A Keystone Methylobacterium Strain in Biofilm Formation in Drinking Water


Abstract
1. Introduction
2. Materials and Methods
2.1. Methylobacterium Culture
2.2. Drinking Water Culture
2.3. Inoculation of Methylobacterium into Drinking Water
2.4. Stagnant Conditions
2.5. Flow Conditions
2.6. Statistical Analysis
2.7. Polymerase Chain Reaction
2.8. Characterisation of Aggregates Using FISH
3. Results
3.1. Influence of Methylobacterium on Aggregation under Stagnant Conditions
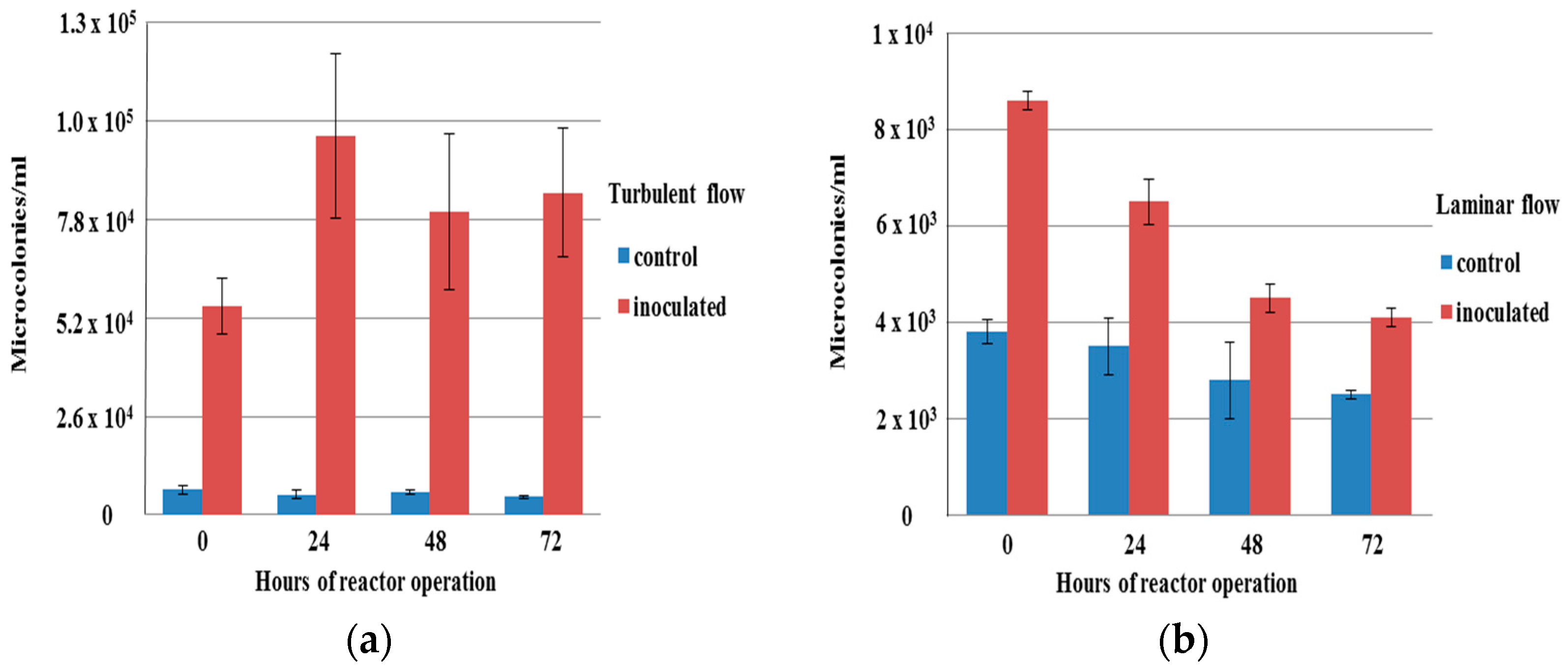
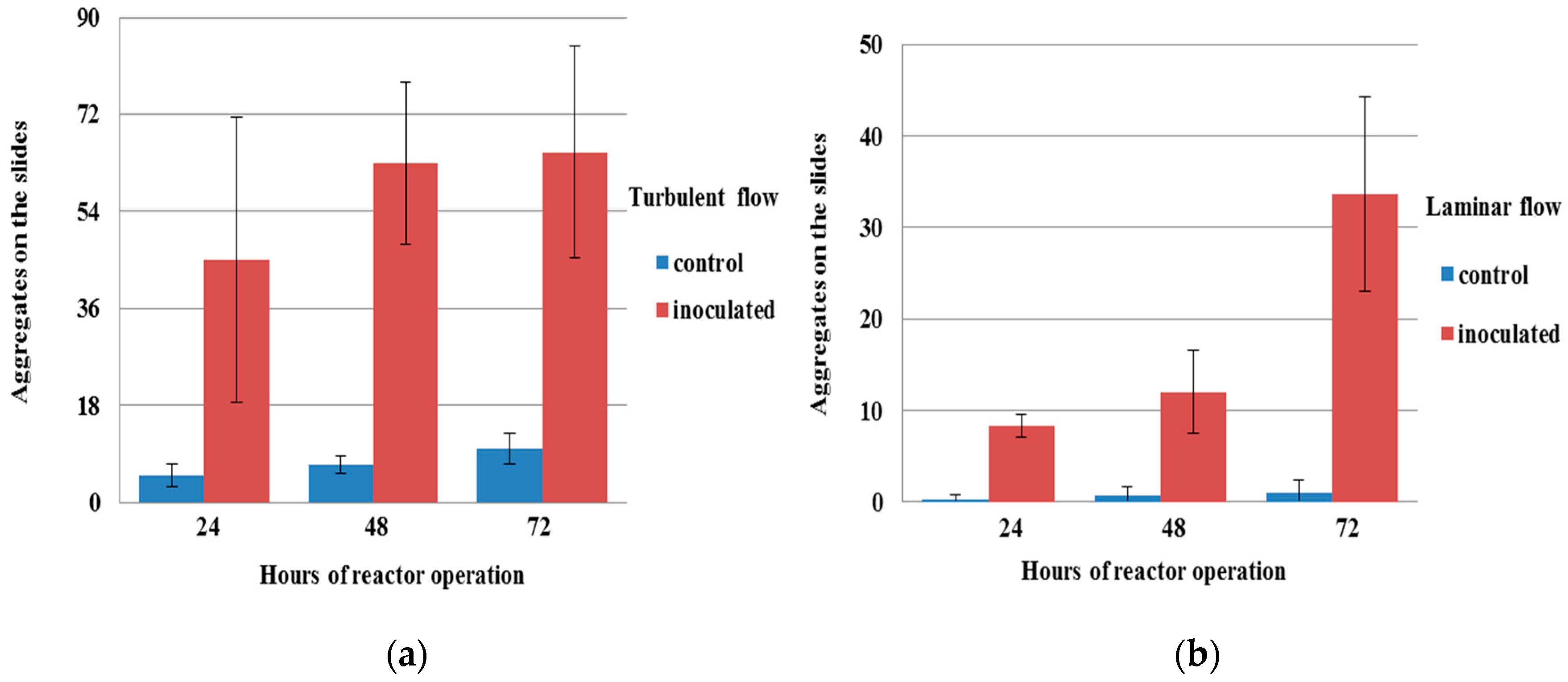
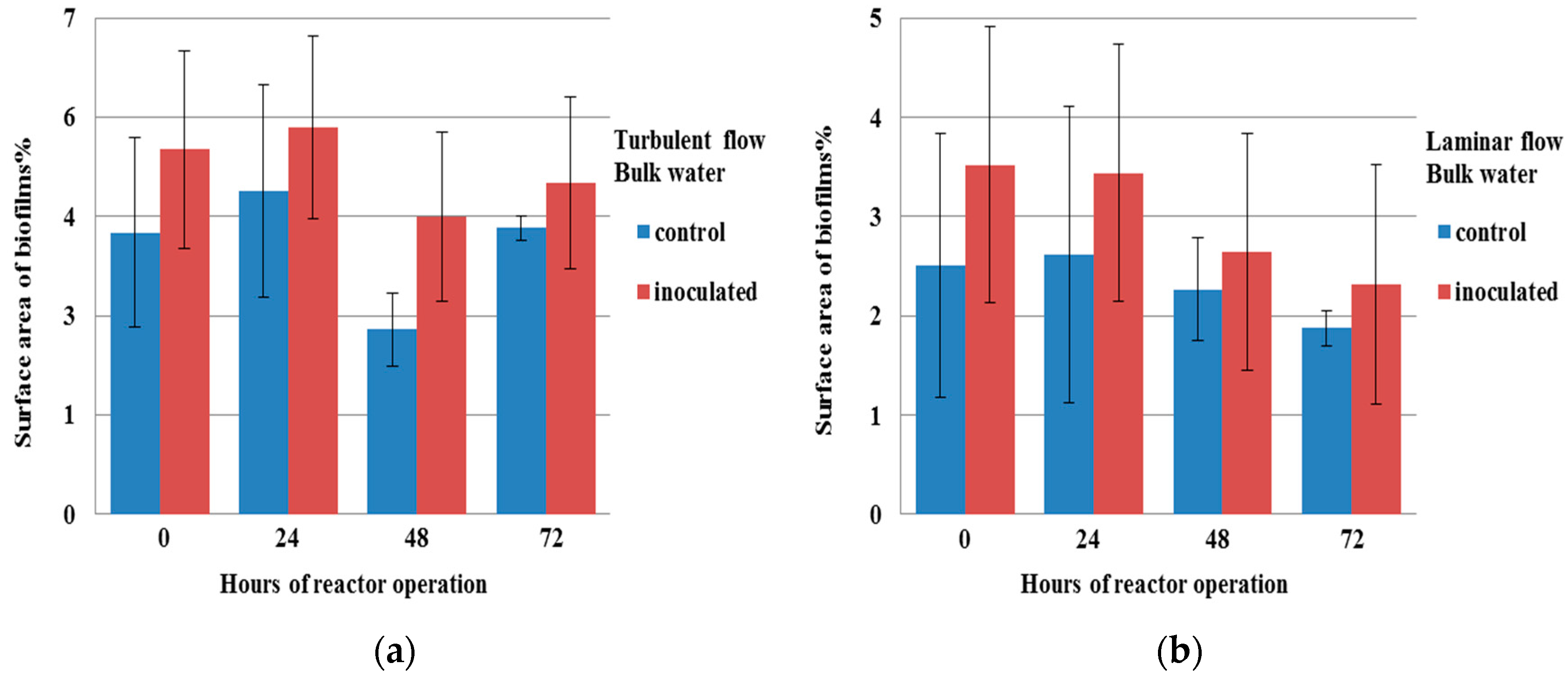
3.2. Influence of Methylobacterium on Aggregation under Flow Conditions
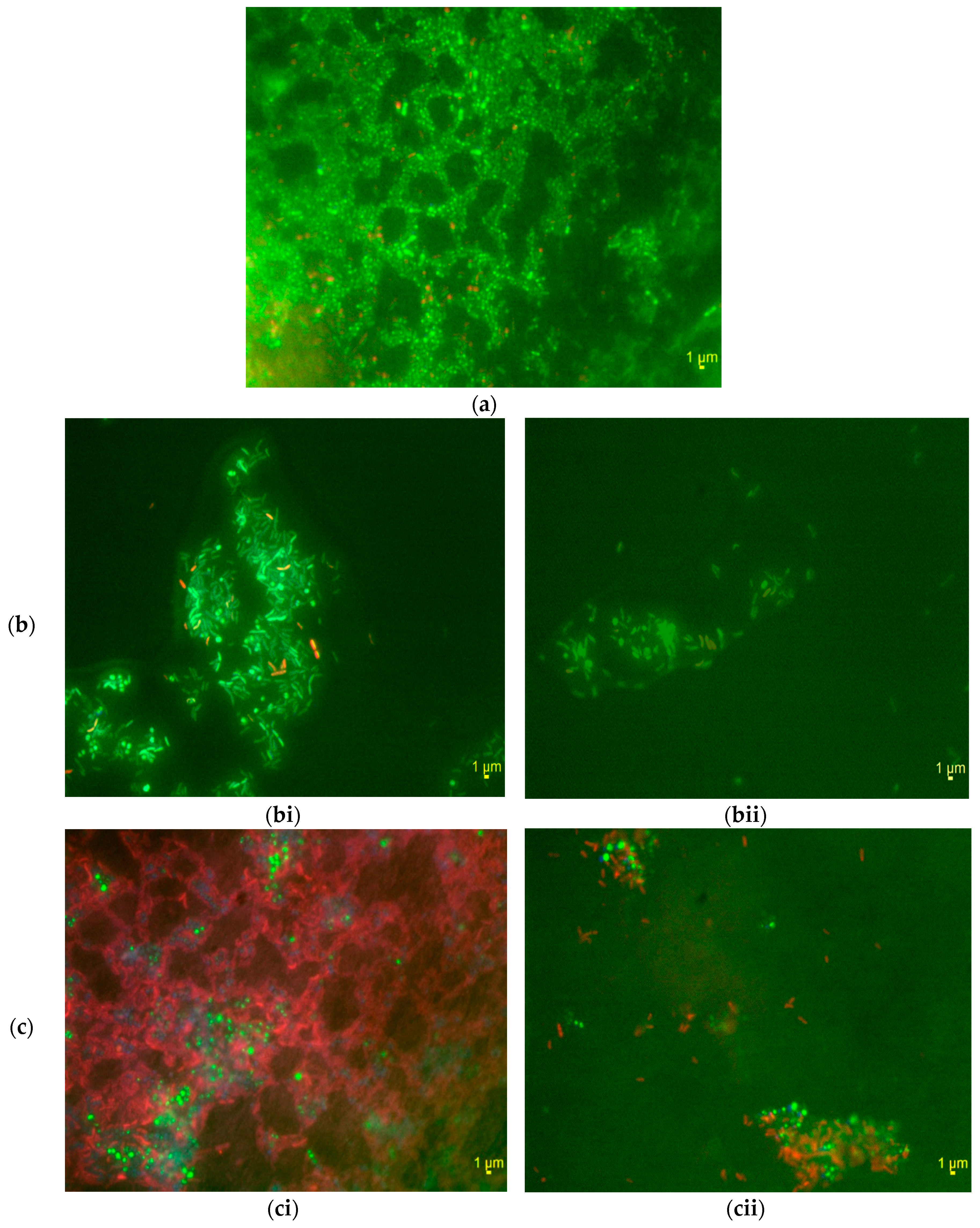
3.3. Structures of Aggregates
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, E.; Mukherjee, J.; Ramalingam, B.; Biggs, C.A. “Biofilmology”: A multidisciplinary review of the study of microbial biofilms. Appl. Microbiol. Biotechnol. 2011, 90, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; McBain, A.J.; Ledder, R.G.; Handley, P.S.; Gilbert, P. Coaggregation between freshwater bacteria within biofilm and planktonic communities. FEMS Microbiol. Lett. 2003, 220, 133–140. [Google Scholar] [CrossRef]
- Simões, L.C.; Simões, M.; Vieira, M.J. Intergeneric coaggregation among drinking water bacteria: Evidence of a role for Acinetobacter calcoaceticus as a bridging bacterium. Appl. Environ. Microbiol. 2008, 74, 1259–1263. [Google Scholar] [CrossRef] [PubMed]
- Sekar, R.; Deines, P.; Machell, J.; Osborn, A.M.; Biggs, C.A.; Boxall, J.B. Bacterial water quality and network hydraulic characteristics: A field study of a small, looped water distribution system using culture-independent molecular methods. J. Appl. Microbiol. 2012, 112, 1220–1234. [Google Scholar] [CrossRef] [PubMed]
- Simoes, L.C.; Simoes, M.; Oliveira, R.; Vieira, M.J. Potential of the adhesion of bacteria isolated from drinking water to materials. J. Basic Microbiol. 2007, 47, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.M.; Domingo, J.W.S.; Meckes, M.C.; Kelty, C.A.; Rochon, H.S. Phylogenetic diversity of drinking water bacteria in a distribution system simulator. J. Appl. Microbiol. 2004, 96, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Kormas, K.A.; Neofitou, C.; Pachiadaki, M.; Koufostathi, E. Changes of the bacterial assemblages throughout an urban drinking water distribution system. Environ. Monit. Assess. 2010, 165, 27–38. [Google Scholar] [CrossRef]
