Abstract
Harmful Algal Blooms (HABs) are a growing environmental concern due to their adverse impacts on aquatic ecosystems, human health, and economic activities. These blooms are driven by a combination of factors, including nutrient enrichment, environmental factors, and hydrological conditions, leading to the excessive growth of algae. HABs produce toxins that threaten aquatic biodiversity, contaminate drinking water, and cause economic losses in fisheries and tourism. The causes of HABs are multifaceted, involving interactions between environmental factors such as temperature, light availability, and nutrient levels. Agricultural runoff, wastewater discharge, and industrial pollution introduce excessive nitrogen and phosphorus into water bodies, fueling bloom formation. Climate change further exacerbates the problem by altering precipitation patterns, increasing water temperatures, and intensifying coastal upwelling events, all of which create favorable conditions for HAB proliferation. This review explores the causes, ecological consequences, and potential mitigation strategies for HABs. Effective monitoring and detection methods, including satellite remote sensing, molecular biotechnology, and artificial intelligence-driven predictive models, offer promising avenues for early intervention. Sustainable management strategies such as nutrient load reductions, bioremediation, and regulatory policies can help mitigate the adverse effects of HABs. Public awareness and community involvement also play a crucial role in preventing and managing HAB events by promoting responsible agricultural practices, reducing waste discharge, and supporting conservation efforts. By examining existing literature and case studies, this study underscores the urgent need for comprehensive and interdisciplinary approaches to regulate HABs.
1. Introduction
Harmful Algal Blooms (HABs) are rapid, excessive growths of algae or, in some cases, cyanobacteria in freshwater or aquatic environments that accumulate on or just below the water’s surface. These blooms can produce toxins, deplete oxygen, discolor water, and pose serious risks to humans, wildlife, ecosystems, and local economies. Their occurrence is driven by a combination of nutrient enrichment (especially nitrogen and phosphorus), favorable hydrological conditions, and climatic factors, and is becoming more frequent and intense under changing environmental conditions [1]. The term ‘bloom’ refers to the rapid increase and subsequent decrease of phytoplankton populations [2]. The term ‘harmful’ refers to the negative impact that the microalgal species have on aquatic organisms and ecosystems. Overall, the occurrence of HABs will cause a decrease in oxygen content and water transparency, an increase in algal toxins, and the death of aquatic organisms [3,4,5,6].
Algae use carbon dioxide and water to synthesize organic food molecules through the process of photosynthesis [7]. During this process, they also release oxygen as a by-product. It is estimated that algal organisms are responsible for producing between 30% and 50% of the global oxygen supply necessary for respiration in humans and other land animals [8]. True algae are eukaryotic organisms that contain nuclei, chloroplasts, mitochondria, and other membrane-bound organelles, but prokaryotes, such as cyanobacteria, which lack a nucleus, are also often referred to as algae. Their cell walls may be composed of cellulose, silica (as in diatoms), or other materials, although some algae lack a rigid outer wall [9].
HABs can profoundly alter trophic dynamics within aquatic ecosystems, primarily driven by nutrient over-enrichment. These nutrient surpluses can disrupt food web structures in multiple ways. For example, HABs often outcompete typical primary producers, such as zooplankton, ultimately leading to a decline in biodiversity. As herbivores depend on these primary producers for sustenance, their populations may also decline, reducing grazing pressure and causing ripple effects throughout the ecosystem. In addition to toxins generated by HABs, the toxins carried by stormwater runoffs [10] can bioaccumulate within the tissues of aquatic organisms, posing risks up the food chain [9].
The problem of HABs in U.S. coastal waters has been recognized for centuries, dating back to reports from early explorers and settlers who suffered poisoning from consuming contaminated seafood [7]. HABs are linked to various human poisoning syndromes through the consumption of molluscs, crustaceans, fish, and other aquatic species that bioaccumulate algal toxins. Besides ingestion, humans can be exposed to these toxins through inhalation of aerosolized particles, direct contact with contaminated waters, or skin exposure during bloom events, all of which pose serious health risks [7].
The ecological consequences of HABs extend beyond food competition. Many harmful algae release potent toxins and bioactive compounds into surrounding waters, leading to the widespread mortality of aquatic plants and animals. Predators, such as large fish and seabirds, that consume smaller, contaminated prey may also accumulate these toxins, making them hazardous throughout the food web. The decomposition of HAB biomass can deplete oxygen levels, leading to hypoxic conditions detrimental to species that rely on well-oxygenated waters. According to Anderson et al. [7], fish mortalities can result from either oxygen depletion or direct exposure to algal toxins, with clinical symptoms including erratic swimming, paralysis, respiratory failure, and death.
An example of the devastating impact of HABs occurred in May 1991, when a major bloom in Vestfjorden and adjacent areas of Northern Norway led to the loss of approximately 743 tons of farmed salmon [11,12]. HAB-related fish kills have also been observed to affect broader wildlife populations, including seabirds, whales, dolphins, and other aquatic mammals. These toxins can be transmitted throughout the food web either by ingestion or through gill uptake [13,14].
Moreover, HABs often result in dense biomass accumulation, which reduces light penetration through the water column. This decrease in light availability can negatively impact seagrass beds, essential nurseries for juvenile fish and shellfish, thus further disrupting coastal ecosystems [7]. Even blooms of non-toxic species can cause ecological harm; when they die off, bacterial decomposition consumes oxygen, leading to hypoxia and stressing both plant and animal communities. Furthermore, the breakdown of large algal blooms can result in elevated nutrient inputs, further exacerbating environmental degradation [7].
Among the different types of algae associated with HABs, cyanobacteria, dinoflagellates, and diatoms are the most frequently observed. Cyanobacteria, often called blue-green algae, typically inhabit freshwater ecosystems like lakes and ponds, while dinoflagellates and diatoms are more commonly found in aquatic environments such as oceans and bays [15].
Cyanobacteria are photosynthetic bacteria that utilize sunlight to generate energy, playing a significant role in global carbon and nitrogen cycles [16]. As noted in a study conducted by Singh et al. [17], Cyanobacteria contribute to the mobility of essential minerals, influence soil structure and moisture retention, and are capable of thriving in both aquatic and damp terrestrial environments. These microorganisms can sometimes produce harmful toxins or proliferate excessively, adversely affecting aquatic ecosystems and human health. Some of the primary toxins produced by cyanobacteria include microcystin, cylindrospermopsin, anatoxin, guanitoxin, saxitoxin, nodularin, and lyngbyatoxin [18]. Additionally, cyanobacteria are recognized for their ability to fix atmospheric nitrogen, aid in soil remediation, and impact the physiological and biochemical properties of their associated host plants. Their presence in freshwater poses significant concerns for drinking water supplies, as HABs have the potential to release cyanotoxins as well as taste and odor (T&O) compounds such as geosmin and 2-methylisoborneol (MIB) [19].
Dinoflagellates and diatoms, sometimes referred to as microalgae, are two diverse varieties of phytoplankton [15]. These are widely known for causing red tides. These algal species can also produce harmful toxins or proliferate excessively. The primary toxins produced by diatoms and dinoflagellates include brevetoxin, azaspiracid, ciguatoxins, okadic acid, saxitoxin, and domoic acid [15]. These toxins can lead to various forms of shellfish poisoning and negatively impact their surrounding environments [20]. Diatoms are unicellular algae with microscopic silica shells that are highly resistant to most acids, body decomposition, and heat. They often form colonies and grow abundantly [21]. Despite the ballast effect of their silica shells, diatoms are known to regulate buoyancy [22].
Although our understanding of HABs has advanced considerably, important research gaps persist. Current methodologies often fail to fully integrate key climate factors such as temperature variability, stratification of the water column, changes in light availability, ocean acidification, and nutrient influx driven by altered precipitation patterns that collectively influence HAB development. Furthermore, there is a pressing need for experimental designs that extend beyond traditional disciplinary and comparative approaches [23]. Given the rising frequency of HAB occurrences and their proven threats to aquatic ecosystems and public health, addressing these research deficiencies is essential. Therefore, this paper aims to investigate the underlying causes and ecological impacts of HABs, review current detection and monitoring techniques, and underscore the critical importance of developing effective mitigation strategies.
This paper is a review of HABs, including their causes, effects, monitoring, and prevention methods. The primary objective of this review is to comprehensively examine the causes, ecological and economic impacts, monitoring techniques, and mitigation strategies related to HABs. This study aims to identify existing knowledge gaps within the current body of research, emphasize the growing frequency and severity of HAB events, and highlight the urgent need for interdisciplinary approaches in HAB prediction, detection, and management. By synthesizing recent literature and case studies, the review seeks to provide a foundation for advancing future research and informing effective policy and practice.
2. Methodology
The methodology employed in this review involved a systematic literature search using Google Scholar and Scopus. Key search terms included “Harmful Algal Blooms”, “HAB”, “HAB impacts”, “HAB monitoring”, “HAB detection”, “HAB mitigation”, and “HAB awareness”. Articles were initially screened based on their abstracts to assess relevance to these themes. Only peer-reviewed studies, official reports, and scientific reviews that directly addressed HAB mechanisms, impacts, and management practices were selected for inclusion. Sources deemed irrelevant or lacking sufficient detail were excluded. The selected literature was analyzed to synthesize the current understanding of HAB dynamics and to identify prevailing trends, technological advancements, and policy approaches in HAB research and management.
3. HAB Causes
3.1. Environmental Conditions
3.1.1. Light
Light is a crucial factor influencing the physiological processes and growth of algae. Previous studies have shown that under low light conditions, algae experience significant impairments in their growth and reproduction [23,24,25]. To cope with limited light availability, algae adjust by modifying their nutrient metabolism and storage patterns [26]. Research has further demonstrated that the proliferation of green tide algae is highly sensitive to changes in light intensity [27].
