Abstract
In an effort to monitor ascidian recruitment in mussel aquaculture facilities, a series of settlement plates (20 × 20 cm) were placed in a mussel farm located in the Amvrakikos Gulf (Ionian Sea). The plates were vertically deployed on floating facilities in the water column at regular intervals (depths of 0.2 m, 1.5 m, and 3 m) to monitor the settlement and proliferation of ascidians. Furthermore, measurements of seawater physicochemical parameters such as temperature, salinity, dissolved oxygen, and chlorophyll-a concentration were conducted together with the record of ascidian species in each sampling from January 2021 to November 2021. The correlation of these parameters with ascidian species provides information on their effect on the periodicity of ascidians’ recruitment. The results demonstrated a significant correlation between ascidian presence and water temperature. The potential influence of other important environmental parameters such as chlorophyll-a was not revealed, likely due to the limited number of values and samples included in the analyses. While increased chlorophyll levels, reflecting increased primary productivity or nutrient availability, are associated with increased growth and reproduction of all ascidian species, the effect of temperature was more potent and species-specific. Ciona robusta, Styela plicata, Microcosmus squamiger, and Phallusia mammillata were mainly detected at temperatures below 25 °C, whereas Clavelina oblonga was prevalent at temperatures above 25 °C. The absence of most ascidians at temperatures above 25 °C was possibly attributed to decreased settlement success and to the increased competition from C. oblonga at higher temperatures. The deployment of settlement plates in correlation with seawater physiochemical parameters can provide valuable data on ascidian settlement dynamics and support the development of targeted management practices for biofouling control.
Keywords:
aquaculture; tunicates; bivalve culture; Greece; chlorophyll; temperature; salinity; oxygen availability; biofouling 1. Introduction
Since ascidians are among the most significant biofouling organisms, they lead to significant economic consequences in mussel farms as these facilities provide an ideal substrate for the proliferation and spread of ascidian species [1,2,3]. The most common ascidian [Ascidiacea (Chordata: Tunicata)] species are found attached to any kind of hard substrate (e.g., human-made infrastructure), as well as epibionts of other organisms (plants and animals). The size of ascidians may vary from 0.5 to 30 cm according to their morphology, i.e., colonial or solitary type, while they can frequently form large colonies, spreading in thin and wide layers allowing their symbiotic unicellular algae to utilize the necessary light [4].
A significant number of ascidian species has been detected in Greece [3,4], including invasive or cryptic species. These species usually attach and grow on bivalve shells competing for food, while at the same time, they add excessive weight to the rearing units’ floating facilities (long line floating culture system) [1,2,5,6,7,8,9,10,11]. In addition, these ascidian species can affect water [12] and oxygen [13,14,15,16,17] exchange levels in bivalves resulting in decreased growth [2,18,19] and increased mortality rates [11,20,21,22], significantly threatening productivity in shellfish farming operations.
An investigation of the ascidian distribution, composition, and spread, which may differ in space and time, in combination with practices for the mitigation of ascidian settlement in mussel farms, is essential for the evaluation of their impact on the aquaculture sector and its subsequent economic losses. Although water temperature is considered to be the primary influencing factor [23,24,25] resulting in increased ascidian spawning during summer’s elevated temperatures [24,26], little is known regarding the effect of other physicochemical parameters on their reproductive cycles [27,28,29]. In addition, a variety of factors including light intensity [30,31,32], the existence of congeneric species [33,34], the characteristics of the host surface [33,34], and the chemical composition of the substrate [35,36] have been proposed to influence the ascidian larval settlement. In this context, it becomes clear that seasonal tracking of the ascidian presence facilitates the estimation of their dispersal patterns and physiological effects on mussels, pinpointing critical periods when biofouling poses a threat to mussel farming [37]. Thus, data regarding the patterns of ascidians’ occurrence and attachment provide valuable information for predicting the emergence of dominant species, the timing of their appearance, and the areas where their settlement will be most pronounced [38].
Since most studies evaluating these factors on ascidians’ presence originate from laboratory experiments or were focused on a single parameter, the aim of the present study was to shed light on the seasonal preference of ascidians, in correlation with the main physicochemical parameters influencing their presence. Their seasonal presence was documented in mussel farm facilities in Western Greece with the subsequent identification of the ascidian species. Monthly recordings of the area’s physicochemical parameters including temperature, salinity, dissolved oxygen, and eutrophication (chlorophyll-a concentration) were also conducted in order to assess their influence on the periodic occurrence of ascidians in the region as they are closely linked to the biological cycles of invasive species.
2. Materials and Methods
2.1. Study Area Description
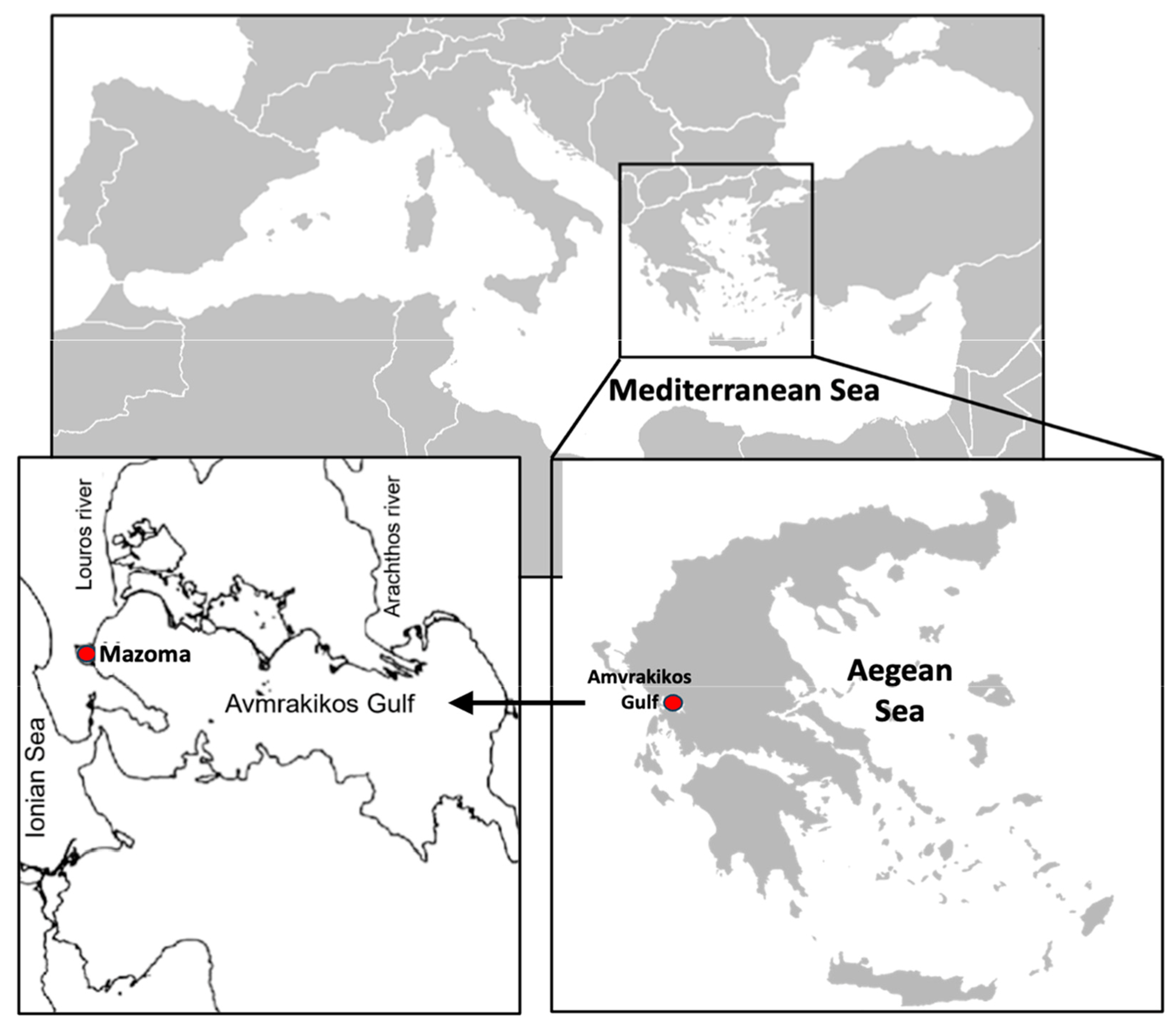
The field experiments were implemented on the installations of a raft installed at the mussel farm to which the experiments were attached (Figure 1), on the outer side of the estuary/gate to the Mazoma Lagoon, located on the northwest coast of the Amvrakikos Gulf, next to the city of Preveza, Epirus (NW Greece) (Figure 2), with the following coordinates: 39°1.538′ N, 20°45.430′ E. The Amvrakikos Gulf resembles a fjord with an area of 525 km2. The renewal of its waters takes place from the Ionian Sea through the long but shallow “mouth” of the Gulf (Preveza—Aktio Channel). The volume of water entering the gulf is insufficient to renew the entire water mass, which is supplemented by the Arachthos and Louros rivers. Specifically, the Louros River (length 80 km, average annual discharge 400 × 103 m3 year−1) provides excessive amounts of nutrients to the gulf, due to the intensive agricultural activity and livestock production occurring upstream [39,40,41,42].