- Emtiazi, F.; Schwartz, T.; Marten, S.M.; Krolla-Sidenstein, P.; Obst, U. Investigation of natural biofilms formed during the production of drinking water from surface water embankment filtration. Water Res. 2004, 38, 1197–1206. [Google Scholar] [CrossRef]
- Martiny, A.C.; Jorgensen, T.M.; Albrechtsen, H.J.; Arvin, E.; Molin, S. Long-term succession of structure and diversity of a biofilm formed in a model drinking water distribution system. Appl. Environ. Microbiol. 2003, 69, 6899–6907. [Google Scholar] [CrossRef] [PubMed]
- Soge, O.O.; Giardino, M.A.; Ivanova, I.C.; Pearson, A.L.; Meschke, J.S.; Roberts, M.C. Low prevalence of antibiotic-resistant gram-negative bacteria isolated from rural south-western Ugandan groundwater. Water SA 2009, 35, 343–347. [Google Scholar] [CrossRef]
- Simões, L.C.; Simões, M.; Vieira, M.J. Biofilm interactions between distinct bacterial genera isolated from drinking water. Appl. Environ. Microbiol. 2007, 73, 6192–6200. [Google Scholar] [CrossRef] [PubMed]
- Scoaris, D.D.; Colacite, J.; Nakamura, C.V.; Ueda-Nakamura, T.; de Abreu, B.A.; Dias, B.P. Virulence and antibiotic susceptibility of Aeromonas spp. isolated from drinking water. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2008, 93, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; McBain, A.J.; Stead, A.T.; Gilbert, P. Shear rate moderates community diversity in freshwater biofilms. Appl. Environ. Microbiol. 2004, 70, 7426–7435. [Google Scholar] [CrossRef] [PubMed]
- Gallego, V.; Garcia, M.T.; Ventosa, A. Methylobacterium isbiliense sp. nov., isolated from the drinking water system of Sevilla, Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 2333–2337. [Google Scholar] [CrossRef] [PubMed]
- Gallego, V.; Garcia, M.T.; Ventosa, A. Methylobacterium variabile sp. nov., a methylotrophic bacterium isolated from an aquatic environment. Int. J. Syst. Evol. Microbiol. 2005, 55, 1429–1433. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O.; Williams, M.D.; Kwait, R.; Lande, L. Methylobacterium spp. as an indicator for the presence or absence of Mycobacterium spp. Int. J. Mycobacteriol. 