



Untangling the Characteristics and Ecological Processes of Microbial Community Assembly in the Source Area of the East Route of the South-to-North Water Diversion Project in China Under Different Water Periods

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
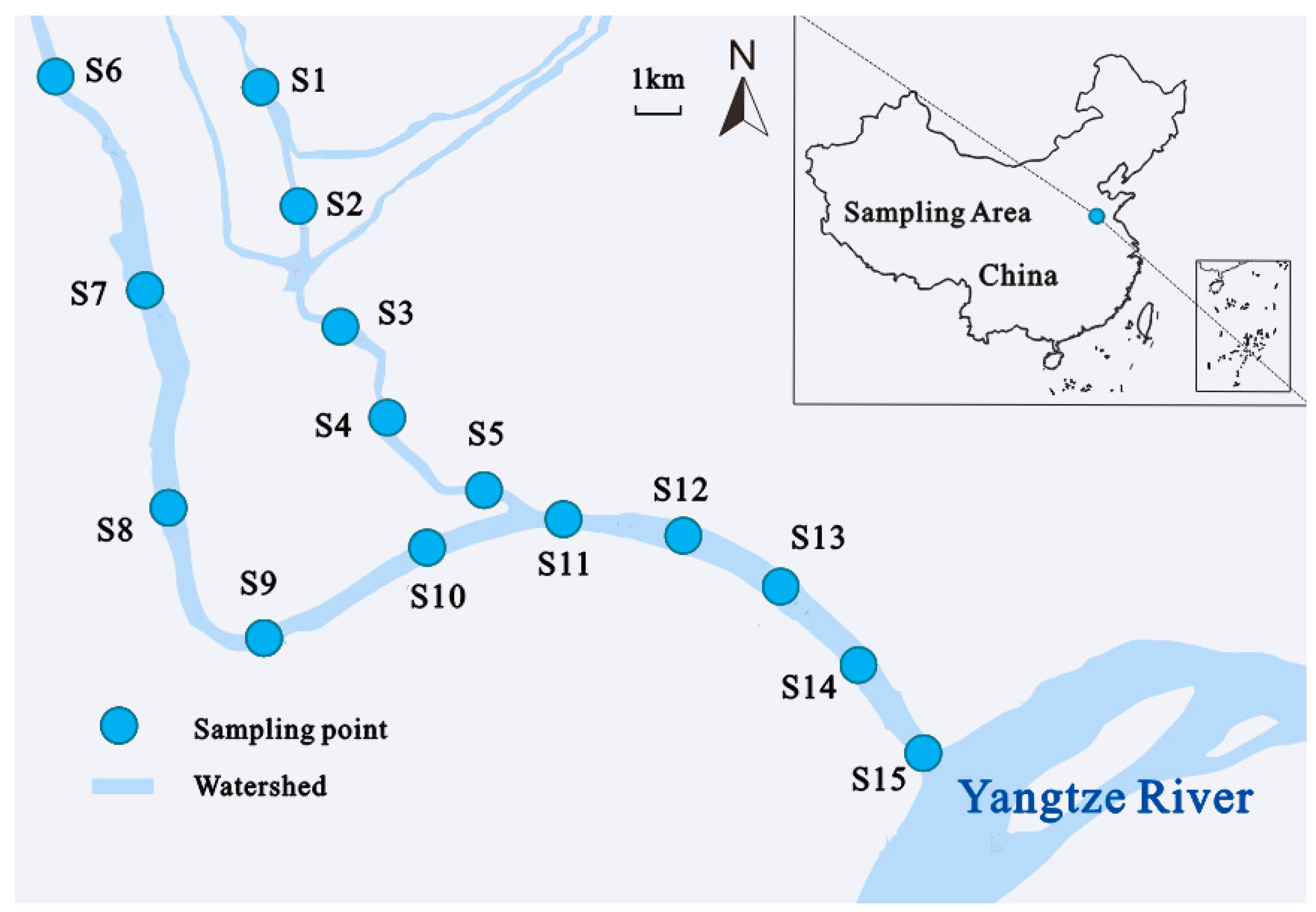
2.1. Study Area
2.2. Sample Collection
2.3. DNA Extraction, PCR Amplification, and Illumina MiSeq Sequencing
2.4. Statistical Analysis
3. Results
3.1. Species Richness, Evenness, and Diversity
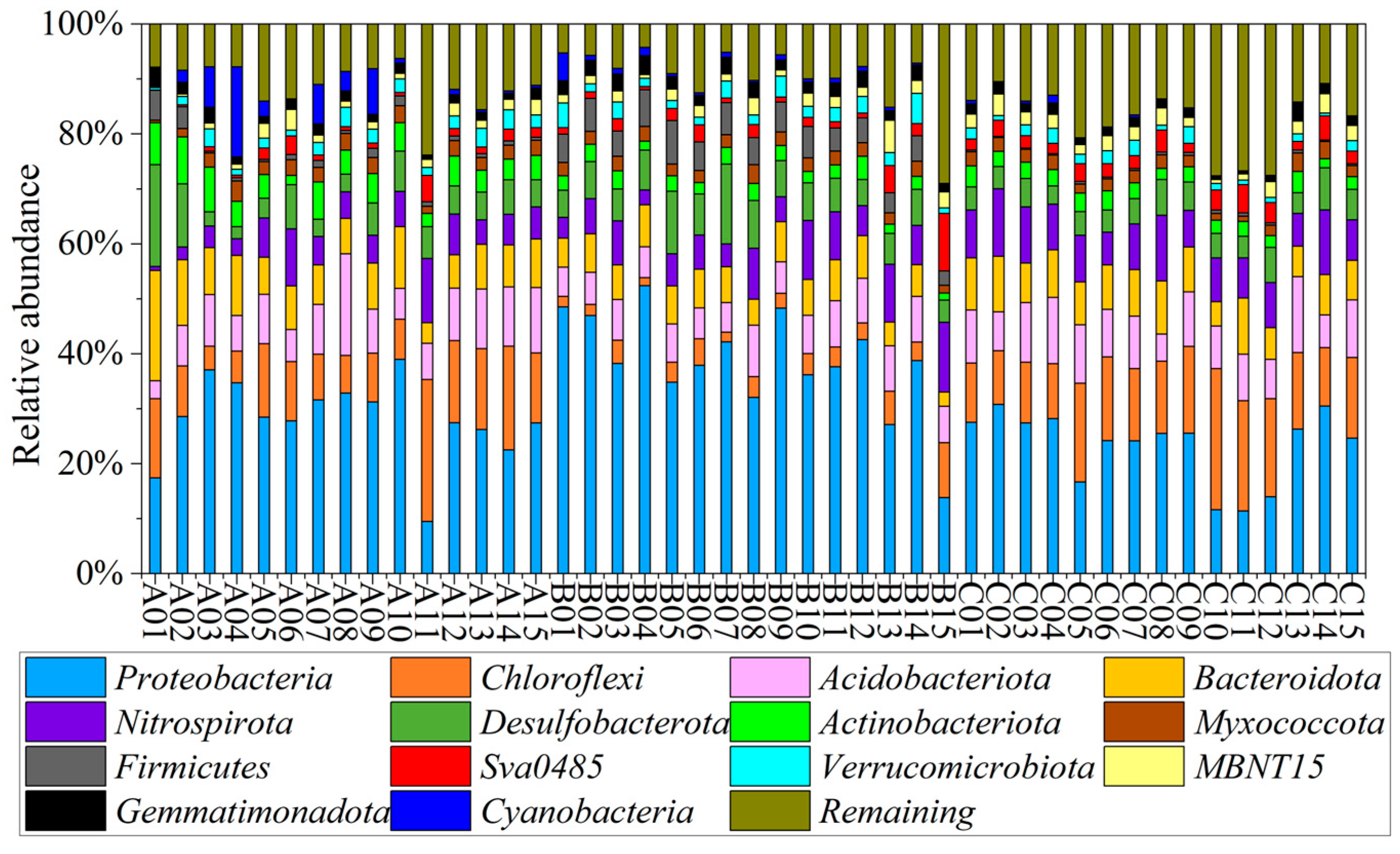
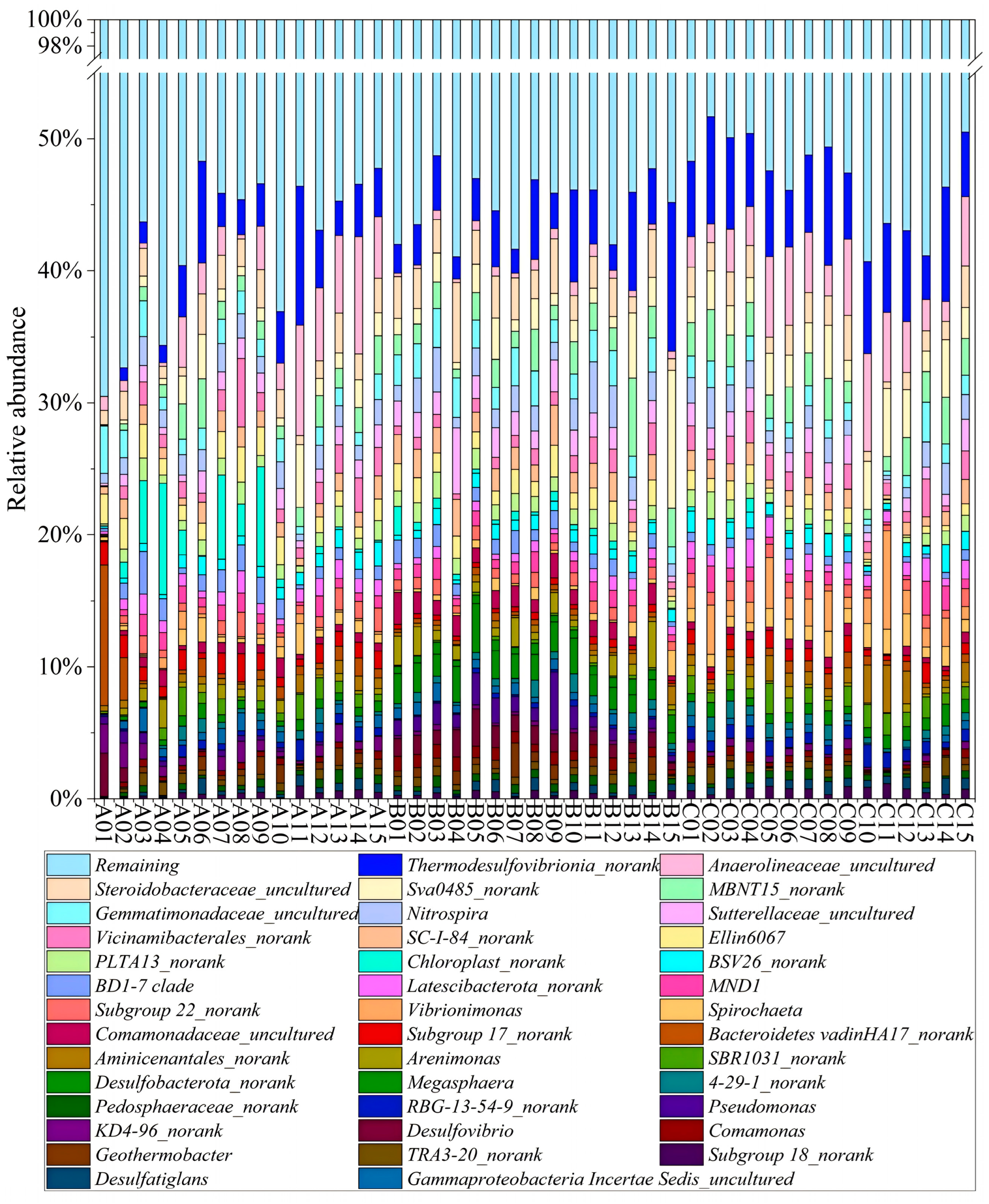
3.2. Microbial Community Composition
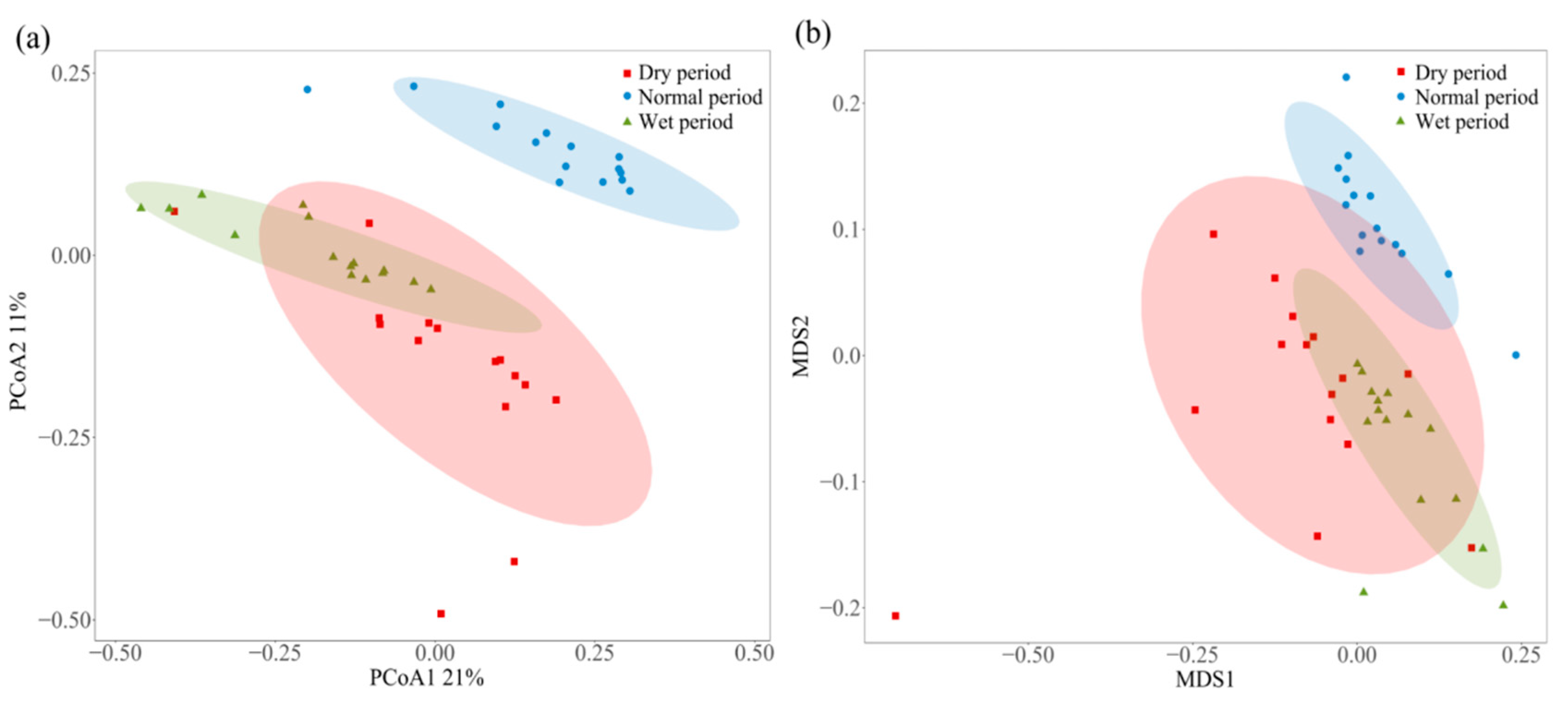
3.3. PCoA Analysis and NMDS Analysis
3.4. β-Diversity Measurement
3.5. Co-Occurrence Networks of Microbial Community
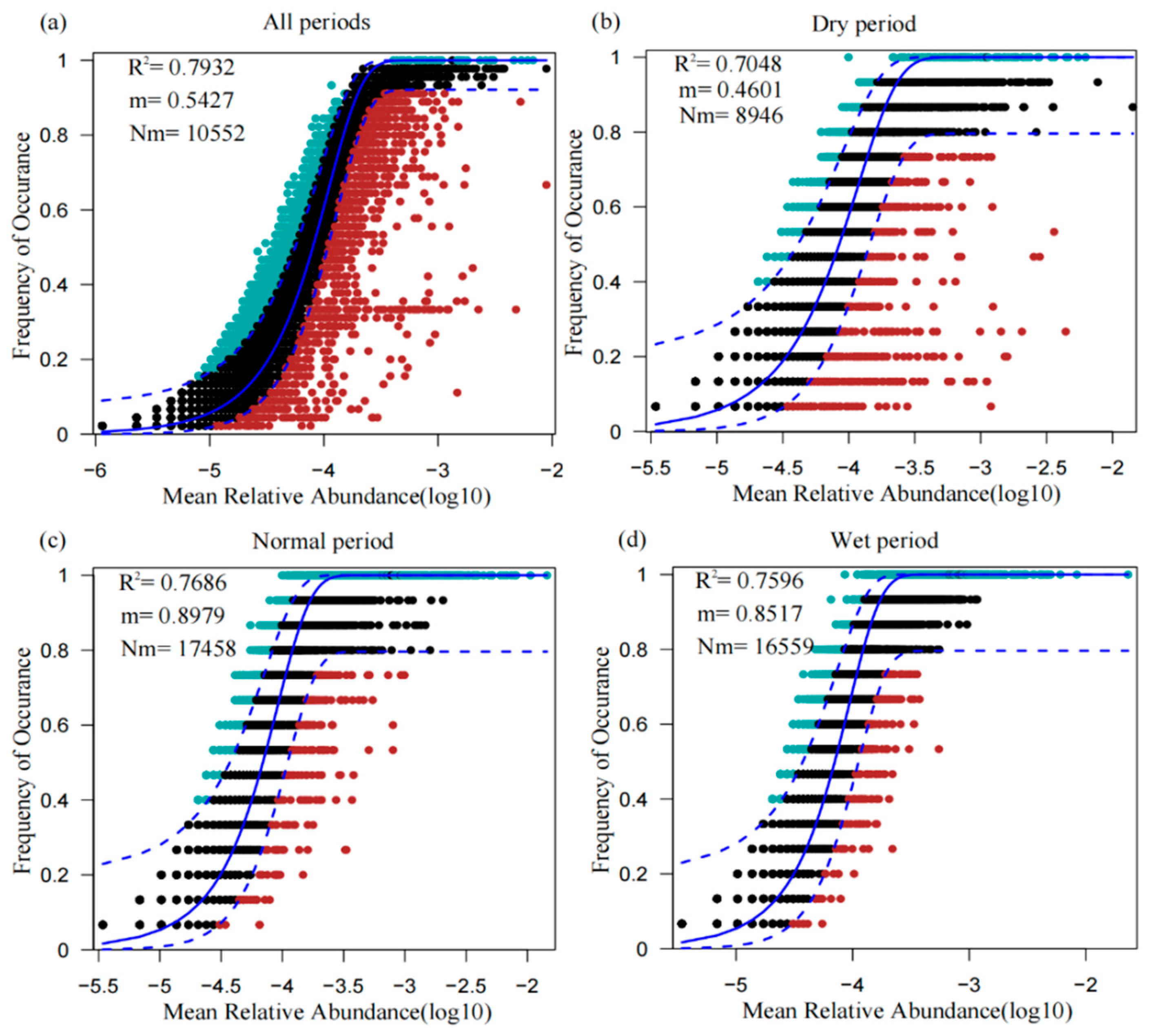
3.6. Fit to the Neutral Model of Community Assembly
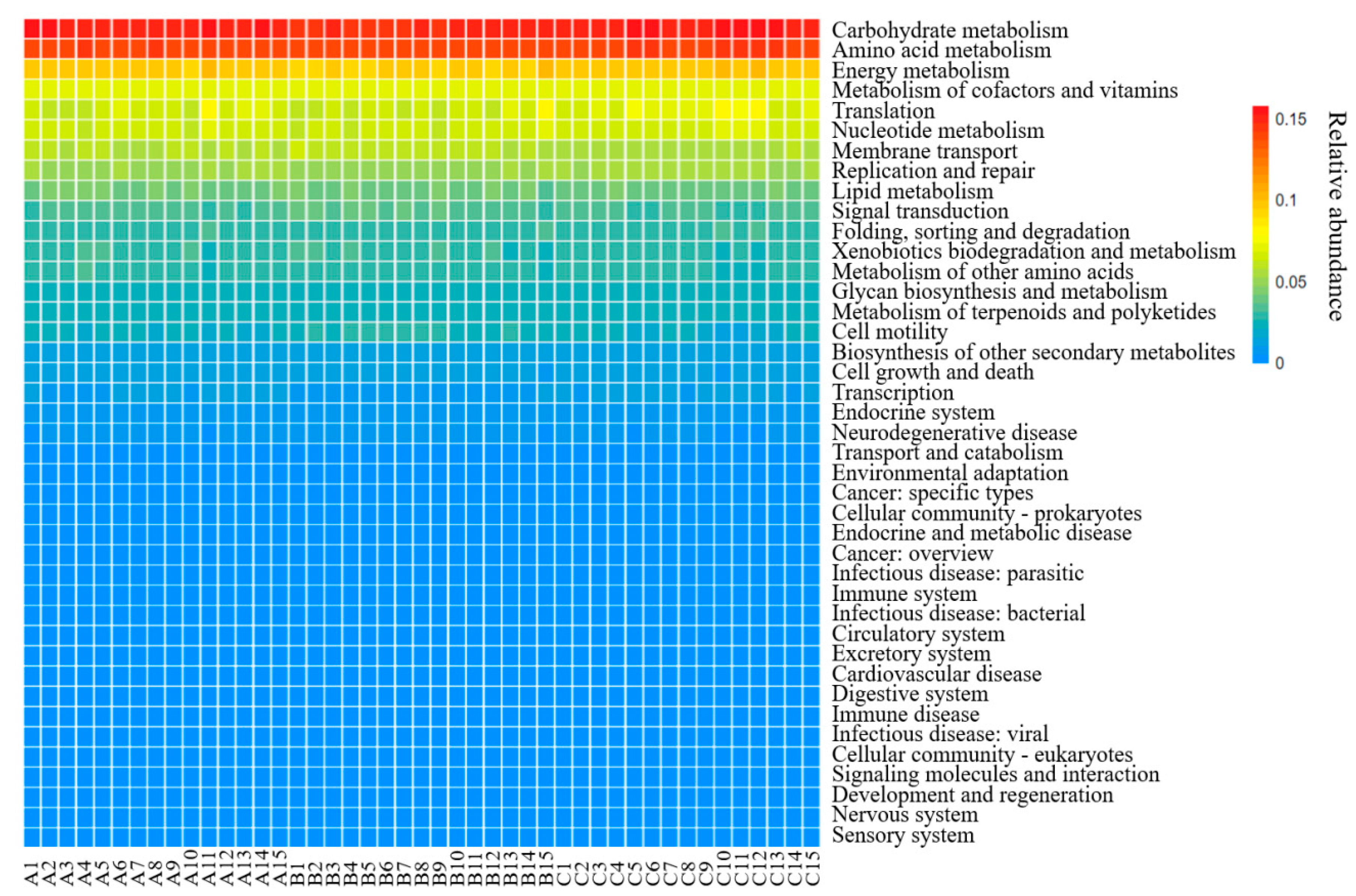
3.7. Functional Analysis from PICRUSt
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Faraji, H.; Shahryari, A. Estimation of Water Quality Index and Factors Affecting Their Changes in Groundwater Resource and Nitrate and Fluoride Risk Assessment. Water Air Soil Pollut. 2023, 234, 608. [Google Scholar] [CrossRef]