The findings suggest that higher light intensities accelerate the inactivation of algal cells (approximately 90%), while even minimal light can significantly alter cellular physiological activity [27]. As light intensity increases, the time needed to deactivate algal cells decreases substantially. When light levels become excessively high, algae are unable to adequately adjust to the external conditions, negatively affecting their seedling growth and maturation [27].
Additionally, a study conducted in 2022 by Li et al. [28] examined how different levels of visible light irradiation influence algal growth during the process of removing algae from water bodies. Their results indicated that light exposure alone rapidly diminished algal viability, allowing little to no time for cellular adaptation. Overall, light intensity plays a key role in the development, persistence, and toxicity of HABs. In conjunction with nutrient enrichment and climate change, increased light availability can exacerbate the frequency and severity of HABs.
3.1.2. Temperature
Temperature is a key factor driving the dynamics of HABs, influencing their growth rates, duration, toxin production, and species composition. With ongoing climate change and rising global temperatures, HAB events are becoming increasingly frequent and severe. Research conducted in 2020 by Ho and Michalak [29] found that higher spring air temperatures were correlated with reduced microcystin concentrations, suggesting that while warmer conditions may extend the growing season and boost cyanobacterial abundance, they could simultaneously lead to lower toxin production. Observations have also indicated that longer summers are associated with shifts in phytoplankton communities, favoring nontoxic taxa [30]. Regarding total abundance, there is evidence that elevated temperatures enhance the growth rates of Microcystis aeruginosa, although the resulting strains may be less toxic [31]. It is further suggested that the increasing threat posed by future blooms may stem from higher biomass or the dominance of certain species, rather than heightened toxicity levels [29]. Overall, rising temperatures intensify HABs by promoting algal proliferation, extending bloom periods, altering species distributions, and potentially affecting toxicity profiles. As the increase in temperature continues, HABs are expected to become more frequent, widespread, and ecologically and socially disruptive.
3.1.3. Hydrodynamic Factors
The influence of hydrodynamic conditions on algal cells can be broadly categorized into two types: direct and indirect effects (Figure 1). Direct impacts include factors such as flow velocity, water turbulence, and shear stress, whereas indirect effects involve water retention, stratification, nutrient distribution, and light availability. Flow velocity, which measures how quickly water moves, plays a critical role in algal transport and aggregation. High flow velocities disrupt algal clumping and inhibit bloom formation, while moderate or low velocities tend to favor algal bloom development [31,32,33,34,35].
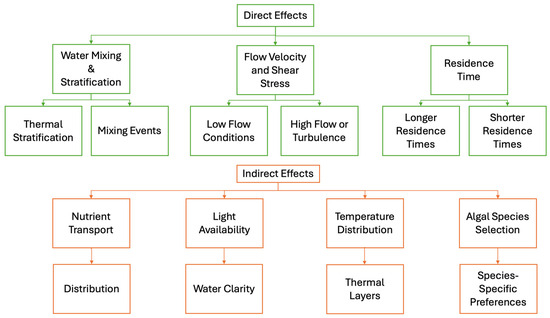
Figure 1.
Flow diagram summarizing the mechanisms by which hydrodynamic conditions affect algal blooms in reservoirs, focusing on both direct and indirect effects.
Research has shown that moderate water turbulence enhances algal growth by improving nutrient uptake and energy metabolism [36,37,38,39,40]. However, excessive turbulence damages algal cells, disrupts colony formation, and inhibits growth [41,42,43]. The complexity of turbulence, due to overlapping physical processes, makes its precise role challenging to fully delineate. Small-scale turbulence at the water surface has been found to significantly contribute to the formation of Microcystis surface scums [44]. Additionally, turbulence influences predator-prey interaction rates, thereby affecting the growth of zooplankton, mixotrophic algae, and autotrophic algae [27,44,45]. Water retention time is another critical factor influencing algal biomass. Studies have confirmed that extended water retention promotes bloom development, while enhanced water exchange, such as through reservoir operations, can suppress blooms [46,47,48,49,50,51].
Stratification of the water column, by reducing vertical mixing and impeding material exchange, allows algae to concentrate near the surface where light is more abundant, thus, supporting their proliferation [45,52,53]. Stratified waters, often associated with extended residence times and low flow velocities, further enhance conditions favorable to bloom formation [50,54].
Nutrients such as nitrogen, phosphorus, and silica are essential for algal blooms. Hydrodynamic processes can modify nutrient transport and release, thereby influencing nutrient availability for algal cells [42,55,56]. For example, enhanced nutrient delivery from tributaries to main water bodies has been shown to increase the likelihood of spring blooms. In reservoirs, nutrient release from sediments, especially under low water levels and strong bottom turbulence, plays a major role in promoting algal growth [52]. Because hydrodynamic and nutrient cycling processes vary between water bodies, case-specific studies are needed to fully understand bloom dynamics.
Finally, light distribution is a dominant driver of algal bloom development, as light is crucial for photosynthesis. Hydrodynamic mixing can expose algal cells to higher irradiance levels, stimulating growth [35,57,58,59].
3.1.4. Climate Change
According to the Environmental Protection Agency (EPA), several climate change-driven factors can influence the occurrence and severity of HABs, including rising water temperatures, changes in salinity, elevated carbon dioxide (CO2) levels, altered rainfall patterns, sea level rise, and shifts in coastal upwelling dynamics [60].
Warmer water temperatures, in particular, create favorable conditions for cyanobacterial HABs (cyanoHABs), which thrive in warm, slow-moving waters. As climate change drives global temperature increase/rise, both magnitude and duration of cyanoHABs are expected to rise. Elevated temperatures also enable cyanobacteria to migrate within the water column, drawing nutrients from cooler, deeper layers and using them to fuel growth in the sunlit, warmer surface layers due to the presence or enhancement of stratification. Furthermore, cyanoHABs absorb sunlight and release heat, which in turn raises surface water temperatures even further, creating a feedback loop that further favors their proliferation [61].
Climate change is also projected to alter salinity levels. In some regions, decreased freshwater runoff caused by intensified droughts and increased evaporation will lead to higher salinity in water bodies. In other places affected by snowfall and freezing rain, the increased use of road salt leads to increased salinity downstream in the watershed [62]. Elevated salinity levels can damage agriculture by affecting irrigation systems, harming freshwater crops, allowing salt-tolerant and aquatic algae to invade inland lakes, and contaminating drinking water supplies.
Higher atmospheric and aquatic CO2 concentrations, another consequence of climate change, can accelerate algal growth by providing more carbon for photosynthesis. At the same time, increased CO2 levels acidify water bodies, altering species competition among algae and impacting organisms that graze on them [61].
Elevated salinity levels can cause significant physiological stress in algal cells. Increased salt concentrations may lead to the leakage of cellular contents by accelerating the excretion processes. Salinity also impacts algal growth by altering the osmotic pressure of the surrounding environment, which can cause contraction of the internal membrane structure and disrupt membrane transport functions. Furthermore, changes in salinity influence the solubility of intracellular gases like CO2 and O2, ultimately decreasing metabolic activity and slowing algal growth [63].
When salinity becomes excessively high, algal cells may experience a rapid efflux of water, leading to elevated internal salt concentrations. This disrupts cellular metabolism, impairs nutrient absorption, and negatively affects the utilization of nutrients within algal cells [64]. Additionally, high salinity can impact the pigment composition of certain algal species, leading to reduced photosynthetic activity [39,65].
Rainfall patterns, already being disrupted by climate change, are expected to become more extreme, with heavier rainfall events and prolonged droughts. Intense rainfall increases nutrient-rich runoff from land into aquatic systems, fueling HABs. Additionally, large volumes of freshwater runoff can stratify coastal waters by layering lighter freshwater over denser saltwater, inhibiting oxygen mixing. It is important to note that in inland freshwater systems, inflowing water can be denser than lake water. As HABs die and sink, their decomposition depletes oxygen in bottom waters, forming hypoxic (low-oxygen) zones that can kill aquatic organisms that are dependent on oxygen. Enhanced freshwater flows may also flush HABs and associated toxins into aquatic ecosystems, worsening existing blooms or promoting new ones [61].
Sea level rise contributes by expanding areas of shallow, stable coastal waters, creating ideal conditions for HAB development. Finally, climate change is expected to intensify and alter the timing of coastal upwelling events. During upwelling, surface waters are pushed offshore by winds, allowing nutrient-rich deep waters to rise. These nutrient surges can stimulate algal growth, particularly HABs, especially when combined with increased nutrient runoff from terrestrial sources [61].
3.1.5. Nutrient Enrichment Caused by Agricultural Practices
Nutrients play a major role in promoting HABs across a wide range of aquatic ecosystems. The three primary nutrients associated with HAB development are nitrogen, phosphorus, and silicon (Table 1). Their influence has been observed in freshwater systems, coastal regions, oceanic environments, coastal lagoons, and estuaries.

Table 1.
Nutrients Depicting their HAB Role, Processes/Conditions, and Dominant Algal Species Types.
Nitrogen significantly stimulates algal growth when available in excess and is often the limiting nutrient in aquatic systems. In coastal areas, nitrogen inputs are typically elevated due to agricultural runoff, urban wastewater discharge, and atmospheric deposition. Consequently, HAB occurrences in these regions are frequently concentrated near estuaries and river mouths. In contrast, nitrogen concentrations in open ocean environments are generally low, with inputs mainly supplied through upwelling or atmospheric deposition [66]. A study of two freshwater lakes in Quebec found that a pulse of nitrogen was associated with an increased presence of Microcystis and increased microcystin concentrations [67].
