Google Scholar]
- Heisey, R.M. Allelopathic and Herbicidal Effects of Extracts from Tree of Heaven (Ailanthus altissima). Am. J. Bot. 1990, 77, 662–670. [Google Scholar] [CrossRef]
- Heisey, R.M. Evidence for allelopathy by tree-of-heaven (Ailanthus altissima). J. Chem. Ecol. 1990, 16, 2039–2055. [Google Scholar] [CrossRef]
- Gomez-Aparicio, L.; Canham, C.D. Neighbourhood analyses of the allelopathic effects of the invasive tree Ailanthus altissima in temperate forests. J. Ecol. 2008, 96, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Csiszár, Á. Allelopathic effects of invasive woody plant species is Hungary. Acta Silv. Lign. Hung. 2009, 5, 9–17. [Google Scholar]
- Csiszár, Á.; Korda, M.; Schmidt, D.; Šporčić, D.; Süle, P.; Teleki, B.; Tiborcz, V.; Zagyvai, G.; Bartha, D. Allelopathic potential of some invasive neophytes occurring in Hungary. Allelopath. J. 2013, 31, 309–318. [Google Scholar]
- Demeter, A.; Falvai, D.; Trenyik, P.; Czóbel, S. Ecological indicator based comparative study of tree of heaven (Ailanthus altissima) stands’ herb layer. Columella J. Agric. Environ. Sci. 2017, 4, 15–20. [Google Scholar] [CrossRef]
- Borhidi, A. Social behaviour types, the naturalness and relative indicator values of the higher plants in the Hungarian Flora. Acta Bot. Hung. 1995, 39, 97–182. [Google Scholar]
- Motard, E.; Muratet, A.; Clair-Maczulajtys, D.; Machon, N. Does Invasive Species Ailanthus altissima Threaten Floristic Diversity of Temperate Peri-Urban Forests? Comptes Rendus Biol. 2011, 334, 872–879. [Google Scholar] [CrossRef]
- Vilà, M.; Tessier, M.; Suehs, C.M.; Brundu, G.; Carta, L.; Galanidis, A.; Lambdon, P.; Manca, M.; Médail, F.; Moragues, E.; et al. Local and regional assessments of the impacts of plant invaders on vegetation structure and soil properties of Mediterranean islands. J. Biogeogr. 2006, 33, 853–861. [Google Scholar] [CrossRef]
- Traveset, A.; Brundu, G.; Carta, L.; Mprezetou, I.; Lambdon, P.; Manca, M.; Médail, F.; Moragues, E.; Rodríguez-P’erez, J.; Siamantziouras, A.-S.D. Consistent performance of invasive plant species within and among islands of the Mediterranean basin. Biol. Invasions 2008, 10, 847–858. [Google Scholar] [CrossRef]
- Constán-Nava, S.; Soliveres, S.; Torices, R.; Serra, L.; Bonet, A. Direct and indirect effects of invasion by the alien tree Ailanthus altissima on riparian plant communities and ecosystem multifunctionality. Biol. Invasions 2015, 17, 1095–1108. [Google Scholar] [CrossRef]
- Mount, H. Impacts of Invasive Ailanthus altissima on Woody Plant Communities in an Old Growth Forest of Southeastern Kentucky. NSF-Research Experiences for Undergraduates, Disturbance Ecology in Central Appalachia 2019. p. 2. Available online: https://www.eku.edu/ (accessed on 24 August 2021).
- Brooks, R.K.; Barney, J.N.; Salom, S.M. The invasive tree, Ailanthus altissima, impacts understory nativity, not seedbank nativity. For. Ecol. Manag. 2021, 489, 119025. [Google Scholar] [CrossRef]
- Gioria, M.; Jarošík, V.; Pyšek, P. Impact of invasions by alien plants on soil seed bank communities: Emerging patterns. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 132–142. [Google Scholar] [CrossRef]
- Strayer, D.L.; Eviner, V.T.; Jeschke, J.M.; Pace, M.L. Understanding the long-term effects of species invasions. Trends Ecol. Evol. 2006, 21, 645–651. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Fierro-Brunnenmeister, N.; González-Muñoz, N.; Gallardo, A. Effects of exotic and native tree leaf litter on soil properties of two contrasting sites in the Iberian Peninsula. Plant Soil 2012, 350, 179–191. [Google Scholar] [CrossRef]
- Montecchiari, S.; Allegrezza, M.; Peliccia, V.; Tesei, G. First syntaxonomical contribution to the invasive Ailanthus altissima (Mill.) Swingle forest communities at its southern limit in Europe. Plant Sociol. 2020, 57, 145–160. [Google Scholar] [CrossRef]
- Montecchiari, S.; Tesei, G.; Allegrezza, M. Ailanthus altissima Forests Determine a Shift in Herbaceous Layer Richness: A Paired Comparison with Hardwood Native Forests in Sub-Mediterranean Europe. Plants 2020, 9, 1404. [Google Scholar] [CrossRef] [PubMed]
- Motard, E.; Dusz, S.; Geslin, B.; Akpa-Vinceslas, M.; Hignard, C.; Babiar, O.; Clair-Maczulajtys, D.; Michel-Salzat, A. How invasion by Ailanthus altissima transforms soil and litter communities in a temperate forest ecosystem. Biol. Invasions 2015, 17, 1817–1832. [Google Scholar] [CrossRef]