- Eliasson, J. The rising pressure of global water shortages. Nature 2015, 517, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-Y.; Zuo, J.; Zillante, G. Transformation of water resource management: A case study of the South-to-North Water Diversion project. J. Clean. Prod. 2017, 163, 136–145. [Google Scholar] [CrossRef]
- Zhuang, W. Eco-environmental impact of inter-basin water transfer projects: A review. Environ. Sci. Pollut. Res. 2016, 23, 12867–12879. [Google Scholar] [CrossRef]
- Davies, B.R.; Thoms, M.; Meador, M. An assessment of the ecological impacts of inter-basin water transfers, and their threats to river basin integrity and conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 1992, 2, 325–349. [Google Scholar] [CrossRef]
- Zhang, S.; Pang, Y.; Xu, H.; Wei, J.; Jiang, S.; Pei, H. Shift of phytoplankton assemblages in a temperate lake located on the eastern route of the South-to-North Water Diversion Project. Environ. Res. 2023, 227, 115805. [Google Scholar] [CrossRef]
- Funes, A.; de Vicente, J.; Cruz-Pizarro, L.; Álvarez-Manzaneda, I.; de Vicente, I. Magnetic microparticles as a new tool for lake restoration: A microcosm experiment for evaluating the impact on phosphorus fluxes and sedimentary phosphorus pools. Water Res. 2016, 89, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Pusch, M.; Fiebig, D.; Brettar, I.; Eisenmann, H.; Ellis, B.K.; Kaplan, L.A.; Lock, M.A.; Naegeli, M.W.; Traunspurger, W. The role of micro-organisms in the ecological connectivity of running waters. Freshw. Biol. 1998, 40, 453–495. [Google Scholar] [CrossRef]
