Abstract
The impact of small-stature alkali bulrush (Bolboschoenus maritimus) and two hydroperiod treatments (early season raised water level or ambient water level) on mosquito production and water quality was studied in replicate 0.09 ha free water surface (FWS) treatment wetlands. Following reconfiguration of a 1-ha constructed wetland into a system with six replicate wetlands, bulrush was planted on 0.5-m centers in three 5-m wide bands in each wetland in summer, 2012. Open water and the low density of emergent vegetation effectively limited mosquito production from the bands of B. maritimus in each wetland during summer and autumn of year one. After the autumnal senescence of the bulrush culms, water levels were raised in half of the wetlands during winter and early spring to enhance sinking of dead bulrush biomass to reduce harborage for mosquitoes. Macrophyte coverage continued to increase in both hydroperiod treatments during year two, but non-bulrush species proliferated and eventually overgrew B. maritimus. Immature mosquito abundance in dipper samples from wetlands in the raised water level treatment was greater than from wetlands in the constant water level treatment. During spring of year two, adult mosquito production was associated with volunteer vegetation in the center of the test cells and averaged 6–18 mosquitoes m−2·day−1, approximately twice that of the other treatment. Hydrological regime did not significantly affect water quality performance (removal of nitrogen, phosphorus and chemical oxygen demand) in the wetlands. Alkali bulrush can persist in shallow water (depth < 0.2 m), but did not persist in deeper zones (mean depth > 0.4 m) of the wetlands and after comparatively large stature grasses and cattails colonized the wetlands. Raised planting beds interspersed with zones of deeper water are recommended to facilitate persistence of alkali bulrush and to limit proliferation of superior competitors.
1. Introduction
Emergent vegetation in free water surface constructed wetlands fulfills important functions that enhance nutrient removal for wastewater treatment. Emergent vegetation can (i) create slack flow and reduce wind-driven mixing which enhance sedimentation; (ii) adsorb particulates; (iii) reduce variation in environmental conditions to promote nutrient removal; (iv) provide physical structure for the attachment of microbes and periphyton important for wastewater treatment; (v) store nutrients; (vi) oxygenate sediments surrounding roots; and (vii) create conditions (i.e., low dissolved oxygen concentration in the water column, provide organic carbon) that promote denitrification [1,2,3]. Emergent vegetation can also provide food and cover for wildlife. However, large (>2.5 m in height) emergent macrophytes planted commonly in constructed treatment wetlands can support large numbers of pestiferous and pathogen-transmitting mosquitoes, especially when the water undergoing treatment carries high concentrations of nutrients and organic matter [4,5,6]. Large-stature emergent macrophytes used frequently in treatment wetlands include bulrushes [Schoenoplectus acutus (G.H.E. Muhlenberg ex J. Bigelow), S. americanus (Persoon) Volkart ex Schinz and R. Keller, S. californicus (C.A. Meyer) Palla], cattail (Typha spp.) and common reed (Phragmites australis (Cav.) Trin. ex Steud.).
Dense stands of live plants, as well as dead plant biomass, can hinder mosquito control efforts by limiting the contact of current formulations of mosquito-specific biorational control agents with the water surface and the target immature mosquitoes [7,8]. Mats of decaying matter protect mosquito larvae from predators and also increase food resources for larval mosquitoes [9,10]. Failure to control mosquitoes in the aquatic stages often necessitates the use of chemical pesticides against the adult stage typically over a geographic area that is much larger than that circumscribed by the wetlands. When human development encroaches on wetlands with environmental conditions conducive for high mosquito production, weekly or semi-weekly applications of adulticides may be required to control mosquito populations [4,8]. If mosquito production cannot be controlled sufficiently by the application of mosquito control agents, then these large macrophytes require often expensive, time-consuming management protocols. In addition to lower management costs, species of emergent vegetation that can be managed by environmental manipulations will reduce the use and impact on non-target organisms of insecticides for mosquito control.
Bolboschoenus (= Schoenoplectus) maritimus (L.) Palla (Cyperaceae), alkali (or cosmopolitan) bulrush, is a widely distributed wetland species [11,12] that may be more amenable than large macrophytes for integrated mosquito management (IMM) strategies for some constructed wetlands. Alkali bulrush is a relatively short (<1.5 m high) bulrush and has been recommended for use in treatment wetlands [13]. A natural senescence in autumn and winter dormancy thin stands of alkali bulrush to create conditions less favorable for mosquito production during the spring when mosquito activity increases annually. Bolboschoenus maritimus is an important waterfowl food that produces large protein-rich achenes and carbohydrate-rich corms [14,15].
We examined the effects of the two hydrological regimens on the sustainability of the emergent plants, water quality performance, and reduction of mosquito populations in experimental wetlands planted with B. maritimus during a two-year study. During the winter between years one and two, water levels were increased in half of the replicate wetlands to enhance the sinking of decaying bulrush biomass. Ambient water levels for river water moving through wetland complex were maintained in the other replicate wetlands. The two hydrological regimes were predicted to differ in the potential to reduce the biomass of the natural annual dieback of B. maritimus and faster sinking of decaying bulrush biomass could result in significantly smaller mosquito populations during the period when arboviruses increase naturally in the bird reservoirs and vector mosquito populations. Fluctuating water levels to enhance removal of decaying bulrush also could eliminate the need to dry the wetlands and reduce costly vegetation management.
