
Could Land Abandonment with Human Intervention Benefit Cropland Restoration? From the Perspective of Soil Microbiota


1
Institute of Land and Urban-Rural Development, Zhejiang University of Finance and Economics, Hangzhou 310058, China
2
School of Public Affairs, Zhejiang University, Hangzhou 310058, China
*
Author to whom correspondence should be addressed.
Land 2021, 10(10), 1049; https://doi.org/10.3390/land10101049
Submission received: 12 August 2021
/
Revised: 3 October 2021
/
Accepted: 3 October 2021
/
Published: 6 October 2021
(This article belongs to the Special Issue Land Abandonment: Positive and Negative Effects on Soil Quality, Ecosystem Services, and Environmental Functioning)
Abstract
:Although agricultural land abandonment (LA) is accompanied by land degradation, it could be considered a kind of self-rehabilitation. Studies have shown that long-term LA has profound ecological and environmental benefits, whereas few studies have compared LA with human intervention (HI), which involves planting and fertilization in agroecosystem restoration. Here, we established four different scenarios based on local livestock husbandry, including LA without HI, LA with slight human intervention (HIS), medium human intervention (HIM), and intensive human intervention (HII). LA experiments were conducted for 3 years and repeatedly sampled three times. The soil bacterial and fungal communities were determined to present the ecological impacts. In this study, LA and HIS could save soil inorganic carbon and total calcium (Ca) contents and benefit soil mycorrhizal fungi and plant growth-promoting rhizobacteria. LA and HIM benefited some microbial communities associated with complicated organic compounds. Human interference methods did not significantly increase soil nutrients after 3 years of farmland abandonment. However, indigenous vegetation increased the risk of plant diseases based on soil microbial communities. Forage grass may control the risk, and HIS was a cost-effective scenario in our study. Moreover, we should maintain a cautious attitude toward HII to prevent excessive intervention.
1. Introduction
Agricultural land abandonment (LA) is emerging because of rapid urbanization and a decrease in the agricultural labor force. An increasing number of young people choose to go to cities for off-farm work, and the loss of agricultural labor is driving cropland abandonment [1]. In addition, the increase in grain storage capacity caused the grain supply to exceed the demand, which was followed by a decline in food prices [2]. Insufficient income from crops exacerbated farmers leaving farm work and led to the abandonment of farmland. Some agricultural policies also seemed to cause the emergence of abandonment. The European Common Agricultural Policy (CAP), modified in the 1980s, led to agricultural overcapacity and then subsidized cropland fallowing. It enhanced the LA phenomenon. Recently, the amendment of subsidy systems, which has taken agricultural and environmental conditions into account, has changed the situation [3]. In some cases, LA was always considered as a negative status for farmland. It is more likely to be present in low-quality areas, such as alkaline land, arid areas, or mountainous areas. High-cost management and poor environmental conditions have not only caused farmland to be abandoned but also led to farmland degradation, such as soil erosion and salinization, which increases the cost of reclamation. The natural recovery of vegetation and soil may be too slow to change the poor environment, which requires decades to centuries to approach natural states [4]. Based on a 10-year field experiment on the Loess Plateau of China, continuous natural recovery increased soil organic carbon, microbial biomass carbon, available potassium, and Steroidobacter (a bacterial genus) [5].
Therefore, some human interventions (HIs) have been applied to shorten ecosystem recovery times, such as revegetation and soil biochar application. However, the restoration method cannot ignore regional industrial development [6]. The selection of HI should follow ecological principles and the local socioeconomic status quo. For example, livestock production systems could be applied in the natural restoration of farmlands [7]. Based on the properties of livestock production systems, there are two ways to improve the effects of fallows. First, using goal-oriented forage grass promotes the benefits of restoration, especially the use of nitrogen-fixing pastures. Rhizosphere microbes showed that colonizing annual plants accelerated the process of soil fertility improvement in a short natural restoration time (1 to 5 years) that potentially boosted vegetation succession [8]. In addition, mixing nitrogen-fixing pastures with other local plants may produce synergistic effects [9]. Plant diversity has been shown to increase soil fertility [10]. Consequently, two or more plant species were applied to enhance the efficiency of natural restoration.
Second, livestock manure input can promote soil nutrient content, especially soil organic carbon. As tillage can cause the loss of significant amounts of carbon, belowground carbon cannot compensate for the negative impact of such losses [11]. Although farmyard manure (N: P: K = 32: 18: 38) and organic manures could increase the soil organic matter content [11,12], little is known about the interaction effects of vegetation and organic manures on microbial composition during the natural restoration period. In addition to the construction of vegetation, the addition of amendments to revegetation systems has been considered in some natural restoration studies. For instance, nitrogen-fixing plants and P+K fertilizer enhanced the nitrogen stock during a natural restoration period [13]. Natural fallow with farmyard manure significantly enhanced microbial biomass nitrogen over a short duration [14]. In this paper, we attempted to determine the effects of human intervention (livestock) on abandoned farmland from the perspective of soil physicochemical and microbial properties.
Although soil microbial indicators are widely used in agricultural research, they are still inadequately applied in LA studies. Soil microbial composition is closely related to the carbon–nitrogen cycle, soil-borne disease resistance, and farmland productivity [15]. The study of soil microbial properties showed that microbial biomass and some microbial communities were significantly affected by human intervention during the natural recovery time (18 months) [16]. Planted fallow farmland with phosphorus applications promoted arbuscular mycorrhizal fungi (AMF) and nutrient uptake [17]. In a long-term agroforestry study, abandoned land that was replanted showed insignificant positive effects on soil microbial properties [18]. However, these studies focused more on the comparison of different types of land use/cover than on ecological restoration studies, such as farmland fallow and crop rotation. Moreover, the resolution of the microbial community structure was indistinct in these studies because the characteristics of the microbial community structure were not analyzed at the genus level. In some short ecological restoration (one year) studies, a molecular biology approach was used to analyze the soil microbial community structure where different effects of human intervention on soil bacterial genera were established [14]. Nevertheless, soil microbial properties may be unstable because of the short restoration duration. It is necessary to extend the ecological restoration duration to determine the effects of human intervention on soil microbial properties, especially on soil microbial communities.
To determine the effects of HI on LA via nutrient cycling and microbial community structure, we analyzed the soil data for 3 untilled years. The bacterial and fungal compositions were determined in this study. We integrated soil physicochemical characteristics and microbial community structure to build three hypotheses:
Hypothesis H1:
The natural restoration of abandoned land may take longer, and it is more beneficial to nitrogen-fixing bacteria than human intervention;
Hypothesis H2:
Planting forage grass or inputting farmyard manure on abandoned land can improve soil fertility and benefit some fungi involved in plant growth;
Hypothesis H3:
Intensive human intervention on abandoned land may not achieve the expected results and may have negative effects.
2. Materials and Methods
2.1. Study Site Description
The research area is located at the Field Scientific Observation and Research Station for Yellow River Delta Land Use Safety in Binzhou, China [E 117°43′, N 37°48′, at an altitude of 5 m above sea level (a.s.l) ] (Figure S1). The 10-year average rainfall was 55.3 mm and the average temperature was 13.9 °C. Having a silty loam texture (approximately 3% clay and 78% silt), the study soil stemmed from diluvial sediments and is classified as a typical saline alluvial soil (Fluvisols, FAO). Based on our investigation, sorghum [Sorghum bicolor (L.) Moench] monoculture was practiced for over 5 years in the study area, and soil fertility has decreased markedly in the study area based on local soil investigation [19]. Before the experiment, the farmlands were compacted by a four-wheeled tractor which is a common phenomenon after harvest. Therefore, the study areas were flat and homogeneous. From 2010 to 2015, the same land management scheme was applied to the experimental plots for 5 years. The manager was planning to abandon the farmland to restore soil fertility, and we rented part of the farmland (approximately 2.6 ha) to carry out our experiment.
2.2. Experimental Design
The experiment commenced in 2016. We have investigated the natural and socioeconomic conditions around the study area. The chemical properties of the topsoil (0–20 cm layer) were as follows: electrical conductivity of 1.8 μS m−1, total nitrogen (TN) content of 0.62 g kg−1, soil organic carbon content (SOC) of 8.39 g kg−1, available potassium (AK) content of 0.11 g kg−1, available nitrogen content of 24.59 mg kg−1, available phosphorus (AP) content of 8.51 mg kg−1, and pH of 8.43. In terms of socioeconomic conditions, the local livestock breeding industry is relatively developed. We found that there was a cattle farm within 1 km of the study area. Therefore, our experiment is designed based on the characteristics of the local cattle industry. Four scenarios established, and three plots were set for each scenario. The size of each plot was 5 × 6 m, and the plots were randomly distributed (more than 1 m intervals each).
We investigated the effects of abandoned farmland (LA), which is the natural regrowth of spontaneous vegetation without human intervention. Taking into account the forage demand of the cattle farm, we can choose local forage planting as a method of human intervention in LA. Alfalfa (Medicago sativa L.) and Dahurian wild rye (Elymus dahuricus Turcz.), which belonged to the indigenous meadow, were selected to create plant mixtures for building pasture meadows without cattle manure. It was considered a scenario of slight human intervention in LA (HIS). Cattle manure is an inevitable output of cattle farms. We believe that its impact on farmland is greater than that of vegetation. Therefore, we fertilized cattle manure in LA, which was considered a scenario of medium human intervention in LA (HIM). Fertilization is an activity that is repeated every year. It could be considered a stronger human intervention than grass planting. Finally, we applied both forage planting and manure fertilization to abandoned farmland to build intensive human intervention in LA (HII). Manual weeding was applied at the beginning of the experiment to ensure the survival of the forages. To avoid the impact of tillage, hand weeding was used. The cattle manure was decomposed before. The cattle manure was mixed with water to prevent hardening. Fertilization was then accompanied by irrigation. No pesticide and little weeding were used in our study region. Moreover, we reaped forage grass in each harvest season (every 3 months).
Management scenarios were summarized and detailed as follows:
- LA, spontaneous vegetation, no manure input, spontaneous vegetation, as agricultural land abandonment;
- HIS, no manure input, spontaneous vegetation sown with alfalfa (Medicago sativa L.) and Dahurian wild rye (Elymus dahuricus Turcz.) in LA, as slight human intervention in LA;
- HIM, spontaneous vegetation, LA with composted cattle manure (400 g kg−1 organic C, 7.0 g kg−1 TN, 11.5 g kg−1 total P, and 9.8 g kg−1 total K), 1500 kg ha−1, as medium human intervention in LA;
- HII, HIS with composted cattle manure (same as HIM), 1500 kg ha−1, as intensive human intervention in LA.
2.3. Preparation of Soil Samples
The first soil sampling occurred in October 2016. The second and third samplings were performed in 2017 and 2018, respectively. Soil samples were collected from six points to a depth of 0–20 cm in each plot; a foil sampler was employed. The approximate amount of soil (500 g) collected per plot. The samples were then mixed and homogenized. Each soil sample was passed through a <2 mm sieve to remove plant roots and stones and was then divided into three different parts. The first part was stored at −40 °C for DNA and enzyme analysis. The remainder was air-dried to assay the soil physicochemical properties.
2.4. Edaphic Properties Analysis
The soil pH was measured by a glass electrode (soil: water = 1:2.5). The soil total organic carbon (SOC) and soil inorganic carbon (SIC) were determined with a CN analyzer (SOC-L Analyzer and SSM-5000A unit, SHIMADZU, Japan). The total nitrogen (TN) was assayed by the Kjeldahl method (Bremner et al. 1996). The carbon to nitrogen ratio (C/N) was based on the SOC and TN. Soil ammonia concentrations (NH4N) and nitrate–nitrogen (NO3N) in each sample were analyzed via a continuous flow analyzer (SKALAR San Plus automated wet chemistry analyzer, Breda, The Netherlands). The β-Glucosidase (Beta) activity was expressed as p-nitrophenol (PNP) [20]. Urease (Ur) activity was measured using a modified method [21]. A UV-Vis spectrophotometer (PGENERAL, Beijing, China) and fluorescence method were employed for enzyme analysis. Soil total calcium (Ca), total sodium (Na), and total heavy metal (Pb/Cu/Zn) contents were analyzed with an atomic absorption spectrometer (Thermo Fisher Scientific M iCE 3000, Waltham, America).
2.5. Bacterial and Fungal Analysis
Total soil genomic DNA was extracted from each soil sample using a FastDNA kit (MoBio Labs, Solana Beach, CA, USA). The DNA concentration and quality were checked using a NanoDrop Spectrophotometer (NanoDrop Technologies Inc., Wilmington, NC, USA). The extracted DNA was diluted to 10 ng μL−1 and stored at −40 °C for downstream analysis.
In bacteria, the primer pair 515F (5’-GTGCCAGCMGCCGCGG-3’) and reverse primer 907R (5’-CCGTCAATTCMTTTRAGTT-3’) with a unique 6-nt barcode were used to amplify the hypervariable V4 region of the 16S rRNA gene [22]. In terms of fungi, the ITS forward primer ITS1F (5’-CTTGGTCATTTAGAGGAAGTAA-3’) and reverse primer ITS2R (5’-GCTGCGTTCTTCATCGATGC-3’) with a unique 12-nt barcode were used to amplify the ITS1 region. All samples were pooled together with an equal molar amount from each sample. The sequencing samples were prepared using the TruSeq DNA kit according to the manufacturer’s instructions. The purified library was diluted, denatured, rediluted, mixed with PhiX (equal to 30% of final DNA amount) as described in the Illumina library preparation protocols, and then applied to an Illumina MiSeq system for sequencing with the Reagent Kit v2 2 × 250 bp as described in the manufacturer manual.
