Distribution Patterns of the Freshwater Oligochaete Limnodrilus hoffmeisteri Influenced by Environmental Factors in Streams on a Korean Nationwide Scale



 ,
, 
Abstract
1. Introduction
2. Materials and Methods
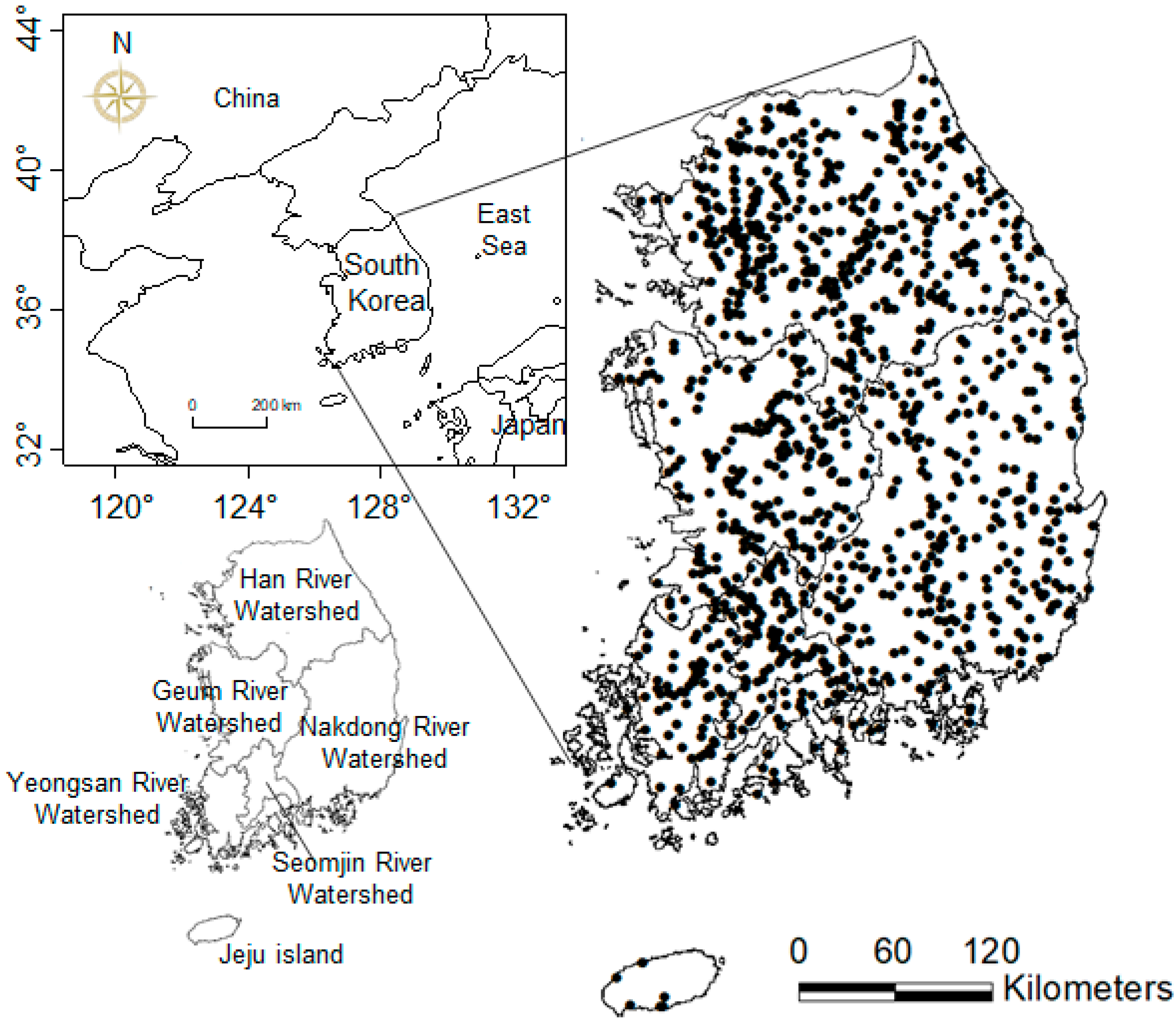
2.1. Ecological Data
2.2. Data Analyses
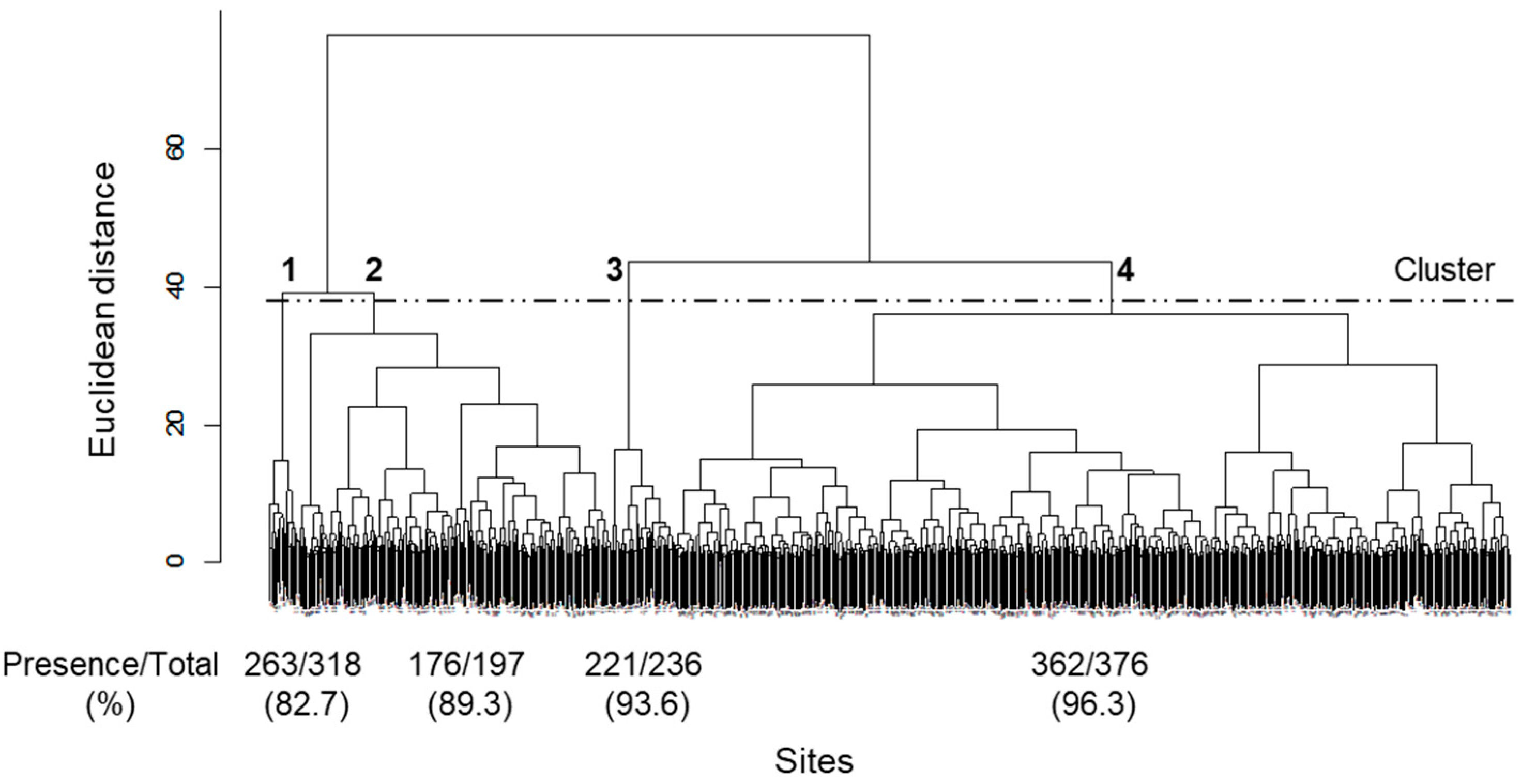
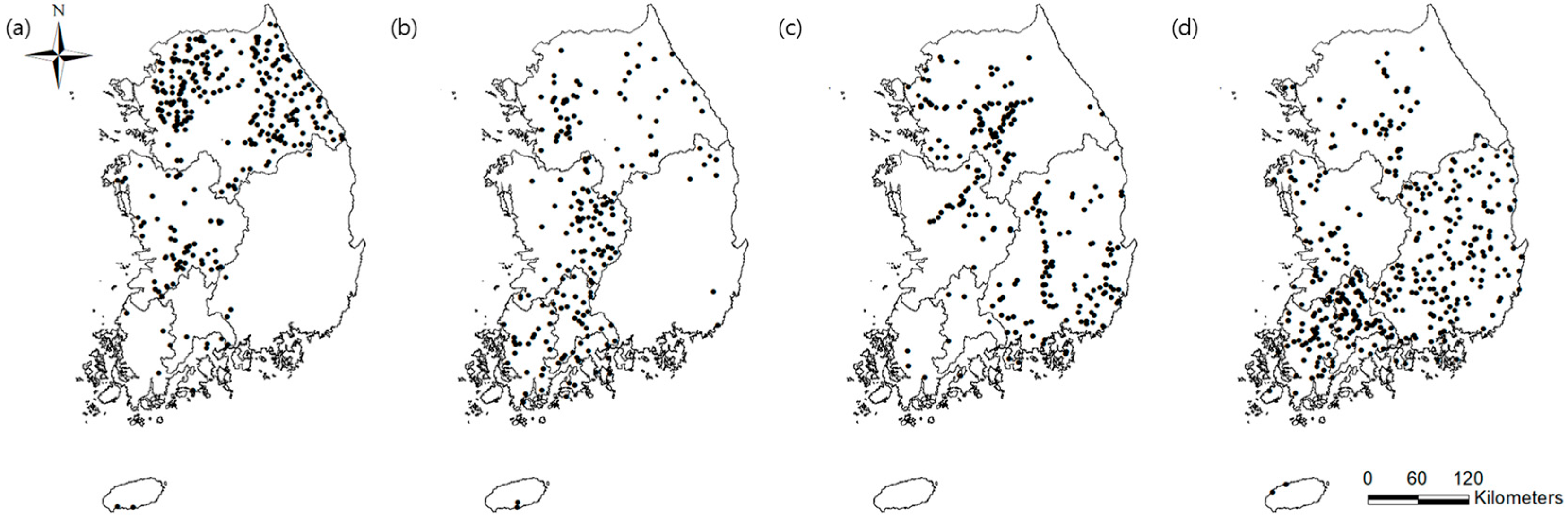
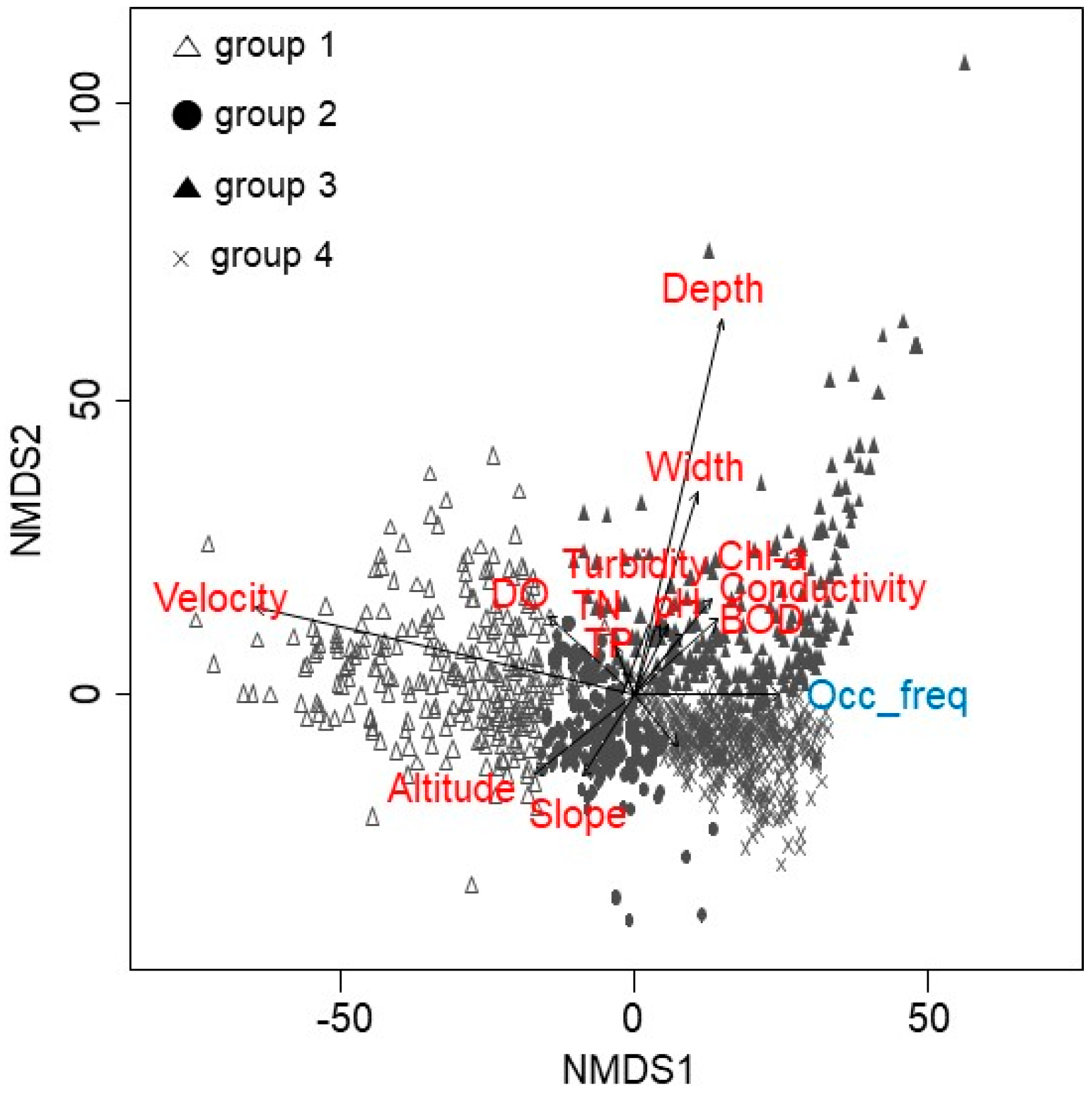
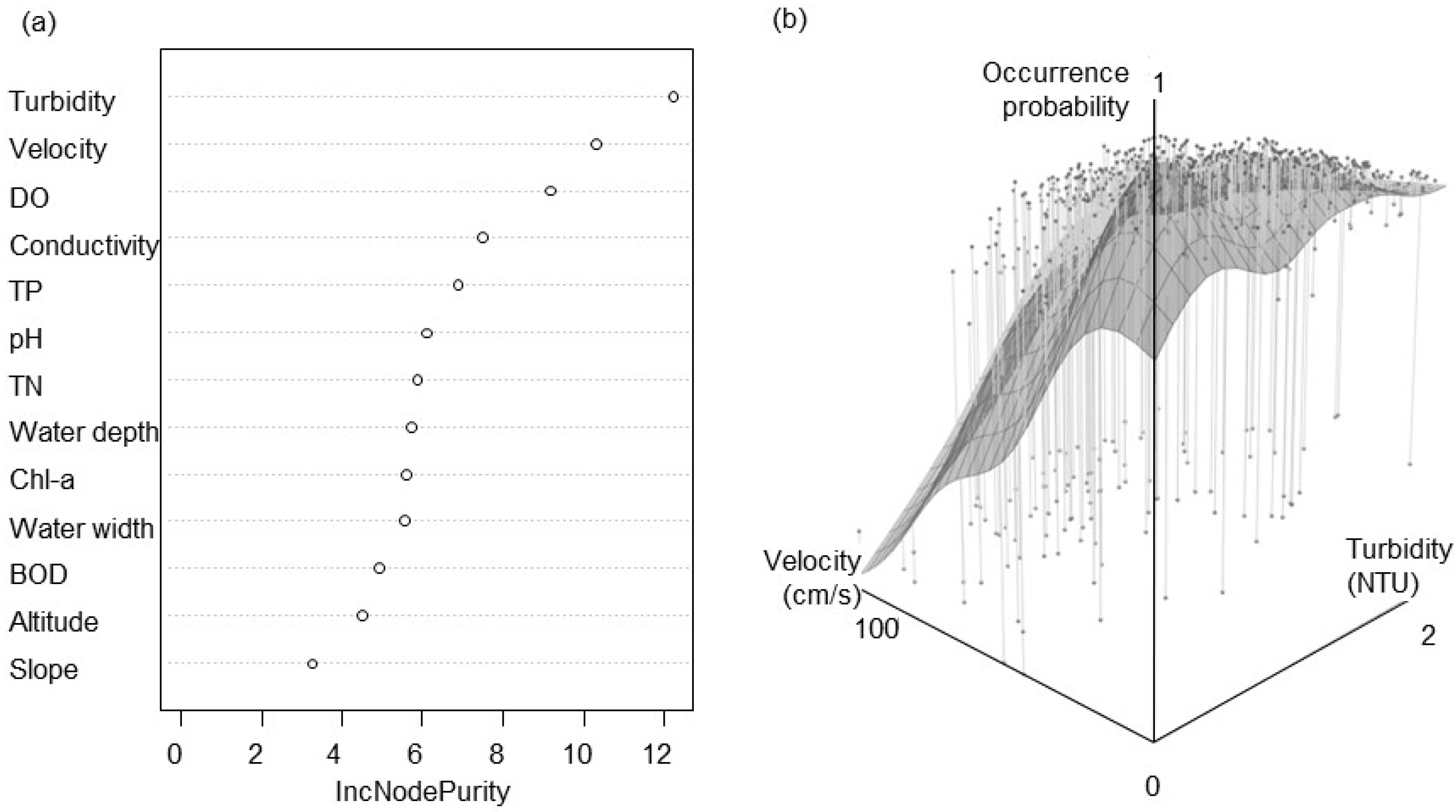
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bae, M.J.; Li, F.; Kwon, Y.S.; Chung, N.; Choi, H.; Hwang, S.J.; Park, Y.S. Concordance of diatom, macroinvertebrate and fish assemblages in streams at nested spatial scales: Implications for ecological integrity. Ecol. Indic. 2014, 47, 89–101. [Google Scholar] [CrossRef]
- Brinkhurst, R.O.; Kennedy, C.R. Studies on the biology of the Tubificidae (Annelida, Oligochaeta) in a polluted stream. J. Anim. Ecol. 1965, 34, 429–443. [Google Scholar] [CrossRef]
- Chapman, P.M.; Farrell, M.A.; rinkhurst, R.O. Relative tolerances of selected aquatic oligochaetes to individual pollutants and environmental factors. Aquat. Toxicol. 1982, 2, 47–67. [Google Scholar] [CrossRef]
- Lin, K.J.; Yo, S.P. The effect of organic pollution on the abundance and distribution of aquatic oligochaetes in an urban water basin, Taiwan. Hydrobiolgia 2008, 596, 213–223. [Google Scholar] [CrossRef]
- Korn, H. Studien zur Ökologie der Oligochaeten in der oberen Donau unter Berücksichtigung der Abwasserwirkungen. Archiv Für Hydrobiol. 1963, 27, 131–182. (In German) [Google Scholar]
- Prenda, J.; Gallardo, A. The influence of environmental factors and microhabitat availability on the distribution of an aquatic oligochaete assemblage in a Mediterranean river basin. Int. Rev. Hydrobiol. 1992, 77, 421–434. [Google Scholar] [CrossRef]
- Verdonschot, P.F.M. Hydrology and substrates: Determinants of oligochaete distribution in lowland streams (The Netherlands). Hydrobiologia 2001, 463, 249–262. [Google Scholar] [CrossRef]