2. Materials and Methods
2.1. Study Site
A 1-ha wetland at the Prado Constructed Wetlands (33.9° N, 117.9° W) was reconfigured into six replicate wetland test cells (test cell length × width: 93 m × 9.8 m), an equalization wetland that supplied water to the six test cells and a receiving wetland (Figure 1a). The Prado Constructed Wetlands receive approximately 50% of the Santa Ana River flow and the primary functions of the wetlands are to reduce nitrate levels in river water destined for groundwater storage basins in Orange County, CA, USA [16], as well as to provide riparian and wetland habitats for wildlife. Each 0.09 m2 test cell had a 0.91-m wide cement weir box at the inflow and outflow. V-notched weir boards were placed into each inflow weir box. Discharge from the equalization wetland through the v-notched weirs was estimated using established relationships between the height of the hydraulic head and flow rate [17]. Weir boards were set so that hydraulic residence time in the test cells was 2–4 days for ambient summer flows in the Santa Ana River.

Figure 1.
(a) Aerial view of the test cells in January 2013; (b) a test cell wetland in October with bands of B. maritimus; (c) following the autumnal dieback of B. maritimus in November; and (d) after sinking of dead bulrush biomass in January 2013. The yellow arrows indicate flow into the equalization wetland and out of the receiving wetland.
Approximately 3800 B. maritimus culms were transplanted on 0.5-m centers into the wetlands in late July and August 2012. Bolboschoenus maritimus was derived from seeds collected from three populations in Riverside County and grown at the UC Riverside Aquatic and Vector Control Facility [18]. Three 5-m wide bands of B. maritimus were planted, approximately equidistantly between the inflow and outflow of each test cell.
Two hydroperiod treatments were applied to the wetlands. Water levels in half of the test cells were either maintained by ambient flows of the Santa Ana River through the Prado Wetlands (test cells 2, 4 and 6) or were increased during winter and early spring and then maintained by ambient river flow (test cells 1, 3 and 5). On average, mean water depth increased from 0.2 m in the vegetation zone nearest the inflow to 0.36 m in the vegetation zone nearest the outflow.
Senescence of the above-ground growth of B. maritimus established in the wetlands during year one began in late autumn. In December 2012, water levels in half of the test cells were increased to about 0.5 m depth in the vegetation zone nearest the outlet weir to promote the degradation of senescent S. maritimus. Water flow was decreased in January to accommodate renovation of wetlands upstream of the test cells; however, elevated water levels were maintained in half of the test cells. Water levels were lowered in late February to pre-December levels. The effects of the two hydrological regimens on the sustainability of the emergent plants, water quality performance and reduction of mosquito populations were assessed.
2.2. Vegetation
Plant species, stem density and height were assessed in November 2012 and July 2013 in thirty 0.09-m2 quadrats in each test cell. Ten locations in each band of vegetation were chosen using a random number generator. The height of up to five plants in each quadrat was measured from the substrate surface to the top of the plant.
During 2013, percent cover of each test cell by emergent vegetation was estimated monthly by two persons (W.W. and D.D.) and from photographs. A mean value for percent cover was calculated for each test cell. The estimates of percent cover for a particular test cell by the two investigators on each sampling date did not differ by >5%.
2.3. Water Quality Monitoring
Grab samples from the water column were taken monthly (November and December 2012; February, April through August 2013) at the inflow and near the outflow of each test cell, placed into 500 mL dark plastic bottles and transported to the laboratory on ice. Nitrogen (total, ammonium-, nitrate- and nitrite-nitrogen) and phosphorus (total) concentrations and chemical oxygen demand were analyzed colorimetrically using APHA protocols [19] with a Hach DR™ 2800 spectrophotometer (TNT Plus tests, Hach Chemical Co., Loveland, CO, USA). Water samples were analyzed within 3 h of collection. Total Kjeldahl nitrogen concentration was determined during June through August 2013.
Inflow rates, estimated from flow through the v-notched weirs, were combined with the temporally corresponding water quality constituent concentration data to calculate mass loading for the various water quality constituents, in kg·ha−1·day−1. Removal efficiencies (RE) for the various constituents were calculated as percent of inflow water quality constituent removed.
Multi-parameter water quality sensor arrays (ICM Water Analyzer, Perstorp Analytical, Wilsonville, OR, USA) were used monthly to measure temperature (°C), dissolved oxygen (DO, mg·L−1), pH and specific conductance (μS·cm−1) at a representative location in the center of a test cell in each of the two hydrological treatments during summer 2013. The sensors were positioned approximately 15 cm below the water surface. Physicochemical variables were recorded every 0.5 h and were measured for 3–4 days.
2.4. Mosquitoes and Invertebrates
Immature mosquitoes and other invertebrates were sampled by taking five 350-mL dipper samples within a 2-m radius at 12 locations (4 locations in each band of emergent vegetation) within each test cell. The composite samples from each location were concentrated using a concentrator cup (mesh opening = 148 μm), transferred into 50-mL plastic vials, preserved with 95% ethanol and transported to the laboratory for enumeration. Mosquitoes were identified using Meyer and Durso [20] and other invertebrates were identified using Merritt et al. [21].