2.6. Sequences Processing and Statistical Analysis
Processing of the raw sequences obtained through Illumina sequencing was performed using the Quantitative Insights into Microbial Ecology (QIIME) pipeline [23]. The resultant high-quality sequences were then clustered into operational taxonomic units (OTUs) at 97% similarity using the UPARSE algorithm [24]. Taxonomic classification of representative sequences from individual OTUs was performed by the Ribosomal Database Project (RDP version 2.12) Classifier and UNITE database [25,26]. Alpha diversity indices were calculated in QIIME, including Chao1 index, Shannon’s diversity index, Simpson index, and Faith’s PD diversity. Bray–Curtis distance metrics were calculated to complete a nonmetric multidimensional scaling (NMDS), analysis of similarities (ANOSIM), and mantel test with monoMDS, anosim, and mantel in the vegan and permute packages in R [27]. For edaphic factors, the Kruskal–Wallis test (K-W test) was employed to determine differences between groups. Spearman’s rank correlation analysis was applied to determine the relationship between edaphic properties and microbial biodiversity in SPSS software (version 13.0). Linear discriminant analysis effect size (LEfSe) was used to elucidate the effects of each management scenario [28]. The microbial communities that could be classified were selected to detail the management effects. To determine the directed or indirect effects of management, the relationship between classified microbial communities and edaphic properties was determined by Spearman’s rank correlation analysis. Moreover, random forest models were employed to determine the relative importance of management and edaphic properties using the randomForest package in R [29]. The factor’s proportion of importance was calculated by the proportion of the factor’s %IncMSE in the sum of %IncMSE of all factors.
3. Results
3.1. The Effects of Management Scenarios on Edaphic Properties
The results of edaphic factors showed that the between-group differences in soil nutrients. The between-group differences in inorganic carbon and calcium (Ca) were stable (Table 1). In terms of inorganic carbon, the contents in land abandonment (LA) and slight human intervention (HIS) were significantly higher than those of moderate human intervention (HIM) in 2016 (K-W test, P = 0.006). After 2 years, HIS was still significantly higher than HIM (K-W test, P = 0.004), but the difference between LA and HIM became insignificant. In terms of Ca, the content in LA was significantly higher than that in HIM in 2016 (K-W test, P = 0.012). Then, the HIS was significantly higher than the HIM and intensive human intervention (HII) in 2017 and 2018 (K-W test, Both P = 0.003). Moreover, the difference between LA and other scenarios became insignificant. In terms of soil nutrients, soil organic carbon decreased from 6.68 g kg−1 to 4.69 g kg−1 in LA, whereas it increased from 6.27 g kg−1 to 6.50 g kg−1 in HIM. The ratio of carbon to nitrogen (C/N) showed that LA was always lower than HIS (K-W test, 2016 P = 0.004 and 2017 P = 0.020). The ammonium nitrogen (NH4N) content was lower in HIS and HII, and the content of HIS was significantly lower than that of HIM in 2017 and 2018 (K-W test, P = 0.031 and P = 0.011). The content of nitrate-nitrogen (NO3N) showed that LA was always lower than HII (K-W test, 2016 P = 0.004 and 2017 P = 0.020). However, the between-group differences in C/N and NO3N were insignificant in 2018. The β-glucosidase in HIS and HII was significantly lower than that in HIM in 2018 (K-W test, P = 0.003). The correlation between factors in different years showed that soil inorganic carbon (SIC) showed a significant positive correlation with Ca (Spearman’s rank correlation, 2016 rho = 0.965 and P = 0.0001, 2017 rho = 0.937 and P = 0.0001, 2018 rho = 0.937 and P = 0.0001) and a negative correlation with sodium (Na) during the study period (2016 rho = −0.800 and P = 0.002, 2017 rho = −0.930 and P = 0.0001, 2018 rho = −0.783 and P = 0.003) (Figure S2).
3.2. The Effects of Management Scenarios on Microbial Biodiversity
Alpha diversity indicators of bacteria and fungi showed insignificant between-group differences within each year. The results of the correlation between alpha diversity and edaphic factors showed that bacteria were less affected by edaphic factors than fungi (Figure 1). This result indicated that bacterial alpha diversity may be affected by different intensities of HI more than edaphic properties. However, the alpha diversity showed more significant relationships with soil carbon-nitrogen cycling in 2018, such as soil organic carbon (SOC) (Bacterial Shannon’s index and Simpson’s index, rho = 0.608, P = 0.036), total nitrogen (TN) (Bacterial Shannon’s index and Simpson’s index, rho = 0.734, P = 0.007 and rho = 0.860, P = 0.0003), ammonium nitrogen (NH4N) (Fungal Chao1 index, rho = 0.606, P = 0.037), and β-glucosidase (Beta) (Fungal Chao1 index, rho = 0.762, P = 0.004).
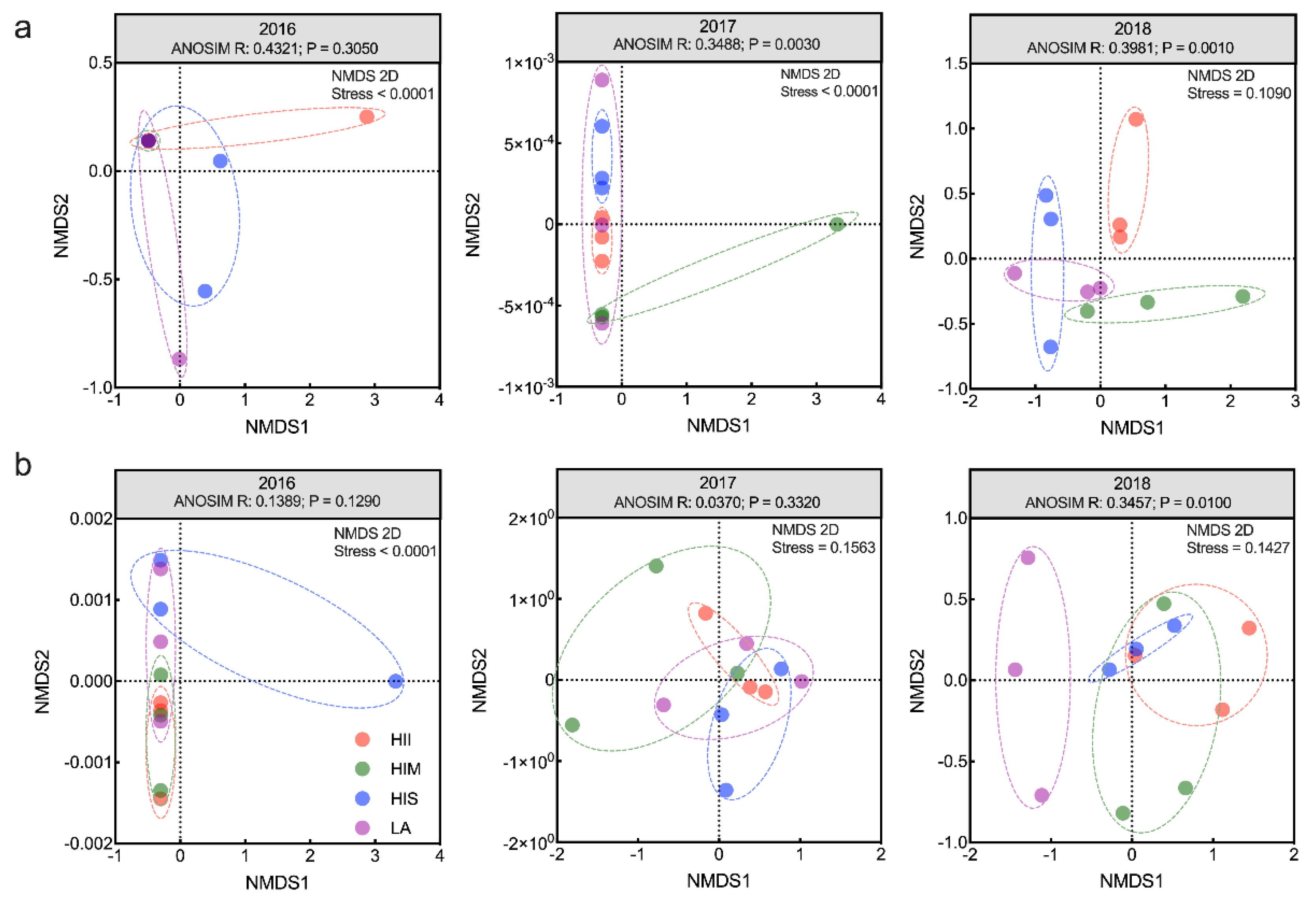
Then, beta diversity was applied to detect the properties of the microbial community structure. Differences in microbial community structure were evident in 2018. For bacteria, ANOSIM showed significant differences among groups from 2017 to 2018 (ANOSIM, R = 0.3488, P = 0.003, and R = 0.3981, P = 0.001). In 2017, the HIS, HIM, and HII began to show between-group differences. In 2018, the between-group differences among LA, HIM, and HII were presented. In addition, the differences among treatments were significant in 2018 (Figure 2a). For fungi, ANOSIM showed significant differences among treatments only in 2018 (R = 0.3457, P = 0.010) (Figure 2b). The LA was different from the other groups, and the HI-groups overlapped in 2018. The results of the Mantel test showed that bacterial community structure was significantly related to NO3N (Mantel statistic r = 0.3279, P = 0.025) in 2016. Then, the bacterial community structure was significantly related to pH (Mantel statistic r = 0.3745, P = 0.035), SIC (Mantel statistic r = 0.3087, P = 0.037), and Ca (Mantel statistic r = 0.3388, P = 0.010) in 2018, and the edaphic properties changed with different levels of HI. The fungal community structure was significantly related to C/N in 2016 (Mantel statistic r = 0.4556, P = 0.033) and was associated with NO3N in 2018 (Mantel statistic r = 0.4287, P = 0.006) (Figure S3). In 2017, the relationship between microbial community structure and edaphic properties was insignificant.
3.3. The Effects of Management Scenarios on Microbial Communities
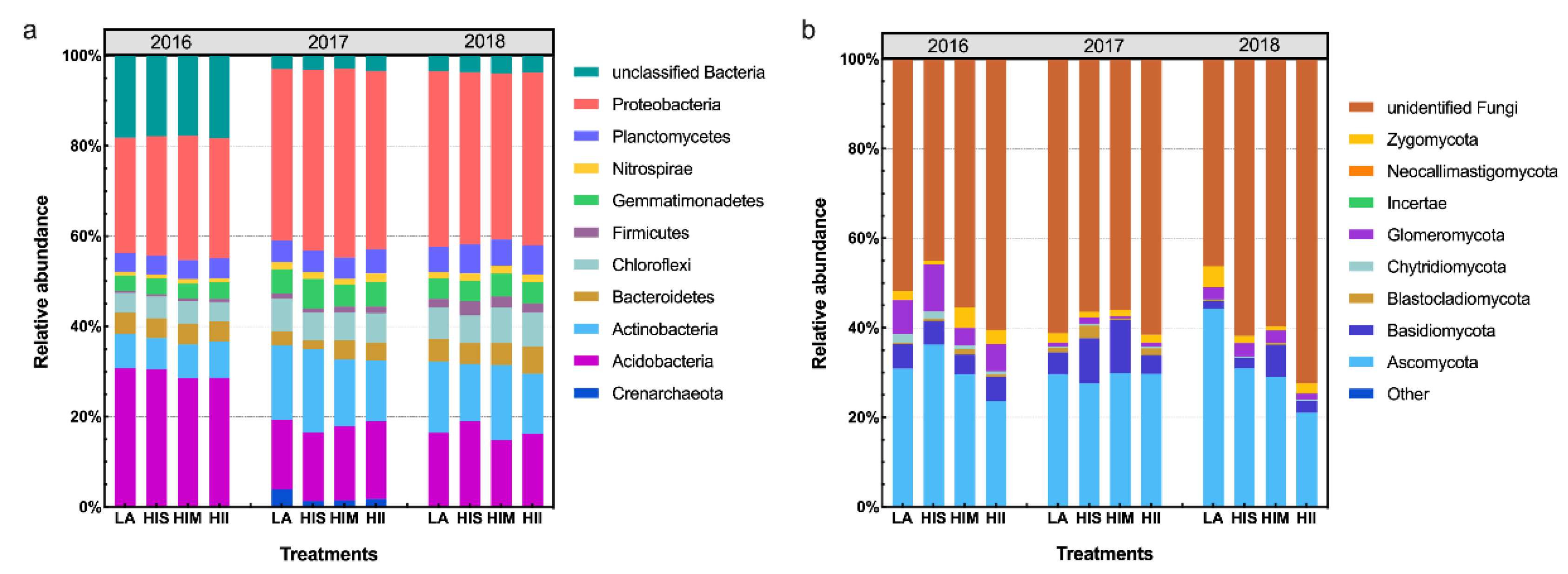
The bacterial community composition and fungal community composition are shown at the phylum level in Figure 3. The LEfSe results showed that the impacts of HI on the microbiota gradually became stronger over time (Figure S4). Notably, fungi were more resistant to HI than bacteria because LA showed dominant effects on fungi in 2016 and 2018. Then, the unclassified microbial communities were ignored, and the microbial communities that could be classified were selected to analyze the management scenario effects.