While phosphorus is less commonly a limiting nutrient in aquatic settings compared to nitrogen, it can still enhance HAB formation. Coastal lagoons and estuaries receiving high phosphorus loads from agricultural runoffs and sewage are particularly vulnerable. In freshwater ecosystems, phosphorus-driven blooms are especially pronounced when nitrogen concentrations are already elevated [66].
Silicon is another critical nutrient, primarily essential for the growth of diatoms. Diatoms often dominate coastal upwelling regions and riverine systems, where silicon availability directly influences their population dynamics and shapes the overall phytoplankton community structure. However, when silicon becomes depleted, other harmful algal groups—such as dinoflagellates and cyanobacteria—may gain a competitive advantage, leading to an increased risk of HABs [66].
Agricultural runoff is recognized as a major source of nutrient enrichment in aquatic ecosystems, introducing substantial amounts of nitrogen and phosphorus. These nutrients, common components of both organic and inorganic fertilizers, become mobilized during rainfall events and are carried via surface runoff into nearby aquatic systems [75,76,77,78,79]. The global demand for nitrogen- and phosphorus-based fertilizers continues to rise. According to the Food and Agriculture Organization (FAO) in 2019, global nitrogen fertilizer consumption increased by approximately 1–2% annually [80]. The widespread use of inorganic fertilizers further elevates nitrogen and phosphorus concentrations in soils, and nutrients not absorbed by crops are highly susceptible to runoff during rainfall or irrigation events, often traveling through drainage systems into aquatic environments [66].
Beyond fertilizer application, animal manure represents another significant source of agricultural nutrient runoff. The expansion of global livestock production has been linked to increased aquatic eutrophication. Industrialized farming operations, including concentrated animal feeding operations (CAFOs), produce large volumes of manure rich in nitrogen and phosphorus. When manure is improperly stored, over-applied to fields, or exposed to heavy rainfall, these nutrients can readily leach into waterways through surface runoff [66].
The extent and composition of nutrient runoff from agricultural lands are influenced by crop type, management practices, and environmental conditions. Certain agricultural practices exacerbate nutrient runoff, such as conventional tillage, improper fertilizer application timing, and inefficient irrigation. Tillage disturbs the soil, promotes erosion, and enhances surface runoff [81], increasing nutrient loss. Similarly, applying fertilizers during non-growing seasons or immediately before heavy rain can lead to substantial nutrient runoff. Poor irrigation practices, especially excessive or insufficient/inadequate watering, can contribute to significant nitrogen leaching and surface runoff. Regional factors such as high rainfall patterns and specific soil types can further amplify these nutrient losses [66].
Residential development also poses a challenge for algal blooms. In rural or marginalized communities without sanitary sewer systems, septic systems are common and represent about one-quarter of U.S. households [82]. Septic systems are privately owned and require regular owner maintenance every 3–5 years [82]. In North Carolina, for example, where half of homes use septic systems, improper maintenance, poorly-draining clay soils, and rising water tables lead to an annual septic system failure rate of 10–20% (likely underreported), leading to sewage appearing in backed up appliances, the soil surface, or in the groundwater [82]. The decentralized nature and private ownership of these results in a watershed management challenge. Of concern is not only the fecal pathogens that pose a hazard to health, but also the unmitigated nutrients that enter the environment [83]. A study by Brewton et al., in 2022 [83] used multiple tracers to confirm that in Florida, where septic systems represent 33% of systems and more than 50% of domestic wastewater, septic system release to groundwater leads to surface water contamination with nutrients and HABs.
4. Impacts of HABs
4.1. Ecological
The ecological impacts of HABs include oxygen depletion, toxin production and bioaccumulation, disruption of food webs, and habitat degradation. When HABs die, their decomposition consumes large amounts of oxygen, which leads to hypoxic (low oxygen) or anoxic (no oxygen) zones in the system. Such low oxygen levels may cause mass mortality of fish, shellfish, and other aquatic organisms due to suffocation. HABs produce toxins that can be released into the marine ecosystem and can even result in bioaccumulation. Algae produce toxins that can be released into the aquatic ecosystem and can even result in bioaccumulation. HABs also disrupt food webs when they outcompete phytoplankton, which reduces the availability for filter feeders such as oysters and mussels. These toxins will then accumulate up the food chain, ultimately affecting the top predators like dolphins, seabirds, and sharks.
Additionally, HABs can cause degradation of habitats. HABs shade out seagrasses and corals, which reduces light penetration and leads to habitat loss. Even more so, some HAB toxins are directly harmful to coral reefs and can contribute to coral bleaching and reef degradation. Since HABs can elevate CO2 levels, leading to increased water acidity and altered chemistry, they can harm aquatic organisms while also outcompeting non-toxic plankton, ultimately resulting in a loss of biodiversity. HAB species can outcompete non-toxic plankton, which will result in a loss of biodiversity. Some HABs can even fix nitrogen, which will further increase their dominance in freshwater systems [7]. HABs create cascading ecological disruptions, affecting oxygen levels, food webs, habitats, and biodiversity. Climate change, nutrient pollution, and altered hydrodynamic conditions are making HABs more frequent and severe.
4.2. Economic
HABs cause many economic impacts, specifically affecting fisheries, tourism, and water quality management. The total economic cost of HABs in the US since 2000 has been estimated at USD 327 million [1]. Table 2 summarizes various historical economic costs for HABs, which are adapted from NOAA 2024. The occurrence with the largest cost was brown tides caused by the prevention of seagrasses from recovering, which resulted in potential loss to the local economy in the Indian River Lagoon, Florida, in 2011. The second most expensive occurrence was fisheries loss due to a toxic bloom of Pseudo-nitzschia, which impacted Dungeness crab landings in 2015 [84].
Table 2.
Cost (Million) of specific occurrences and events based on year and location.
A study conducted in 2024 by Alvarez et al. [85] estimated that Florida’s coastal counties experienced approximately USD 2.7 billion in tourism-related economic losses due to the 2018 red tide HAB. The study noted that unlike sudden disasters, such as flooding events, the economic impacts of HABs often unfold gradually and require comprehensive assessments to accurately quantify [85]. Figure 2 illustrates the aggregated economic effects of the red tide event, comparing observed sales data with simulated projections under HAB and non-HAB conditions, based on six coastal counties in Florida [85].
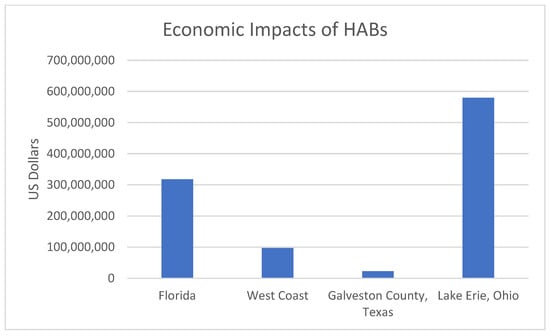
Figure 2.
The Economic Impacts of HABs Measured in Millions of U.S. dollars for Various Locations.
A pseudo-nitzschia bloom in 2015 in the West Coast resulted in USD 97.5 Million in lost Dungeness crab landing revenue [86]. In 2000, a red tide event in Galveston County, Texas resulted in an average of USD 22.9 Million in total economic impacts, which affected tourism and commercial oyster fisheries [84]. In Lake Erie, Ohio it cost around USD 580 million to protect drinking water from HABs [87]. Lastly, for Florida, the total economic impact caused by HABs was estimated at USD 318 million for the Airbnb Market [84].
Overall, HABs have far-reaching economic consequences that ripple across multiple industries, ecosystems, and communities. HABs can have significant economic impacts across various sectors, particularly in coastal and freshwater communities. The costs stem from both direct damages and indirect effects on industries, public health, and ecosystems. HABs cause economic damages to fisheries and aquaculture, tourism and recreation, increased burdens on public health systems and water treatment infrastructure, property values, and ecosystem services and restoration efforts [84].
Commercial fisheries tend to have losses due to the closures of shellfish harvesting areas due to toxins. Aquaculture industry impacts include fish farms suffering from catastrophic losses due to oxygen depletion or toxins. For tourism and recreation, HABs can lead to beach closures and negative media coverage, which results in tourism cancellations, lower visitation rates, and declines in water quality [88]. HABs can cause public health costs, in the form of medical expenses, lost wages, and increased burden on healthcare systems, to skyrocket due to the exposure to HAB toxins that can cause illness through seafood consumption, aerosolized toxins, and skin irritation from contact with water. Additionally, HABs can reduce property values by reducing the aesthetic and recreational appeal of waterfront properties. Lastly, ecosystem services and restoration costs will increase due to loss of biodiversity, a decline in water quality and habitat degradation. There is an expense when there are restoration efforts, such as nutrient reduction programs, and wetland restoration [85].
4.3. Public: Toxins Produced by HABs and Their Effects on Human Health
According to the EPA (2024), there are numerous toxins produced by HABs that can be harmful and cause negative effects on human health. Table 3 summarizes the health effects in humans of selected toxins produced by HABs. The most common health effects include gastroenteritis symptoms, cardiovascular symptoms, and neurological symptoms. Many of the humans exposed reported abdominal pain, diarrhea, vomiting, headaches, respiratory difficulty and more.