- Mohapatra, M.; Behera, P.; Kim, J.Y.; Rastogi, G. Seasonal and spatial dynamics of bacterioplankton communities in a brackish water coastal lagoon. Sci. Total Environ. 2020, 705, 134729. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, Z.; Liu, K.; Cheng, L.; Wang, L.; Ye, A. Impacts of the eastern route of the South-to-North Water Diversion Project emergency operation on flooding and drainage in water-receiving areas: An empirical case in China. Nat. Hazards Earth Syst. Sci. 2019, 19, 555–570. [Google Scholar] [CrossRef]
- Bagra, K.; Bellanger, X.; Merlin, C.; Singh, G.; Berendonk, T.U.; Klümper, U. Environmental stress increases the invasion success of antimicrobial resistant bacteria in river microbial communities. Sci. Total Environ. 2023, 904, 166661. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Li, S.; Hou, K.; Ji, G. Spatial and seasonal bacterioplankton community dynamics in the main channel of the Middle Route of South-to-North Water Diversion Project. Res. Microbiol. 2019, 170, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Guo, F.; Xie, P.; Zhang, W.; Hu, H.; Wu, J.; Yang, Z. Differences in sediment microbial community structure and co-occurrence network in different seasons. J. Soils Sediments 2023, 23, 3539–3549. [Google Scholar] [CrossRef]