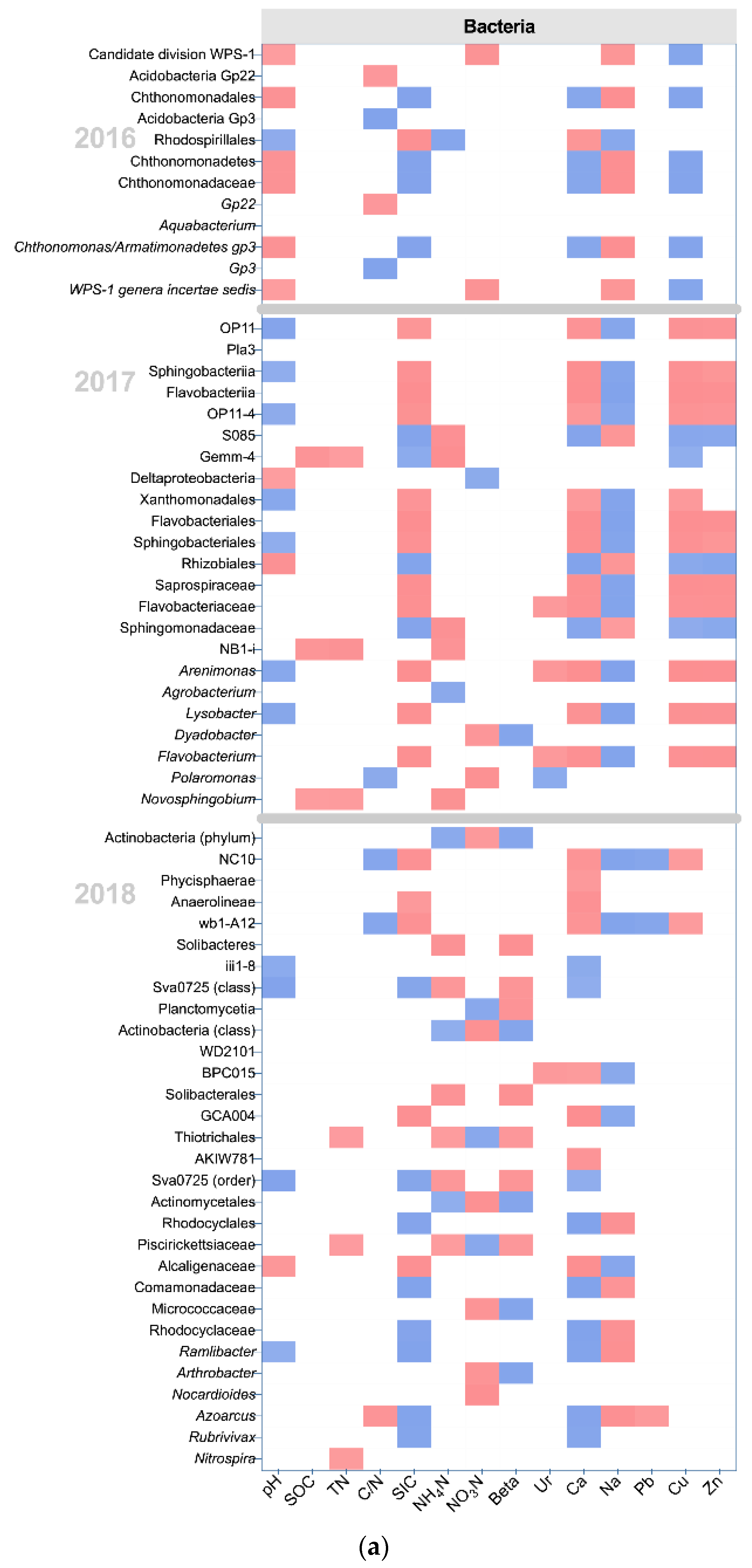
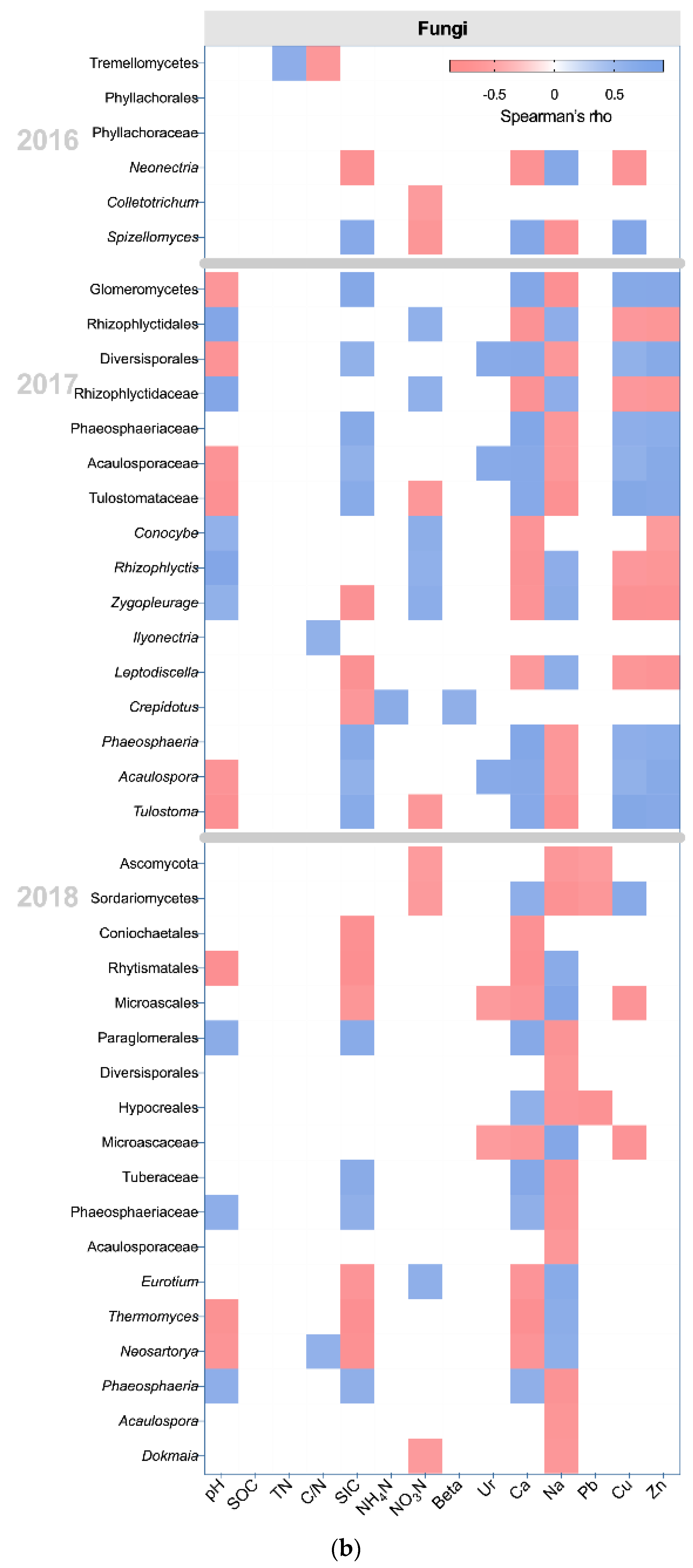
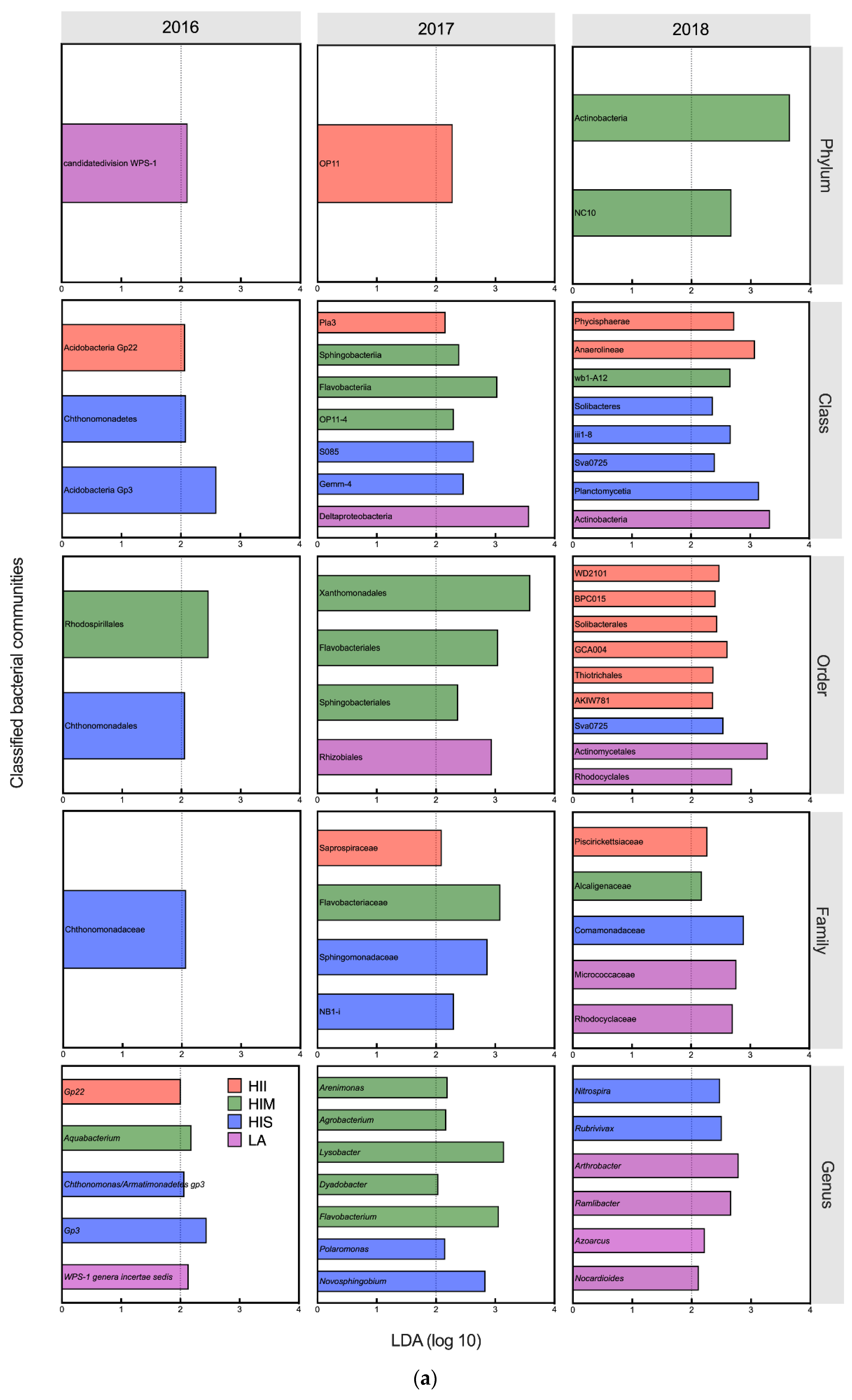
In 2016 and 2017, LA had significant effects on the WPS-1 genera incertae sedis, which prefers natural soil, and on the nitrogen-fixing bacterium Rhizobiales (LDA score > 2, K-W test P < 0.05) [30,31,32] (Figure 4b). In 2018, some nitrogen-fixing bacteria and plant-growth promotion bacteria increased in the LA, including Actinobacteria (class), Actinomycetales, Nocardioides, Azoarcus, Ramlibacter, and Arthrobacter (LDA score > 2, K-W test P < 0.05) [33,34,35,36,37,38,39,40,41] (Figure 4a). In terms of fungi, the abundance of some plant pathogens increased in LA in 2016, including Phyllachoraceae, Colletotrichum, and Spizellomyces (LDA score > 2, K-W test P < 0.05) [42,43]. In 2018, the abundance of some plant pathogens reappeared in LA, including Paraglomerales, Hypocreales, and Phaeosphaeria (LDA score > 2, K-W test P < 0.05) [44,45]. However, some fungi associated with AMF and endomycorrhizae formation were also promoted by LA in 2018, including Diversisporales and Acaulospora (LDA score > 2, K-W test P < 0.05) [46,47].
In 2016 and 2017, HIS had significant effects on some oligotrophic bacteria, including Acidobacteria Gp3, Gemm-4, NB1-I, Polaromonas, and Novosphingobium (LDA score > 2, K-W test P < 0.05) [14,48,49,50]. In 2018, some bacteria involved in nitrogen cycling and plant-growth promotion increased in the HIS, including Solibacteres, Planctomycetia, Sva0725, Rubrivivax, and Nitrospira (LDA score > 2, K-W test P < 0.05) [51,52,53]. In terms of fungi, some fungi associated with AMF and endomycorrhizae formation were promoted in HIS in 2017 and 2018, including Diversisporales, Tuberaceae, and Acaulospora (LDA score > 2, K-W test P < 0.05) [46,47,54]. The plant pathogenic fungus Phaeosphaeria (LDA score = 2.46, K-W test P = 0.049) increased in HIS in 2017 [45].