2016, 5, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.W.; Martin, J.W.; Hooke, M.; Hooke, J. Methylobacterium mesophilicum infection: Case report and literature review of an unusual opportunistic pathogen. Clin. Infect. Dis. 2000, 30, 936–938. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.; Herrera-Rimann, K.; Blanc, D.S.; Greub, G. Biodiversity of amoebae and amoeba-resisting bacteria in a hospital water network. Appl. Environ. Microbiol. 2006, 72, 2428–2438. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; Leach, S.A.; Buswell, C.M.; High, N.J.; Handley, P.S. Coaggregation between aquatic bacteria is mediated by specific-growth-phase-dependent lectin-saccharide interactions. Appl. Environ. Microbiol. 2000, 66, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Dohner, H.; Stilgenbauer, S.; Benner, A.; Leupolt, E.; Krober, A.; Bullinger, L.; Dohner, K.; Bentz, M.; Lichter, P. Genomic aberrations and survival in chronic lymphocytic leukemia. N. Engl. J. Med. 2000, 343, 1910–1916. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Shu, W.; Li, P. Fluorescence In situ Hybridization: Cell-Based Genetic Diagnostic and Research Applications. Front. Cell Dev. Biol. 2016, 4, 89. [Google Scholar] [CrossRef] [PubMed]
- Douterelo, I.; Boxall, J.B.; Deines, P.; Sekar, R.; Fish, K.E.; Biggs, C.A. Methodological approaches for studying the microbial ecology of drinking water distribution systems. Water Res. 2014, 65, 134–156. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.L.; Klausen, C.; Nielsen, P.H.; Burford, M.; Jorgensen, N.O.G. Detection of activity among uncultured Actinobacteria in a drinking water reservoir. FEMS Microbiol. Ecol. 2006, 55, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Wilhartitz, I.; Mach, R.L.; Teira, E.; Reinthaler, T.; Herndl, G.J.; Farnleitner, A.H. Prokaryotic community analysis with CARD-FISH in comparison with FISH in ultra-oligotrophic ground- and drinking water. J. Appl. Microbiol. 2007, 103, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Deines, P.; Sekar, R.; Husband, P.S.; Boxall, J.B.; Osborn, A.M.; Biggs, C.A. A new coupon design for simultaneous analysis of in situ microbial biofilm formation and community structure in drinking water distribution systems. Appl. Microbiol. Biotechnol. 2010, 87, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, B. The Role of Cell to Cell Interactions and Quorum Sensing in Formation of Biofilms by Drinking Water Bacteria, in Chemical and Biological Engineering. Ph.D. Thesis, Univeristy of Sheffield, United Kingdom, Sheffield, UK, 2012. [Google Scholar]