- Lv, J.; Niu, Y.; Yuan, R.; Wang, S. Different Responses of Bacterial and Archaeal Communities in River Sediments to Water Diversion and Seasonal Changes. Microorganisms 2021, 9, 782. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Jia, C.; Liu, Q.; Li, Z.; Liu, P.; Yang, M.; Zhao, M.; Li, W.; Zhu, H.; Zhang, Q. Dynamics of Bacterial Community Diversity and Structure in the Terminal Reservoir of the South-To-North Water Diversion Project in China. Water 2018, 10, 709. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, P.; Wang, C. The Influence on Contaminant Bioavailability and Microbial Abundance of Lake Hongze by the South-to-North Water Diversion Project. Int. J. Environ. Res. Public Health 2019, 16, 3068. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pan, B.; Zhu, X.; Zhao, X.; Sun, H.; He, H.; Jiang, W. Patterns of microbial communities and their relationships with water quality in a large-scale water transfer system. J. Environ. Manage. 2022, 319, 115678. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, C.; Colin, Y.; Debret, M.; Copard, Y.; Gardes, T.; Jacq, K.; Ayrault, S.; Berthe, T. Shifts in sediment bacterial communities reflect changes in depositional environments in a fluviatile context. Sci. Total Environ. 2023, 885, 163890. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Liu, J.; Sun, X.; Zuo, Y.; Hu, Q.; He, X. Plant species shape the bacterial communities on the phyllosphere in a hyper-arid desert. Microbiol. Res. 2023, 269, 127314. [Google Scholar] [CrossRef] [PubMed]
- Newton Ryan, J.; Jones Stuart, E.; Eiler, A.; McMahon Katherine, D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [PubMed]
- Colatriano, D.; Tran, P.Q.; Guéguen, C.; Williams, W.J.; Lovejoy, C.; Walsh, D.A. Genomic evidence for the degradation of terrestrial organic matter by pelagic Arctic Ocean Chloroflexi bacteria. Commun. Biol. 2018, 1, 90. [Google Scholar] [CrossRef] [PubMed]
- Roberto, A.A.; Van Gray, J.B.; Leff, L.G. Sediment bacteria in an urban stream: Spatiotemporal patterns in community composition. Water Res. 2018, 134, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.J.; Tate Iii, R.L. Effects of Heavy Metal Contamination and Remediation on Soil Microbial Communities in the Vicinity of a Zinc Smelter. J. Environ. Qual. 1998, 27, 609–617. [Google Scholar] [CrossRef]
- Abiriga, D.; Jenkins, A.; Klempe, H. Microbial assembly and co-occurrence network in an aquifer under press perturbation. Ann. Microbiol. 2022, 72, 41. [Google Scholar] [CrossRef]