In 2016 and 2017, some bacteria involved in plant diseases or disease resistance were increased in HIM, including Flavobacteriales, Sphingobacteriales, Agrobacterium, Lysobacter, and Dyadobacter (LDA score > 2, K-W test P < 0.05) [55]. In 2018, HIM had significant effects on the phylum Actinobacteria (LDA score = 3.66, K-W test P = 0.030) associated with organic decomposition [56]. In terms of fungi, HIM had significant effects on plant pathogenic fungi in 2017, including Ilyonectria and Leptodiscella (LDA score > 2, K-W test P < 0.05) [57,58]. Moreover, HIM also increased Crepidotus in 2017 (LDA score = 2.23, K-W test P = 0.047), which was associated with the decay of plant litter [59]. In 2018, HIM simultaneously promoted the plant pathogen Rhytismatales and antagonistic fungus Neosartorya (LDA score > 2, K-W test P < 0.05) [59,60].
In 2016 and 2017, the HII had significant effects on some bacteria that preferred nutritious soil, including Acidobacteria Gp22, OP11, and Saprospiraceae (LDA score > 2, K-W test P < 0.05) [61,62]. In 2018, some bacteria associated with soil nitrogen cycling, including Solibacterales, Thiotrichales, and Piscirickettsiaceae, increased in HII (LDA score > 2, K-W test P < 0.05) [63,64,65]. In terms of fungi, the plant pathogen Conocybe (LDA score = 2.90, K-W test P = 0.042) increased in HII in 2017 [43].
3.4. The Effects of Management and Edaphic Properties on Classified Microbial Communities
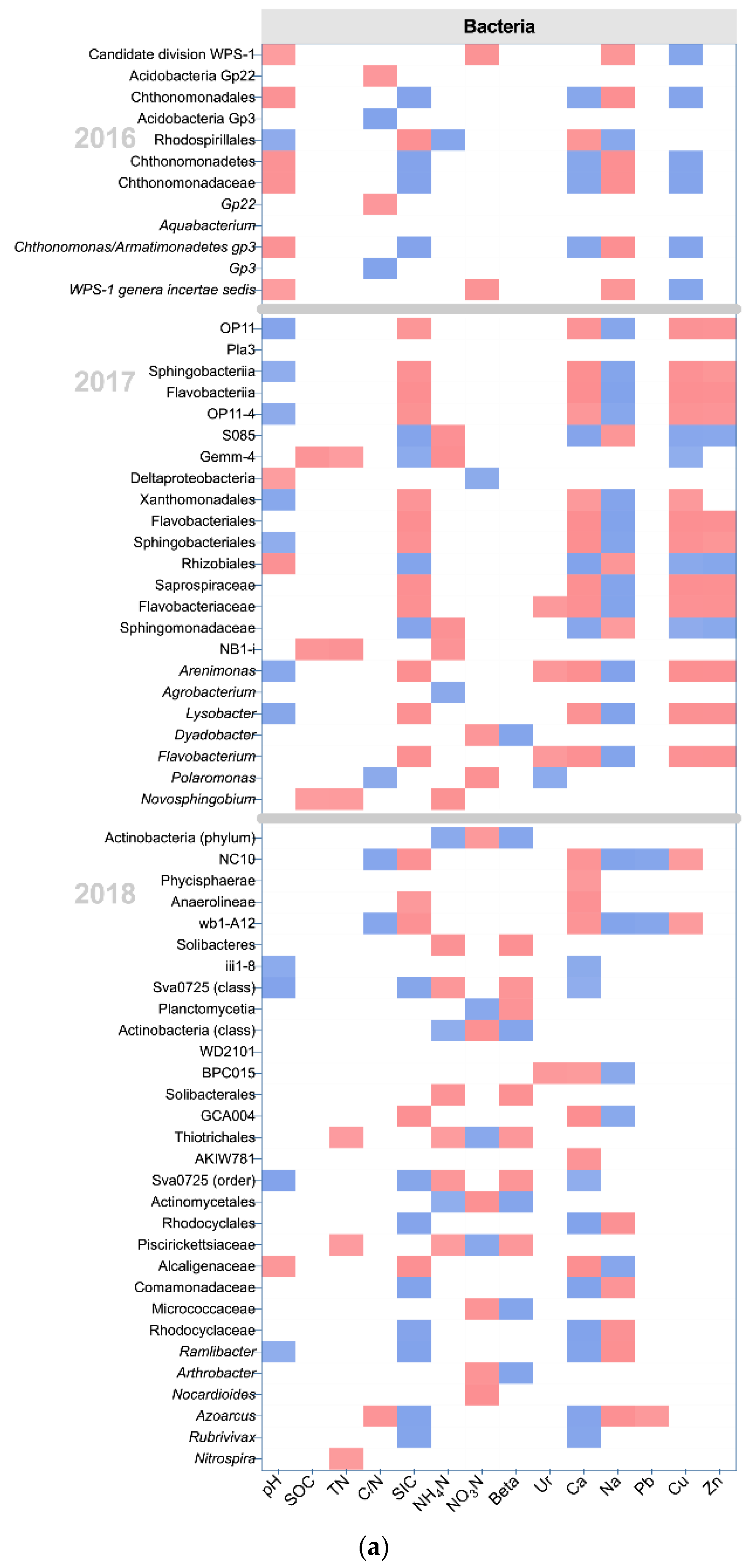
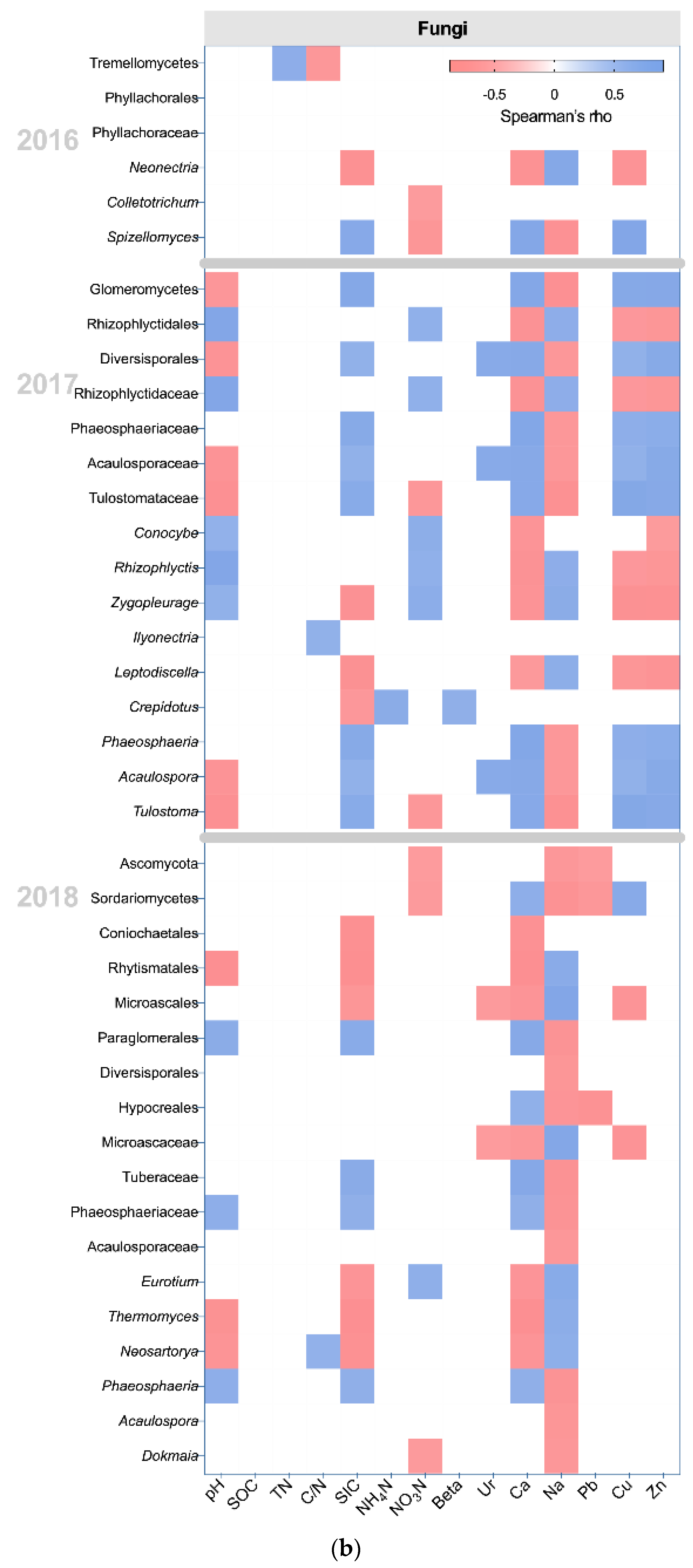
The results of the correlation analysis showed the relationship between microbial communities, which showed between-group differences, and edaphic properties (Figure 5). SOC and TN, the main sources of soil nutrients, were negatively correlated with Gemm-4 (SOC rho = −0.692, P = 0.013 and TN rho = −0.588, P = 0.044), NB1-I (SOC rho = −0.701, P = 0.011 and TN rho = −0.746, P = 0.005), and Novosphingobium (SOC rho = −0.593, P = 0.042 and TN rho = −0.603, P = 0.038) in 2017 (Figure 5a). Those bacterial communities were increased in HIS. Moreover, there were 3 bacterial communities that showed a negative correlation with TN in 2018, including Thiotrichales (rho = −0.601, P = 0.039), Piscirickettsiaceae (rho = −0.601, P = 0.039), and Nitrospira (rho = −0.615, P = 0.033). Thiotrichales and Piscirickettsiaceae were increased by HII, and Nitrospira was promoted by HIS. The activity of β-glucosidase showed a positive correlation with Dyadobacter (rho = 0.761, P = 0.004), phylum Actinobacteria (rho = 0.720, P = 0.008), class Actinobacteria (rho = 0.734, P = 0.007), Actinomycetales (rho = 0.734, P = 0.007), Micrococcaceae (rho = 0.727, P = 0.007), and Arthrobacter (rho = 0.727, P = 0.007) in 2017 and 2018. Dyadobacte and the phylum Actinobacteria were promoted in HIM, and the rest of the bacterial communities were all improved in LA. Carbon cycling may be promoted by spontaneous grass (LA and HIM), especially LA. Negative correlations between β-glucosidase and bacterial communities were exhibited in 2018, including Sva0725 (rho = −0.685, P = 0.014), Planctomycetia (rho = −0.720, P = 0.008), Solibacteres (rho = −0.776, P = 0.003), Solibacterales (rho = −0.769, P = 0.003), and Piscirickettsiaceae (rho = −0.657, P = 0.020). The Sva0725, Planctomycetia, and Solibacteres were increased in HIS, and Solibacterales and Piscirickettsiaceae were increased in HII. The activity of β-glucosidase may be stronger in spontaneous grass (LA and HIM) than in forage grass (HIS and HII). Crepidotus showed a positive correlation with β-glucosidase in 2017 (rho = 0.606, P = 0.037), and HIM promoted the genus (Figure 5b). These fungal relationships complemented the results of the bacterial relationship. The results of relative importance analysis showed that the effects of organic transformation (Beta) on bacteria increased in 2018 (Figure S5).
4. Discussion
4.1. The Risks and Benefits of LA
Agricultural LA requires a long and slow process to restore SOC storage. Therefore, LA does not present significant benefits for organic carbon storage in our study. However, it controlled the loss of SIC, which may be caused by irrigation [66]. The total Ca content also accumulated in LA, which was also consistent with a previous study [66]. In terms of microbiota, LA rapidly resisted continuous cropping obstacles and showed characteristics of natural soils in 2016 [30,31]. LA also benefitted the soil nitrogen-fixing capacity and some plant-growth promotion rhizobacteria which is consistent with our hypothesis 1 [32,33,34,35,36,37,38,39,40,41]. Spontaneous grass may be sensitive to a lack of external nitrogen source in the short term, consistent to a previous study [67]. Some bacteria associated with β-glucosidase activity were also improved in LA. Although the β-glucosidase activity of LA was not significantly different from that of the other groups, the relevant bacteria implied that LA may benefit β-glucosidase activity. In addition, some fungi associated with arbuscular mycorrhizal fungi (AMF) formation and endomycorrhizae formation were also improved in LA [46,47]. LA showed potential for carbon-nitrogen storage and plant growth promotion in our study. These advantages were also mentioned in previous studies [17,38]. However, our study showed that plant pathogenic fungi present in LA repeatedly may increase the risk of plant disease [42,43,44,45]. A previous study indicated that spontaneous grass could provide fungal pathogens with more chances for survival than green manure, so short-term LA increased crop disease risk [64]. Therefore, we should prevent crop diseases when short-term LA is recultivated. Compared with a previous study on LA, our study showed few negative impacts on soil physicochemical properties, such as soil erosion [68]. This result may be because our study area is flat, covered with herbs, and has moderate precipitation.