- Schenková, J.; Helešic, J. Habitat preferences of aquatic oligochaeta (Annelida) in the Rokytná River, Czech Republic—A small highland stream. Hydrobiologia 2006, 564, 117–126. [Google Scholar] [CrossRef]
- Pascar-Gluzman, C.; Dimentman, C. Distribution and habitat characteristics of Naididae and Tubificidae in the inland waters of Israel and the Sinai Peninsula. Hydrobiologia 1984, 115, 197–205. [Google Scholar] [CrossRef]
- Fischer, J.A.; Beeton, A.M. The effect of dissolved oxygen on the burrowing behavior of Limnodrilus hoffmeisteri (Oligochaeta). Hydrobiologia 1975, 47, 273–290. [Google Scholar] [CrossRef]
- Van Duinen, G.A.; Timm, T.; Smolders, A.J.; Brock, A.M.; Verberk, W.C.; Esselink, H. Differential response of aquatic oligochaete species to increased nutrient availability—A comparative study between Estonian and Dutch raised bogs. Hydrobiologia 2006, 564, 143–155. [Google Scholar] [CrossRef]
- Martínez-Ansemil, E.; Collado, R. Distribution patterns of aquatic oligochaetes inhabiting watercourses in the Northwestern Iberian Peninsula. Hydrobiologia 1999, 334, 73–83. [Google Scholar] [CrossRef]
- Dumnicka, E. Communities of oligochaetes in mountain streams of Poland. Hydrobiologia 1994, 278, 107–110. [Google Scholar] [CrossRef]
- Sauter, G.; Güde, H. Influence of grain size on the distribution of tubificid oligochaete species. Hydrobiologia 1996, 334, 97–101. [Google Scholar] [CrossRef]
- Syrovátka, V.; Schenkova, J.; Brabec, K. The distribution of chironomid larvae and oligochaetes within a stony-bottomed river stretch: The role of substrate and hydraulic characteristics. Fundam. Appl. Limnol. Archiv Fur Hydrobiol. 2009, 174, 43–62. [Google Scholar] [CrossRef]
- Ratsak, C.H.; Verkuijlen, J. Sludge reduction by predatory activity of aquatic oligochaetes in waste water treatment plants: Science or fiction? A review. Hydrobiologia 2006, 564, 197–211. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Nogaro, G.; Datry, T.; Malard, F.; Gibert, J. Do tubificid worms influence the fate of organic matter and pollutants in stormwater sediments? Environ. Pollut. 