Adult mosquito production was measured monthly from May through August 2013 using ten pyramidal emergence traps per test cell deployed for a four-day period. The pyramidal emergence traps sampled an area equal to 0.25 m2 and emerging insects were collected in a jar at the top of the trap [22]. Chironomid midges and other insects in the samples also were quantified. Mosquito, as well as other invertebrate, production was not assessed in 2012 because immature mosquitoes were not collected in surveys.
ANOVA or repeated-measures ANOVA was carried out using SYSTAT (Ver. 9) to discern the effects of hydroperiod regime on water quality performance, plant characteristics and invertebrate abundance in the test cells. Wilk’s lambda, a multivariate test, was computed when the number of within subject levels (date and date × treatment effects) was >2. Numbers of mosquitoes were ln (x + 1) transformed before analysis. Chironomid midge production was ln (x + 1) transformed prior to analysis. The statistical significance of differences of mosquito abundance or production on a particular date was analyzed using one-way ANOVA.
3. Results
3.1. Vegetation
Bolboschoenus maritimus culms expanded across the 5-m planting zones within the three months after planting, increasing from 4 culms m−2 to approximately 47 culms m−2 in November (Table 1; Figure S1). The large variation relative to the mean of culm density indicates that there were places in the planting zones that were not colonized or supported low densities of bulrush; nevertheless, three robust stands of bulrush were evident in each test cell (Figure 1b). The density and mean height of B. maritimus (weighted mean: 81.7 cm) did not differ significantly between the hydroperiod treatments or among the test cells (F tests, P > 0.05; Table S1). The height of bulrush plants tended to decrease towards the outflow of the test cells and differed significantly among the vegetation bands within test cells (Table S1).

Table 1.
Height and density of Bolboschoenus maritimus culms and Echinocloa crus-galli in 0.09-ha test cells subjected to two hydroperiod treatments.
A natural senescence and dieback of the bulrush biomass above the substrate began in late October and by November dead culms were present in the test cells (Figure 1c). Duckweed (Lemna minor L.) was present in the test cells during this period. By February, most of the dead bulrush sunk below the water surface, the duckweed disappeared and extensive open water was present in the test cells (Figure 1d). Test cells in both hydroperiod treatments were nearly devoid of emergent vegetation (Figure 2).
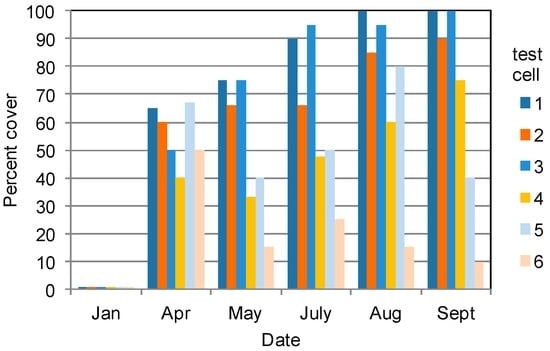
Figure 2.
Percent cover of the water surface by emergent vegetation in six 0.09 m2 test cell wetlands subjected to two hydroperiod treatments (blue histograms: early season water level increase; orange histograms: ambient water level in flow from the Santa Ana River) from January through September 2013.
Emergent vegetation increased markedly during spring. Even though the mean height of B. maritimus culms in both hydroperiod treatments continued to increase to about 122 cm, the density of B. maritimus declined by more than two-thirds to about 15 culms m−2 in July (Table 1). Barnyardgrass (Echinochloa crus-galli (L.) P. Beauv.) was dominant in four of the test cells. The percent of test cell surface area covered by emergent vegetation increased during year two in four of the test cells and either fluctuated or declined in the remaining two test cells (Figure 2). Barnyardgrass in the test cells in the raised water-level treatment was slightly taller than and, on average, twice as dense as in the ambient water-level treatment (Table 1). Grasses overgrew and replaced B. maritimus in many of the planting zones (Figures S2 and S3).
Other plants found in the test cells in July include Polygonum lapathifolium (L.) Gray (curlytop knotweed), Typha latifolia L. (cattail), Schoenolplectus americanus (Pers.) Volk. ex. Schinz and R. Keller (American three square), and S. californicus (C.A. Mey.) Palla (California bulrush). The abundance of curlytop knotweed was intermediate to the grass and the other plants and was present in 8% of the quadrats. Cattail and other large bulrush species were rare (≤2% of the quadrats) in July, but increased over time. Duckweed reappeared in May and completely covered the test cells by July.
3.2. Water Quality
Hydrological treatment did not significantly affect the removal efficiency of the test cells for any of the water quality constituents during the summer (Table S2). Tertiary-treated municipal effluent is the primary source of water in the Santa Ana River during the summer. Loading rates ranged seasonally between 3.5 to 10.2 kg N ha−1·day−1 and 0.8–2.4 kg P ha−1·day−1; the upper end of the range characterized the late spring and early summer loading rates into the test cells. Nitrate-nitrogen was the predominant form of nitrogen entering the test cells (Table 2). Nitrate-nitrogen removal efficiency ranged between 36% and 44% during late spring and summer. Whereas, the RE for various forms of nitrogen differed between the treatments, RE for total nitrogen was similar for test cells in both hydroperiod treatments during spring.