4.2. The Risks and Benefits of Planting Forage Grass on LA
Forage grass did not cause strong changes in LA. In terms of soil physicochemical properties, HIS also accumulated Ca and inorganic carbon, similar to LA. However, HIS increased nitrogen consumption, which was the same as in HIM and HII. Although the nutrient consumption of HIS did not cause significant between-group differences in our study, the relationship between soil nitrogen content and bacterial communities involved in nitrogen cycling indicated that HIS exhibited oligotrophic status and increased nitrogen consumption [14,48,49,50,51,52]. The reason may be that forage grass enhances nitrogen cycling and causes nitrogen loss via nitrite reduction [69]. Moreover, the negative relationship between β-glucosidase activity and some bacteria showed ineffective cellulose decomposition in HIS. This may be due to harvesting, which took away forage grass each year and reduced plant residues in the soil. However, HIS could improve the abundance of the genus Rubrivivax, which belongs to plant growth-promoting rhizobacteria [53], and it also increased some fungi associated with AMF and endomycorrhizae formation [46,47,54,70]. Compared to LA, the plant pathogens promoted in HIS were only present in 2017, whereas they vanished in 2018 [45]. Phaeosphaeriaceae and Phaeosphaeria decreased from 0.26 and 0.27% to 0.16% in HIS. Therefore, compared to LA, the main issue of HIS is nutrient deficiency, whereas HIS may contribute to the control of plant disease risks. Nevertheless, the results of HIS cannot fully support hypothesis 2. HIS caused oligotrophic soil and nitrogen loss. Therefore, we considered that reducing the seeding density of forage grass may solve this issue. Spontaneous grass may provide nitrogen-fixing bacteria in unfertilized soil. On the other hand, we also designed the HII, which considers the effects of external nutrients and may solve the nutrient deficiency caused by HIS.
4.3. The Risks and Benefits of Cow Manure Inputs on LA
In terms of soil physicochemical properties, cow manure input increased the loss of SIC and total Ca content because of its accompanying irrigation. However, it could resist the loss of soil organic carbon and total nitrogen because of fertilization. HIM also increased the soil ammonium nitrogen content and activity of β-glucosidase. This result implied that HIM benefitted soil fertility. Moreover, HIM could improve some bacteria and fungi associated with soil carbon cycling [56,71,72,73,74]. However, HIM increased the risk of plant disease because of pathogenic bacteria, disease-resistant bacteria, and pathogenic fungi [55,57,58,59,60]. The disease risk it brings may be more than that of LA. Compared to HIS, HIM significantly improved soil fertility. Compared to LA, the advantages of HIM were insignificant in the short term. Nevertheless, the disadvantages of HIM were significant, including the increased disease risk and disappearance of fungi associated with AMF and endomycorrhizae formation and plant growth-promoting rhizobacteria. The reason seems to be that nutrient enrichment has negative effects on fungi associated with AMF and endomycorrhizae formation, which was mentioned in a previous study [75].
4.4. The Risks and Benefits of Intensive Human Interference on LA
When forage grass planting and fertilization appeared in the abandoned land at the same time, the loss of SIC and total Ca content also occurred, which was similar to HIM. However, the activity of β-glucosidase was relatively lower than that of LA and HIM. The relationship between the activity of β-glucosidase and bacterial communities also showed lower cellulose decomposition in HII. This result indicated that HII met the same problem of HIS: less plant cellulose remained in the forage grass participant plots. In addition, the risk of plant diseases only presented in 2016 and then disappeared [42]. The HII also increased nitrogen consumption in 2018 [63,64,65]. These were similar to HIS. Nevertheless, HII and HIS are different in other aspects. The HII solved the issue of nutrient deficiency in HIS [61,70], whereas it failed to improve soil fertility. Moreover, loss of SIC and total Ca content occurred and the fungi associated with AMF and endomycorrhizae formation disappeared, similar to HIM. Compared to LA, the benefit of HII lies in the control of disease risk. Unfortunately, it cannot benefit soil fertility and microbiota associated with mycorrhizal formation and plant growth and fails to improve soil fertility. The results of HII showed that some shortcomings of HIM appeared in HII, and it lost the advantages of HIS. It is consistent with hypothesis 3. Intensive human interference was helpless for restoration duration in the short term, and it caused the loss of soil nutrients.
Compared with LA and HIM, the risks of plant diseases were relatively lower in HIS and HII. In contrast, a previous study showed that LA could decrease the risks of plant disease [76]. This result may be because the LA duration in our study was much shorter than that in the previous study, and the main vegetation was herbaceous plants, which are different from shrubs. Moreover, a relevant study showed that spontaneous grass could provide fungal pathogens with more chances for survival than green manure which increased crop disease risk in the short term [64]. In our study, the disease resistance of forage grass may be because new vegetation changed the monoculture, which altered the fungal community composition and evolution path [77]. The interspecies competition of fungi provides more possibilities for fungal disease resistance.
5. Conclusions
Based on our hypotheses, natural restoration of abandoned land has enhanced some microbial communities involved in nitrogen fixation, plant growth promotion, AMF, and endomycorrhizae formation. Nevertheless, it may face plant disease risk because plant pathogens repeatedly presented in LA during the study period. LA may need more time to solve the problem, and our results showed that the risks of plant diseases could be controlled by appropriate vegetation planting. However, relying on oriented vegetation cannot improve soil fertility and increase soil nitrogen consumption. Farmyard manure inputs caused Ca and SIC loss, and some fungi and bacteria associated with soil fertility and plant growth cannot benefit from manure. This result indicated that short-term farmyard manure inputs have limited effects on improving soil fertility and microbial community structure. Although LA could restore farmland soil, it is still considered to waste resources and is risky in some Chinese areas [78]. When combining abandoned farmland with local animal husbandry, it will be easier for farmers to understand and accept. If there are transportation costs, HIS is the first choice of cattle farm managers. If there is no transportation cost, cattle farm managers may prefer HII. Considering the risk of overgrazing, we believe that HII should be accompanied by a supervision system.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/land10101049/s1, Figure S1: The study area and experimental plots. Figure S2: The correlation analysis of soil properties in each year, based on Spearman’s Rank correlation analysis. Only significant Spearman’s coefficients are shown (p < 0.05). Figure S3: Mantel test between soil properties and microbial community structure. *, **, and *** are used to show statistical significance at the 0.05, 0.01, and 0.001 levels, respectively. Figure S4: The results of linear discriminant analysis effect size (LEfSe) on bacterial and fungal communities. The cutoff of the LDA score is 2. Figure S5: The relative importance ranks of edaphic and human factors on microbial communities, based on %IncMSE of random forests model.
Author Contributions
Conceptualization, T.H. and G.L.; methodology, G.L.; software, T.H.; validation, T.H., G.L., and M.Z.; formal analysis, T.H.; investigation, G.L.; resources, M.Z.; data curation, G.L.; writing—original draft preparation, G.L.; writing—review and editing, M.Z.; visualization, T.H.; supervision, C.W.; project administration, C.W.; funding acquisition, C.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Major Program of the National Social Science Foundation of China (Grant number: 14ZDA039).
Data Availability Statement
All raw sequences data from this study are available at the DDBJ. BioProject: PRJDB11923.
Acknowledgments
We are grateful for the support of the Binzhou Land Improvement and Remediation Engineering Technology Research Center and to all colleagues in Zhejiang University who assisted.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Xu, D.; Deng, X.; Guo, S.; Liu, S. Labor migration and farmland abandonment in rural China: Empirical results and policy implications. J. Environ. Manag. 2019, 232, 738–750. [Google Scholar] [CrossRef]
- Ferrucci, G.; Jiménez-Rodríguez, R.; Onorantea, L. Food Price Pass-Through in the Euro Area: Non-Linearities and the Role of the Common Agricultural Policy. Int. J. Cent. Bank. 2012, 8, 179–218. [Google Scholar]
- Lasanta, T.; Arnáez, J.; Pascual, N.; Ruiz, P.; Errea, M.; Lana-Renault, N. Space–time process and drivers of land abandonment in Europe. CATENA 2017, 149, 810–823. [Google Scholar] [CrossRef]
- Yang, Y.; Hobbie, S.E.; Hernandez, R.R.; Fargione, J.; Grodsky, S.M.; Tilman, D.; Zhu, Y.-G.; Luo, Y.; Smith, T.M.; Jungers, J.M.; et al. Restoring Abandoned Farmland to Mitigate Climate Change on a Full Earth. One Earth 2020, 3, 176–186. [Google Scholar] [CrossRef]
- Tian, H.; Wang, H.; Hui, X.; Wang, Z.; Drijber, R.A.; Liu, J. Changes in soil microbial communities after 10 years of winter wheat cultivation versus fallow in an organic-poor soil in the Loess Plateau of China. PLoS ONE 2017, 12, e0184223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurui, L.; Xuanchang, Z.; Zhi, C.; Zhengjia, L.; Zhi, L.; Yansui, L. Towards the progress of ecological restoration and economic development in China’s Loess Plateau and strategy for more sustainable development. Sci. Total. Environ. 2021, 756, 143676. [Google Scholar] [CrossRef]
- Tarjuelo, R.; Margalida, A.; Mougeot, F. Changing the fallow paradigm: A win–win strategy for the post-2020 Common Agricultural Policy to halt farmland bird declines. J. Appl. Ecol. 2020, 57, 642–649. [Google Scholar] [CrossRef]
- Hauchhum, R.; Tripathi, S.K. Impact of Rhizosphere Microbes of Three Early Colonizing Annual Plants on Improving Soil Fertility During Vegetation Establishment Under Different Fallow Periods Following Shifting Cultivation. Agric. Res. 2019, 9, 213–221. [Google Scholar] [CrossRef]
- Joslin, A.; Markewitz, D.; Morris, L.A.; Oliveira, F.D.A.; Kato, O. Improved fallow: Growth and nitrogen accumulation of five native tree species in Brazil. Nutr. Cycl. Agroecosystems 2016, 106, 1–15. [Google Scholar] [CrossRef]
- Sun, B.; Peng, Y.; Yang, H.; Li, Z.; Gao, Y.; Wang, C.; Yan, Y.; Liu, Y. Alfalfa (Medicago sativa L.)/Maize (Zea mays L.) Intercropping Provides a Feasible Way to Improve Yield and Economic Incomes in Farming and Pastoral Areas of Northeast China. PLoS ONE 2014, 9, e110556. [Google Scholar] [CrossRef]
- Antil, R.S.; Gerzabek, M.H.; Haberhauer, G.; Eder, G. Long-term effects of croppedvs. fallow and fertilizer amendments on soil organic matter I. Organic carbon. J. Plant Nutr. Soil Sci. 2005, 168, 108–116. [Google Scholar] [CrossRef]
- Shirani, H.; Hajabbasi, M.; Afyuni, M.; Hemmat, A. Effects of farmyard manure and tillage systems on soil physical properties and corn yield in central Iran. Soil Tillage Res. 2002, 68, 101–108. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wu, C.; Gao, W. Effects of short-term fallow managements on soil microbial properties: A case study in China. Appl. Soil Ecol. 2018, 125, 128–137. [Google Scholar] [CrossRef]
- Lammel, D.R.; Feigl, B.J.; Cerri, C.C.; Nüsslein, K. Specific microbial gene abundances and soil parameters contribute to C, N, and greenhouse gas process rates after land use change in Southern Amazonian Soils. Front. Microbiol. 2015, 6, 1057. [Google Scholar] [CrossRef] [Green Version]
- Bossio, D.; Girvan, M.S.; Verchot, L.; Bullimore, J.; Borelli, T.; Albrecht, A.; Scow, K.; Ball, A.; Pretty, J.; Osborn, A.M. Soil Microbial Community Response to Land Use Change in an Agricultural Landscape of Western Kenya. Microb. Ecol. 2005, 49, 50–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchane, M.N.; Jama, B.; Othieno, C.; Okalebo, R.; Odee, D.; Machua, J.; Jansa, J. Influence of improved fallow systems and phosphorus application on arbuscular mycorrhizal fungi symbiosis in maize grown in western Kenya. Agrofor. Syst. 2010, 78, 139–150. [Google Scholar] [CrossRef]
- Moghimian, N.; Hosseini, S.M.; Kooch, Y.; Darki, B.Z. Impacts of changes in land use/cover on soil microbial and enzyme activities. CATENA 2017, 157, 407–414. [Google Scholar] [CrossRef]
- Lv, K. Spatial and Temporal Variation of Soil Nutrients and Its Influencing Factors—A Case Study of Wudi County. Master’s Thesis, Zhejiang University, Hangzhou, China, 2018. [Google Scholar]
- Dick, R.P.; Breakwell, D.P.; Turco, R.F. Soil Enzyme Activities and Biodiversity Measurements as Integrative Microbiological Indicators. In Methods for Assessing Soil Quality; Doran, J.W., Jones, A.J., Eds.; the Soil Science Society of America, Inc.: Madison, WI, USA, 2015; pp. 247–271. [Google Scholar] [CrossRef]