2005, 134, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Pelegrí, S.P.; Blackburn, T.H. Effects of Tubifex tubifex (Oligochaeta: Tubificidae) on N-mineralization in freshwater sediments, measured with 15N isotopes. Aquat. Microb. Ecol. 1995, 9, 289–294. [Google Scholar] [CrossRef]
- Rensink, J.H.; Rulkens, W.H. Using metazoa to reduce sludge production. Water Sci. Technol. 1997, 36, 171–179. [Google Scholar]
- Elissen, H.J.H.; Mulder, W.J.; Hendrickx, T.L.G.; Elbersen, H.W.; Beelen, B.; Temmink, H.; Buisman, C.J.N. Aquatic worms grown on biosolids: Biomass composition and potential applications. Bioresour. Technol. 2010, 101, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H. The Blood Worm (Limnodrilus socialis) Using Capacity for Treatment of Aquaculture Wastewater. Master’s Thesis, Chungju University, Chungju-si, Korea, 2005. [Google Scholar]
- Jun, T.S.; Park, J.H. The blood worm, Limnodrilus socialis’s using capacity for treatment of aquaculture wastewater. Chungju Univ. Theses Collect. 2005, 40, 201–206. [Google Scholar]
- Macedo-Sousa, J.; Gerhardt, A.; Brett, C.M.A.; Nogueira, A.; Soares, A.M.V.M. Behavioural responses of indigenous benthic invertebrates (Echinogammarus meridionalis, Hydropsyche pellucidula and Choroterpes picteti) to a pulse of acid mine drainage: A laboratorial study. Environ. Pollut. 2008, 156, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Dermott, R.; Munawar, M. A simple and sensitive assay for evaluation of sediment toxicity using Lumbriculus variegatus (Muller). In Sediment/Water Interactions; Springer: Dordrecht, The Netherlands, 1992; pp. 407–414. [Google Scholar]
- Phipps, G.L.; Ankley, G.T.; Benoit, D.A. Use of the aquatic oligochaeta Lumbriculus variegatus for assessing the toxicity and bioaccumulation of sediment-associated contaminants. Environ. Toxicol. Chem. 1993, 12, 269–279. [Google Scholar] [CrossRef]