Figure 1.
Ascidian species on mussel farm infrastructures in Mazoma, Amvrakikos Gulf.
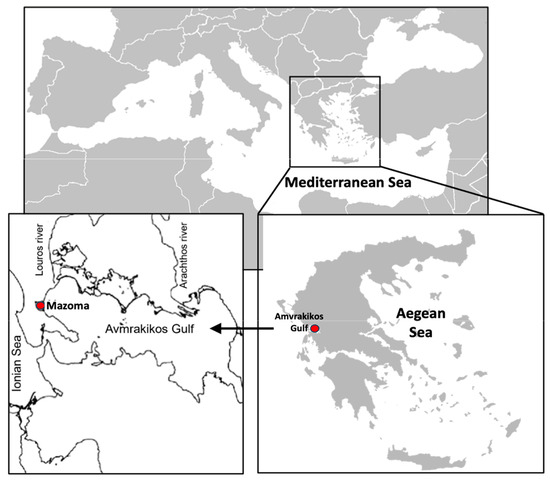
Figure 2.
Map location of the Amvrakikos Gulf and the area of Mazoma Lagoon where the mussel farm in which the field experimental procedure was conducted is located.
The hydrological nutrient profile, combined with limited seawater renewal, induces vertical stratification at 6–10 m, creating hypoxic (<2 mg L−1 O2) or anoxic (<0.2 mg L−1 O2) conditions near the riverbed [43,44]. Zooplankton, including bivalve larvae, decrease in abundance with depth, while autumn mixing (October–November) leads to a uniform vertical distribution [41,45,46]. Some zooplankton adjust their vertical migration patterns to avoid low-oxygen zones, leading to altered distribution and potential impacts on feeding and predator–prey interactions. Research indicates that hypolimnetic anoxia can modify zooplankton migration behaviors, biomass, and overall community structure [47].
2.2. Assessment of Abiotic and Biotic Environmental Factors
A total of 9 samplings were carried out between January 2021 and November 2021 every 1 or 2 months (Table 1) for both water and settlement plates. The water samplings were carried out at two different depths, 0.2 m and 3 m. Water samples were taken to the laboratory where phytoplankton analysis was carried out using a compound light microscope and identification keys summarized by Santhanam and Perumal (2008) [48]. Along with obtaining water samples for quality analysis, temperature, dissolved oxygen, and salinity were also recorded during sampling on-site. Measurements of temperature and dissolved oxygen were performed using a WTWoxi 196 portable meter (Xylem, Weilheim, Germany) and the EasyLog-USB-1-PRO Industrial USB Temperature Data Logger (Lascar SE Asia, Hong Kong). Prior to each sampling, a 5% Na2SO3 solution was employed to set the zero value, and well-aerated water to set the maximum value. The salinity measurements were carried out using a WTWLF 191 salinometer (Xylem, Weilheim, Germany). Distilled water was used for the initial zero setting and two measurements were taken, the first at a depth of 0.2 m and the second at 3 m from the surface in duplicate.

Table 1.
Timeline of the settlement plates’ placement, samplings, and total duration of the settlement plates in the sea.
A Shimadzu UV 1800 spectrophotometer with stoppered cuvettes with a path length of 1 cm was used for the trichromatic determination (630 nm, 647 nm, 664 nm, 750 nm) of chlorophyll-a. The concentration of chlorophyll-a was calculated according to the equations of Jeffrey and Humphrey [49]:
where:
E = the absorbance intensity;
Ve = extraction volume (mL);
Vf = filtered volume (L).
The extraction was performed by grinding the filters with 90% acetone.
2.3. Settlement Plate Placement
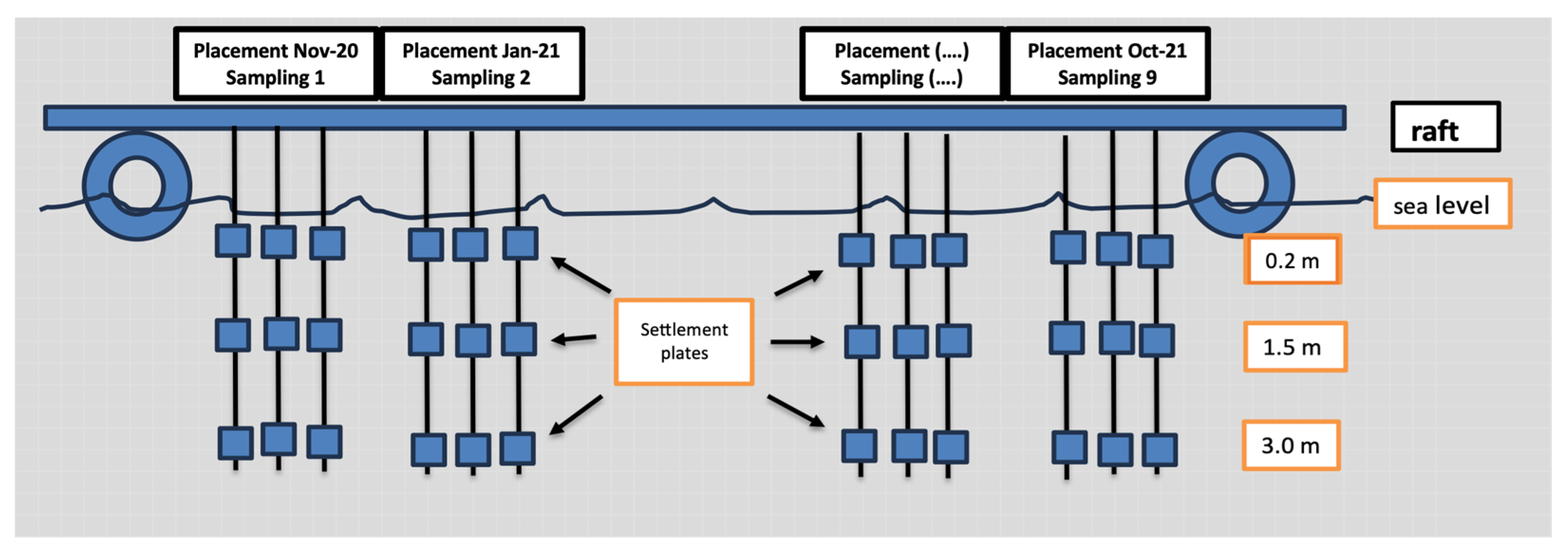
A floating unit-rearing platform (raft) was employed for the hanging of the settlement plates. These settlement plates consisted of 20 × 20 cm polyester tiles with quite rough (about 3 mm thick) surfaces (not flexible), secured to the rope with plastic clips. Each rope was 4 m long with a weight at the lower end to stay vertical in the water surface (as much as possible). Three settlement plates per unit (Figure 3) were attached to each rope at depths of 0.2 m, 1.5 m, and 3 m. Each sampling was performed in triplicate (Figure 4).

Figure 3.
Settlement plates’ construction.
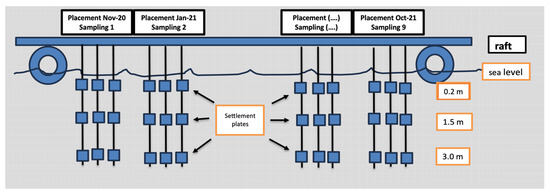
Figure 4.
Schematic representation of experimental floating unit-rearing platform (raft) hanging the settlement plates. In the first 6 months, settlement plates were installed every 2 months, while for the next 6 months, settlement plates were installed every month.
2.4. In Situ Species Assessment and Systematic Classification
The frequency of settlement plates’ installation and sampling are presented in Table 1. In the first 6 months, settlement plates were installed every 2 months and, correspondingly, the sampling was performed every 2 months, while for the next 6 months, settlement plates were installed every month and sampled on a monthly basis (Table 1). For each group, the settlement plates remained suspended for the entire period (1–2 months), while after that period, the plates were removed and renewed with new ones.
For each sampling, visual inspection and photography of the two surfaces of each plate were conducted for the identification of ascidians’ presence. Ascidian settlement was determined according to Casso et al. (2018) [50]. Specifically, all the attached ascidians of different species were collected using forceps and scissors and were transferred to the laboratory preserved in formaldehyde for their systematic classification. Plates were inspected visually and photographed in each sampling. The presence of ascidian species was recorded in situ, and species identity was confirmed using collected samples.
2.5. Redundancy Analysis (RDA) and ANOVA Permutation Test on the Influence of Environmental Variables on Species Presence
Redundancy analysis (RDA) was performed to investigate the relationship between environmental variables and species presence/absence data. The environmental variables included in the analyses were chlorophyll-a, temperature, salinity, and dissolved oxygen, while species presence/absence data were used as response variables. The analysis included five ascidians identified at the species level: Ciona robusta, Clavelina oblonga, Styela plicata, Microcosmus squamiger, and Phallusia mammilata. Prior to the analysis, all values of the environmental parameters were standardized using z-scores to ensure comparability of the variables with different units. The RDA was conducted using the vegan package in R (Version 4.4.2; R Foundation for Statistical Computing, Vienna, Austria), with species presence data modeled as a function of the standardized environmental parameters. Afterward, a permutation test with 999 iterations was performed to assess the significance of each environmental parameter and test the overall model fit. The significance of each parameter was determined based on permutation p-values, with p-values less than 0.05 considered statistically significant. The RDA scores of the first and second redundancy axes (RDA1 and RDA2) both for environmental parameters and for the ascidian species were extracted from the RDA analysis in R. These scores were then visualized using GraphPad Prism version 8.4.2 for further interpretation and presentation.