- Hoffmann, G.; Teicher, K. Ein kolorimetrisches Verfahren zur Bestimmung der Ureaseaktivität in Böden. J. Plant Nutr. Soil Sci. 1961, 95, 55–63. [Google Scholar] [CrossRef]
- Angenent, L.T.; Kelley, S.T.; Amand, A.S.; Pace, N.R.; Hernandez, M.T. Molecular identification of potential pathogens in water and air of a hospital therapy pool. Proc. Natl. Acad. Sci. USA 2005, 102, 4860–4865. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, I.J.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naїve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarenkov, K.; Nilsson, H.; Larsson, K.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. R News 2002, 2, 18–22. [Google Scholar]
- Sánchez-Marañón, M.; Miralles, I.; Garrido, J.F.A.; Anguita-Maeso, M.; Millán, V.; Ortega, R.; García-Salcedo, J.A.; Martínez-Abarca, F.; Soriano, M. Changes in the soil bacterial community along a pedogenic gradient. Sci. Rep. 2017, 7, 14593. [Google Scholar] [CrossRef]
- Tan, G.; Liu, Y.; Peng, S.; Yin, H.; Meng, D.; Tao, J.; Gu, Y.; Li, J.; Yang, S.; Xiao, N.; et al. Soil potentials to resist continuous cropping obstacle: Three field cases. Environ. Res. 2021, 200, 111319. [Google Scholar] [CrossRef]
- Garrido-Oter, R.; Nakano, R.T.; Dombrowski, N.; Ma, K.-W.; McHardy, A.C.; Schulze-Lefert, P. Modular Traits of the Rhizobiales Root Microbiota and Their Evolutionary Relationship with Symbiotic Rhizobia. Cell Host Microbe 2018, 24, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef]
- Sathya, A.; Vijayabharathi, R.; Gopalakrishnan, S. Plant growth-promoting actinobacteria: A new strategy for enhancing sustainable production and protection of grain legumes. 3 Biotech 2017, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Zheng, C.; Liang, L.; Yi, Z.; Xue, S. Quantitative assessment of the potential for soil improvement by planting Miscanthus on saline-alkaline soil and the underlying microbial mechanism. GCB Bioenergy 2021, 13, 1191–1205. [Google Scholar] [CrossRef]
- Abdulla, H.M.; El Shatoury, S. Actinomycetes in rice straw decomposition. Waste Manag. 2007, 27, 850–853. [Google Scholar] [CrossRef]
- Pedrinho, A.; Mendes, L.W.; Merloti, L.F.; Andreote, F.D.; Tsai, S.M. The natural recovery of soil microbial community and nitrogen functions after pasture abandonment in the Amazon region. FEMS Microbiol. Ecol. 2020, 96, fiaa149. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhang, M.; Wu, C. Short-term fallow practices drive soil bacterial community changes: A case study from China. Appl. Soil Ecol. 2021, 165, 103988. [Google Scholar] [CrossRef]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Banach, A.; Błaszczyk, M. Indicators of arable soils fatigue—Bacterial families and genera: A metagenomic approach. Ecol. Indic. 2018, 93, 490–500. [Google Scholar] [CrossRef]
- Safdarian, M.; Askari, H.; Nematzadeh, G. Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci. Rep. 2019, 9, 1792. [Google Scholar] [CrossRef]
- Tang, X.; Zhong, R.; Jiang, J.; He, L.; Huang, Z.; Shi, G.; Wu, H.; Liu, J.; Xiong, F.; Han, Z.; et al. Cassava/peanut intercropping improves soil quality via rhizospheric microbes increased available nitrogen contents. BMC Biotechnol. 2020, 20, 13. [Google Scholar] [CrossRef] [Green Version]
- Intra, B.; Mungsuntisuk, I.; Nihira, T.; Igarashi, Y.; Panbangred, W. Identification of actinomycetes from plant rhizospheric soils with inhibitory activity against Colletotrichum spp., the causative agent of anthracnose disease. BMC Res. Notes 2011, 4, 98. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Yu, H.; Zhou, X.; Wu, F. Cucumber (Cucumis sativus L.) Seedling Rhizosphere Trichoderma and Fusarium spp. Communities Altered by Vanillic Acid. Front. Microbiol. 2018, 9, 2195. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Jurado, I.; Fernández-Bravo, M.; Campos, C.; Quesada-Moraga, E. Diversity of entomopathogenic Hypocreales in soil and phylloplanes of five Mediterranean cropping systems. J. Invertebr. Pathol. 2015, 130, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.A.; Lee, K.; Park, B.; Park, H.; Park, K.; Choi, I.-G.; Chang, I.S. Comparative study of the airborne microbial communities and their functional composition in fine particulate matter (PM2.5) under non-extreme and extreme PM2.5 conditions. Atmospheric Environ. 2018, 194, 82–92. [Google Scholar] [CrossRef]
- Jansa, J.; Mozafar, A.; Anken, T.; Ruh, R.; Sanders, I.R.; Frossard, E. Diversity and structure of AMF communities as affected by tillage in a temperate soil. Mycorrhiza 2002, 12, 225–234. [Google Scholar] [CrossRef]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Saixi, Y.; Yi, R.; Baoyin, T. Characterization of soil microbes associated with a grazing-tolerant grass species, Stipa breviflora, in the Inner Mongolian desert steppe. Ecol. Evol. 2020, 10, 10607–10618. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Li, J.; Bai, P.; Li, Q.; Shen, M.; Li, R.; Li, T.; Zhao, J. Comparative Genomics of Degradative Novosphingobium Strains with Special Reference to Microcystin-Degrading Novosphingobium sp. THN1. Front. Microbiol. 2018, 9, 2238. [Google Scholar] [CrossRef] [Green Version]
- Yagi, J.M.; Sims, D.; Brettin, T.; Bruce, D.; Madsen, E.L. The genome of Polaromonas naphthalenivorans strain CJ2, isolated from coal tar-contaminated sediment, reveals physiological and metabolic versatility and evolution through extensive horizontal gene transfer. Environ. Microbiol. 2009, 11, 2253–2270. [Google Scholar] [CrossRef] [PubMed]
- Mackelprang, R.; Grube, A.M.; Lamendella, R.; Jesus, E.D.C.; Copeland, A.; Liang, C.; Jackson, R.D.; Rice, C.W.; Kapucija, S.; Parsa, B.; et al. Microbial Community Structure and Functional Potential in Cultivated and Native Tallgrass Prairie Soils of the Midwestern United States. Front. Microbiol. 2018, 9, 1775. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Yang, J.; Wang, X.; Xie, W.; Zheng, F.; Li, H.; Tang, C.; Zhu, H. Response of soil characteristics and bacterial communities to nitrogen fertilization gradients in a coastal salt-affected agroecosystem. Land Degrad. Dev. 2021, 32, 338–353. [Google Scholar] [CrossRef]
- Mujahid, M.; Sasikala, C.; Ramana, C.V. Production of indole-3-acetic acid and related indole derivatives from L-tryptophan by Rubrivivax benzoatilyticus JA2. Appl. Microbiol. Biotechnol. 2011, 89, 1001–1008. [Google Scholar] [CrossRef]
- Hrynkiewicz, K.; Haug, I.; Baum, C. Ectomycorrhizal community structure under willows at former ore mining sites. Eur. J. Soil Biol. 2008, 44, 37–44. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, X.; Zhang, J.; Cai, Z. Highly connected taxa located in the microbial network are prevalent in the rhizosphere soil of healthy plant. Biol. Fertil. Soils 2019, 55, 299–312. [Google Scholar] [CrossRef]
- Abbott, S.P.; Sigler, L.; Currah, R.S. Microascus brevicaulis sp. nov., the teleomorph of Scopulariopsis brevicaulis, supports placement of Scopulariopsiswith the Microascaceae. Mycologia 1998, 90, 297–302. [Google Scholar] [CrossRef]
- Vitale, A.; Aiello, D.; Guarnaccia, V.; Perrone, G.; Stea, G.; Polizzi, G. First Report of Root Rot Caused by Ilyonectria (=Neonectria) macrodidyma on Avocado (Persea americana) in Italy. J. Phytopathol. 2012, 160, 156–159. [Google Scholar] [CrossRef]
- Hernández-Restrepo, M.; Bezerra, J.D.P.; Tan, Y.P.; Wiederhold, N.; Crous, P.; Guarro, J.; Gené, J. Re-evaluation of Mycoleptodiscus species and morphologically similar fungi. Persoonia Mol. Phylogeny Evol. Fungi 2019, 42, 205–227. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.-L.; Piepenbring, M. New species and new records of Rhytismatales from Panama. Mycologia 2009, 101, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Pattarasaikul, W.; Soytong, K.; Poeaim, S. Biological control of anthracnose disease on banana var ‘Namwa Mali-Ong’ by Neosartorya species. Int. J. Agric. Technol. 2018, 14, 1589–1598. [Google Scholar]
- Li, H.; Cai, X.; Gong, J.; Xu, T.; Ding, G.-C.; Li, J. Long-Term Organic Farming Manipulated Rhizospheric Microbiome and Bacillus Antagonism Against Pepper Blight (Phytophthora capsici). Front. Microbiol. 2019, 10, 342. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.; Chu, L.; Wang, J. Biological treatment of actual petrochemical wastewater using anaerobic/anoxic/oxic process and the microbial diversity analysis. Appl. Microbiol. Biotechnol. 2016, 100, 10193–10202. [Google Scholar] [CrossRef]
- Mason, L.; Eagar, A.; Patel, P.; Blackwood, C.; DeForest, J. Potential microbial bioindicators of phosphorus mining in a temperate deciduous forest. J. Appl. Microbiol. 2021, 130, 109–122. [Google Scholar] [CrossRef]
- Wiggins, B.E.; Kinkel, L.L. Green manures and crop sequences influence alfalfa root rot and pathogen inhibitory activity among soil-borne streptomycetes. Plant Soil 2005, 268, 271–283. [Google Scholar] [CrossRef]
- Burns, A.S.; Padilla, C.C.; Pratte, Z.A.; Gilde, K.; Regensburger, M.; Hall, E.; Dove, A.D.M.; Stewart, F.J. Broad Phylogenetic Diversity Associated with Nitrogen Loss through Sulfur Oxidation in a Large Public Marine Aquarium. Appl. Environ. Microbiol. 2018, 84, e01250–e01218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Wang, X.; Li, X.; Xu, M.; Li, Y.; Zheng, H.; Luo, Y.; Smith, P. Impacts of land use and salinization on soil inorganic and organic carbon in the middle-lower Yellow River Delta. Pedosphere 2021, 31, 839–848. [Google Scholar] [CrossRef]
- Huhe; Borjigin, S.; Cheng, Y.; Nomura, N.; Nakajima, T.; Nakamura, T.; Uchiyama, H. Effect of Abandonment on Diversity and Abundance of Free-Living Nitrogen-Fixing Bacteria and Total Bacteria in the Cropland Soils of Hulun Buir, Inner Mongolia. PLoS ONE 2014, 9, e106714. [Google Scholar] [CrossRef] [Green Version]
- Koulouri, M.; Giourga, C. Land abandonment and slope gradient as key factors of soil erosion in Mediterranean terraced lands. CATENA 2007, 69, 274–281. [Google Scholar] [CrossRef]
- Liu, M.; Han, G.; Zhang, Q. Effects of agricultural abandonment on soil aggregation, soil organic carbon storage and stabilization: Results from observation in a small karst catchment, Southwest China. Agric. Ecosyst. Environ. 2020, 288, 106719. [Google Scholar] [CrossRef]
- Taniguchi, T.; Yuzawa, T.; HuiPing, M.; Yamamoto, F.; Yamanaka, N. Plantation soil inoculation combined with straw checkerboard barriers enhances ectomycorrhizal colonization and subsequent growth of nursery grown Pinus tabulaeformis seedlings in a dryland. Ecol. Eng. 2021, 163, 106191. [Google Scholar] [CrossRef]
- Aburto-Medina, A.; Adetutu, E.M.; Aleer, S.; Weber, J.; Patil, S.S.; Sheppard, P.J.; Ball, A.S.; Juhasz, A.L. Comparison of indigenous and exogenous microbial populations during slurry phase biodegradation of long-term hydrocarbon-contaminated soil. Biodegradation 2012, 23, 813–822. [Google Scholar] [CrossRef]
- Thammannagowda, S.; Magnusson, L.; Jo, J.H.; Maness, P.C.; Seibert, M. Renewable Hydrogen from Biomass. In Encyclopedia of Biological Chemistry, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press: Waltham, MA, USA, 2013; pp. 72–75. [Google Scholar] [CrossRef]
- Raggi, L.; García-Guevara, F.; Godoy-Lozano, E.E.; Martínez-Santana, A.; Escobar-Zepeda, A.; Gutierrez-Rios, R.M.; Loza, A.; Merino, E.; Sanchez-Flores, A.; Licea-Navarro, A.; et al. Metagenomic Profiling and Microbial Metabolic Potential of Perdido Fold Belt (NW) and Campeche Knolls (SE) in the Gulf of Mexico. Front. Microbiol. 2020, 11, 1825. [Google Scholar] [CrossRef]
- Zak, D.R.; Argiroff, W.; Freedman, Z.B.; Upchurch, R.A.; Entwistle, E.M.; Romanowicz, K. Anthropogenic N deposition, fungal gene expression, and an increasing soil carbon sink in the Northern Hemisphere. Ecology 2019, 100, e02804. [Google Scholar] [CrossRef]
- Ma, X.; Geng, Q.; Zhang, H.; Bian, C.; Chen, H.Y.H.; Jiang, D.; Xu, X. Global negative effects of nutrient enrichment on arbuscular mycorrhizal fungi, plant diversity and ecosystem multifunctionality. New Phytol. 2021, 229, 2957–2969. [Google Scholar] [CrossRef] [PubMed]
- Bosso, L.; Lacatena, F.; Varlese, R.; Nocerino, S.; Cristinzio, G.; Russo, D. Plant pathogens but not antagonists change in soil fungal communities across a land abandonment gradient in a Mediterranean landscape. Acta Oecologica 2017, 78, 1–6. [Google Scholar] [CrossRef]
- Umaerus, V.R.; Scholte, K.; Turkensteen, L.J. Crop Rotation and the Occurrence of Fungal Diseases in Potatoes. Effects of Crop Rotation on Potato Production in the Temperate Zones. In Proceedings of the International Conference on Effects of Crop Rotation on Potato Production in the Temperate Zones, Wageningen, The Netherlands, 14–19 August 1988; Vos, J., Van Loon, C.D., Bollen, G.J., Eds.; Springer: Dordrecht, The Netherlands, 1989; pp. 171–189. [Google Scholar] [CrossRef]
- Xiao, G.; Zhu, X.; Hou, C.; Xia, X. Extraction and analysis of abandoned farmland: A case study of Qingyun and Wudi counties in Shandong Province. J. Geogr. Sci. 2019, 29, 581–597. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The correlation between soil properties and microbial alpha diversity in each year, based on Spearman’s Rank correlation analysis. *, ** are used to show statistical significance at the 0.05, 0.01 levels, respectively. The colors indicate the values of Spearman’s rho.