- Kubota, K. CARD-FISH for Environmental Microorganisms: Technical Advancement and Future Applications. Microbes Environ. 2013, 28, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttila, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Yano, T.; Kubota, H.; Hanai, J.; Hitomi, J.; Tokuda, H. Stress Tolerance of Methylobacterium Biofilms in Bathrooms. Microbes Environ. 2013, 28, 87–95. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Biosafety Manual, 3rd ed.; World Health Organization (WHO) Laboratory: Geneva, Switzerland, 2004. [Google Scholar]
- Thiel, T. Streaking Microbial Cultures on Agar Plates, in Science in the Real World: Microbes in Action; Department of Biology, University of Missuri-St. Louis: St. Louis, MO, USA, 1999. [Google Scholar]
- Reasoner, D.J.; Geldreich, E.E. A New Medium for the Enumeration and Subculture of Bacteria from Potable Water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [PubMed]
- Sandle, T. An approach for the reporting of microbiological results from water systems. PDA J. Pharm. Sci. Technol. 2004, 58, 231–237. [Google Scholar] [PubMed]
- Kalmbach, S.; Manz, W.; Szewzyk, U. Isolation of new bacterial species from drinking water biofilms and proof of their in situ dominance with highly specific 16S rRNA probes. Appl. Environ. Microbiol. 1997, 63, 4164–4170. [Google Scholar] [PubMed]
- Gallego, V.; Garcia, M.T.; Ventosa, A. Methylobacterium hispanicum sp. nov and Methylobacterium aquaticum sp. nov., isolated from drinking water. Int. J. Syst. Evolut. Microbiol. 2005, 55, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Furuhata, K.; Matsumoto, A.; Koike, K.A.; Fukuyama, M.; Tabuchi, K. Phenotypic and Genetic Diversity of Chlorine-Resistant Methylobacterium Strains Isolated from Various Environments. Appl. Environ. Microbiol. 1995, 61, 2099–2107. [Google Scholar] [PubMed]
- Kepner, R.L.; Pratt, J.R. Use of Fluorochromes for Direct Enumeration of Total Bacteria in Environmental-Samples—Past and Present. Microbiol. Rev. 1994, 58, 603–615. [Google Scholar] [PubMed]