- Ingersoll, C.G.; Brunson, E.L.; Wang, N.; Dwyer, F.J.; Ankley, G.T.; Mount, D.R.; Huckins, J.N.; Petty, J.; Landrum, P.F. Uptake and depuration of nonionic organic contaminants from sediment by the oligochaete, Lumbriculus variegatus. Environ. Toxicol. Chem. 2003, 22, 872–885. [Google Scholar] [CrossRef] [PubMed]
- Lucan-Bouché, M.L.; Biagianti-Risbourg, S.; Arsac, F.; Vernet, G. An original decontamination process developed by the aquatic oligochaete Tubifex tubifex exposed to copper and lead. Aquat. Toxicol. 1999, 45, 9–17. [Google Scholar] [CrossRef]
- Rathore, R.S.; Khangarot, B.S. Effects of temperature on the sensitivity of sludge worm Tubifex tubifex Müller to selected heavy metals. Ecotoxicol. Environ. Saf. 2002, 53, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Bae, M.-J.; Park, Y.-S. Ecotoxicological studies using aquatic oligochaetes: Review. Korean J. Ecol. Environ. 2016, 49, 343–353. [Google Scholar] [CrossRef]
- Lee, J.; Jung, J. Four unrecorded species of tubificid oligochaetes (Annelida: Clitellata) in Korea. Anim. Syst. Evol. Divers. 2014, 30, 240–247. [Google Scholar] [CrossRef]
- Park, H.J.; Timm, T.; Bae, Y.J. Aquatic oligochaete (Annelida: Clitellata) fauna from the Jungnang Stream in Seoul, Korea, with eight new Korean records. Korean J. Ecol. Environ. 2013, 46, 507–512. [Google Scholar]
- Park, H.J.; Timm, T.; Bae, Y.J. Taxonomy of the Korean freshwater Oligochaeta (Annelida) with eight species new to Korea. Entomol. Res. Bull. 2013, 29, 180–188. [Google Scholar]
- Park, Y.-S.; Chang, J.; Lek, S.; Cao, W.; Brosse, S. Conservation strategies for endemic fish species threatened by the Three Gorges Dam. Conserv. Biol. 2003, 17, 1748–1758. [Google Scholar] [CrossRef]
- Park, Y.-S.; Tison, J.; Lek, S.; Coste, M.; Giraudel, J.; Delmas, F. Application of a self-organizing map in ecological informatics: Selection of representative species from large community dataset. Ecol. Inf. 2006, 1, 247–257. [Google Scholar] [CrossRef]
- Park, Y.-S.; Céréghino, R.; Compin, A.; Lek, S. Applications of artificial neural networks for patterning and predicting aquatic insect species richness in running waters. Ecol. Model. 2003, 160, 265–280. [Google Scholar] [CrossRef]