Figure 1.
The correlation between soil properties and microbial alpha diversity in each year, based on Spearman’s Rank correlation analysis. *, ** are used to show statistical significance at the 0.05, 0.01 levels, respectively. The colors indicate the values of Spearman’s rho.
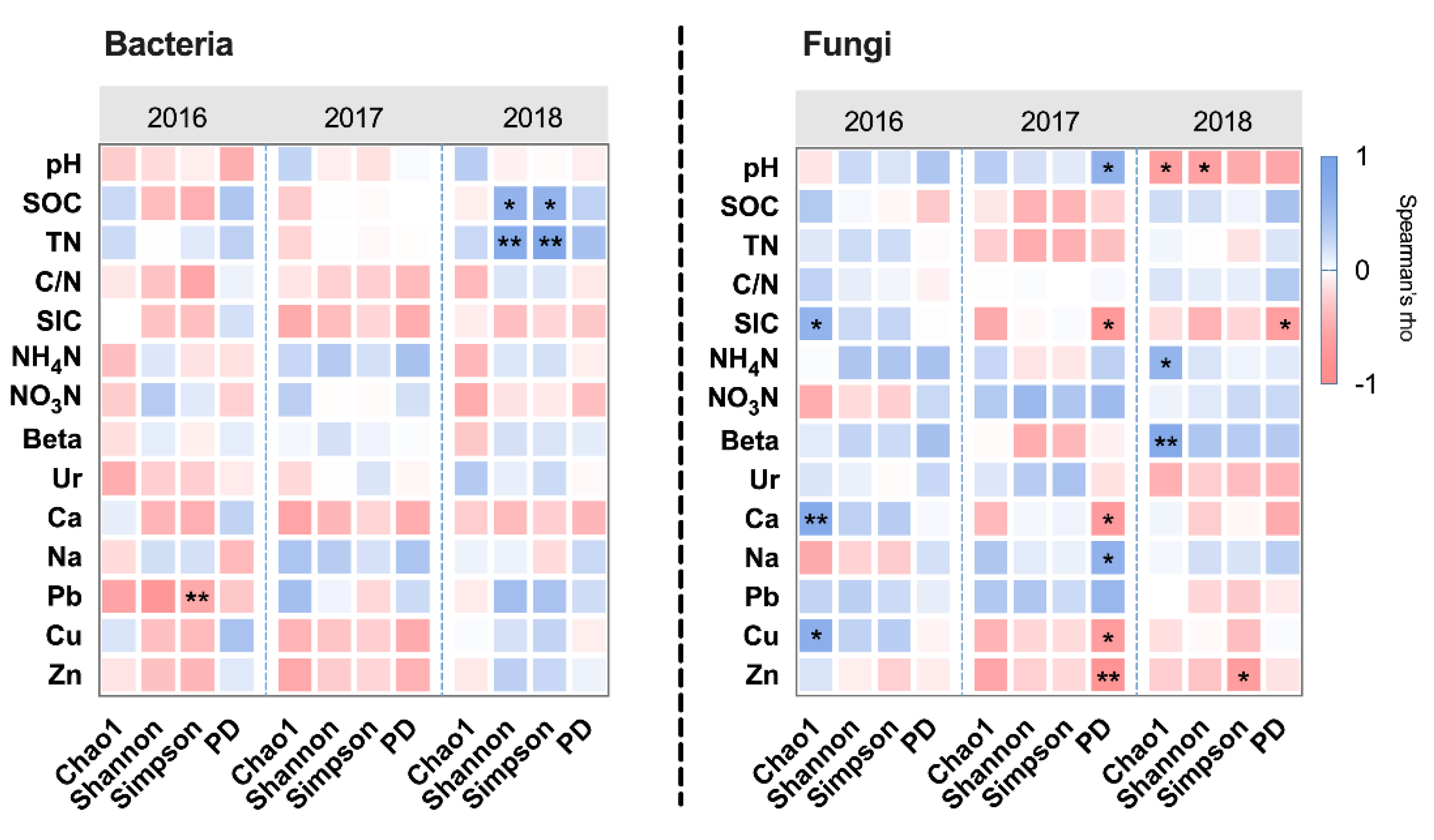
Figure 2.
NMDS and ANOSIM of the bacterial community (a), and NMDS and ANOSIM of the fungal community (b).
Figure 2.
NMDS and ANOSIM of the bacterial community (a), and NMDS and ANOSIM of the fungal community (b).
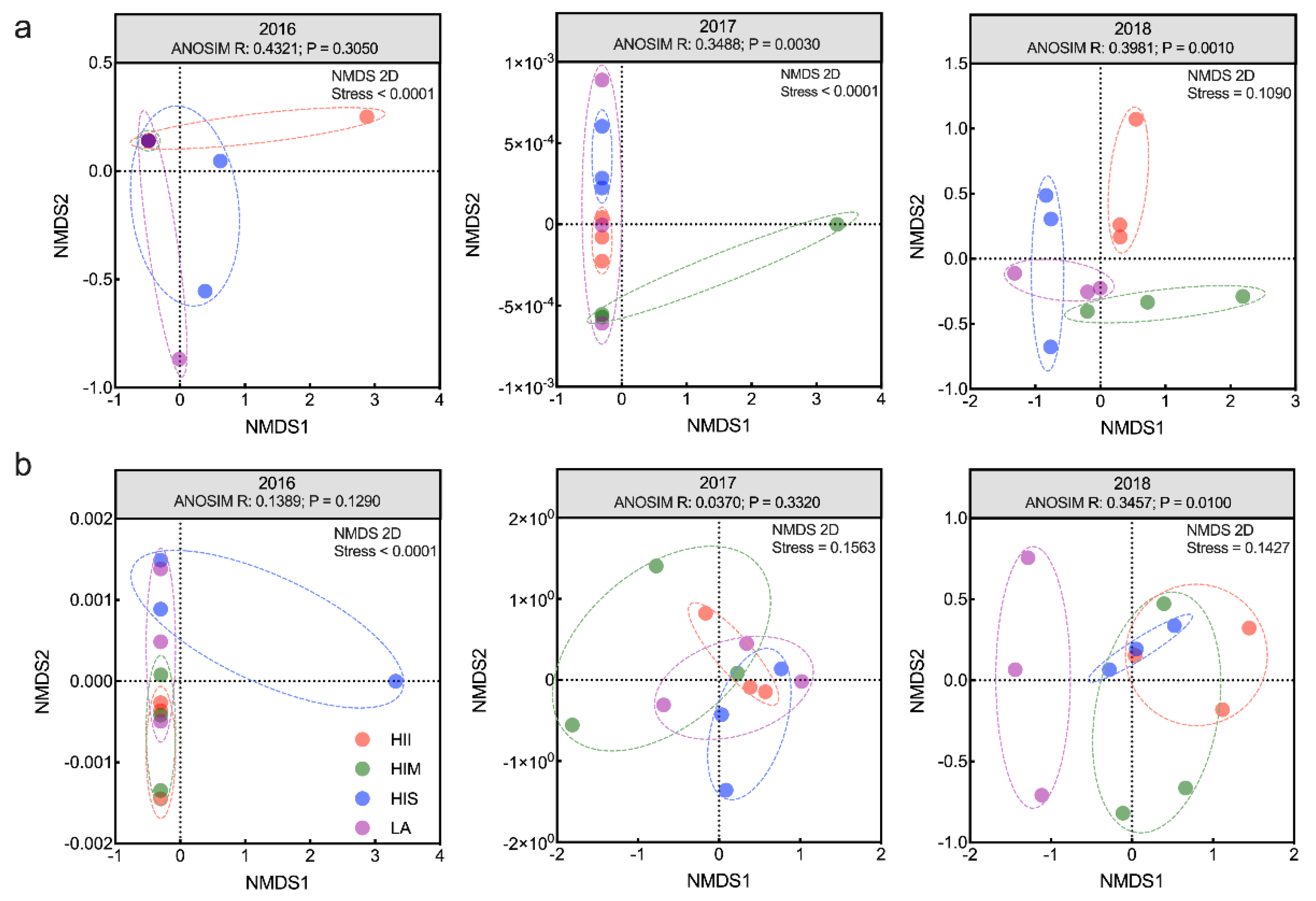
Figure 3.
Bacterial community composition at the phylum level (a), and fungal community composition at the phylum level (b).
Figure 3.
Bacterial community composition at the phylum level (a), and fungal community composition at the phylum level (b).
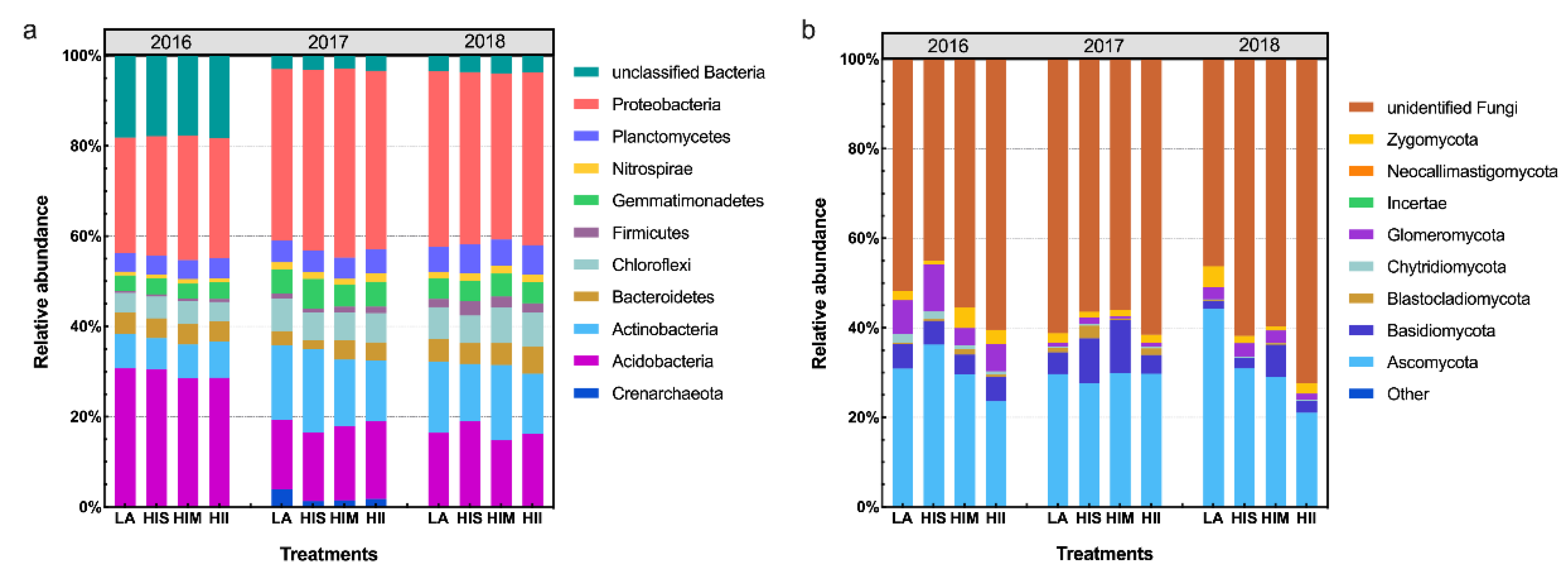
Figure 4.
The Linear Discriminant Analysis (LDA) results of microbial communities at the genus level. Only the classified genera (selected genera) were exhibited in the figure. The cutoff of LDA score is 2. (a) The effects of management groups on bacterial genera; (b) The effects of management groups on fungal genera.
Figure 4.
The Linear Discriminant Analysis (LDA) results of microbial communities at the genus level. Only the classified genera (selected genera) were exhibited in the figure. The cutoff of LDA score is 2. (a) The effects of management groups on bacterial genera; (b) The effects of management groups on fungal genera.