Table 2.
Comparison of water quality parameters (mg·L−1) in the inflow and outflow, and removal efficiencies (RE), of 0.09-ha test cells before, during and after hydroperiod treatments.
Phosphorus (P) removal declined in the test cells with raised water levels as compared to the other treatment during spring (Table 2). Removal efficiency for P was ≤6% in test cells in both hydrological treatments during the summer.
Duckweed and emergent vegetation coverage of the test cells during summer presumably moderated diel variation in temperature (range during July and August: 23 °C to 30.2 °C), pH (6.36 to 8.0) and DO (range: 0 to 2.4 mg·L−1). DO was >7 mg·L−1 when duckweed did not surround the sensor array, indicating that photosynthesis in the water column was greatly reduced in the test cells by L. minor. Mean specific conductance was approximately 1040 μS·cm−1.
3.3. Mosquitoes and Invertebrates
Immature mosquitoes were not collected between July and November 2012 when B. maritimus density was increasing following transplantation and open water was prevalent in the test cells.
Mosquito abundance increased in 2013 and differed between the two hydroperiod treatments across the experiment (F1,54 = 19.26, P < 0.001) and on three dates (May, August, November: Figure 3a). Significantly more immature Culex mosquitoes were collected from test cells in the raised-water-level treatment in May (F1,66 = 34.19, P < 0.001) and August (F1,66 = 7.29, P < 0.009) than from the ambient-water-level treatment. Culex tarsalis Coquillett dominated (81%) the collections of 3rd and 4th in stars and Cx. quinquefasciatus Say was about 11% of the late-instar larvae collected. Mosquito larvae were primarily associated with grasses that invaded the test cells. The mean number of mosquitoes in the raised-water-level treatment exceeded 1.2 larvae per composite sample (0.24 larva per dipper sample) on both dates but was ≤0.1 larva per sample in the ambient-water-level treatment. The difference in mosquito abundance between the hydroperiod treatments in November was not statistically significant (F1,50 = 3.33, P > 0.07). For both hydroperiod treatments, significantly more mosquitoes were collected from test cells 1, 2, 3, and 4 than from test cells 5 and 6 (F tests: test cells within treatment effect: P < 0.025). Anopheles hermsi Barr and Guptavanji larvae were present in the test cells but at very low abundance (<0.05 larva per dipper sample).
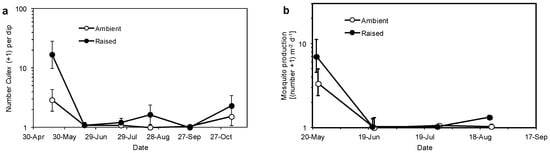
Figure 3.
(a) Abundance of immature mosquitoes in 350-mL dipper samples (backtransformed mean ± 95% CI) and (b) adult mosquito production (backtransformed mean ± 95% CI) from two hydroperiod treatments in 0.09-ha wetlands from May through November 2013.
Adult mosquito production from the raised-water-level treatment was about six individuals m−2·day−1 and was twice that of the ambient treatment during May (Figure 3b). Mosquito production from two replicate cells in the raised-water-level treatment exceeded 18 mosquitoes m−2·day−1 in the center band of vegetation in the test cells during May (F1,34 = 6.18; P < 0.02), whereas, fewer mosquitoes were collected from the bands of vegetation closer to the inflow and outflow of the test cells and no adult mosquitoes were collected from the open water in the third replicate test cell (treatment × position interaction: F4,54 = 3.63; P < 0.04). Mosquito production decreased during the summer and did not differ significantly between the treatments across the entire study (F1,12 = 1.72; P > 0.2).
The abundance of other invertebrate taxa did not differ significantly between the two hydroperiod treatments (RM ANOVAs, P > 0.05). Microcrustaceans (cladocerans, copepods, and ostracods) predominated (~85% of individuals) in the dipper samples. The predatory taxa were dominated by notonectids and zygopterans, but were comparatively rare (<4% individuals). The abundance of notonectids, corixids and mayflies (Callibaetis sp.) was greatest in May and declined across year two as coverage of emergent vegetation increased in the test cells. Amphipods (44%) and gastropods (23%) were numerically abundant among the herbivorous/detritivorous macroinvertebrates.
The temporal trend for chironomid midge production was similar to that observed for mosquito production. The greatest production occurred in May (~600 adults m−2·day−1) and declined to <10 individuals m−2·day−1 by August. During summer 2013, midge production did not differ between the hydroperiod treatments (F1,16 = 0.41, P > 0.5).
4. Discussion
Although alkali bulrush possesses many favorable characteristics for ease of management and for reducing mosquito production [18,23], B. maritimus did not persist in the renovated test cells due to a combination of factors, including the influx of other competitively superior macrophyte species and the effects of relatively deep (>0.4 m) water in places in the wetlands. In addition, the proliferation of duckweed mats covering the majority of the cells during the growing season of B. maritimus might have prevented light access to the new shoots.