3. Results
3.1. Physicochemical Parameters
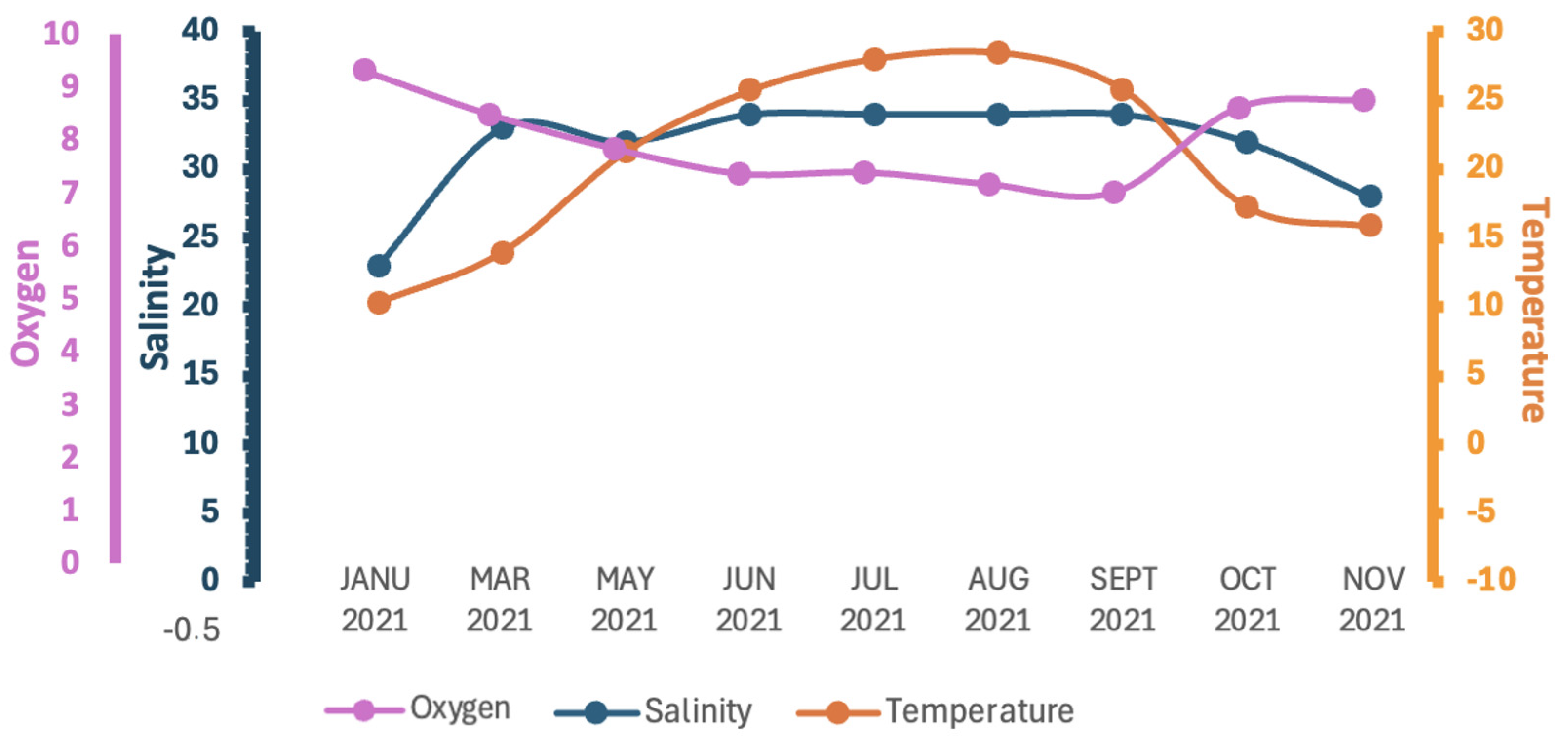
The water temperature significantly fluctuated throughout the year, ranging from 10.28 °C in the winter period (January) to 28.47 °C in the summer (August) (mean values ± SD) (Figure 5; Table 2).
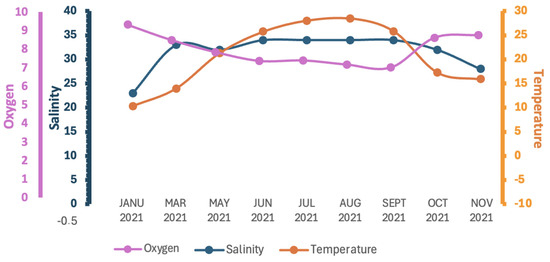
Figure 5.
Seasonal seawater temperature, dissolved oxygen, and salinity levels in Amvrakikos Gulf study area. Oxygen (mg L−1); salinity (ppt); temperature (°C).
Table 2.
Physicochemical parameters of the seawater obtained from the water samples in the study area (mean values ± SD).
Salinity values also varied seasonally, ranging from 18 ppt in December up to 35 ppt in July at a depth of 0.2 m (Figure 5). Similar observations in the Amvrakikos Gulf have revealed seasonal and depth-related salinity stratification and variation. The stratified surface layer, typically 10–20 m thick, exhibits salinity levels ranging from 25 to 30 ppt, depending on the season and specific location. In the northern regions of the Amvrakikos Gulf, where riverine input is more pronounced, salinity levels are lower compared to the southern regions. Beneath this stratified layer, the water is more saline and originates from the Ionian Sea [44,45,46].
Dissolved oxygen levels remained consistently high throughout the measurement period, ranging from a peak of 9.30 mg/L in January to a minimum of 6.99 mg/L in August (Figure 5). These oxygen concentrations are well above the 3 mg/L threshold required for well-oxygenated waters. These findings align with previous studies indicating that the surface waters of the Amvrakikos Gulf are rich in oxygen concentration. This phenomenon can be attributed to the continuous oxygenation of riverine waters as they flow toward the Gulf, coupled with the enhanced solubility of oxygen in low-salinity waters [11,51].
According to the measurements, it appears that the values of phytoplankton abundance and chlorophyll concentration that have been measured in the Amvrakikos Gulf exhibit a seasonal pattern, with their high values being similar to those of the most eutrophic areas of Greece (Table 3) [52].
Table 3.
Additional physicochemical parameters of the seawater obtained from the water samples in the study area.
3.2. Species Recruitment
During the deployment of the settlement plates, seven ascidian species were observed adhering to the surfaces. Of these, five species were identified based on the work of Geladakis et al. [53], including one endemic species (P. mammillata) and four invasive species (S. plicata, C. robusta, C. oblonga, and M. squamiger).
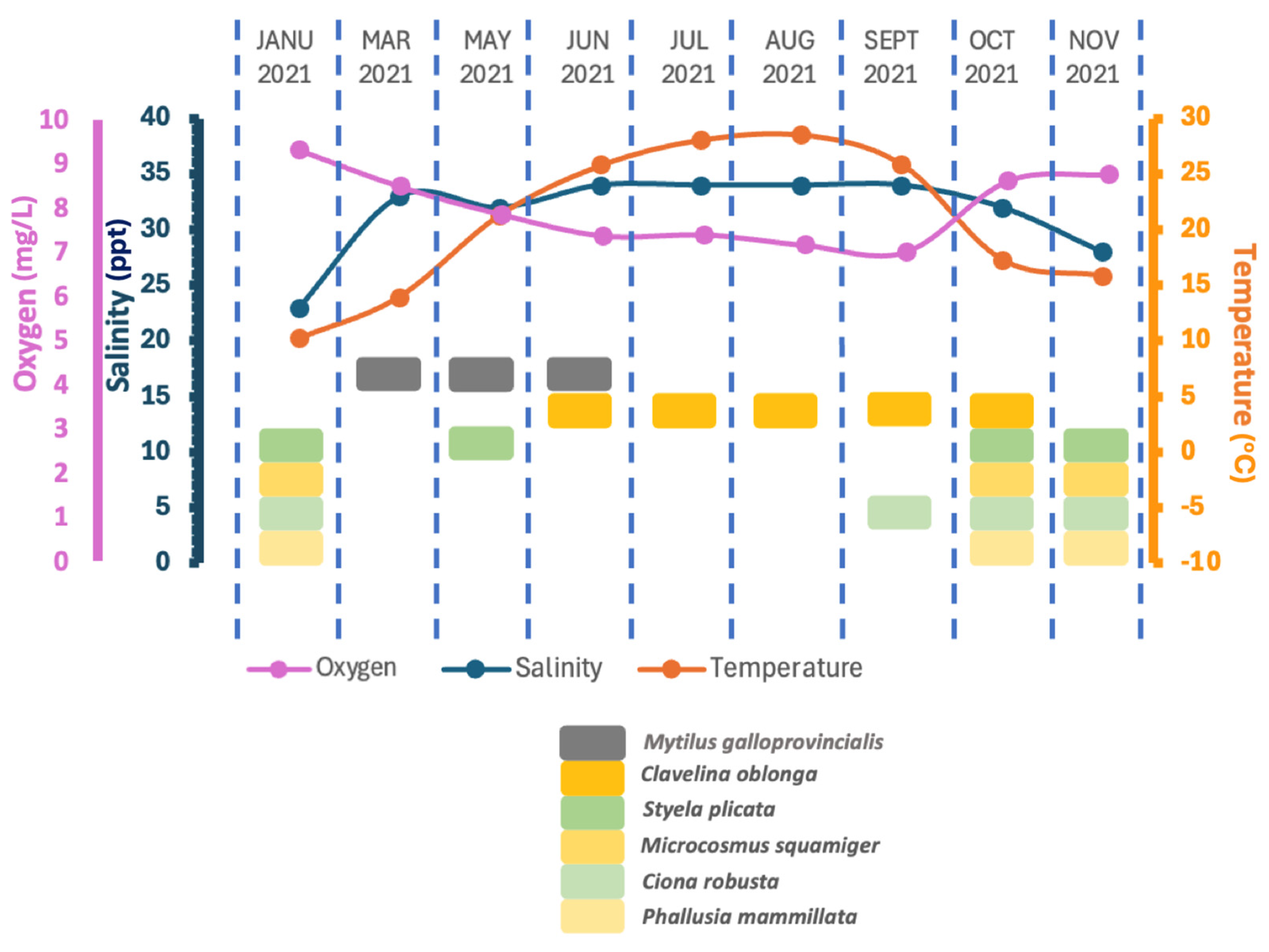
The seasonal patterns of ascidian colonization varied across species. C. robusta, M. squamiger, and P. mammillata were primarily found on the settlement plates deployed late in the autumn and early in the winter. S. plicata was also detected on settlement plates sampled in May, except for October, November, and January indicating broader seasonal activity (Figure 6).