- Park, Y.-S.; Kwon, Y.-S.; Hwang, S.-J.; Park, S.-K. Characterizing effects of landscape and morphometric factors on water quality of reservoirs using a Self-organizing map. Environ. Model. Softw. 2014, 55, 214–221. [Google Scholar] [CrossRef]
- Peters, J.; De Baets, B.; Verhoest, N.E.; Samson, R.; Degroeve, S.; De Becker, P.; Huybrechts, W. Random forests as a tool for ecohydrological distribution modelling. Ecol. Model. 2007, 207, 304–318. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Thomaes, A.; Kervyn, T.; Maes, D. Applying species distribution modelling for the conservation of the threatened saproxylic Stag Beetle (Lucanus cervus). Biol. Conserv. 2008, 141, 1400–1410. [Google Scholar] [CrossRef]
- Bae, M.J.; Kwon, Y.; Hwang, S.J.; Chon, T.S.; Yang, H.J.; Kwak, I.S.; Park, J.H.; Ham, S.A.; Park, Y.S. Relationships between three major stream assemblages and their environmental factors in multiple spatial scales. Ann. Limnol.-Int. J. Limnol. 2011, 47, S91–S105. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Li, F.; Chung, N.; Bae, M.J.; Hwang, S.J.; Byoen, M.S.; Park, S.J.; Park, Y.S. Response of fish communities to various environmental variables across multiple spatial scales. Int. J. Environ. Res. Public Health 2012, 9, 3629–3653. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.M.; Iverson, L.R.; Liaw, A. Newer classification and regression tree techniques: Bagging and random forests for ecological prediction. Ecosystems 2006, 9, 181–199. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Environment (MOE); National Institute of Environmental Research (NIER). The Survey and Evaluation of Aquatic Ecosystem Health in Korea; NIER: Incheon, Korea, 2008; pp. 43–72.
- Brinkhurst, R.O.; Jamieson, B.G.M. Aquatic Oligochaeta of the World; Oliver and Body: Edinburgh, Scotland, 1971. [Google Scholar]
- Brinkhust, R.O. Guide to the Freshwater Aquatic Microdrile Oligachaetes of North America; Canadian Special Publication of Fisheries and Aquatic Sciences: Ottawa, ON, Canada, 1986; Volume 84, p. 259. [Google Scholar]