Bolboschoenus maritimus tends to be a pioneering species that colonizes newly exposed soil and occurs at mid to low elevations in marshes, at wetland margins and in ephemeral wet spots [13]. It has wide pH tolerances, occurs commonly in alkaline and saline sites, and can tolerate fluctuating water levels, including total inundation up to 1 m in depth for brief periods [13,24]. Bolboschoenus maritimus can survive in a wide range of nitrogen loading rates [23,25] making it suitable for wastewater treatment applications. High levels of ammonium nitrogen (up to 60 mg NH4-N L−1) did not affect stem height and stem density; however, above- and below-ground biomass declined at high ammonium loading rates. Nitrogen uptake increased directly with enrichment across an enrichment gradient of <0.3 to 60 mg NH4-N L−1 [23] and, per unit mass, was comparable to other large macrophytes used in constructed treatment wetlands [3,23]. Comparatively small culm height at maturity (<1.5 m), shallow but well-developed rhizomes, pliable stems, an annual cycle that naturally reduces above-ground plant biomass, and dead stems that sink more rapidly than do those of taller related species (i.e., S. acutus, S. californicus which possess well-developed aerenchymatous tissue) are expected to reduce the cost of vegetation management and mosquito abatement as well as enhance the efficacy of ecologically-based mosquito control strategies that include hydroperiod manipulation.
Water levels and salinity are important determinants for the establishment and distribution of alkali bulrush [26,27,28,29,30]. Bolboschoenus maritimus established best in shallow (depth: 5–15 cm) water, but persisted in moderately deep water (30–40 cm) and was found in water as deep as 60 cm [27]. Negative effects of plant density (20 plants m−2) were observed in shallow water, yet dense stands of B. maritimus in moderately deep water were better able to withstand uprooting by wave action following transplantation [27]. Bolboschoenus maritimus dominated in managed wildlife marshes in the Western U.S., where soil submergence occurred for 3 to 11 months; 7 to 8 months of flooding provided optimum conditions for vegetative growth [28]. Alkali bulrush did not occur naturally in soils inundated for less than 60 days per year [28].
The competitive ability of B. maritimus against taller macrophytes (Typha angustifolia L. [28], P. australis [29,30]) declined when plants were continuously inundated with freshwater. In general, sedges are tolerant of anaerobic soils that limit the occurrence of some other macrophytes in continuously flooded habitats [31]. Bolboschoenus maritimus can persist in continuously flooded environments; however, rhizome biomass and aerial shoot length and biomass decline with longer periods of daily inundation in saline tidal environments [27,29,32]. Mean stem length of B. maritimus and two grass species increased directly with water depth [33]. Seed yield of alkali bulrush is greatest under moderately saline conditions (7‰–14‰ salinity [28]); salinity >15‰ caused high mortality in B. maritimus clones [28,34]. Bolboschoenus maritimus is adapted to sites with frequent changes of water levels and higher levels of salinity [26,27,35] better than to continuously flooded freshwater sites, such as the test cells where salinity is estimated to have been ≤0.5‰.
In the Prado Wetlands test cells, alkali bulrush expanded rapidly following transplantation but stem densities declined rapidly following colonization of competitively superior large emergent plant species in year two of our study. Bolboschoenus maritimus stem densities were approximately 50 stems m−2 three months after transplanting but stem density declined by 70% by the middle of following summer. Large-stature grasses rapidly colonized the test cells during the spring and summer after water levels were increased in half the test cells during winter. By July of year two, E. crus-galli dominated the emergent vegetation in the test cells of both hydroperiod treatments. Regardless of hydroperiod treatment, the grasses, cattail and large bulrushes overgrew and greatly reduced Bolboschoenus maritimus abundance in the test cells. The test cells had been used previously as a treatment wetland. Consequently, natural colonization of inundated soil by the large macrophytes was probably enhanced by seeds and rhizomes already present in the test cell soil. Given the similarity of the plant communities and flow regimes in the test cells, it is not surprising that removal efficiency of nutrients by the test cells did not differ between the two hydroperiod treatments.
Mosquito production was not detected during the summer and autumn of year one when B. maritimus was the only macrophyte present in the test cells but increased during year two. Mosquito production was concentrated in the center of the test cells where B. maritimus was replaced by grasses. The daily areal production of adult Culex and Anopheles mosquitoes from the grasses in four of the test cells (#1–4) in May was nearly two orders of magnitude greater than from the Prado Wetlands complex under standard operations. Assuming an annual mean between 0.07 and 0.093 adult mosquitoes m−2·day−1 are produced from the emergent vegetation [36,37], after correcting for emergence trap collection efficiency [38], the 142 ha Prado Wetlands produces between 7,600,000 and 10,120,000 female mosquitoes from July through September. These numbers reflect the low level of mosquito production per unit area, but the large total area of the constructed wetlands. Mosquito production per unit area was considerably greater in the grass-dominated test cells and can be even more substantial following some types of vegetation management. For example, before reconfiguration into the test cells, inundating clippings of S. californicus and S. americanus would have produced nearly 500,000,000 female mosquitoes from the 1-ha wetland during six weeks. Drying a subset of wetlands and then removing dried, downed bulrush with bulldozers before re-inundation effectively eliminates such high levels of mosquito production related to vegetation management.