There is a variety of toxins produced by HABs, but one of the most detrimental is microcystins (MCs) which are a class of hepatotoxins produced by Microcystis. Although MCs are classified as hepatotoxins, the effects are not limited to just the liver. There are also effects on the renal system including necrosis, hyalinization of renal interstitium, hemorrhages, and infiltration of leukocytes [89]. Additionally, there are effects on the gastrointestinal system which include apoptosis in gastrointestinal cells and colorectal/rectal carcinomas [90,91,92]. In regards to the cardiovascular system, exposure to MCs can result in pathophysiological alterations like decline in heart rate/blood pressure, cardiac output, stroke volume, and oxygen consumption, which can lead to hypovolemic shock, hypertension, and cardiac arrest [91,92]. Lastly, MCs can also affect the neurological system, leading to neurodegenerative disorders or an enhancement in neurotoxicity [93].
Table 3.
Impact on Human Health.
Table 3.
Impact on Human Health.
| Toxin | Health Effects in Humans | References |
|---|---|---|
| Anatoxin-a group | Tingling, burning, numbness, respiratory paralysis, death, drowsiness, speech disturbances, excessive salivation, and muscle pain | [17,94,95] |
| Brevetoxin | Neurotoxic Shellfish Poisoning, asthma-like symptoms, coughing, mild ciguatera or paralytic shellfish poisoning, wheezing, shortness of breath, and chest tightness | [17,93,94] |
| Ciguatoxin | Diarrhea, nausea and vomiting, sensation in the mouth, aching teeth, abdominal cramps, muscular aches and pain, itching, dizziness, difficulty concentrating or hallucinations, sweating, numbness and tingling of the mouth, toes, or fingers, fatigue, reversal of temperature, joint pain, and bradycardia | [94,96,97,98] |
| Cylindrospermopsin | Fever, headache, induced DNA strand breaks, vomiting, bloody diarrhea, inhibits eukaryotic protein synthesis, hepatocellular damage, oxidative stress, lung hemorrhage, inflammation, apoptosis, hepatic necrosis, embryo toxicity, micronucleus formation, and tubular necrosis | [94,99,100] |
| Domoic Acid | Nausea, vomiting, abdominal cramps, respiratory secretions, cardiac arrhythmias, diarrhea, dizziness, profuse respiratory difficulty, confusion and disorientation, headache, seizures, hallucinations, possibly coma, short-term memory loss, and death | [94,101] |
| Microcystin-LR | Vomiting and nausea, abdominal pain, liver cancer, hepatomegaly, intrahepatic bleeding, liver necrosis, impairs kidney function, alters hormone levels, reduces sperm quality, induces embryotoxicity, synaptic damage, disrupts immune cell function, headache, sore throat, dry cough, intrahepatic bleeding, liver necrosis, diarrhea, blistering around the mouth, liver fibrosis, cirrhosis, impairs kidney function, pneumonia, and liver cancer | [94,102,103,104] |
| Okadaic Acid | Diarrhea, nausea and vomiting, inhibits protein phosphatases and disrupts cellular regulation, abdominal cramps, induces DNA damage and cell death, chills, headache, impairs immune system function, fever, affects neuronal cells, and carcinogenesis | [96,105,106] |
| Saxitoxin | Tingling, numbness, and burning of the mouth and lips, ataxia, muscle weakness, loss of coordination, irregular heartbeat, giddiness, drowsiness, difficulty breathing, respiratory paralysis, fever, rash, diarrhea, respiratory arrest, staggering, abdominal pain, neurological symptoms, headache, nausea, vomiting, and dizziness | [96,98,107] |
A majority of the cases of human health impacts are due to cyanotoxin. The first known major outbreak occurred in 1979 in Palm Island, Australia, as a result of the outbreak of HABs induced by hepatoenteritis. This event included a total of 140 children and 10 adults who were hospitalized for symptoms including malaise, anorexia, vomiting, headache, bloody diarrhea, dehydration, painful hepatomegaly, acute kidney disease, liver failure, and elevated serum enzyme levels [108,109]. The outbreak was traced to a contaminated potable water reservoir containing toxic cyanobacteria.
5. Monitoring and Detecting HABs
For the prediction and control of HABs, it is imperative to monitor HABs and their precursors in water bodies and the watershed. There are both new and traditional methods for monitoring HABs. Traditional methods include field monitoring and laboratory analysis, and online monitoring, and new methods include remote sensing monitoring and molecular biotechnology.
Field investigations continue to serve as the most traditional and direct method for monitoring HABs [110]. This approach offers several advantages, including the ability to assess visual, olfactory, physical-chemical, and biological indicators that may influence bloom development and dynamics [41]. Laboratory analysis complements field studies by measuring nutrient concentrations and analyzing algal community structures [111]. Common laboratory methods include microscopic species identification, cell counting, and pigment analysis [112], with light microscopy being the primary tool for detecting and quantifying HAB taxa [113]. However, laboratory-based monitoring has limitations, such as high operational costs, sensitivity to weather disruptions, and challenges in providing dynamic, real-time spatiotemporal data [114].
Remote sensing has become an essential technique for HAB surveillance, offering high efficiency and cost-effectiveness, especially in regions at high risk for blooms [115]. It allows for the sustained collection of high spatial and temporal resolution observations [116]. Despite its advantages, remote sensing faces limitations, including cloud cover interference, which restricts temporal and spatial data acquisition. Moreover, the complexity of optical properties in freshwater systems, variability in sensor performance, and differences in model algorithms present additional challenges in accurately retrieving water quality parameters [117].
Online monitoring systems represent another effective strategy, capable of automatically tracking phytoplankton abundance. These systems are valued for their operational simplicity and high stability. Since water quality parameters such as temperature, turbidity, pH, dissolved oxygen (DO), total phosphorus (TP), total nitrogen (TN), chlorophyll-a (chl-a), phycocyanin (a pigment correlated with cyanobacteria), and adenosine triphosphate (ATP) are indicators of HAB activity, continuous online monitoring offers considerable promise for early detection [19]. Looking forward, innovations such as bloom-specific online fluorometers incorporating fluorescence sensing, embedded micro-control units, and real-time data acquisition technologies could enable real-time dynamic monitoring of HABs [117]. Online monitoring provides an important advantage in that it can help identify benthic (solid-water interface) cyanobacteria blooms or actinobacteria blooms, which often do not have visible blooms like pelagic cyanobacteria (water-air interface) blooms [19]. Online monitoring systems, such as moored sensor buoys and automated water quality stations, provide continuous, real-time tracking of phytoplankton and water quality parameters. These systems are equipped with sensors for temperature, turbidity, pH, dissolved oxygen, nutrients, and algal pigments like chlorophyll-a and phycocyanin are vital for early detection of harmful algal blooms [19].
A further advantage of online monitoring systems is that they provide real-time data that can feed process-based (PB) models, which use mathematical relationships that describe the physics, chemistry, and biology of HAB formation to predict blooms [116]. Broadly, these can be classified as ecological or hydrodynamic. Ecological models like the Corps of Engineers Quality-Integrated Compartment Model (CE-QUAL-ICM) focus on processes such as nutrient cycling in zooplankton, phytoplankton, and cyanobacteria. Hydrodynamic models, such as the Corps of Engineers-Water Quality-Two, focus on physical parameters and processes such as fluid flow, sediment transport, and salinity and temperature gradients [116]. These two model types can also be coupled as hydrodynamic-ecological models, with the hydrodynamic component providing input for the ecological model [116]. Computationally intensive PB models with sufficient data and accounting for the correct processes can provide long-term prediction as well as a mechanistic understanding of HAB dynamics [116].
Molecular detection technologies have recently emerged as a promising alternative, particularly for distinguishing algal taxa that are difficult to identify using traditional microscopy [118]. Molecular methods allow for rapid and accurate identification, particularly when processing large numbers of samples. Despite these advantages, molecular detection is still in its early stages of development, with limited application in field-based monitoring due to a primary focus on system development rather than practical deployment [117].
Of the available detection methods, satellite remote sensing remains the most commonly used for large-scale HAB monitoring. Available satellites include Worldview-2, Worldview-3, Sentinel-2, and Landsat-8 Operational Land Imager [116]. When integrated with simple transport models, remote sensing enables the forecasting of HAB landfall and exposure events [119,120]. Remote sensing applications typically rely on identifying water masses containing HAB cells, often through measurements such as sea surface temperature.
In addition, molecular probes constructed using antibodies or DNA sequences specific to targeted HAB species have been developed to improve the speed and accuracy of species detection compared to microscopy alone. Analytical methods, particularly the combination of chromatography and mass spectrometry, are also increasingly utilized to detect and quantify HAB-associated toxins with high sensitivity [121].
A survey of 37 North American and Australian drinking water treatment plants classified the above strategies into three levels. The first tier, identifying microbial activity at the source, includes satellites, drones, cameras, and citizen scientists for observations of blooms as well as online measurement of water quality parameters. Observations in Tier 1 trigger Tier 2 methods, which are more expensive but are aimed at positively identifying cyanobacteria: measuring phycocyanin and microbial counting and speciation. Positive identification of cyanobacteria with Tier 2 measurements triggers active monitoring for cyanotoxins as well as genes that promote cyanotoxin production [19].
Several advanced technologies have recently emerged for monitoring HABs, including flow imaging microscopy (FIM), drone-based multispectral sensing, and machine learning (ML) models. FIM is an innovative method that merges the capabilities of both flow cytometry and microscopy to analyze and identify HAB-forming species. Instruments such as FlowCam enable real-time imaging, detection, and classification of potential cyanotoxin-producing organisms. Numerous case studies have demonstrated the successful application of FlowCam in HAB monitoring initiatives, particularly for reservoir management and water treatment systems [122].