Figure 6.
Population coverage of settlement plates with Styela plicata species in January at 0.2 m, 1.5 m, and 3 m.
During February, March, and April, no ascidians were observed on the settlement plates; only mussel larvae were present. With the increase in sea temperature exceeding 25 °C in June, the first appearance of C. oblonga was recorded. This species exhibited a significant population outbreak during the warmest months, July and August, followed by a decrease in September, minimum presence in October, and complete absence by November (Figure 7).

Figure 7.
Settlement plates from (a) June (b) August (c) September, and (d) November samplings. Settlement plates indicate the cumulative settlement and development of ascidian colonies (Clavelina oblonga) throughout the interval period.
The proliferation of C. oblonga in both mussel farming structures and the floating facility of the plant poses a substantial challenge to production processes due to its high abundance during the summer months. The species identified and their seasonal distribution are summarized in Figure 8.
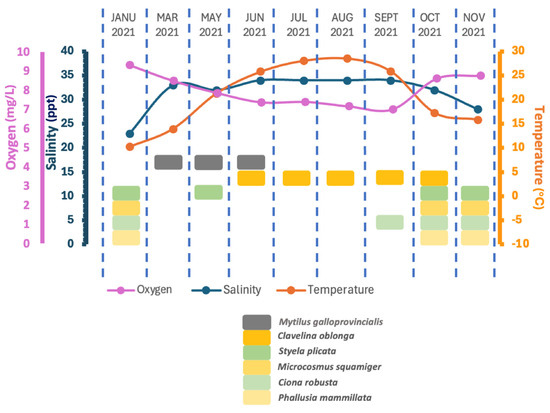
Figure 8.
Ascidian species and Mytilus galloprovincialis recruitment on the settlement plates during the study period. In the present figure, the ascidian species and abiotic parameters are matched only to seasonal data. There is no correlation between ascidian species and abiotic parameters.
3.3. Influence of Environmental Variables on Species Presence/Absence: Insights from Redundancy Analysis (RDA)
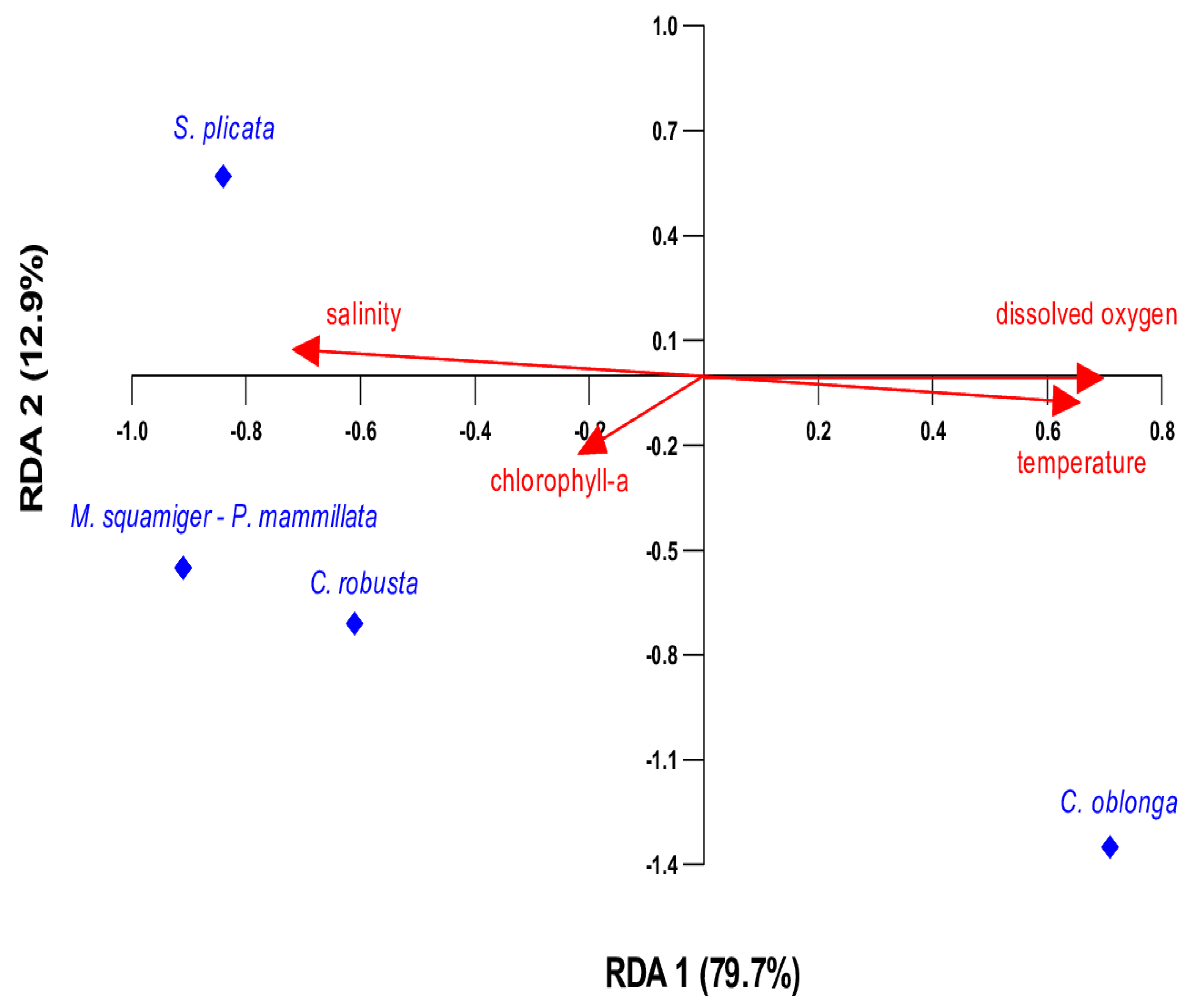
The redundancy analysis (RDA) illustrates how the distribution of ascidian species is influenced by environmental parameters (temperature, chlorophyll-a, dissolved oxygen, and salinity) (Figure 9). The five species C. robusta, M. squamiger, P. mammillata, C. oblonga, and S. plicata were used for the analyses. According to the RDA biplot, RDA successfully captured the majority of the variation in the data in the first two axes, which explains 92.6% of the data variability, indicating a strong relationship between species presence and environmental variables. More specifically, RDA1 explains most of the variation (79.7%), primarily separating the species based on temperature and dissolved oxygen. Accordingly, RDA2 captures a smaller fraction (12.9%), likely distinguishing species based on secondary factors like chlorophyll-a and salinity.
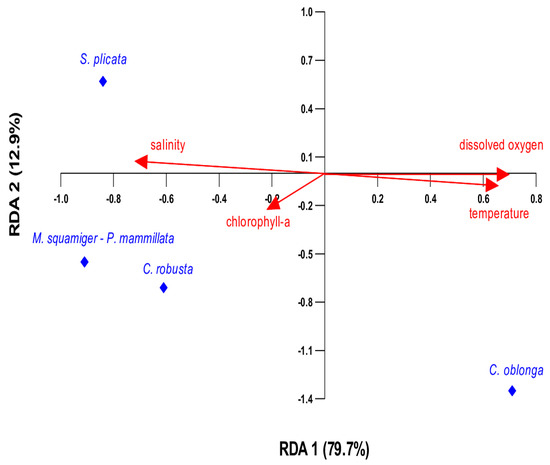
Figure 9.