Whereas, we did not detect mosquito production from monotypic stands of B. maritimus in year 1 of our study, other studies observed mosquito production from mesocosms containing alkali bulrush [18,23,39]. Relative to unenriched controls, the abundance of immature mosquitoes increased with ammonium nitrogen additions in mesocosms containing B. maritimus [23], suggesting that enrichment enhanced resources for larval mosquitoes. Despite a propensity of mosquitoes to lay more eggs on water within B. maritimus stands than within S. californicus stands, mosquito abundance in monotypic stands of B. maritimus was however less than in S. californicus [18,23]. These differences in mosquito abundance in stands of the two bulrushes were associated with greater abundance of mosquito predators in stands of B. maritimus [18,23,39]. Endophytic oviposition preferences of some predatory aquatic insects and the relatively greater structural complexity of B. maritimus might have been responsible for the increased invertebrate predator communities, especially nymphal odonate predators [18,39,40].
Managing vegetation in constructed treatment wetlands is controversial (see discussion in [41]) and can be expensive. Large macrophytes require thinning if water quality performance declines or if mosquito production reaches problematic levels [6,41,42,43]. Drying the wetland is necessary if heavy equipment is used to knock down and move dried vegetation. Maintaining water in the wetlands and harvesting emergent vegetation by aquatic harvesters can be expensive. For example, the cost of removal of vegetation from the periphery of the 6-ha constructed wetland at Valley Sanitary District (Indio, CA, USA) was $130,000 per harvest.
The approach to managing vegetation that was studied in the test cells is not applicable to all applications of free water surface constructed wetlands. Bolboschoenus maritimus has been used successfully for wetland restoration and water conservation in southern California in wetlands with discontinuous inundation (summer drying [44]) and with mixed success under continuous inundation in constructed treatment wetlands in Texas [25,45] and this study. Bolboschoenus maritimus is the most valuable plant resource for waterfowl in wetlands near the San Francisco Bay [28,46] and would constitute a valuable resource for waterfowl utilizing multipurpose constructed treatment wetlands. Under favorable conditions (shallow water depths, alkaline or slightly saline conditions), B. maritimus will produce dense vegetative growth that requires annual reduction [28,46]. Mowing with flail mowers is preferable to discing, bulldozing or burning to increase wildlife value [28]. Sinking decaying B. maritimus by manipulating water levels during cool periods of the year when mosquito activity is low or nonexistent is possible in shallow, comparatively alkaline conditions. The decaying bulrush can also provide organic carbon for denitrifying bacteria in wetlands polishing comparatively high-quality wastewater. After establishment, dense stands of B. maritimus are somewhat resistant to invasion by other macrophyte species [28,44]. Raised planting beds (hummocks) separated by deeper open water zones [41] is one wetland design that might facilitate proliferation of B. maritimus in new constructed wetlands and should be conducive to IMM strategies designed to reduce the use of chemical pesticides [6,42]. Bolboschoenus maritimus and closely related species (i.e., B. robustus (Pursh) M.T. Strong) generally prefer shallow (<0.30 m) water [14,24,27] and, depending on the configuration of the constructed treatment wetland, may not require the labor-intensive, costly methods used in large-scale wetlands to manage large emergent macrophytes.
Supplementary Materials
The following are available online at http://www.mdpi.com/2073-4441/8/10/421/s1.
Acknowledgments
Funding and in-kind assistance were provided by the Orange County Water District. This work also was supported by the Mosquito Research Foundation, Northwest Mosquito and Vector Control District, West Valley Mosquito and Vector Control District and U.S. Department of Agriculture National Institute of Food and Agriculture, Hatch project CA-R*-ENT-5029-H. Min Tao was supported by the International Cooperation Program of the China Scholarship Council. Jordan Greer, Tristan Hallum, Bonnie Johnson, Lynn Luu, Steven Merkley and Chris Wilson provided invaluable assistance with field and laboratory aspects of the study. We thank Andrew Sanders of the Herbarium at U.C. Riverside for assistance with identification of the plants. We benefitted from discussions with Peter Bowler, William Bretz, Michael Lopanec and Loretta Mokry.
Author Contributions
William E. Walton is the primary author for the study. William E. Walton, Dagne Duguma, David A. Popko, Min Tao and Scott Nygren designed the study, carried field and laboratory aspects of the study and contributed to writing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brix, H. Do macrophytes play a role in constructed wetland treatment wetlands? Water Sci. Technol. 1997, 35, 11–17. [Google Scholar] [CrossRef]
- Stottmeister, U.; Wießner, A.; Kuschk, P.; Kappelmeyer, U.; Kästner, M.; Bederski, O.; Müller, R.A.; Moormann, H. Effects of plants and microorganisms in constructed wetlands for wastewater treatment. Biotechnol. Adv. 2003, 22, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, R.H.; Wallace, S.D. Treatment Wetlands, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Walton, W.E. Multipurpose constructed treatment wetlands in the arid southwestern United States: Are the benefits worth the risks? In Treatment Wetlands for Water Quality Improvement, Proceedings of the Quebec 2000 Conference, Quebec City, QC, Canada, 6–12 August 2000; Pries, J., Ed.; Selected Papers. CH2M Hill Canada Limited, Pandora Press: Waterloo, ON, Canada, 2002; pp. 115–123. [Google Scholar]
- Walton, W.E. Managing Mosquitoes in Surface-Flow Constructed Treatment Wetlands; Publication Number 8117; Division of Agriculture and Natural Resources, University of California: Davis, CA, USA, 2003; Available online: http://anrcatalog.ucdavis.edu/pdf/8117.pdf (accessed on 20 August 2016).