Drone-based multispectral sensors, often referred to as unpiloted aerial systems (UAS), have also shown high efficiency, exceeding 90%, in detecting cyanobacterial blooms. Research conducted in New Hampshire’s water bodies confirmed that UAS platforms offer a safer and quicker alternative to traditional water sampling methods. Equipped with specialized sensors, these drones capture spectral images from above water surfaces, allowing researchers to estimate cyanobacteria concentrations based on the spectral signatures. Depending on the size of the waterbody, data collection can be completed within 10 to 15 min [123].
Hyperspectral imaging (HSI) is another emerging technology that expands upon the capabilities of multispectral imaging. HSI combines data not only from visible (red-green-blue) light data, but also wavelengths ranging to near-infrared, a wider range than multispectral [124]. Multispectral data is recorded by older satellites, like Landsat, but NASA’s Plankton, Aerosol, Cloud, Ocean Ecosystem (PACE) mission includes the Ocean Color Instrument (OCI), and other European missions have wavelength ranges of 400–2500 nm [124]. In addition to satellites, HSI can also be applied from manned aircraft, UAVs, or handheld devices, and remains a technology that continues to grow both in the scope of its application and its accuracy [124]. The increased details captured in the data require special data structures and processing techniques, but the main advantage is reliable identification of species with up to 90% accuracy by identifying signatures of pigments such as chlorophyll-a and phycocyanin [124]. The main limit of HSI is the amount of data generated and the computational power required to analyze it, but faster cellular communication and cloud computing are reducing these barriers [124]. Machine learning models are increasingly being utilized to predict and monitor HAB events due to their cost-effectiveness and modeling efficiency. ML algorithms, particularly for object detection, are emerging as valuable tools for developing predictive models that require less dependence on manual parameter selection. Various ML techniques, such as deep learning, artificial neural networks (ANNs), and ensemble models, are actively being applied in this context [125,126]. While ML approaches handle complex, nonlinear datasets effectively, they often demand substantial computational resources [127]. Typically, ML models are divided into regression models, which predict continuous variables, and classification models, which categorize observations into distinct groups. Regression models can include linear regression, generalized linear mixed models (GLMMs), logistic regression, support vector machine (SVM), and multiadaptive regression splines (MARS) [116]. Neural networks can also predict continuous variables based on correlations and include models such as artificial neural networks (ANNs), recurrent neural networks (RNNs), convolutional neural networks (CNNs), and general regression neural networks (GRNNs) [116]. An emerging and promising technique is generative adversarial networks (GANs) in which one neural network predicts an image of HABs distribution (e.g., as measured by chlorophyll-a distribution), and another neural network assesses how realistic the image appears [128]. Genetic Programming (GP), often applied on top of neural networks, generates predictions iteratively through mutation and selection of candidates towards optimized predictions [116]. Classification models include decision trees and probabilistic models such as Bayesian networks (BNs) [116]. Input variables frequently include hydrological, meteorological, and other water quality parameters [129]. These data-driven models have the advantage of being able to identify complex patterns and adapt to new data, but require adequate data, and predictions can be hard to interpret mechanistically [116].
6. Mitigation Strategies for HABs
6.1. Policies
During the 1990s, the detection of high densities of dinoflagellates and the associated HABs captured the attention of the U.S. Congress due to their links to extensive fish kills and negative impacts on human health [130,131,132,133,134,135]. In response, Congress passed the Harmful Algal Bloom and Hypoxia Research and Control Act (HABHRCA). This legislation authorized and directed funding to the National Oceanic and Atmospheric Administration (NOAA) to spearhead the development of a National Research Plan addressing coastal HABs. Implementation of this plan led to significant scientific advancements in understanding HABs across NOAA’s areas of focus, including oceans, estuaries, and the Great Lakes. Notably, it also contributed to the successful prediction of the 2008 red tide event in the Gulf of Maine [134].
Policy mechanisms for managing HABs generally fall into four categories: prevention, restoration, amelioration, and no action [88]. Among these, prevention is regarded as the most cost-effective and environmentally beneficial strategy. A detailed overview of these policy approaches, along with examples, is presented in Table 4.
Table 4.
Policy Mechanisms for the management of HABs.
6.2. Awareness/Knowledge
One of the most critical strategies for combating HABs is raising public awareness about the associated risks. Promoting concern and motivating behavioral changes necessary to mitigate HABs relies heavily on educating the public about the dangers they pose. Public awareness is particularly crucial for illness prevention efforts. However, there is limited available data assessing the general public awareness and concern regarding HABs [137].
A study conducted in 2024 by Jacobi et al. [137] utilized Porter Novelli’s Fall 2020 Consumer Styles survey to gather information on this topic [137]. The survey was distributed to 4548 individuals aged 18 years and older, achieving an 80% response rate with 3625 participants completing it. Approximately 59% of respondents indicated awareness that HABs can pose threats to human and animal health. Notably, awareness regarding impacts on human health was slightly higher. About 49% of participants recognized that HAB-contaminated waters are unsafe for swimming, while 48% acknowledged that exposure to HABs could cause illness in humans. In contrast, only 42% were aware that HABs could be fatal to fish or wildlife, 31% recognized the risk to pets, and just 22% noted potential threats to livestock.
Demographic factors influenced awareness levels. Higher awareness was associated with individuals reporting better health status, older age, white race, non-Hispanic ethnicity, higher levels of education, absence of children in the household, and higher income levels [137].
6.3. Practices
Several management practices are available to help reduce the occurrence and severity of HABs. These waterbody management strategies are generally categorized into two groups: physical controls and chemical controls [136]. Physical controls include techniques such as aeration, hydrologic manipulation, mechanical mixing (or circulation), reservoir drawdown and desiccation, surface skimming, and the use of ultrasound. On the other hand, chemical controls involve the application of algaecides, barley straw, and processes like coagulation and flocculation. Figure 3 provides a visual overview of these waterbody management approaches.
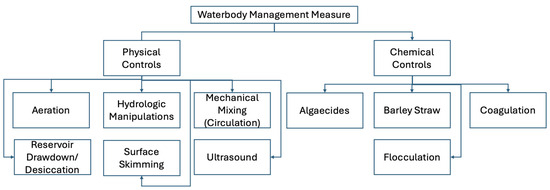
Figure 3.
Waterbody management measures for combating HABs.
Aerators operate by pumping air through a diffuser positioned near the bottom of the waterbody. This process creates plumes that rise to the surface and generate vertical circulation cells as they spread outward from the aeration point. The resulting water column mixing disrupts the vertical migration patterns of cyanobacteria and limits their access to essential nutrients. Aeration is most commonly and effectively applied in small ponds and reservoirs. Additionally, this method can create more favorable conditions for the growth of competing, non-harmful aquatic organisms [136]. This remains among the most popular methods for utilities to control HABs, but its effectiveness can be limited by inadequate or intermittent mixing [19].
Hydrologic manipulation involves adjusting the inflow and outflow of water within a system to break down thermal stratification, thereby inhibiting cyanobacterial proliferation. This technique is particularly well-suited for controlled environments such as reservoirs, dams, and water treatment facilities. However, it requires an adequate water supply and the ability to manage flow rates. A notable drawback of this approach is its potential expense, as well as the risk of unintended impacts on other aquatic organisms [136]. Operators have noted that reservoirs that have otherwise been successfully treated for blooms can have blooms recur due to the reintroduction of cyanobacteria via fill water [19].
Mechanical mixing, also referred to as circulation, typically involves the use of surface-mounted devices designed to disrupt cyanobacterial behavior by altering their vertical movement and limiting their access to nutrients. These systems either pump surface water downward or draw deeper water upward, helping to break thermal stratification and interrupt bloom development. However, mechanical mixing devices often have a limited effective range, meaning areas farther from the equipment may remain stratified. Additionally, mixing can sometimes create conditions that favor cyanobacterial regrowth if not carefully managed [136].
Reservoir drawdown and desiccation involve lowering water levels in controlled waterbodies to expose cyanobacterial accumulations above the surface. Following exposure, desiccation or physical removal of cyanobacterial layers attached to sediments or rocks is typically necessary before refilling the reservoir. This technique is most applicable in managed systems such as reservoirs, dams, and treatment facilities. While effective, it can significantly impact other aquatic organisms and is resource-intensive and costly [136].
Surface skimming targets cyanobacterial blooms that have formed dense surface scums, particularly during the later stages of a bloom. Techniques similar to oil-spill recovery methods are employed, often in combination with the application of coagulants or flocculants to assist removal. Surface skimming is beneficial once surface accumulations are clearly present; however, it is less useful during the early stages of a bloom when damage to the ecosystem may already be underway. Additionally, this method requires specialized equipment and preparation [136].
Ultrasound technology offers a non-chemical approach to controlling HABs. Devices emit ultrasonic waves at specific frequencies that disrupt the cellular structure of cyanobacteria by collapsing their internal gas vesicles, which are critical for buoyancy regulation. This method is particularly effective in ponds and small waterbodies, with a single device capable of treating areas up to 8 acres. Advantages of ultrasound include its chemical-free operation and relatively low cost. However, drawbacks include potential impacts on green algae and the method’s reliance on waterbody shape and the specific cyanobacteria species present (US EPA, 2018). Units have effective radii of about 300 m, and a survey of 37 utilities found that those which applied this technique often had undersized units. Some utilities have reported sonication to be ineffective, with blooms persisting during treatment [19].