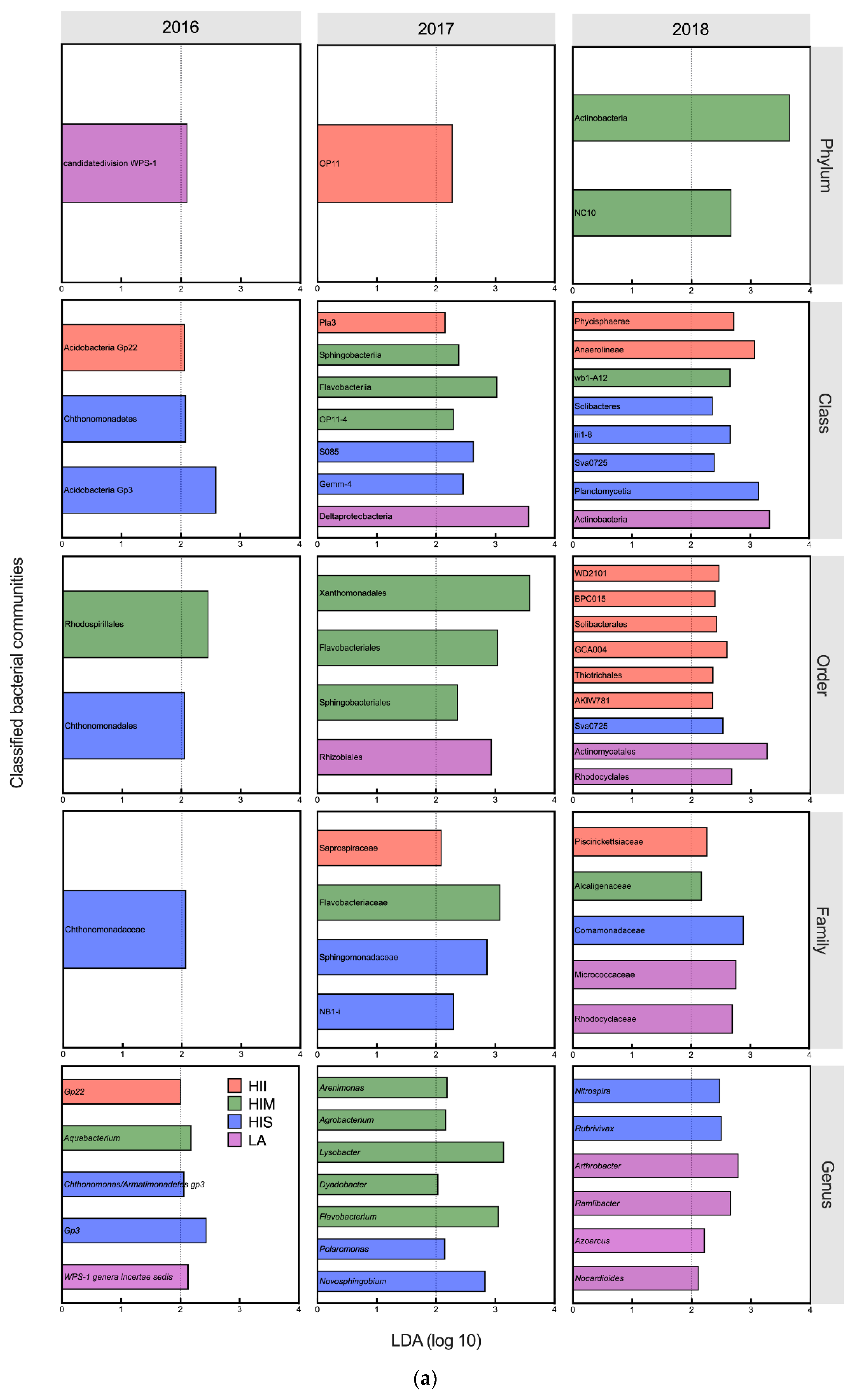

Figure 5.
The correlation between edaphic properties and classified microbial communities, based on Spearman’s Rank correlation analysis. Only significant Spearman’s coefficients (rho) are shown (p < 0.05). The blue color represents a positive correlation, and the red color represents a negative correlation. (a) The relationships between edaphic properties and classified bacterial communities; (b) The relationships between edaphic properties and classified fungal communities.
Figure 5.
The correlation between edaphic properties and classified microbial communities, based on Spearman’s Rank correlation analysis. Only significant Spearman’s coefficients (rho) are shown (p < 0.05). The blue color represents a positive correlation, and the red color represents a negative correlation. (a) The relationships between edaphic properties and classified bacterial communities; (b) The relationships between edaphic properties and classified fungal communities.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The differences in edaphic properties are caused by human interferences.
| Edaphic Properties | LA (n = 3) | HIS (n = 3) | HIM (n = 3) | HII (n = 3) | K-W Statistic (Significance) |
|---|---|---|---|---|---|
| 2016 | |||||
| pH | 8.40 ± 0.04 a | 8.42 ± 0.08 a | 8.56 ± 0.10 a | 8.58 ± 0.14 a | 4.66, ns |
| Ca (g kg−1) | 50.45 ± 3.40 a | 49.25 ± 2.64 a | 43.25 ± 1.69 b | 43.58 ± 0.91 b | 8.33, * |
| Na (g kg−1) | 12.58 ± 0.46 b | 13.18 ± 0.37 ab | 14.07 ± 0.08 a | 14.35 ± 0.42 a | 9.26, ** |
| SIC (g kg−1) | 13.03 ± 1.23 a | 12.78 ± 0.61 a | 11.41 ± 0.52 b | 11.69 ± 0.52 ab | 8.69, ** |
| SOC (g kg−1) | 6.68 ± 0.91 a | 6.47 ± 0.77 a | 6.27 ± 0.39 a | 5.46 ± 0.52 a | 4.44, ns |
| TN (g kg−1) | 1.08 ± 0.13 a | 0.71 ± 0.19 a | 0.84 ± 0.06 a | 0.86 ± 0.02 a | 6.15, ns |
| C/N | 6.20 ± 0.18 b | 9.40 ± 1.95 a | 7.45 ± 0.45 ab | 6.35 ± 0.52 b | 8.95, ** |
| NH4N (mg kg−1) | 0.93 ± 0.35 a | 0.47 ± 0.41 a | 1.68 ± 0.85 a | 0.90 ± 0.14 a | 4.51, ns |
| NO3N (mg kg−1) | 2.37 ± 0.18 b | 6.67 ± 4.07 ab | 8.91 ± 1.46 ab | 13.65 ± 2.85 a | 8.74, ** |
| Beta (PNP mg g−1) | 33.99 ± 2.94 a | 33.04 ± 11.40 a | 38.01 ± 14.08 a | 33.59 ± 14.23 a | 0.54, ns |
| Ur (NH4N mg g−1) | 0.43 ± 0.06 a | 0.47 ± 0.05 a | 0.48 ± 0.21 a | 0.42 ± 0.05 a | 1.77, ns |
| Pb (mg kg−1) | 19.03 ± 3.84 a | 25.56 ± 8.36 a | 17.07 ± 0.53 a | 16.91 ± 1.02 a | 0.90, ns |
| Cu (mg kg−1) | 42.16 ± 2.77 a | 32.33 ± 0.54 ab | 30.36 ± 0.77 b | 28.63 ± 1.77 b | 9.67, *** |
| Zn (mg kg−1) | 87.37 ± 7.95 a | 82.7 ± 8.24 a | 81.1 ± 1.64 a | 82.35 ± 2.73 a | 1.56, ns |
| 2017 | |||||
| pH | 8.98 ± 0.01 b | 9.11 ± 0.01 ab | 9.13 ± 0.11 ab | 9.29 ± 0.05 a | 9.23, ** |
| Ca (g kg−1) | 48.39 ± 6.83 ab | 56.73 ± 3.05 a | 38.45 ± 0.38 b | 38.00 ± 0.91 b | 9.05, ** |
| Na (g kg−1) | 11.03 ± 0.20 ab | 10.71 ± 0.76 a | 13.55 ± 0.45 b | 13.58 ± 0.88 b | 8.44, * |
| SIC (g kg−1) | 13.71 ± 0.51 ab | 14.92 ± 0.98 a | 11.87 ± 0.12 b | 12.17 ± 0.62 b | 9.05, ** |
| SOC (g kg−1) | 6.27 ± 0.38 a | 5.56 ± 0.10 ab | 6.33 ± 0.64 a | 4.96 ± 0.57 b | 8.64, ** |
| TN (g kg−1) | 0.98 ± 0.06 a | 0.73 ± 0.04 b | 0.84 ± 0.08 ab | 0.69 ± 0.04 b | 9.46, ** |
| C/N | 6.39 ± 0.41 b | 7.66 ± 0.26 a | 7.56 ± 0.15 a | 7.13 ± 0.42 ab | 7.82, * |
| NH4N (mg kg−1) | 4.50 ± 1.09 a | 0.82 ± 0.73 b | 6.08 ± 2.06 a | 3.44 ± 0.56 ab | 8.44, * |
| NO3N (mg kg−1) | 2.51 ± 0.83 b | 9.09 ± 5.64 a | 8.51 ± 2.55 ab | 14.96 ± 1.84 a | 9.46, ** |
| Beta (PNP mg g−1) | 37.74 ± 17.18 a | 26.60 ± 12.03 a | 51.07 ± 16.87 a | 21.93 ± 5.58 a | 6.08, ns |
| Ur (NH4N mg g−1) | 0.33 ± 0.05 a | 0.46 ± 0.11 a | 0.37 ± 0.06 a | 0.32 ± 0.05 a | 5.97, ns |
| Pb (mg kg−1) | 19.09 ± 2.25 a | 19.29 ± 1.54 a | 20.76 ± 3.59 a | 21.25 ± 2.30 a | 2.48, ns |
| Cu (mg kg−1) | 32.00 ± 0.48 ab | 35.13 ± 0.80 a | 29.37 ± 0.92 b | 28.51 ± 1.99 b | 9.05, ** |
| Zn (mg kg−1) | 89.1 ± 1.71 ab | 94.37 ± 1.49 a | 83.16 ± 2.06 b | 80.83 ± 4.73 b | 9.46, ** |
| 2018 | |||||
| pH | 8.97 ± 0.04 a | 9.06 ± 0.11 a | 8.77 ± 0.10 a | 8.91 ± 0.12 a | 6.73, ns |
| Ca (g kg−1) | 45.40 ± 1.55 ab | 48.25 ± 2.19 a | 36.57 ± 0.60 b | 34.55 ± 4.26 b | 9.51, ** |
| Na (g kg−1) | 11.52 ± 0.18 b | 11.82 ± 0.54 ab | 12.56 ± 0.21 a | 12.81 ± 0.96 a | 7.62, * |
| SIC (g kg−1) | 12.29 ± 0.22 ab | 12.69 ± 0.38 a | 10.67 ± 0.74 c | 11.18 ± 0.78 bc | 8.95, ** |
| SOC (g kg−1) | 4.69 ± 1.79 a | 4.66 ± 1.05 a | 6.50 ± 2.29 a | 5.34 ± 1.49 a | 1.36, ns |
| TN (g kg−1) | 0.80 ± 0.06 a | 0.65 ± 0.10 a | 0.80 ± 0.21 a | 0.68 ± 0.15 a | 2.74, ns |
| C/N | 5.84 ± 2.13 a | 7.13 ± 0.59 a | 7.98 ± 0.83 a | 7.76 ± 0.69 a | 4.33, ns |
| NH4N (mg kg−1) | 3.29 ± 0.80 ab | 2.56 ± 0.38 b | 5.39 ± 1.75 a | 1.84 ± 1.10 b | 7.40, * |
| NO3N (mg kg−1) | 4.06 ± 1.74 a | 15.61 ± 4.09 a | 12.02 ± 4.16 a | 15.45 ± 4.11 a | 6.90, ns |
| Beta (PNP mg g−1) | 40.45 ± 5.18 ab | 25.12 ± 0.62 b | 59.23 ± 25.91 a | 24.37 ± 0.90 b | 9.05, ** |
| Ur (NH4N mg g−1) | 0.43 ± 0.07 a | 0.50 ± 0.20 a | 0.36 ± 0.07 a | 0.34 ± 0.13 a | 2.69, ns |
| Pb (mg kg−1) | 8.36 ± 2.01 a | 10.64 ± 2.75 a | 17.15 ± 8.17 a | 13.36 ± 4.70 a | 5.36, ns |
| Cu (mg kg−1) | 29.20 ± 0.82 a | 28.48 ± 2.97 a | 26.88 ± 0.88 a | 25.87 ± 4.81 a | 2.59, ns |
| Zn (mg kg−1) | 78.57 ± 5.48 a | 79.98 ± 7.17 a | 79.25 ± 3.69 a | 79.08 ± 13.10 a | 0.54, ns |
Values in the same row followed by the same letter are not different (P > 0.05) according to a Kruskal–Wallis test; *, ** and *** are used to show statistical significance at the 0.05, 0.01, and 0.001 levels, respectively; ns, not significant.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, G.; He, T.; Zhang, M.; Wu, C. Could Land Abandonment with Human Intervention Benefit Cropland Restoration? From the Perspective of Soil Microbiota. Land 2021, 10, 1049. https://doi.org/10.3390/land10101049
AMA Style
Li G, He T, Zhang M, Wu C. Could Land Abandonment with Human Intervention Benefit Cropland Restoration? From the Perspective of Soil Microbiota. Land. 2021; 10(10):1049. https://doi.org/10.3390/land10101049
Chicago/Turabian StyleLi, Guangyu, Tingting He, Maoxin Zhang, and Cifang Wu. 2021. "Could Land Abandonment with Human Intervention Benefit Cropland Restoration? From the Perspective of Soil Microbiota" Land 10, no. 10: 1049. https://doi.org/10.3390/land10101049
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.