- Bautista-de Los Santos, Q.M.; Schroeder, J.L.; Blakemore, O.; Moses, J.; Haffey, M.; Sloan, W.; Pinto, A.J. The impact of sampling, PCR, and sequencing replication on discerning changes in drinking water bacterial community over diurnal time-scales. Water Res. 2016, 90, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, E.; Adams, C. Oxidation of sulfonamides, macrolides, and carbadox with free chlorine and monochloramine. Water Res. 2006, 40, 2517–2526. [Google Scholar] [CrossRef] [PubMed]
- Machell, J.; Boxall, J.; Saul, A.; Bramley, D. Improved Representation of Water Age in Distribution Networks to Inform Water Quality. J. Water Resour. Plan. Manag. ASCE 2009, 135, 382–391. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Miettinen, I.; Schaule, G. Surveillance and control of microbiological stability in drinking water distribution networks. Handbook for Analytical Methods and Operational Criteria for Biofilm Reactors. EU Project SAFER, Contract Number: EVK1-CT-2002-00108. 2003. Available online: http://www.oieau.org/eaudoc/system/files/documents/40/204680/204680_doc.pdf (accessed on 10 October 2017).
- Fuhrman, J.A. Microbial community structure and its functional implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Douterelo, I. Bacterial community dynamics during the early stages of biofilm formation in a chlorinated experimental drinking water distribution system: Implications for drinking water discolouration. J. Appl. Microbiol. 2014, 117, 286–301. [Google Scholar] [CrossRef] [PubMed]
- Douterelo, I.; Husband, S.; Boxall, J.B. The bacteriological composition of biomass recovered by flushing an operational drinking water distribution system. Water Res. 2014, 54, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, B.; Sekar, R.; Boxall, J.B.; Biggs, C.A. Aggregation and biofilm formation of bacteria isolated from domestic drinking water. Water Sci. Technol. Water Supply 2013, 13, 1016–1023. [Google Scholar] [CrossRef]
- Szabo, J.G.; Rice, E.W.; Bishop, P.L. Persistence and decontamination of Bacillus atrophaeus subsp. globigii spores on corroded iron in a model drinking water system. Appl. Environ. Microbiol. 2007, 73, 2451–2457. [Google Scholar] [CrossRef] [PubMed]