- Timm, T. Distribution of freshwater oligochaetes in the west and east coastal regions of the North Pacific Ocean. Hydrobiologia 1999, 406, 67–81. [Google Scholar] [CrossRef]
- Environmental Systems Research Incorporated (ESRI). ArcGIS 10.1; Environmental Systems Research Incorporated: Redlands, CA, USA, 2012; Available online: https://www.esri.com (accessed on 26 November 2017).
- Water Resource Management Information System (WAMIS). Seoul, Republic of Korea. Available online: http://www.wamis.go.kr (accessed on 3 March 2017).
- Legendre, P.; Legendre, L.F. Numerical Ecology, 3rd ed.; Elsevier: Oxford, UK, 2012; pp. 508–520. ISBN 9780444538680. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; ISBN 3-900051-07-0. Available online: https://www.r-project.org/ (accessed on 3 March 2017).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.4-4. 2017. Available online: https://CRAN.R-project.org/package=vegan (accessed on 24 April 2017).
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. Available online: https://cran.r-project.org/web/packages/randomForest/ (accessed on 7 June 2017).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 2nd ed.; SAGE: Thousand Oaks, CA, USA, 2011; Available online: https://cran.r-project.org/web/packages/car/index.html (accessed on 3 March 2017).
- Poff, N.L. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Lamouroux, N.; Dolédec, S.; Gayraud, S. Biological traits of stream macroinvertebrate communities: Effects of microhabitat, reach, and basin filters. J. N. Am. Benthol. Soc. 2004, 23, 449–466. [Google Scholar] [CrossRef]
- Timm, T.; Martin, P.J. Clitellata: Oligochaeta. In Freshwater Invertebrates, 4th ed.; Thorp, J., Rogers, D.C., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 529–549. ISBN 9780123850263. [Google Scholar]
- Avel, M.A.R.C.E.L. Classe des annélides oligochètes. Trait. De Zool. 1959, 5, 224–470. (In French) [Google Scholar]