- Walton, W.E. Design and management of free water surface constructed wetlands to minimize mosquito production. Wetl. Ecol. Manag. 2012, 20, 173–195. [Google Scholar] [CrossRef]
- Walton, W.E.; Popko, D.A.; Van Dam, A.R.; Merrill, A.; Lythgoe, J.; Hess, B. Width of planting beds for emergent vegetation influences mosquito production from a constructed wetland in California (USA). Ecol. Eng. 2012, 42, 150–159. [Google Scholar] [CrossRef]
- Walton, W.E.; Popko, D.A.; Van Dam, A.R.; Merrill, A. Distribution of Culex species in vegetation bands of a constructed wetland undergoing integrated mosquito management. J. Am. Mosq. Control Assoc. 2013, 29, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Berkelhamer, R.C.; Bradley, T.J. Mosquito larval development in container habitats: The role of rotting Scirpus californicus. J. Am. Mosq. Control Assoc. 1989, 5, 258–260. [Google Scholar] [PubMed]
- Walton, W.E.; Jiannino, J.A. Vegetation management to stimulate denitrification increases mosquito abundance in multipurpose constructed treatment wetlands. J. Am. Mosq. Control Assoc. 2005, 21, 22–27. [Google Scholar] [CrossRef]
- Smith, G.C.; Kukkonen, I. A new lectotype for Scirpus maritimus (Cyperaceae). Taxon 1999, 48, 355–357. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA); National Resources Conservation Service (NRCS). The PLANTS Database; National Plant Data Team: Greensboro, NC, USA, 2011. Available online: http://plants.usda.gov/java/profile?symbol=SCMA8 (accessed on 20 August 2016).
- Hoag, C. Plant Guide. Alkali Bulrush Schoenoplectus maritimus (L.) Lye; USDA-Natural Resources Conservation Service, Idaho Plant Materials Center: Aberdeen, ID, USA, 2000. Available online: http://plants.usda.gov/plantguide/pdf/pg_scma8.pdf (accessed on 20 August 2016).
- Kantrud, H.A. The Alkali (Scirpus maritimus L.) and Saltmarsh (S. robustus Pursh) Bulrushes: A Literature Review. Information and Technology Report 6; National Biological Service, Northern Prairie Wildlife Research Center: Jamestown, ND, USA, 1996. Available online: https://web.archive.org/web/20130902225308/http://www.npwrc.usgs.gov/resource/plants/bulrush (accessed on 24 August 2016). [Google Scholar]
- Miller, M.R.; Burns, E.G.; Wickland, B.E.; Eadie, J.M. Diet and body mass of wintering ducks in adjacent brackish and freshwater habitats. Waterbirds 2009, 32, 374–378. [Google Scholar] [CrossRef]
- Mills, W.R.; Bradford, S.M.; Rigby, M.; Wehner, M.P. Groundwater recharge at the Orange County Water District. In Wastewater Reclamation and Reuse; Asano, T., Ed.; Technomic Publishing Company: Lancaster, PA, USA, 1998; pp. 1105–1141. [Google Scholar]
- Skrenter, R.G. Instrumentation Handbook for Water and Wastewater Treatment Plants; ATSM D 5242-92; Lewis Publishers: Chelsea, MI, USA, 1988. [Google Scholar]
- Popko, D.A.; Walton, W.E. Small-stature emergent macrophytes and crepuscular sprinkler disturbance reduce mosquito abundance in wetland mesocosms. J. Vector Ecol. 2013, 38, 379–389. [Google Scholar] [CrossRef] [PubMed]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 19th ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Meyer, R.P.; Durso, S.L. Identification of the Mosquitoes of California; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 1998. [Google Scholar]
- Merritt, R.W.; Cummins, K.W.; Berg, M.B. An Introduction to the Aquatic Insects of North America, 4th ed.; Kendall Hunt Publishing Co.: Dubuque, IA, USA, 2008. [Google Scholar]
- Walton, W.E.; Workman, P.D.; Keiper, J.B. An inexpensive collapsible pyramidal emergence trap for the assessment of wetland insect populations. In Proceedings of the 67th Annual Conference of the Mosquito and Vector Control Association of California, Anaheim, CA, USA, 24–27 January 1999; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 1999; pp. 15–17. [Google Scholar]
- Duguma, D.; Walton, W.E. Effects of nutrients on mosquitoes and an emergent macrophyte, Schoenoplectus maritimus, for use in treatment wetlands. J. Vector Ecol. 2014, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Rafferty, P.S. Clonal variation in response to salinity and flooding stress in four marsh macrophytes of the northern gulf of Mexico, USA. Environ. Exp. Bot. 2006, 56, 301–313. [Google Scholar] [CrossRef]
- Noack, T.J.; Pierce, R.M. The swamps of Cactus: Year 1 lessons learned treating high-strength wastewater with a constructed wetland. In Proceedings of the Texas Water Conference, San Antonio, TX, USA, 10–13 April 2012.