Algaecides involve the direct application of chemical agents to eliminate cyanobacteria and terminate blooms. Commonly used algaecides include copper-based compounds, potassium permanganate, chlorine, and lime. This method has a long history of application, with well-documented properties and effects. Algaecides act quickly and are considered a reliable option. However, risks include the lysis of cyanobacterial cells, which can release toxins into the water. As a result, algaecide application is typically recommended during the early stages of a bloom. Additionally, some compounds can be toxic to non-target species, including zooplankton, invertebrates, and fish [136]. Hydrogen peroxide is more specific to cyanobacteria than copper-based compounds and has been shown to be effective, while cyanobacteria can become resistant to copper-based compounds after decades of application [19].
The barley straw method involves placing straw bales around the perimeter of a waterbody. When exposed to sunlight and oxygen, decomposing barley straw produces natural chemicals that inhibit algal reproduction. Research has confirmed that decomposed barley straw can effectively prevent cyanobacterial growth. However, this method does not eliminate existing algae, and it requires a lead time of 2–8 weeks for the straw to begin producing the active inhibitory compounds. Moreover, the decomposition process can lower dissolved oxygen levels, posing a risk of fish mortality [136].
In the coagulation method, chemical coagulants are used to promote the aggregation and sedimentation of cyanobacterial cells, causing them to sink to the anoxic bottom layers of the water column. Deprived of sunlight, oxygen, and other essential resources, the cyanobacteria are unable to survive and eventually die. One of the advantages of this approach is that it allows for the removal of cyanobacterial cells without causing immediate cellular damage, and it can be effectively applied in various water treatment facilities. However, a major limitation is its dependency on water depth, as coagulation is less effective in very shallow systems. Additionally, over time, stressed coagulated cells may lyse, leading to the release of intracellular toxins into the surrounding water [136]. A study of coagulation in Chlorella vulgaris blooms with aluminum chloride, ferric chloride, poly-aluminum chloride (PACl), chitosan, and polydimethyldiallylammonium chloride (PDADMAC) showed that the different coagulants worked by different mechanisms and PDADMAC was recommended as particularly effective [138].
Flocculation is often synonymous with coagulation, but the US EPA separates the mechanism as using the same physical-chemical process to aggregate and settle nutrients to the anoxic bottom layers. By removing key nutrients from the photic zone, this method limits the resources available for cyanobacterial growth, thereby reducing the likelihood of bloom formation. Flocculation is particularly effective in larger lakes and ponds. However, like coagulation, it faces depth limitations, which can reduce its effectiveness in shallower or highly dynamic water bodies [136]. The nutrient-limited conditions created by this process can create >90-day recovery times for blooms, especially when paired with aeration, longer than the <30-day times reported for algaecides [19].
While numerous reviews have explored the causes, ecological effects, and traditional monitoring methods of HABs, several critical research gaps remain unaddressed in the current body of literature. Previous reviews have primarily focused on nutrient enrichment, meteorological factors, and classical mitigation techniques [7,70], yet they often overlook the integration of advanced technologies such as hyperspectral imaging, drone-based multispectral sensing, machine learning (ML), and molecular biotechnology for real-time HAB detection and prediction [19,117,124]. Additionally, many studies fail to incorporate climate-induced variables like ocean acidification, altered salinity, and intensified upwelling into predictive models, which limits their robustness under evolving environmental conditions [29,96].
Notably, fully coupled hydrodynamic–ecological models, which combine physical and biological processes to simulate bloom dynamics at multiple scales, remain sparsely implemented despite their capacity to improve forecasting accuracy [86,116]. Furthermore, while molecular detection methods have demonstrated strong potential in laboratory applications, there is limited research evaluating their deployment in field conditions or integration into tiered early warning systems [19,20].
Another underexplored area is the interaction between microplastic pollution and HABs. Emerging evidence suggests that microplastics may enhance algal growth and toxin production by acting as vectors for nutrient and microbial exchange, especially when transported via stormwater runoff [10,81], yet this interaction remains poorly characterized in current models [31,83]. Economic assessments of HABs also tend to be retrospective and event-specific, lacking standardized, forward-looking models that account for long-term, multisector financial risks [1,85].
Geographically, existing literature remains concentrated in North America and Europe, with limited research focused on Asia, Africa, and South America regions that are increasingly experiencing HAB events due to rapid environmental change and inadequate monitoring infrastructure [7]. This review addresses these shortcomings by synthesizing recent literature on emerging detection tools, climate stressors, and underrepresented geographies, while emphasizing overlooked themes such as field-deployable molecular diagnostics and microplastic interactions. In doing so, it offers a more integrative, technologically current, and globally relevant framework for advancing HAB prediction, detection, and management.
7. Conclusions
HABs represent a significant and growing environmental challenge with wide-ranging ecological, economic, and human health consequences. As anthropogenic activities continue to contribute to nutrient enrichment in aquatic ecosystems, the frequency and intensity of HABs are expected to rise. These blooms disrupt aquatic and freshwater ecosystems by depleting oxygen levels, releasing harmful toxins, and leading to mass mortality of aquatic organisms. Additionally, they impose economic burdens on fisheries, tourism, and water treatment facilities, while also posing serious health risks to humans and animals through contaminated water and seafood.
Addressing HABs requires a multifaceted approach that integrates scientific research, policymaking, and community involvement. Continuous monitoring and early detection systems can help mitigate their impact, while regulatory policies aimed at reducing nutrient runoff from agricultural, industrial, and urban sources are essential for long-term prevention. Advancements in biotechnology and artificial intelligence also offer promising solutions for predicting and managing HAB outbreaks with greater accuracy.
Public awareness and education play a crucial role in combating HABs by encouraging sustainable agricultural practices, responsible waste disposal, and active participation in conservation efforts. Collaborative efforts among governments, environmental organizations, researchers, and local communities are imperative to developing effective mitigation strategies and safeguarding aquatic ecosystems.
While progress has been made in understanding the mechanisms driving HAB formation, further interdisciplinary research is necessary to develop innovative solutions that balance environmental protection with economic and social needs. By prioritizing sustainability and proactive management, we can work towards minimizing the adverse effects of HABs and ensuring the long-term health of our water bodies.
Author Contributions
Conceptualization, C.M.B., M.P.J. and A.D.B.; Writing—original draft, C.M.B.; Writing—review and editing, M.P.J., A.D.B., W.H.P. and F.A.; Validation, M.P.J.; Supervision, A.D.B.; All authors have read and agreed to the published version of the manuscript.
Funding
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Data Availability Statement
Dataset available upon request from the authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- NOAA Harmful Algae Blooms (HABs). National Environmental Satellite, Data, and Information Service. Available online: https://www.nesdis.noaa.gov/harmful-algae-blooms-habs (accessed on 24 June 2025).
- Chattopadhyay, J.; Sarkar, R.R.; Pal, S. Mathematical Modelling of Harmful Algal Blooms Supported by Experimental Findings. Ecol. Complex. 2004, 1, 225–235. [Google Scholar] [CrossRef]
- Capuzzo, E.; Stephens, D.; Silva, T.; Barry, J.; Forster, R.M. Decrease in Water Clarity of the Southern and Central North Sea during the 20th Century. Glob. Change Biol. 2015, 21, 2206–2214. [Google Scholar] [CrossRef]
- Goes, J.I.; Gomes, H.D.R.; Al-Hashimi, K.; Buranapratheprat, A. Ecological Drivers of Green Noctiluca Blooms in Two Monsoonal-Driven Ecosystems. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Springer International Publishing: Cham, Germany, 2018; pp. 327–336. ISBN 978-3-319-70069-4. [Google Scholar]
- Moy, N.J.; Dodson, J.; Tassone, S.J.; Bukaveckas, P.A.; Bulluck, L.P. Biotransport of Algal Toxins to Riparian Food Webs. Environ. Sci. Technol. 2016, 50, 10007–10014. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Ruan, R.; Addy, M.; Mack, S.; Chen, P.; Zhou, Y. Life Cycle Assessment and Nutrient Analysis of Various Processing Pathways in Algal Biofuel Production. Bioresour. Technol. 2017, 230, 33–42. [Google Scholar] [CrossRef]
- Anderson, D.M.; Fensin, E.; Gobler, C.J.; Hoeglund, A.E.; Hubbard, K.A.; Kulis, D.M.; Landsberg, J.H.; Lefebvre, K.A.; Provoost, P.; Richlen, M.L.; et al. Marine Harmful Algal Blooms (HABs) in the United States: History, Current Status and Future Trends. Harmful Algae 2021, 102, 101975. [Google Scholar] [CrossRef]
- Leibach, J. Ten Fun Facts About Algae|Audubon. Available online: https://www.audubon.org/magazine/ten-fun-facts-about-algae (accessed on 12 May 2025).
- Lewin, R.; Andersen, R. Algae—Photosynthesis, Diversity, Ecology. Available online: https://www.britannica.com/science/algae/Physical-and-ecological-features-of-algae (accessed on 10 February 2025).
- Parameswarappa Jayalakshmamma, M.; Na Nagara, V.; Borgaonkar, A.; Sarkar, D.; Sadik, O.; Boufadel, M. Characterizing Microplastics in Urban Runoff: A Multi-Land Use Assessment with a Focus on 1–125 Μm Size Particles. Sci. Total Environ. 2023, 904, 166685. [Google Scholar] [CrossRef] [PubMed]
- Aure, J.; Rey, F. Oceanographic Conditions in the Sandsfjord System, Western Norway, after a Bloom of the Toxic Prymnesiophyte Prymnesium Parvum Carter in August 1990. Sarsia 1992, 76, 247–254. [Google Scholar] [CrossRef]
- Rey, F. Oppblomstringen av Chrysochromulina leadbeateri i Vestfjorden, mai-juni 1991: Rapport fra et faglig arbeidsseminar. 122 S. 1991. Available online: https://core.ac.uk/download/pdf/225955132.pdf (accessed on 24 May 2025).