- Håkanson, L. The relationship between salinity, suspended particulate matter and water clarity in aquatic systems. Ecol. Res. 2006, 21, 75–90. [Google Scholar] [CrossRef]
- Lazim, M.N.; Learner, M.A. The influence of sediment composition and leaf litter on the distribution of tubificid worms (Oligochaeta). Oecologia 1987, 72, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.; Martinez-Madrid, M.; Arrate, J.A.; Navarro, E. Selective feeding by the aquatic oligochaete Tubifex tubifex (Tubificidae, Clitellata). Hydrobiologia 2001, 463, 133–140. [Google Scholar] [CrossRef]
- Kang, H.; Bae, M.-J.; Park, Y.-S. Behavioral response of Tubifex tubifex to changes of water temperature and substrate composition. Korean J. Ecol. Environ. 2017, 50, 355–361. [Google Scholar]
- Marchand, J. The influence of seasonal salinity and turbidity maximum variations on the nursery function of the Loire estuary (France). Aquat. Ecol. 1993, 27, 427–436. [Google Scholar] [CrossRef]
- Bilotta, G.S.; Brazier, R.E. Understanding the influence of suspended solids on water quality and aquatic biota. Water Res. 2008, 42, 2849–2861. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.R. The life history of Limnodrilus hoffmeisteri Clap. (Oligochaeta: Tubificidae) and its adaptive significance. Oikos 1966, 17, 158–168. [Google Scholar] [CrossRef]
- Poddubnaya, T.L. Life cycles of mass species of Tubificidae (Oligochaeta). In Aquatic Oligochaete Biology; Springer: Boston, MA, USA, 1980; pp. 175–184. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Variable | Group | |||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||
| Geography | Altitude (m) | 183.64 (9.80) a | 140.96 (9.83) b | 69.12 (4.10) d | 99.69 (5.37) c |
| Slope (°) | 2.76 (0.35) a | 1.99 (0.40) a | 0.59 (0.13) b | 0.75 (0.11) b | |
| Hydrology | Water width (m) | 32.05 (1.89) c | 30.04 (2.93) c | 136.05 (11.12) a | 54.25 (5.81) b |
| Water depth (cm) | 26.15 (0.54) b | 22.97 (0.47) c | 49.53 (1.15) a | 25.51 (0.27) b | |
| Velocity (cm/s) | 64.97 (0.75) a | 37.38 (0.46) b | 18.26 (0.83) c | 14.95 (0.42) d | |