RDA biplot showing the relationship between environmental variables (red arrows) and species presence (blue points). The first redundancy axis (RDA1) accounts for 79.7% of the variation, while the second axis (RDA2) explains 12.9%.
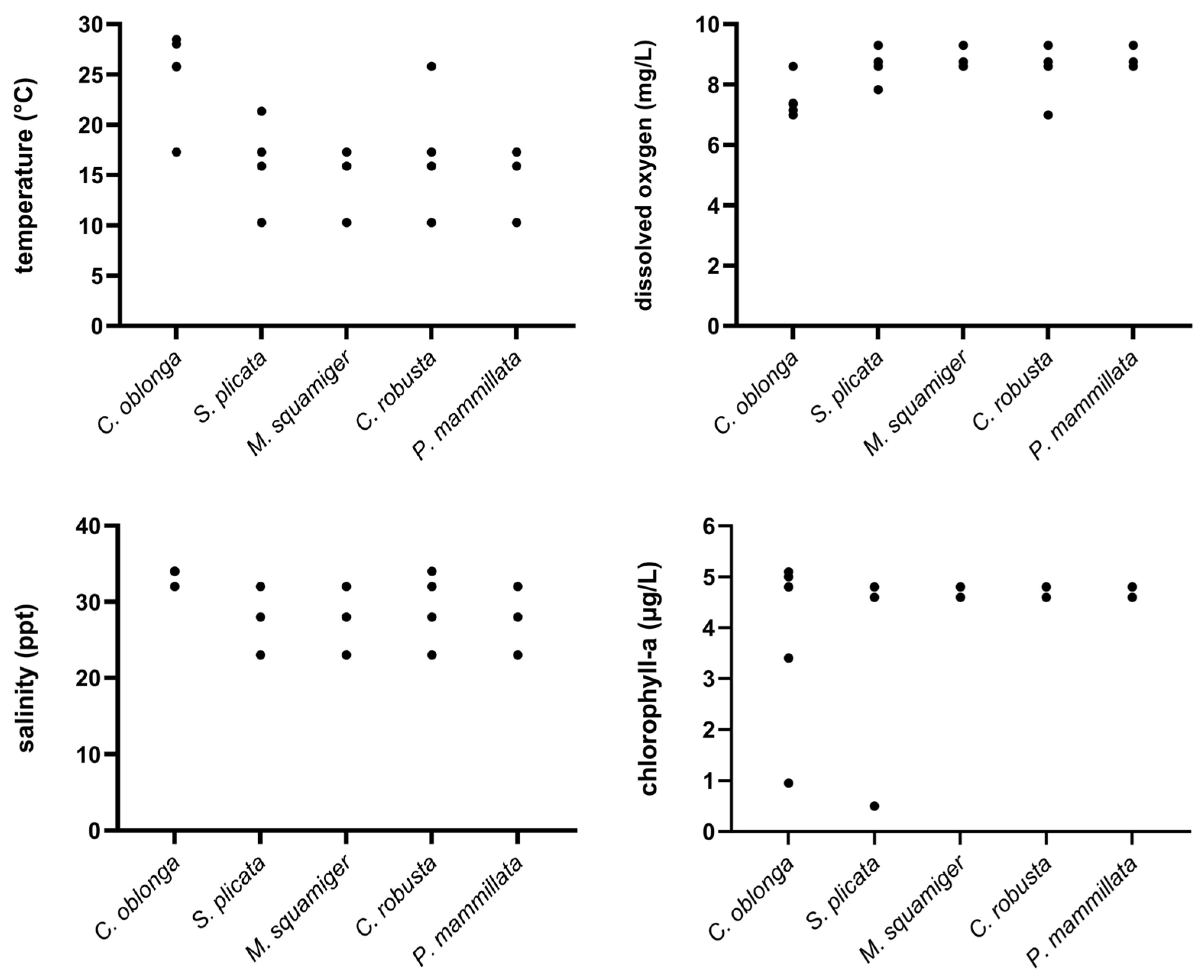
Most environmental parameters yielded high p-values (Table 4) after the ANOVA permutation test, suggesting that their potential effects may not be statistically detectable, likely due to the limited sample size. As a result, their true influence may not be reflected in the statistical results. Among the tested environmental parameters, temperature appears to be the most influential factor, significantly shaping species presence (Table 4). This suggests that temperature is the dominant environmental factor shaping species’ presence. Temperature is positively correlated, influencing species distribution along RDA1 (Figure 9). C. oblonga is positioned along the positive RDA1 axis, indicating a strong association with elevated temperatures. S. plicata aligns more with salinity and chlorophyll-a, suggesting it thrives in many different conditions compared to the other species. M. squamiger, P. mammillata, and C. robusta were positioned in the lower-left part of the plot, indicating an increased presence at lower temperatures. The presence/absence of ascidian species according to physicochemical parameters is also depicted in Figure 10.
Table 4.
Results of the ANOVA permutation test (999 permutations) evaluating the influence of environmental variables on species presence. p-values indicate the statistical significance of each variable, with p < 0.05 considered significant and indicated with a * in the Table.
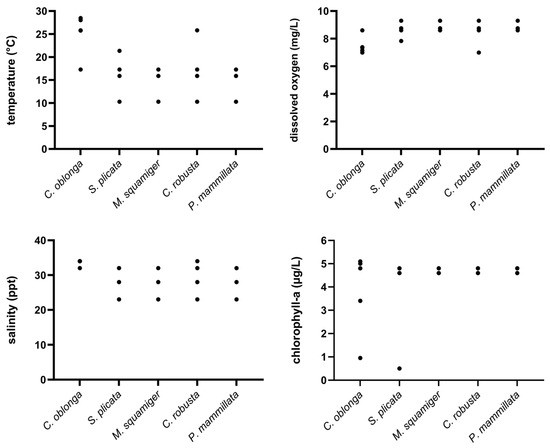
Figure 10.
Diagram showing the range of values for the tested environmental variables (temperature, dissolved oxygen, salinity, and chlorophyll-a) on the x-axis, where different ascidian species were present (y-axis).
4. Discussion
Seven different ascidian species were detected on the settlement plates, highlighting the significant role of aquaculture units as vectors for the introduction and spread of non-indigenous species [54,55]. Five out of those seven species were identified at a species level and were considered for the analyses. The use of settlement plates has been extensively documented in previous studies of ascidian settlement [48,56], both for descriptive purposes and experimental investigations [57,58,59]. In the present study, species such as C. robusta, S. plicata, M. squamiger, and P. mammillata were detected in samples collected during periods corresponding to the placement of settlement plates in November 2020 and October–November 2021 (Samplings 1, 8, and 9). S. plicata was also identified during Sampling 4, conducted in June. Meanwhile, C. oblonga was mainly found in settlement plates deployed in June, July, August, and September (Samplings 5, 6, 7, and 8).
The species distribution was closely linked to temperature variations, which was highlighted by the RDA biplot. The seasonal presence of S. plicata aligns with findings from Qiu et al. [60], which indicate that this species thrives in temperatures between 18 and 24 °C, while larvae fail to reach settlement stages at temperatures of 26–30 °C. This fact explains its absence during warmer months and its highly negative RDA1 score. Additionally, the absence of S. plicata in plates sampled in March is likely due to temperatures dropping below 18 °C during winter, as larvae experience prolonged metamorphosis times, depleted energy reserves, and ultimately fail to survive [61]. Furthermore, the species S. plicata, C. robusta, M. squamiger, and P. mammillata were detected at temperatures below 25 °C, whereas C. oblonga presence was prevalent at temperatures exceeding 25 °C. Cooler months may favor different species due to higher oxygen levels and lower temperatures, coinciding with less competition from C. oblonga, which has the potential to become a dominant competitor after an initial growth phase under favorable conditions [62,63]. The ANOVA permutation test reinforced these findings, highlighting the importance of temperature in structuring ascidian communities.
Salinity also plays a critical role in the survival and settlement of S. plicata larvae, since levels below 25 ppt inhibit the development of fertilized eggs, as reported by Thiyagarajian and Quian [64]. These observations are consistent with embryonic patterns observed across various ascidian species [65,66,67,68].
The slight variations in settlement periods, colony numbers, and species composition on floating aquaculture structures compared to the settlement plates could be attributed to differences in substrate properties, such as material composition and surface roughness, which influence larval settlement preferences [69]. For instance, S. plicata was detected on mooring ropes in June. Newly installed settlement plates, lacking biofilm, surface irregularities, and potential shelters that accumulate over time, may not provide suitable conditions for the establishment of some species. Certain ascidian species require unoccupied substrates for recruitment and survival, while others benefit from the structural complexity created by pre-existing epibiont organisms [58]. Additionally, interactions between population dynamics and environmental factors influence competitive outcomes among ascidians on artificial substrates [70]. Over time, environmental factors contribute to achieving ecological balance [71]. Laboratory experiments on larval behavior can complement field studies by clarifying whether differences in settlement density, pre-settlement mortality, or post-settlement growth contribute to observed population dynamics. Such studies indicate that many invertebrates, including ascidians, preferentially settle on shaded surfaces [31,32]. This behavior may decrease competition and the risk of macroalgal overgrowth, enhancing survival and establishment. Furthermore, substrate microtopography possesses a key role in settlement by influencing water flow, individual stress, and the availability of dissolved gases and nutrients [72]. Rough surfaces, which promote turbulent flow, have been shown to facilitate settlement for various aquatic organisms, including barnacles [73,74], bivalves [75], and polychaetes [76,77]. However, other species, including ascidians, bryozoans, polychaetes [78,79], and barnacles [80], exhibit a preference for settling on smooth substrates. Substrate characteristics, such as color and chemical composition, are also known to influence larval settlement, although these factors have been investigated in relatively few studies [36,81].