- Landsberg, J.H. The Effects of Harmful Algal Blooms on Aquatic Organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Shumway, S. A Review of the Effects of Algal Blooms on Shellfish and Aquaculture. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1749-7345.1990.tb00529.x (accessed on 5 February 2025).
- CDC Types of Harmful Algal Blooms. Available online: https://www.cdc.gov/harmful-algal-blooms/about/types-of-harmful-algal-blooms.html (accessed on 5 December 2024).
- Yannarell, A.C.; Kent, A.D. Bacteria, Distribution and Community Structure. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Oxford, UK, 2009; pp. 201–210. ISBN 978-0-12-370626-3. [Google Scholar]
- Singh, V.K.; Singh, S.K.; Singh, P.K.; Verma, H.; Pandey, K.D.; Singh, P.K.; Kumar, A. Chapter 10—Impact of Pesticides Applications on the Growth and Function of Cyanobacteria. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 151–161. ISBN 978-0-12-819311-2. [Google Scholar]
- CDC Harmful Algal Blooms and Your Health. Available online: https://www.cdc.gov/harmful-algal-blooms/about/index.html (accessed on 5 December 2024).
- Kibuye, F.A.; Almuhtaram, H.; Zamyadi, A.; Gaget, V.; Owen, C.; Hofmann, R.; Wert, E.C. Utility Practices and Perspectives on Monitoring and Source Control of Cyanobacterial Blooms. AWWA Water Sci. 2021, 3, e1264. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Zhang, H.; Zhang, Y.; Zhang, S.-F. Marine Dinoflagellate Proteomics: Current Status and Future Perspectives. J. Proteomics 2014, 105, 121–132. [Google Scholar] [CrossRef]
- Houck, M.M.; Crispino, F.; McAdam, T. Chapter 6.4—Other Types of Evidence. In The Science of Crime Scenes; Houck, M.M., Crispino, F., McAdam, T., Eds.; Academic Press: San Diego, CA, USA, 2012; ISBN 978-0-12-386464-2. [Google Scholar]
- Assmy, P.; Smetacek, V.; Montresor, M.; Ferrante, M.I. Algal Blooms☆. In Encyclopedia of Microbiology, 4th ed.; Schmidt, T.M., Ed.; Academic Press: Oxford, UK, 2019; pp. 61–76. ISBN 978-0-12-811737-8. [Google Scholar]
- Research Gaps Identified to Improve Future Harmful Algal Bloom Forecasts. Available online: https://coastalscience.noaa.gov/news/research-gaps-identified-improve-future-harmful-algal-bloom-forecasts/ (accessed on 5 February 2025).
- Muller-Feuga, A.; Le Guedes, R.; Herve, A.; Durand, P. Comparison of Artificial Light Photobioreactors and Other Production Systems Using Porphyridium Cruentum. J. Appl. Phycol. 1998, 10, 83–90. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of Temperature and Light on the Growth of Algae Species: A Review. Renew. Sustain. Energy Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Rao, Q.; Su, H.; Ruan, L.; Deng, X.; Wang, L.; Rao, X.; Liu, J.; Xia, W.; Xu, P.; Shen, H.; et al. Stoichiometric and Physiological Mechanisms That Link Hub Traits of Submerged Macrophytes with Ecosystem Structure and Functioning. Water Res. 2021, 202, 117392. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhang, J.; Huo, Y.; Zhou, L.; Wu, Q.; Chen, L.; Yu, K.; He, P. Adaptability of Free-Floating Green Tide Algae in the Yellow Sea to Variable Temperature and Light Intensity. Mar. Pollut. Bull. 2015, 101, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, Y.; Song, X.; Jiang, M.; Zhao, X.; Cao, X. The Inhibitory Effects of Simulated Light Sources on the Activity of Algae Cannot Be Ignored in Photocatalytic Inhibition. Chemosphere 2022, 309, 136611. [Google Scholar] [CrossRef]
- Ho, J.C.; Michalak, A.M. Exploring Temperature and Precipitation Impacts on Harmful Algal Blooms across Continental U.S. Lakes. Limnol. Oceanogr. 2020, 65, 992–1009. [Google Scholar] [CrossRef]
- Anneville, O.; Gammeter, S.; Straile, D. Phosphorus Decrease and Climate Variability: Mediators of Synchrony in Phytoplankton Changes among European Peri-Alpine Lakes. Freshw. Biol. 2005, 50, 1731–1746. [Google Scholar] [CrossRef]
- Peng, X.; Chen, M.; Chen, S.; Dasgupta, S.; Xu, H.; Ta, K.; Du, M.; Li, J.; Guo, Z.; Bai, S. Microplastics Contaminate the Deepest Part of the World’s Ocean. Geochem. Perspect. Lett. 2018, 9, 1–5. [Google Scholar] [CrossRef]
- Huang, J.; Xu, Q.; Xi, B.; Wang, X.; Li, W.; Gao, G.; Huo, S.; Xia, X.; Jiang, T.; Ji, D.; et al. Impacts of Hydrodynamic Disturbance on Sediment Resuspension, Phosphorus and Phosphatase Release, and Cyanobacterial Growth in Lake Tai. Environ. Earth Sci. 2015, 74, 3945–3954. [Google Scholar] [CrossRef]
- Li, F.; Zhang, H.; Zhu, Y.; Xiao, Y.; Chen, L. Effect of Flow Velocity on Phytoplankton Biomass and Composition in a Freshwater Lake. Sci. Total Environ. 2013, 447, 64–71. [Google Scholar] [CrossRef]
- Matson, P.G.; Boyer, G.L.; Bridgeman, T.B.; Bullerjahn, G.S.; Kane, D.D.; McKay, R.M.L.; McKindles, K.M.; Raymond, H.A.; Snyder, B.K.; Stumpf, R.P.; et al. Physical Drivers Facilitating a Toxigenic Cyanobacterial Bloom in a Major Great Lakes Tributary. Limnol. Oceanogr. 2020, 65, 2866–2882. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Zhang, H.; Lei, P.; Tang, W.; Yin, W.; Li, J.; Zhong, H.; Li, K. Algal Blooms in the Middle and Lower Han River: Characteristics, Early Warning and Prevention. Sci. Total Environ. 2020, 706, 135293. [CrossRef]
- Zhu, C.; Chi, Z.; Bi, C.; Zhao, Y.; Cai, H. Hydrodynamic Performance of Floating Photobioreactors Driven by Wave Energy. Biotechnol. Biofuels 2019, 12, 54. [Google Scholar] [CrossRef] [PubMed]
- San, L.; Long, T.; Liu, C.C.K. Algal Bioproductivity in Turbulentwater: An Experimental Study. Water 2017, 9, 304. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, T.; Liang, D.; Chen, J.M. Experimental Study of Water and Dissolved Pollutant Runoffs on Impervious Surfaces. J. Hydrodyn. 2016, 28, 162–165. [Google Scholar] [CrossRef]
- Yang, G.; Tang, X.; Wilhelm, S.W.; Pan, W.; Rui, Z.; Xu, L.; Zhong, C.; Hu, X. Intermittent Disturbance Benefits Colony Size, Biomass and Dominance of Microcystis in Lake Taihu under Field Simulation Condition. Harmful Algae 2020, 99, 101909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xiao, Y.; Li, Z.; Wang, S.; Guo, J.; Lu, L. Turbulence Exerts Nutrients Uptake and Assimilation of Bloom-Forming Dolichospermum through Modulating Morphological Traits: Field and Chemostat Culture Studies. Sci. Total Environ. 2019, 671, 329–338. [Google Scholar] [CrossRef]
- Zhao, G.; Gao, X.; Zhang, C.; Sang, G. The Effects of Turbulence on Phytoplankton and Implications for Energy Transfer with an Integrated Water Quality-Ecosystem Model in a Shallow Lake. J. Environ. Manag. 2020, 256, 109954. [Google Scholar] [CrossRef]
- Kang, L.; He, Y.; Dai, L.; He, Q.; Ai, H.; Yang, G.; Liu, M.; Jiang, W.; Li, H. Interactions between Suspended Particulate Matter and Algal Cells Contributed to the Reconstruction of Phytoplankton Communities in Turbulent Waters. Water Res. 2019, 149, 251–262. [Google Scholar] [CrossRef]
- Liu, P.; Qian, L.; Wang, H.; Zhan, X.; Lu, K.; Gu, C.; Gao, S. New Insights into the Aging Behavior of Microplastics Accelerated by Advanced Oxidation Processes. Environ. Sci. Technol. 2019, 53, 3579–3588. [Google Scholar] [CrossRef]
- Wu, X.; Noss, C.; Liu, L.; Lorke, A. Effects of Small-Scale Turbulence at the Air-Water Interface on Microcystis Surface Scum Formation. Water Res. 2019, 167, 115091. [Google Scholar] [CrossRef]
- Marrasé, C.; Costello, J.H.; Granata, T.; Strickler, J.R. Grazing in a Turbulent Environment: Energy Dissipation, Encounter Rates, and Efficacy of Feeding Currents in Centropages Hamatus. Proc. Natl. Acad. Sci. USA 1990, 87, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Shen, L.; Zhang, L.; Li, J.; Chen, M. Study of a Hydrodynamic Threshold System for Controlling Dinoflagellate Blooms in Reservoirs. Environ. Pollut. 2021, 278, 116822. [Google Scholar] [CrossRef] [PubMed]