- Gyalog, L.; Síkhegyi, F. (Eds.) Surface Geological Maps of Hungary; Scale = 1:100.000; Hungarian Geological Institute: Budapest, Hungary, 2015. [Google Scholar]
- Bihari, Z.; Babolcsai, G.; Bartholy, J.; Ferenczi, Z.; Gerhátné Kerényi, J.; Haszpra, L.; Homokiné Ujváry, K.; Kovács, T.; Lakatos, M.; Németh, Á.; et al. Climate—Precipitation. In National Atlas of Hungary; Kocsis, K., Ed.; MTA CSFK Geographical Institute: Budapest, Hungary, 2018; Volume 2—Natural Environment, pp. 62–63. [Google Scholar]
- Bihari, Z.; Babolcsai, G.; Bartholy, J.; Ferenczi, Z.; Gerhátné Kerényi, J.; Haszpra, L.; Homokiné Ujváry, K.; Kovács, T.; Lakatos, M.; Németh, Á.; et al. Climate—Temperature. In National Atlas of Hungary; Kocsis, K., Ed.; MTA CSFK Geographical Institute: Budapest, Hungary, 2018; Volume 2—Natural Environment, pp. 60–61. [Google Scholar]
- Buzás, I. (Ed.) Soil and agrochemical test method book 2. In Physico-Chemical and Chemical Test Methods for Soils; Mezőgazdasági Kvk: Budapest, Hungary, 1988; p. 243. (In Hungarian) [Google Scholar]
- Egner, J.; Riehm, H.; Domingo, W. Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährst-offzustandes der Böden II. Chemische Extraktionsmethoden zur Phosphor-und Kaliumbestimmung. K. Lantbrukshögskolans Ann. 1960, 26, 199–215. (In German) [Google Scholar]
- MSZ-08-0210-1977; Testing Organic Carbon Content in Soils; Hungarian Standard; Hungarian Standards Institution: Budapest, Hungary, 1977; p. 6. (In Hungarian)
- Hammer, Ø. PAST—PAleontological STatictics Version 3.21 Reference Manual; Natural History Museum, University of Oslo: Oslo, Norway, 1999–2018; p. 225. [Google Scholar]
- Hammer, Ø. PAST—PAleontological STatictics Version 4.05 Reference Manual; Natural History Museum, University of Oslo: Oslo, Norway, 1999–2021; p. 284. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST—Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Harper, D.A.T. (Ed.) Numerical Palaeobiology; John Wiley & Sons: New York, NY, USA, 1999; p. 468. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Average Annual Rainfall [mm] | Average Annual Temperature [°C] | Soil Type | Average Altitude MASL [m] | Vegetation Belt |
|---|---|---|---|---|---|
| B | 500–550 | 10–11 | quicksand | 115 | lowlands |
| F | 500–550 | 9–10 | sand | 186 | lowlands |
| G | 500–550 | 9–10 | loess | 160 | lowlands |
| GY | 550–600 | 9–10 | loess | 183 | lowlands |
| I | 500–550 | 9–10 | sand | 210 | hills |
| M | 500–550 | 10–11 | river sediment | 96 | lowlands |
| T | 550–600 | 10–11 | loess | 300 | hills |
| TL | 500–550 | 10–11 | sand | 100 | lowlands |
| V | 700–750 | 10–11 | proluvial sediment | 178 | lowlands |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demeter, A.; Saláta, D.; Tormáné Kovács, E.; Szirmai, O.; Trenyik, P.; Meinhardt, S.; Rusvai, K.; Verbényiné Neumann, K.; Schermann, B.; Szegleti, Z.; et al. Effects of the Invasive Tree Species Ailanthus altissima on the Floral Diversity and Soil Properties in the Pannonian Region. Land 2021, 10, 1155. https://doi.org/10.3390/land10111155
Demeter A, Saláta D, Tormáné Kovács E, Szirmai O, Trenyik P, Meinhardt S, Rusvai K, Verbényiné Neumann K, Schermann B, Szegleti Z, et al. Effects of the Invasive Tree Species Ailanthus altissima on the Floral Diversity and Soil Properties in the Pannonian Region. Land. 2021; 10(11):1155. https://doi.org/10.3390/land10111155
Chicago/Turabian StyleDemeter, András, Dénes Saláta, Eszter Tormáné Kovács, Orsolya Szirmai, Petra Trenyik, Sarolta Meinhardt, Katalin Rusvai, Krisztina Verbényiné Neumann, Balázs Schermann, Zsófia Szegleti, and et al. 2021. "Effects of the Invasive Tree Species Ailanthus altissima on the Floral Diversity and Soil Properties in the Pannonian Region" Land 10, no. 11: 1155. https://doi.org/10.3390/land10111155
APA StyleDemeter, A., Saláta, D., Tormáné Kovács, E., Szirmai, O., Trenyik, P., Meinhardt, S., Rusvai, K., Verbényiné Neumann, K., Schermann, B., Szegleti, Z., & Czóbel, S. (2021). Effects of the Invasive Tree Species Ailanthus altissima on the Floral Diversity and Soil Properties in the Pannonian Region. Land, 10(11), 1155. https://doi.org/10.3390/land10111155