- Douterelo, I.; Sharpe, R.L.; Boxall, J.B. Influence of hydraulic regimes on bacterial community structure and composition in an experimental drinking water distribution system. Water Res. 2013, 47, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Childs, P.R.N. Rotating Cylinders, Annuli, and Spheres. In Rotating Flow; Elsevier Science & Technology Books: China, 2011; Chapter 6; Available online: http://www.homepages.ed.ac.uk/shs/Climatechange/Flettner%20ship/Childs%20Annuli.pdf (accessed on 10 October 2017).
- Bird, R.B.; Steward, E.W.; Lightfoot, E.N. Transport Phenomena; Wiley: New York, NY, USA, 1960; 780p. [Google Scholar]
- Kaye, J.; Elgar, E.C. Modes of Adiabatic and Diabatic Fluid Flow in an Annulus with an Inner Rotating Cylinder; M.I.T. Research Laboratory of Heat Transfer in Electronics: Cambridge, MA, USA, 1957; pp. 753–765. [Google Scholar]
- SAWater. Allowable Pipe Size, Class and Materials for Water Mains; South Australian (SA) Water Corporation: Adelaide, Australia, 2011. [Google Scholar]
- Chowdhury, S. Heterotrophic bacteria in drinking water distribution system: A review. Environ. Monit. Assess. 2012, 184, 6087–6137. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.Z.; He, C.G.; He, Q. Fate of free chlorine in drinking water during distribution in premise plumbing. Ecotoxicology 2015, 24, 2151–2155. [Google Scholar] [CrossRef] [PubMed]
- Garny, K.; Horn, H.; Neu, T.R. Interaction between biofilm development, structure and detachment in rotating annular reactors. Bioprocess. Biosyst. Eng. 2008, 31, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.Q.; Zhang, T. Optimization of fixation methods for observation of bacterial cell morphology and surface ultrastructures by atomic force microscopy. Appl. Microbiol. Biotechnol. 2011, 92, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Ragot, S.A.; Kertesz, M.A.; Bunemann, E.K. phoD Alkaline Phosphatase Gene Diversity in Soil. Appl. Environ. Microbiol. 2015, 81, 7281–7289. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, P.; Tyson, G.W.; Blackall, L.L. Design and evaluation of 16S rRNA-targeted oligonucleotide probes for fluorescence in situ hybridization. Methods Mol. Biol. 2002, 179, 29–42. [Google Scholar] [PubMed]
- Herndl, G. CARD-FISH and Microautoradiography Protocol for Bacteria and Archaea. 2007. Available online: http://www.microbial-oceanography.eu/pdf/MicroCardFish_Protocol.pdf (accessed on 10 October 2017).