- Santos, A.; Rachid, C.; Pacheco, A.B.; Magalhães, V. Biotic and abiotic factors affect microcystin-LR concentrations in water/sediment interface. Microbiol. Res. 2020, 236, 126452. [Google Scholar] [CrossRef] [PubMed]
- Meziti, A.; Tsementzi, D.; Rodriguez-R, L.M.; Hatt, J.K.; Karayanni, H.; Kormas, K.A.; Konstantinidis, K.T. Quantifying the changes in genetic diversity within sequence-discrete bacterial populations across a spatial and temporal riverine gradient. ISME J. 2018, 13, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, Z.; Sun, Q.; Ding, Y.; Ding, Z.; Sun, L. The spatial and seasonal variations of bacterial community structure and influencing factors in river sediments. J. Environ. Manag. 2019, 248, 109293. [Google Scholar] [CrossRef]
- Ma, X.; Li, Y.; Niu, L.; Shang, J.; Yang, N. Microbial community structure and denitrification responses to cascade low-head dams and their contribution to eutrophication in urban rivers. Environ. Res. 2023, 221, 115242. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.; Yager, P.L.; Moran, M.A.; Coles, V.J.; Fortunato, C.S.; Krusche, A.V.; Medeiros, P.M.; Payet, J.P.; Richey, J.E.; Satinsky, B.M.; et al. Bacterial Biogeography across the Amazon River-Ocean Continuum. Front. Microbiol. 2017, 8, 882. [Google Scholar] [CrossRef]