It should be also noted that the present study’s data show that chlorophyll-a levels are associated with specific biological or environmental processes that vary independently of temperature and dissolved oxygen. Chlorophyll levels could reflect seasonal changes in primary productivity or nutrient availability that impact biological dynamics in the marine area, with increased food likely increasing the growth and reproduction of the ascidian species. Chlorophyll-a concentration in water is directly associated with phytoplankton abundance, as chlorophyll-a is the primary pigment involved in photosynthesis within phytoplankton cells. Higher concentrations of chlorophyll-a typically indicate greater phytoplankton biomass and productivity. The position of the ascidians in the RDA biplot represents patterns in ascidian species’ presence/absence that align with the environmental factors. C. obonga was detected from June to October in the study area exhibiting its peak densities during August and September when, apart from the high temperature, chlorophyll levels were also increased compared to June and July. In line with our results, the growth of C. oblonga has been associated with rising seawater temperatures and increased chlorophyll-a [50,82]. All other ascidian species were present in the study area from October to January, and their presence was accompanied by increased chlorophyll levels. Accordingly, the total absence of ascidian species during March and only the presence of S. plicata in May is possibly attributed to the low temperatures and low chlorophyll levels in the previous months. However, the potential effects of chlorophyll-a on species presence were not statistically detectable, presumably owing to the limited sample size.
Seasonal shifts in dominant species and early-stage biotic interactions pose significant implications for species coexistence and the diversity of ascidians and other fouling organisms [83,84]. Beyond substrate properties and interspecies interactions, the timing of settlement plate deployment is a crucial factor influencing settlement. Aligning substrate availability with reproductive cycles is vital for successful species establishment, as the deployment date of settlement plates has a pronounced effect on species coverage [71]. The physical stability of installations, as well as the hardness, rigidity of materials, and prevailing currents, likely play a more significant role in larval settlement than the material composition of the settlement plates themselves [60]. In mussel aquaculture, handling and cleaning processes inadvertently create clean surfaces when mussels are returned to the water, providing an ideal substrate for ascidian settlement. Larvae that settle on concealed surfaces, such as within rigging or rearing ropes, often go unnoticed until they grow large enough to become visible [60]. Consequently, information regarding ascidian settlement often becomes available only after the fouling has already occurred.
These results emphasize the need for proactive aquaculture management. Monthly deployment of settlement plates provides the early detection of fouling species, enabling timely mitigation strategies. Understanding the environmental and biological factors influencing settlement can inform targeted control measures, reducing the impact of invasive ascidians on aquaculture operations.
5. Conclusions
Mussel farms in the Amvrakikos Gulf have become hotspots for invasive species, with distinct temporal patterns observed for the appearance of each species. Among the environmental factors influencing these patterns, temperature and chlorophyll-a levels play a pivotal role. The rapid and substantial rise in water temperature during the summer and autumn months significantly promotes the reproduction and abundance of C. oblonga, both in aquaculture facilities and the bivalves being farmed. The intensity of this phenomenon raises concerns about its potential long-term impacts, particularly in light of the ongoing sea temperature increases as a result of global warming. Therefore, implementing a comprehensive monitoring program to track the occurrence of ascidians in aquaculture facilities is imperative. While coverage data were only obtainable for certain species, occurrence data were collected for all the species observed. Notably, species like C. oblonga demonstrated widespread and persistent presence, appearing on multiple settlement plates over several months, warranting more attention. Occurrence rates have proven to be a reliable indicator of species abundance. Moreover, simple monitoring of species presence on settlement plates—a faster approach than detailed coverage analysis—can serve as an effective tool for tracking population dynamics. Continuous year-round monitoring is crucial for the early detection of newly introduced species, which is essential for implementing timely and effective mitigation measures. This approach also provides valuable insights into the settlement preferences of key species, enabling strategies to minimize ecological and operational impacts. Annual ascidian monitoring can facilitate the development of a robust database, offering more accurate predictions of problematic species occurrences, aiming for more targeted preventive measures, and thus enhancing the sustainability and productivity of aquaculture facilities, such as the ones in the Amvrakikos Gulf.
Author Contributions
Conceptualization, D.T. and J.A.T.; methodology, D.T., M.V.A. and I.A.G.; software, D.K.P. and K.F.; validation, M.V.A. and K.F.; formal analysis, M.V.A. and I.A.G.; investigation, D.T. and V.B.; resources, D.T. and J.A.T.; data curation, K.F. and I.A.G.; writing—original draft preparation, D.T., M.V.A., and D.K.P.; writing—review and editing, I.A.G., K.F. and J.A.T.; visualization, M.V.A. and J.A.T.; supervision, I.A.G. and J.A.T.; project administration, I.A.G. and J.A.T.; funding acquisition, J.A.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the EU-GREECE OPERATIONAL PROGRAM OF FISHERIES, EPAL 2014–2020, grant number MIS: 5048463. The present work is a part of the project “Development of the best control practices of invasive ascidians in mussel farming infrastructures and remediation of economic effects of invasion”.
Institutional Review Board Statement
This study did not require ethical approval since it concerns animals that do not require ethical approval for their study according to Directive 2010/63/EU, nor are they endangered.
Data Availability Statement
The data supporting the reported results of this study are included in this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Papadopoulos, D.K.; Lattos, A.; Giantsis, I.A.; Theodorou, J.A.; Michaelidis, B.; Feidantsis, K. The impact of ascidian biofouling on the farmed Mediterranean mussel Mytilus galloprovincialis physiology and welfare, revealed by stress biomarkers. Biofouling 2023, 39, 271–288. [Google Scholar] [CrossRef]
- Feidantsis, K.; Papadopoulos, D.K.; Lattos, A.; Theodorou, J.A.; Michaelidis, B.; Giantsis, I.A. Effects of biofouling by ascidians on cultured mussels: Apoptosis, autophagy, and antioxidant defense. J. Shellfish Res. 2023, 42, 199–213. [Google Scholar] [CrossRef]
- Alvanou, M.; Feidantsis, K.; Papadopoulos, D.; Lattos, A.; Theodorou, J.; Michaelidis, B.; Giantsis, I. Major ascidian species with negative impacts on bivalve aquaculture: Current knowledge and future research aims. Open Geosci. 2024, 16, 20220660. [Google Scholar] [CrossRef]
- Antoniadou, C.; Gerovasileiou, V.; Bailly, N. Ascidiacea (Chordata: Tunicata) of Greece: An updated checklist. Biodivers Data J. 2016, 4, e9273. [Google Scholar] [CrossRef]
- Duggan, W. Growth and survival of the bay scallop, Argopectenirradians, at various locations in the water column and at various densities. Proc. Natl. Shellfish. Ass. 1973, 63, 68–72. [Google Scholar]
- Leighton, D. A growth profile for the rock scallop Hinnitesmultirugosus held at several depths off La Jolla, California. Mar. Biol. 1978, 51, 229–232. [Google Scholar] [CrossRef]