- Pirttila, A.M.; Laukkanen, H.; Pospiech, H.; Myllyla, R.; Hohtola, A. Detection of intracellular bacteria in the buds of Scotch fine (Pinus sylvestris L.) by in situ hybridization. Appl. Environ. Microbiol. 2000, 66, 3073–3077. [Google Scholar] [CrossRef] [PubMed]
- Podolich, O.; Laschevskyy, V.; Ovcharenko, L.; Kozyrovska, N.; Pirttila, A.M. Methylobacterium sp. resides in unculturable state in potato tissues in vitro and becomes culturable after induction by Pseudomonas fluorescens IMGB163. J. Appl. Microbiol. 2009, 106, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Krumholz, L.; Stahl, D.A. Fluorescent-Oligonucleotide Probing of Whole Cells for Determinative, Phylogenetic, and Environmental-Studies in Microbiology. J. Bacteriol. 1990, 172, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Stoecker, K.; Dorninger, C.; Daims, H.; Wagner, M. Double labeling of oligonucleotide probes for fluorescence in situ hybridization (DOPE-FISH) improves signal intensity and increases rRNA accessibility. Appl. Environ. Microbiol. 2010, 76, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Barai, P.; Kumar, A.; Mukherjee, P.P. Modeling of Mesoscale Variability in Biofilm Shear Behavior. PLoS ONE 2016, 11, e0165593. [Google Scholar] [CrossRef] [PubMed]
- Duddu, R.; Chopp, D.L.; Moran, B. A Two-Dimensional Continuum Model of Biofilm Growth Incorporating Fluid Flow and Shear Stress Based Detachment. Biotechnol. Bioeng. 2009, 103, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Knapp, J.S.; Wales, D.S.; Edyvean, R.G.J. The effect of turbulent flow and surface roughness on biofilm formation in drinking water. J. Ind. Microbiol. Biotechnol. 1999, 22, 152–159. [Google Scholar] [CrossRef]
- Manuel, C.M.; Nunes, O.C.; Melo, L.F. Dynamics of drinking water biofilm in flow/non-flow conditions. Water Res. 2007, 41, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Harshey, R.M. Bacterial motility on a surface: Many ways to a common goal. Ann. Rev. Microbiol. 2003, 57, 249–273. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gunawan, C.; Barraud, N.; Rice, S.A.; Harry, E.J.; Amal, R. Understanding, Monitoring, and Controlling Biofilm Growth in Drinking Water Distribution Systems. Environ. Sci. Technol. 2016, 50, 8954–8976. [Google Scholar] [CrossRef] [PubMed]
- Saur, T.; Morin, E.; Habouzit, F.; Bernet, N.; Escudie, R. Impact of wall shear stress on initial bacterial adhesion in rotating annular reactor. PLoS ONE 2017, 12, e0172113. [Google Scholar] [CrossRef] [PubMed]
- Bos, R.; Vandermei, H.C.; Busscher, H.J. A Quantitative Method to Study Co-Adhesion of Microorganisms in a Parallel-Plate Flow Chamber. 2. Analysis of the Kinetics of Co-Adhesion. J. Microbiol. Methods 1995, 23, 169–182. [Google Scholar] [CrossRef]
- Perni, S.; Jordan, S.J.; Andrew, P.W.; Shama, G. Biofilm development by Listeria innocua in turbulent flow regimes. Food Control 2006, 17, 875–883. [Google Scholar] [CrossRef]
- Brugnoni, L.I.; Cubitto, M.A.; Lozano, J.E. Role of shear stress on biofilm formation of Candida krusei in a rotating disk system. J. Food Eng. 2011, 102, 266–271. [Google Scholar] [CrossRef]
- Bos, R.; Vandermei, H.C.; Meinders, J.M.; Busscher, H.J. A Quantitative Method to Study Co-Adhesion of Microorganisms in a Parallel-Plate Flow Chamber—Basic Principles of the Analysis. J. Microbiol. Methods 1994, 20, 289–305. [Google Scholar] [CrossRef]
- Kolenbrander, P.E.; Palmer, R.J.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell-cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Purevdorj, B.; Costerton, J.W.; Stoodley, P. Influence of Hydrodynamics and Cell Signaling on the Structure and Behavior of Pseudomonas aeruginosa Biofilms. Appl. Environ. Microbiol. 2002, 68, 4457–4464. [Google Scholar] [CrossRef] [PubMed]