- Baldassi, A.C.; Balbuena, T.S. The Eucalyptus grandis chloroplast proteome: Seasonal variations in leaf development. PLoS ONE 2022, 17, e0265134. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Yuan, R.; Wang, S. Water diversion induces more changes in bacterial and archaeal communities of river sediments than seasonality. J. Environ. Manag. 2021, 293, 112876. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.J.; Theis, K.R.; Uzarski, D.G.; Learman, D.R. Microbial community structure and microbial networks correspond to nutrient gradients within coastal wetlands of the Laurentian Great Lakes. FEMS Microbiol. Ecol. 2019, 95, fiz033. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Network Features | Dry Period | Normal Period | Wet Period |
|---|---|---|---|
| Total nodes | 485 | 343 | 650 |
| Todal edges | 845 | 323 | 1197 |
| Positive correlations | 808 (95.62%) | 261 (80.80%) | 923 (77.11%) |
| Negative correlations | 37 (4.38%) | 62 (19.20%) | 274 (274) |
| Average clustering coefficient (avgCC) | 0.298 | 0.284 | 0.303 |
| Average degree (avgK) | 3.48 | 1.883 | 3.683 |
| Average path distance (GD) | 6.691 | 7.927 | 5.547 |
| Density | 0.007 | 0.006 | 0.006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, W.; Wen, X.; Zhao, Y.; Wu, X.; Zheng, H.; Chen, J.; Hu, Z.; Zhong, Q.; Wu, J. Untangling the Characteristics and Ecological