- Leighton, D. Control of sabellid infestation in green and pink abalones, Haliotis fulgens and H. corrugate, by exposure to elevated water temperatures. J. Shellfish Res. 1998, 17, 701–705. [Google Scholar]
- Côté, J.; Himmelman, J.H.; Claereboudt, M.; Bonardelli, J.C. Influence of density and depth in the growth of juvenile sea scallps (Placopectenmagellanicus) in suspended culture. Can. J. Fish. Aquat. Sci. 1993, 50, 1857–1869. [Google Scholar] [CrossRef]
- Wilson, U.A.W. The Potential for Cultivation and Restocking of Pecten maximus (L.) and Aequipectenopercularis (L.) on Manx Inshore Fishing Grounds. Ph.D. Thesis, Port Erin Marine Laboratory, University of Liverpool, Isle of Man, UK, 1994. [Google Scholar]
- Lodeiros, C.J.M.; Himmelman, J.H. Influence of fouling on the growth and survival of the tropical scallop, Euvola (Pecten) ziczac (L. 1758) in suspended culture. Aquacult. Res. 1996, 27, 749–756. [Google Scholar] [CrossRef]
- Tsotsios, D.; Papadopoulos, D.K.; Alvanou, M.V.; Georgoulis, I.; Lattos, A.; Michaelidis, B.; Feidantsis, K.; Giantsis, I.A.; Theodorou, J.A. Environmentally friendly and efficient methods for mitigating the density of ascidian fouling in mediterranean mussel farming. Fishes 2024, 9, 135. [Google Scholar] [CrossRef]
- Claereboudt, M.R.; Bureau, D.; Côte, J.; Himmelman, J.H. Fouling development and its effect on the growth of juvenile giant scallops (Placopectenmagellanicus) in suspended culture. Aquaculture 1994, 121, 327–342. [Google Scholar] [CrossRef]
- Huguenin, J.E.; Huguenin, S.S. Biofouling resistant shellfish trays. J. Shellfish Res. 1982, 2, 41–46. [Google Scholar]
- Enright, C. Control of biofouling in aquaculture. World Aquac. 1993, 24, 44–46. [Google Scholar]
- Lu, Y.T.; Blake, N.J. The culture of the southern bay scallop in Tampa bay, an urban Florida estuary. Aquac. Int. 1997, 5, 439–450. [Google Scholar] [CrossRef]
- Cronin, E.R.; Cheshire, A.C.; Clarke, S.M.; Melville, A.J. An investigation into the composition, biomass and oxygen budget of the fouling community on a tuna aquaculture farm. Biofouling 1999, 13, 279–299. [Google Scholar] [CrossRef]
- Mazouni, N.; Gaertner, J.C.; Deslou-Paoli, J.M. Composition of biofouling communities on suspended oyster cultures: An in situ study of their interactions with the water column. Mar. Ecol. Progr. Ser. 2001, 214, 93–102. [Google Scholar] [CrossRef]
- Boyd, C.E. Water Quality Management for Pond Fish Culture; Elsevier Science: New York, NY, USA, 1982. [Google Scholar]
- Laired, L.; Needham, T. Salmon and Trout Farming; Ellis Horwood Limited: Chichester, UK, 1988. [Google Scholar]
- Daigle, R.M.; Herbinger, C.M. Ecological interactions between the vase tunicate (Ciona intestinalis) and the farmed blue mussel (Mytilus edulis) in Nova Scotia, Canada. Aquat. Invasions 2009, 4, 177–187. [Google Scholar] [CrossRef]
- Theodorou, A.J.; Perdikaris, C.; Spinos, E. On the occurrence of rayed pearl oyster Pinctada imbricata radiata (Leach, 1814) in Western Greece (Ionian Sea) and its biofouling potential. Biharean Biol. 2019, 13, 4–7. [Google Scholar]
- Tsotsios, D.; Moutopoulos, D.K.; Lattos, A.; Michaelidis, B.; Theodorou, J.A. Impacts of the Establishment of Biofoulants on Greek Aquaculture: Farmers’ Expert Knowledge. J. Mar. Sci. Eng. 2023, 11, 1077. [Google Scholar] [CrossRef]
- Bates, W.R. Environmental factors affecting reproduction and development in ascidians and other prochordates. Can. J. Zool. 2005, 83, 51–61. [Google Scholar] [CrossRef]
- Lambert, G. Ecology and natural history of the protochordates. Can. J. Zool. 2005, 83, 34–50. [Google Scholar] [CrossRef]
- Lambert, G. Invasive sea squirts: A growing global problem. J. Exp. Mar. Biol. Ecol. 2007, 1, 3–4. [Google Scholar] [CrossRef]
- Coma, R.; Ribes, M.; Gili, J.M.; Zabala, M. Seasonality in coastal benthic ecosystems. Trends Ecol. Evol. 2000, 15, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Turon, X. The ascidians of Tossa de Mar (NE Spain) 2. Biological cycles of the colonial species. Cah. Biol. Mar. 1988, 29, 407–418. [Google Scholar]
- Turon, X. Periods of non-feeding in Polysyncratonlacazei (Ascidiacea: Didemnidae): A rejuvenative process? Mar. Biol. 1992, 112, 647–655. [Google Scholar] [CrossRef]
- Perez-Portela, R.; Palacin, C.; Duran, S.; Turon, X. Biological traits of three closely related species of Pycnoclavella (Ascidiacea) in the Western Mediterranean. Mar. Biol. 2007, 152, 1031–1038. [Google Scholar] [CrossRef]
- Thorson, G. Light as an ecological factor in the dispersal and settlement of larvae of marine bottom invertebrates. Ophelia 1964, 1, 167–208. [Google Scholar] [CrossRef]
- Durante, K.M. Larval behavior, settlement preference, and induction of metamorphosis in the temperate solitary ascidian Molgulacitrina Alder & Hancock. J. Exp. Mar. Biol. Ecol. 1991, 145, 175–187. [Google Scholar]
- Rius, M.; Branch, G.M.; Griffiths, C.L.; Turon, X. Larval settlement behaviour in six gregarious ascidians in relation to adult distribution. Mar. Ecol. Prog. Ser. 2010, 418, 151–163. [Google Scholar] [CrossRef]
- Burke, R. Pheromones and the gregarious settlement of marine invertebrate larvae. Bull. Mar. Sci. 1986, 39, 323–331. [Google Scholar]
- Pawlik, J. Chemical ecology of the settlement of benthic marine-invertebrates. Oceanogr. Mar. Biol. 1992, 30, 273–335. [Google Scholar]
- Kerr, A.; Beveridge, C.M.; Cowling, M.J.; Hodgkiess, T.; Parr, A.C.S.; Smith, M.J. Some physical factors affecting the accumulation of biofouling. J. Mar. Biol. Assoc. UK 1999, 79, 357–359. [Google Scholar] [CrossRef]
- Bavestrello, G.; Bianchi, C.N.; Calcinai, B.; Cattaneo-Vietti, R.; Cerrano, C.; Morri, C.; Puce, S.; Sara, M. Bio-mineralogy as a structuring factor for marine epibenthic communities. Mar. Ecol. Prog. Ser. 2000, 193, 241–249. [Google Scholar] [CrossRef]
- Ramsay, A.; Davidson, J.; Bourque, D.; Stryhn, H. Recruitment patterns and population development of the invasive ascidian Ciona intestinalis in Prince Edward Island, Canada. Aquat. Invasions. 2009, 4, 169–176. [Google Scholar] [CrossRef]
- Woods, C.M.C.; Floerl, O.; Hayden, B.J. Biofouling on Greenshell™ mussel (Perna canaliculus) farms: A preliminary assessment and potential implications for sustainable aquaculture practices. Aquacult. Int. 2012, 20, 537–557. [Google Scholar] [CrossRef]
- Katselis, G.N.; Moutopoulos, D.K.; Dimitriou, E.N.; Koutsikopoulos, C. Long-term changes of fisheries landings in enclosed gulf lagoons (Amvrakikos gulf, W Greece): Influences of fishing and other human impacts. Est. Coast. Shelf Sci. 2013, 131, 31–40. [Google Scholar] [CrossRef]
- Kountoura, K.; Zacharias, I. Temporal and spatial distribution of hypoxic/seasonal anoxic zone in Amvrakikos gulf, Western Greece. Estuar. Coast. Shelf Sci. 2011, 94, 123–128. [Google Scholar] [CrossRef]
- Kountoura, K.; Zacharias, I. Trophic state and oceanographic conditions of Amvrakikos Gulf: Evaluation and monitoring. Desalination Water Treat. 2013, 51, 2934–2944. [Google Scholar] [CrossRef]
- Nicolaidou, A.; Moraitou-Apostolopoulou, M.; Ignatiades, L.A. Survey of estuarine benthic, zooplanktonic and phytoplanktonic communities of Amvrakikos Gulf, Ionian Sea. Mar. Ecol.-Pubbl. Stn. Zool. Napoli. 1983, 4, 197–209. [Google Scholar] [CrossRef]
- Panayotidis, P.; Pancucci, M.A.; Balopoulos, E.; Gotsis-Skretas, O. Plankton distribution patterns in a Mediterranean dilution basin: Amvrakikos Gulf (Ionian Sea, Greece). Mar. Ecol.-Pubbl. Stn. Zool. Napoli 1994, 15, 93–104. [Google Scholar] [CrossRef]
- Friligos, N.; Balopoulos, E.T.; Psillidou-Giouranovits, R. Eutrophication and hydrography in the Amvrakikos Gulf, Ionian Sea. Fresenius Environ. Bull. 1997, 6, 21–26. [Google Scholar]
- Ferentinos, G.; Papatheodorou, G.; Geraga, M.; Iatrou, M.; Fakiris, E.; Christodoulou, D.; Dimitriou, E.; Koutsikopoulos, C. Fjord water circulation patterns and dysoxic/anoxic conditions in a Mediterranean semi-enclosed embayment in the Amvrakikos Gulf, Greece. Estuar. Coast. Shelf Sci. 2010, 88, 473–481. [Google Scholar] [CrossRef]
- Kehayias, G.; Aposporis, M. Zooplankton variation in relation to hydrology in an enclosed hypoxic bay (Amvrakikos Gulf, Greece). Mediterr. Mar. Sci. 2014, 15, 554–568. [Google Scholar] [CrossRef]