| Physicochemistry | Biochemical oxygen demand (BOD) (mg/L) | 1.62 (0.07) c | 1.70 (0.08) bc | 2.10 (0.07) a | 1.89 (0.05) ab |
| Total nitrogen (TN) (mg/L) | 2.83 (0.09) a | 2.52 (0.09) bc | 2.67 (0.07) ab | 2.40 (0.05) c | |
| Total phosphorus (TP) (mg/L) | 0.10 (0.01) | 0.10 (0.01) | 0.10 (0.01) | 0.08 (0.01) | |
| Chlorophyll a (Chl-a) (mg/L) | 2.65 (0.19) b | 3.17 (0.25) b | 5.05 (0.38) a | 3.26 (0.14) b | |
| Dissolved oxygen (DO) (mg/L) | 9.29 (0.08) a | 8.86 (0.08) b | 8.74 (0.09) b | 8.41 (0.06) c | |
| pH | 7.77 (0.02) b | 7.75 (0.03) b | 7.88 (0.03) a | 7.80 (0.02) ab | |
| Electric conductivity (μS/cm) | 195.05 (10.08) b | 239.53 (21.24) b | 764.06 (202.54) a | 285.68 (64.57) b | |
| Turbidity (Nephelometric Turbidity Units, NTU) | 11.67 (0.77) a | 7.74 (0.80) b | 12.64 (0.60) a | 13.02 (0.85) a | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.; Bae, M.-J.; Lee, D.-S.; Hwang, S.-J.; Moon, J.-S.; Park, Y.-S. Distribution Patterns of the Freshwater Oligochaete Limnodrilus hoffmeisteri Influenced by Environmental Factors in Streams on a Korean Nationwide Scale. Water 2017, 9, 921. https://doi.org/10.3390/w9120921
Kang H, Bae M-J, Lee D-S, Hwang S-J, Moon J-S, Park Y-S. Distribution Patterns of the Freshwater Oligochaete Limnodrilus hoffmeisteri Influenced by Environmental Factors in Streams on a Korean Nationwide Scale. Water. 2017; 9(12):921. https://doi.org/10.3390/w9120921
Chicago/Turabian StyleKang, Hyejin, Mi-Jung Bae, Dae-Seong Lee, Soon-Jin Hwang, Jeong-Suk Moon, and Young-Seuk Park. 2017. "Distribution Patterns of the Freshwater Oligochaete Limnodrilus hoffmeisteri Influenced by Environmental Factors in Streams on a Korean Nationwide Scale" Water 9, no. 12: 921. https://doi.org/10.3390/w9120921
APA StyleKang, H., Bae, M.-J., Lee, D.-S., Hwang, S.-J., Moon, J.-S., & Park, Y.-S. (2017). Distribution Patterns of the Freshwater Oligochaete Limnodrilus hoffmeisteri Influenced by Environmental Factors in Streams on a Korean Nationwide Scale. Water, 9(12), 921. https://doi.org/10.3390/w9120921