Processes of Microbial Community Assembly in the Source Area of the East Route of the South-to-North Water Diversion Project in China Under Different Water Periods. Water 2025, 17, 649. https://doi.org/10.3390/w17050649
Cai W, Wen X, Zhao Y, Wu X, Zheng H, Chen J, Hu Z, Zhong Q, Wu J. Untangling the Characteristics and Ecological Processes of Microbial Community Assembly in the Source Area of the East Route of the South-to-North Water Diversion Project in China Under Different Water Periods. Water. 2025; 17(5):649. https://doi.org/10.3390/w17050649
Chicago/Turabian StyleCai, Wei, Xin Wen, Yueru Zhao, Xiusen Wu, Haoran Zheng, Jiangtao Chen, Zhengyang Hu, Qin Zhong, and Jun Wu. 2025. "Untangling the Characteristics and Ecological Processes of Microbial Community Assembly in the Source Area of the East Route of the South-to-North Water Diversion Project in China Under Different Water Periods" Water 17, no. 5: 649. https://doi.org/10.3390/w17050649
APA StyleCai, W., Wen, X., Zhao, Y., Wu, X., Zheng, H., Chen, J., Hu, Z., Zhong, Q., & Wu, J. (2025). Untangling the Characteristics and Ecological Processes of Microbial Community Assembly in the Source Area of the East Route of the South-to-North Water Diversion Project in China Under Different Water Periods. Water, 17(5), 649. https://doi.org/10.3390/w17050649