- Kirisits, M.J.; Margolis, J.J.; Purevdorj-Gage, B.L.; Vaughan, B.; Chopp, D.L.; Stoodley, P.; Parsek, M.R. Influence of the hydrodynamic environment on quorum sensing in Pseudomonas aeruginosa biofilms. J. Bacteriol. 2007, 189, 8357–8360. [Google Scholar] [CrossRef] [PubMed]
- Poonguzhali, S.; Madhaiyan, M.; Sa, T. Production of acyl-homoserine lactone quorum-sensing signals is widespread in gram-negative Methylobacterium. J. Microbiol. Biotechnol. 2007, 17, 226–233. [Google Scholar] [PubMed]
- Penalver, C.G.N.; Cantet, F.; Morin, D.; Haras, D.; Vorholt, J.A. A plasmid-borne truncated luxI homolog controls quorum-sensing systems and extracellular carbohydrate production in Methylobacterium extorquens AM1. J. Bacteriol. 2006, 188, 7321–7324. [Google Scholar] [CrossRef] [PubMed]
- Jischa, M. Transfer of Momentum, Heat and Mass in Turbulent Flows of Binary-Mixtures 1. Reynolds Equations and Transport-Equations. Warme Stoffubertrag.-Thermo Fluid Dyn. 1976, 9, 173–178. [Google Scholar] [CrossRef]
- Jaberi, F.A.; Colucci, P.J. Large eddy simulation of heat and mass transport in turbulent flows. Part 2: Scalar field. Int. J. Heat Mass Transf. 2003, 46, 1827–1840. [Google Scholar] [CrossRef]
- Jaberi, F.A.; Colucci, P.J. Large eddy simulation of heat and mass transport in turbulent flows. Part 1: Velocity field. Int. J. Heat Mass Transf. 2003, 46, 1811–1825. [Google Scholar] [CrossRef]
- Hughmark, G.A. Heat, Mass, and Momentum Transport with Turbulent-Flow in Smooth and Rough Pipe. Aiche J. 1975, 21, 1033–1035. [Google Scholar] [CrossRef]
- Hondzo, M.; Wuest, A. Do Microscopic Organisms Feel Turbulent Flows? Environ. Sci. Technol. 2009, 43, 764–768. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measures | 24 h | 48 h | 72 h |
|---|---|---|---|
| Size of aggregates (mm2) | |||
| C/I | 0.2/0.3 | 0.3/0.6 | 0.1/0.2 |
| Number of aggregates | |||
| C/I | 1/4 | 5/6 | 1/2 |
| Total Area of aggregates (mm2) | |||
| C/I | 0.2/1.2 | 1.5/3.6 | 0.1/0.4 |
| Aggregation scores | |||
| C/I | 1/2 | 2/3 | 2/3 |
| p < 0.05 = X | Control vs. Inoculated | ||
|---|---|---|---|
| p < 0.05 = Y | Turbulent vs. Laminar | ||
| Aggregation measure | microcolonies per mL | aggregates on slides | biofilms% from the bulk |
| Turbulent/Laminar | X/X | X/X | X/- |
| Control/Inoculated | Y/Y | Y/Y | Y/Y |
| p < 0.05 = X | Microcolonies per mL | Aggregates on Slides | Biofilms% from the Bulk | |
|---|---|---|---|---|
| 0–24 h | Turbulent C/I | /X | /X | |
| Laminar C/I | /X | |||
| 24–48 h | Turbulent C/I | /X | /X | X/X |
| Laminar C/I | /X | /X | X/X | |
| 48–72 h | Turbulent C/I | X/X | ||
| Laminar C/I | /X | |||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsagkari, E.; Keating, C.; Couto, J.M.; Sloan, W.T. A Keystone Methylobacterium Strain in Biofilm Formation in Drinking Water. Water 2017, 9, 778. https://doi.org/10.3390/w9100778
Tsagkari E, Keating C, Couto JM, Sloan WT. A Keystone Methylobacterium Strain in Biofilm Formation in Drinking Water. Water. 2017; 9(10):778. https://doi.org/10.3390/w9100778
Chicago/Turabian StyleTsagkari, Erifyli, Ciara Keating, Jillian M. Couto, and William T. Sloan. 2017. "A Keystone Methylobacterium Strain in Biofilm Formation in Drinking Water" Water 9, no. 10: 778. https://doi.org/10.3390/w9100778
APA StyleTsagkari, E., Keating, C., Couto, J. M., & Sloan, W. T. (2017). A Keystone Methylobacterium Strain in Biofilm Formation in Drinking Water. Water, 9(10), 778. https://doi.org/10.3390/w9100778