- Doubek, J.P.; Campbell, K.L.; Doubek, K.M.; Hamre, K.D.; Lofton, M.E.; McClure, R.P.; Ward, N.K.; Carey, C.C. The effects of hypolimnetic anoxia on the diel vertical migration of freshwater crustacean zooplankton. Ecosphere 2018, 9, e02332. [Google Scholar] [CrossRef]
- Santhanam, P.; Perumal, P. Marine plankton in Indian waters. In Training Manual on GIS and Marine Biodiversity; John Milton, M.C., Ed.; Lyola College Pub: New Delhi, India, 2008; p. 492. [Google Scholar]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Und Physiol. Der Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Casso, M.; Navarro, M.; Ordóñez, V.; Fernández-Tejedor, M.; Pascual, M.; Turon, X. Seasonal patterns of settlement and growth of introduced and native ascidians in bivalve cultures in the Ebro Delta (NE Iberian Peninsula). Reg. Stud. Mar. Sci. 2018, 23, 12–22. [Google Scholar] [CrossRef]
- Theodorou, A.J.; Spinos, E.; Ramfos, A.; Tsamadias, E.I.; Bekiari, V.; Kamilari, M.; Ntouni, M.-M.; Tsotsios, D.; Feidantsis, K.; Lattos, A.; et al. Fan Mussel (Pinna nobilis L.) Spat Collection, Monitoring of Early Growth and Conservation Implications by Deploying Conventional Aquaculture Methodology. J. Mar. Sci. Eng. 2024, 12, 2070. [Google Scholar] [CrossRef]
- Koukaras, K.; Nikolaidis, G. Dinophysis blooms in Greek coastal waters (Thermaikos Gulf, NW Aegean Sea). J. Plankton Res. 2004, 26, 445–457. [Google Scholar] [CrossRef]
- Geladakis, G.; Kommata, V.; Kamilari, M.; Papaioannou, C.; Papadopoulos, D.K.; Lattos, A.; Michaelidis, B.; Theodorou, I.A.; Batargias, C. Genetic identification of Ascidian species in mussel farm facilities of Greece. In Proceedings of the 18th Panhellenic Ichthyologist Conference, Messolonghi, Greece, 3–6 November 2022; pp. 436–439. (In Greek). [Google Scholar]
- Rius, M.; Heasman, K.G.; McQuaid, C.D. Long-term coexistence of nonindigenous species in aquaculture facilities. Mar. Pollut. Bull. 2011, 62, 2395–2403. [Google Scholar] [CrossRef]
- Fitridge, I.; Dempster, T.; Guenther, J.; de Nys, R. The impact and control of biofouling in marine aquaculture: A review. Biofouling 2012, 28, 649–669. [Google Scholar] [CrossRef]
- Cordell, J.R.; Levy, C.; Toft, J.D. Ecological implications of invasive tunicates associated with artificial structures in puget sound, Washington, USA. Biol. Invasions 2013, 15, 1303–1318. [Google Scholar] [CrossRef]
- Simkanin, C.; Dower, J.F.; Filip, N.; Jamieson, G.; Therriault, T.W. Biotic resistance to the infiltration of natural benthic habitats: Examining the role of predation in the distribution of the invasive ascidian Botrylloides violaceus. J. Exp. Biol. Ecol. 2013, 439, 76–83. [Google Scholar] [CrossRef]
- Simkanin, C.; Davidson, I.C.; Therriault, T.W.; Jamieson, G.; Dower, J.F. Manipulating propagule pressure to test the invasibility of subtidal marine habitats. Biol. Invasions 2017, 19, 1565–1575. [Google Scholar] [CrossRef]
- Kremer, L.P.; Rocha, R.M. The biotic resistance role of fish predation in fouling communities. Biol. Invasions 2016, 18, 3223–3237. [Google Scholar] [CrossRef]
- Qiu, J.W.; Thiyagarajan, V.; Leung, A.W.Y.; Qian, P.Y. Development of a Marine Subtidal Epibiotic Community in Hong Kong: Implications for Deployment of Artificial Reefs. Biofouling 2003, 19, 37–46. [Google Scholar] [CrossRef]
- Lezzi, M.; Del Pasqua, M.; Pierri, C.; Giangrande, A. Seasonal non-indigenous species succession in a marine macrofouling invertebrate community. Biol. Invasions 2017, 20, 937–961. [Google Scholar] [CrossRef]
- Rocha, R.M.D. Replacement of the compound ascidian species in a southeastern Brazilian fouling community. Bol. Do Inst.Ocean. 1991, 39, 141–153. [Google Scholar] [CrossRef]
- Ordóñez, V.; Pascual, M.; Fernández-Tejedor, M.; Turon, X. When invasion biology meets taxonomy: Clavelina oblonga (Ascidiacea) is an old invader in the Mediterranean Sea. Biol. Invasions 2016, 18, 1203–1215. [Google Scholar] [CrossRef]
- Thiyagarajan, V.; Qian, P.-Y. Effect of temperature, salinity and delayed attachment on development of the solitary ascidian Styela plicata (Lesueur). J. Exp. Mar. Biol. Ecol. 2003, 290, 133–146. [Google Scholar] [CrossRef]
- Anderson, D.T.; White, M.B.; Egan, A.E. The larval development and metamorphosis of the ascidians Pyurapraeputialis (Heller) and Pyurapachydermatina (Herdman) (Pleurogona, family Pyuridae). Linn. Soc. New South Wales 1976, 100, 205–217. [Google Scholar]
- Cloney, A.R. Ascidian Larvae and the Events of Metamorphosis. Am. Zoolog. 1982, 22, 817–826. [Google Scholar] [CrossRef]
- Satoh, N. Embryogenesis: Developmental Biology of Ascidians; Developmental and Cell Biology Series; Cambridge University Press: New York, NY, USA, 1994; p. 234. [Google Scholar]
- Degnan, M.B.; Souter, D.; Degnan, M.S.; Long, C.S. Induction of metamorphosis with potassium ions requires development of competence and an anterior signallingcentre in the ascidian Herdmaniamomus. Dev. Genes Evol. 1996, 206, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Chase, A.L.; Dijkstra, J.A.; Harris, L.G. The influence of substrate material on ascidian larval settlement. Mar. Poll. Bull. 2016, 106, 35–42. [Google Scholar] [CrossRef]
- Bouchemousse, S.; Bishop, J.D.D.; Viard, F. Contrasting global genetic patterns in two biologically similar, widespread and invasive Ciona species (Tunicata, Ascidiacea). Sci. Rep. 2016, 6, 24875. [Google Scholar] [CrossRef]
- Fortic, A.; Mavric, B.; Pitacco, V.; Lipej, L. Temporal changes of a fouling community: Colonization patterns of the benthic epifauna in the shallow northern Adriatic Sea. Reg. Stud. Mar. Sci. 2021, 45, 101818. [Google Scholar] [CrossRef]
- Lins, D.M.; de Marco, P., Jr.; Andrade, A.F.; Rocha, R.M. Predicting global ascidian invasions. Divers. Distrib. 2018, 24, 692–704. [Google Scholar] [CrossRef]
- Vogel, S. Life in Moving Fluids: The Physical Biology of Flow; Princeton University Press: Princeton, NJ, USA, 1996. [Google Scholar]
- Mullineaux, L.; Butman, C. Initial contact, exploration and attachment of barnacle (Balanus amphitrite) cyprids settling in flow. Mar. Biol. 1991, 110, 93–103. [Google Scholar] [CrossRef]
- Wright, J.R.; Boxshall, A.J. The influence of small-scale flow and chemical cues on the settlement of two congeneric barnacle species. Mar. Ecol. Prog. Ser. 1999, 183, 179–187. [Google Scholar] [CrossRef]
- Bologna, P.a.X.; Heck, K.L. Impacts of seagrass habitat architecture on bivalve settlement. Estuaries 2000, 23, 449–457. [Google Scholar] [CrossRef]
- Hurlbut, C. Community recruitment—Settlement and juvenile survival of 7 cooccurring species of sessile marine-invertebrates. Mar. Biol. 1991, 109, 507–515. [Google Scholar] [CrossRef]
- Walters, L.J.; Miron, G.; Bourget, E. Endoscopic observations of invertebrate larval substratum exploration and settlement. Mar. Ecol. Prog. Ser. 1999, 182, 95–108. [Google Scholar] [CrossRef]
- Osman, R.W.; Whitlatch, R.B. The influence of resident adults on recruitment: A comparison to settlement. J. Exp. Mar. Biol. Ecol. 1995, 190, 169–198. [Google Scholar] [CrossRef]
- Osman, R.W.; Whitlatch, R.B. The influence of resident adults on larval settlement: Experiments with four species of ascidians. J. Exp. Mar. Biol. Ecol. 1995, 190, 199–220. [Google Scholar] [CrossRef]
- Lemire, M.; Bourget, E. Substratum heterogeneity and complexity influence microhabitat selection of Balanus sp. and Tubulariacrocea larvae. Mar. Ecol. Prog. Ser. 1996, 135, 77–87. [Google Scholar] [CrossRef]
- Satheesh, S.; Wesley, S.G. Influence of substratum colour on the recruitment of macrofouling communities. J. Mar. Biol. Assoc. UK 2010, 90, 941–946. [Google Scholar] [CrossRef]
- Dijkstra, J.A.; Harris, L.G. Maintenance of diversity altered by a shift in dominant species: Implications for species coexistence. Mar. Ecol. Prog. Ser. 2009, 387, 71–80. [Google Scholar] [CrossRef]
- Ordóñez, V.; Rius, M.; McQuaid, C.D.; Pineda, M.C.; Pascual, M.; Turon, X. Early biotic interactions among introduced and native species reveal cryptic predation and shifts in larval behaviour. Mar. Ecol. Prog. Ser. 2013, 488, 65–79. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).