- Deegan, B.; Harrington, T.J.; Dundon, P. Effects of salinity and inundation regime on growth and distribution of Schoenoplectus triqueter. Aquat. Bot. 2005, 81, 199–211. [Google Scholar] [CrossRef]
- Clevering, O.A.; van Gulik, W.M.G. Restoration of Scirpus lacustris and Scirpus maritimus stands in a former tidal area. Aquat. Bot. 1997, 55, 229–246. [Google Scholar] [CrossRef]
- Mall, R.E. Soil-Water-Salt Relationships of Waterfowl Food Plants in the Suisun Marsh of California; Wildlife Bulletin Number 1; California Department of Fish and Game: Sacramento, CA, USA, 1969.
- Lieffers, V.J.; Shay, J.M. The effects of water level on the growth and reproduction of Scirpus maritimus var. paludosus. Can. J. Bot. 1981, 59, 118–121. [Google Scholar] [CrossRef]
- Shay, J.M.; Shay, C.T. Prairie marshes in western Canada, with specific reference to the ecology of five emergent macrophytes. Can. J. Bot. 1986, 64, 443–454. [Google Scholar] [CrossRef]
- Crawford, R.M.M. Oxygen availability as an ecological limit to plant distribution. Adv. Ecol. Res. 1992, 23, 93–171. [Google Scholar]
- Clevering, O.A.; Hundscheid, M.P. Plastic and non-plastic variation in growth of newly established clones of Scirpus (Bolboschoenus) maritimus L. grown at different water depths. Aquat. Bot. 1998, 62, 1–17. [Google Scholar] [CrossRef]
- Coops, H.; van den Brink, F.W.B.; van der Velde, G. Growth and morphological responses of four helophyte species in an experimental water-depth gradient. Aquat. Bot. 1996, 54, 11–24. [Google Scholar] [CrossRef]
- Lillebø, A.I.; Pardal, M.A.; Neto, J.M.; Marques, J.C. Salinity as the major factor affecting Scirpus maritimus annual dynamics. Evidence from field data and greenhouse experiment. Aquat. Bot. 2003, 77, 111–120. [Google Scholar] [CrossRef]
- Clevering, O.A.; Blom, C.W.P.M.; van Vierssen, W. Growth and morphology of Scirpus lacustris and S. maritimus seedlings as affected by water level and light availability. Funct. Ecol. 1996, 10, 289–296. [Google Scholar] [CrossRef]
- Keiper, J.B.; Jiannino, J.A.; Beehler, J.; Walton, W.E. Distribution and abundance of Culicidae and Chironomidae (Diptera) following storm damage in a southern California constructed wetlands. In Proceedings of the 67th Annual Conference of the Mosquito and Vector Control Association of California, Anaheim, CA, USA, 24–27 January 1999; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 1999; pp. 47–54. [Google Scholar]
- Keiper, J.B.; Jiannino, J.A.; Sanford, M.R.; Walton, W.E. Effect of vegetation management on the abundance of mosquitoes at a constructed treatment wetland in southern California. In Proceedings of the 70th Annual Conference of the Mosquito and Vector Control Association of California, Fish Camp, CA, USA, 27–30 January 2002; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 2003; pp. 35–43. [Google Scholar]
- Walton, W.E. Effectiveness of pyramidal emergence traps for estimating production of Culex mosquitoes (Diptera: Culicidae). J. N. Am. Benthol. Soc. 2009, 28, 676–682. [Google Scholar] [CrossRef]
- Duguma, D.; Rugman-Jones, P.; Kaufman, M.G.; Hall, M.W.; Neufeld, J.D.; Stouthamer, R.; Walton, W.E. Bacterial communities associated with Culex mosquito larvae and two emergent aquatic plants of bioremediation importance. PLoS ONE 2013, 8, e72522. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa-Garro, D.; Mason, C.F.; Underwood, G.J.C. Influence of macrophyte spatial architecture on periphyton and macroinvertebrate community structure in shallow water bodies under contrasting land management. Fundam. Appl. Limnol. 2010, 177, 19–37. [Google Scholar] [CrossRef]
- Thullen, J.S.; Sartoris, J.J.; Walton, W.E. Effects of vegetation management in constructed wetland treatment cells on water quality and mosquito production. Ecol. Eng. 2002, 18, 441–457. [Google Scholar] [CrossRef]
- Knight, R.L.; Walton, W.E.; O’Meara, G.F.; Reisen, W.K.; Wass, R. Strategies for effective mosquito control in constructed treatment wetlands. Ecol. Eng. 2003, 21, 211–232. [Google Scholar] [CrossRef]
- Rey, J.R.; Walton, W.E.; Wolfe, R.J.; Connelly, C.R.; O’Connell, S.M.; Berg, J.A.; Sakolsky-Hoopes, G.E.; Laderman, A.D. North American wetlands and mosquito control. Int. J. Environ. Res. Public Health 2012, 9, 4537–4605. [Google Scholar] [CrossRef] [PubMed]
- Bowler, P.; Bretz, W.; (University of California-Irvine, Irvine, CA, USA). Personal communication, 2012.
- Mokry, L.; (Alan Plummer Associates Inc., College Station, TX, USA). Personal communication, 2014.
- Rollins, G.L. A Guide to Waterfowl Habitat Management in Suisun Marsh; California Department of Fish and Game: Sacramento, CA, USA, 1981.
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).