- Burford, M.A.; Johnson, S.A.; Cook, A.J.; Packer, T.V.; Taylor, B.M.; Townsley, E.R. Correlations between Watershed and Reservoir Characteristics, and Algal Blooms in Subtropical Reservoirs. Water Res. 2007, 41, 4105–4114. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Mu, Z.; Wang, H.; Zhao, F.; Li, Y.; Lin, L. Water Residence Time in a Typical Tributary Bay of the Three Gorges Reservoir. Water 2019, 11, 1585. [Google Scholar] [CrossRef]
- Chuo, M.; Ma, J.; Liu, D.; Yang, Z. Effects of the Impounding Process during the Flood Season on Algal Blooms in Xiangxi Bay in the Three Gorges Reservoir, China. Ecol. Model. 2019, 392, 236–249. [Google Scholar] [CrossRef]
- Gao, X.; Kong, B.; Vigil, R.D. Multiphysics Simulation of Algal Growth in an Airlift Photobioreactor: Effects of Fluid Mixing and Shear Stress. Bioresour. Technol. 2018, 251, 75–83. [Google Scholar] [CrossRef]
- Soares, M.C.S.; Marinho, M.M.; Huszar, V.L.M.; Branco, C.W.C.; Azevedo, S.M.F.O. The Effects of Water Retention Time and Watershed Features on the Limnology of Two Tropical Reservoirs in Brazil. Lakes Reserv. Sci. Policy Manag. Sustain. Use 2008, 13, 257–269. [Google Scholar] [CrossRef]
- Song, Y. Hydrodynamic Impacts on Algal Blooms in Reservoirs and Bloom Mitigation Using Reservoir Operation Strategies: A Review. J. Hydrol. 2023, 620, 129375. [Google Scholar] [CrossRef]
- Amorim, C.A.; Moura, A.D.N. Ecological Impacts of Freshwater Algal Blooms on Water Quality, Plankton Biodiversity, Structure, and Ecosystem Functioning. Sci. Total Environ. 2021, 758, 1585. [Google Scholar] [CrossRef]
- Raine, R.; Berdalet, E.; Yamazaki, H.; Jenkinson, I.; Reguera, B. Key Questions and Recent Research Advances on Harmful Algal Blooms in Stratified Systems. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Springer International Publishing: Cham, UK, 2018; pp. 165–186. ISBN 978-3-319-70069-4. [Google Scholar]
- Azadi, F.; Ashofteh, P.-S.; Chu, X. Evaluation of the Effects of Climate Change on Thermal Stratification of Reservoirs. Sustain. Cities Soc. 2021, 66, 102531. [Google Scholar] [CrossRef]
- Ishikawa, M.; Gurski, L.; Bleninger, T.; Rohr, H.; Wolf, N.; Lorke, A. Hydrodynamic Drivers of Nutrient and Phytoplankton Dynamics in a Subtropical Reservoir. Water 2022, 14, 1544. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, P.; Liu, D.; Ma, J.; Ji, D.; Cui, Y. Hydrodynamic Mechanisms Underlying Periodic Algal Blooms in the Tributary Bay of a Subtropical Reservoir. Ecol. Eng. 2018, 120, 6–13. [Google Scholar] [CrossRef]
- Hadiyanto, H.; Elmore, S.; Van Gerven, T.; Stankiewicz, A. Hydrodynamic Evaluations in High Rate Algae Pond (HRAP) Design. Chem. Eng. J. 2013, 217, 231–239. [Google Scholar] [CrossRef]
- Sarat Chandra, T.; Aditi, S.; Maneesh Kumar, M.; Mukherji, S.; Modak, J.; Chauhan, V.S.; Sarada, R.; Mudliar, S.N. Growth and Biochemical Characteristics of an Indigenous Freshwater Microalga, Scenedesmus Obtusus, Cultivated in an Airlift Photobioreactor: Effect of Reactor Hydrodynamics, Light Intensity, and Photoperiod. Bioprocess Biosyst. Eng. 2017, 40, 1057–1068. [Google Scholar] [CrossRef]
- US EPA. Climate Change and Freshwater Harmful Algal Blooms. Available online: https://www.epa.gov/habs/climate-change-and-freshwater-harmful-algal-blooms (accessed on 12 May 2025).
- Gobler, C.J. Climate Change and Harmful Algal Blooms: Insights and Perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Likens, G.E.; Pace, M.L.; Reimer, J.E.; Maas, C.M.; Galella, J.G.; Utz, R.M.; Duan, S.; Kryger, J.R.; Yaculak, A.M.; et al. Freshwater Salinization Syndrome: From Emerging Global Problem to Managing Risks. Biogeochemistry 2021, 154, 255–292. [Google Scholar] [CrossRef]
- Qiu, Y.; Ma, Z.; Liu, X.; Zheng, R.; Xiao, Y.; Wang, M. The Detrimental Effect of High Salinity on the Growth and Microcystins Contamination of Microcystis Aeruginosa. Water 2022, 14, 2871. [Google Scholar] [CrossRef]
- Pan, Y.; Amenorfenyo, D.K.; Dong, M.; Zhang, N.; Huang, X.; Li, C.; Li, F. Effects of Salinity on the Growth, Physiological and Biochemical Components of Microalga Euchlorocystis Marina. Front. Mar. Sci. 2024, 11, 1402071. [Google Scholar] [CrossRef]
- Yao, C.-H.; Ai, J.-N.; Cao, X.-P.; Xue, S. Salinity Manipulation as an Effective Method for Enhanced Starch Production in the Marine Microalga Tetraselmis subcordiformis. Bioresour. Technol. 2013, 146, 663–671. [Google Scholar] [CrossRef]
- Lan, J.; Liu, P.; Hu, X.; Zhu, S. Harmful Algal Blooms in Eutrophic Marine Environments: Causes, Monitoring, and Treatment. Water 2024, 16, 2525. [Google Scholar] [CrossRef]
- Le, K.T.N.; Maldonado, J.F.G.; Nguyen, T.-L.; Goitom, E.; Trigui, H.; Ndiaye, N.A.; Terrat, Y.; Shapiro, B.J.; Husk, B.; Zamyadi, A.; et al. The Short-Term Effect of Nitrogen on Freshwater Cyanobacteria and Cyanotoxins. Front. Water 2024, 6, 1432183. [Google Scholar] [CrossRef]
- Kudela, R.M.; Seeyave, S.; Cochlan, W.P. The Role of Nutrients in Regulation and Promotion of Harmful Algal Blooms in Upwelling Systems. Prog. Oceanogr. 2010, 85, 122–135. [Google Scholar] [CrossRef]
- Oduor, N.A.; Munga, C.N.; Ong’anda, H.O.; Botwe, P.K.; Moosdorf, N. Nutrients and Harmful Algal Blooms in Kenya’s Coastal and Marine Waters: A Review. Ocean Coast. Manag. 2023, 233, 106454. [Google Scholar] [CrossRef]
- Wang, J.; Dong, J.; Tang, M.; Yao, J.; Li, X.; Kong, D.; Zhao, K. Identification and Detection of Microplastic Particles in Marine Environment by Using Improved Faster R–CNN Model. J. Environ. Manag. 2023, 345, 118802. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Onodera, S.-I.; Saito, M.; Shimizu, Y. Long-Term Variations in Water Balance by Increase in Percent Imperviousness of Urban Regions. J. Hydrol. 2021, 602, 126767. [Google Scholar] [CrossRef]
- Wells, M.L.; Karlson, B. Harmful Algal Blooms in a Changing Ocean. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 77–90. ISBN 978-3-319-70069-4. [Google Scholar]
- Ragueneau, O.; Conley, D.J.; Leynaert, A.; Longphuirt, S.N.; Slomp, C.P. Role of Diatoms in Silica Cycling and Coastal Marine Food Webs. SCOPE 2006, 66, 163–195. [Google Scholar]
- Shetye, S.; Pratihary, A.; Shenoy, D.; Kurian, S.; Gauns, M.; Uskaikar, H.; Naik, B.; Nandakumar, K.; Borker, S. Rice Husk as a Potential Source of Silicate to Oceanic Phytoplankton. Sci. Total Environ. 2023, 879, 162941. [Google Scholar] [CrossRef]
- Davidson, K.; Gowen, R.J.; Harrison, P.J.; Fleming, L.E.; Hoagland, P.; Moschonas, G. Anthropogenic Nutrients and Harmful Algae in Coastal Waters. J. Environ. Manag. 2014, 146, 206–216. [Google Scholar] [CrossRef]
- Heffer, P.; Prud’homme, M. Global Nitrogen Fertiliser Demand and Supply: Trend, Current Level and Outlook. In Proceedings of the International Nitrogen Initiative Conference, Melbourne, Australia, 4 December 2016. [Google Scholar]
- Paerl, H.W. Coastal Eutrophication and Harmful Algal Blooms: Importance of Atmospheric Deposition and Groundwater as “New” Nitrogen and Other Nutrient Sources. Limnol. Oceanogr. 2003, 42, 1154–1165. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G.; Kudela, R. Mitigating the Expansion of Harmful Algal Blooms Across the Freshwater-to-Marine Continuum. Environ. Sci. Technol. 2018, 52, 5519–5529. [Google Scholar] [CrossRef] [PubMed]
- Wurtsbaugh, W.A.; Paerl, H.W.; Dodds, W.K. Nutrients, Eutrophication and Harmful Algal Blooms along the Freshwater to Marine Continuum. WIREs Water 2019, 6, e1373. [Google Scholar] [CrossRef]
- FAO Inorganic Fertilizers 1961–2019. Available online: https://www.fao.org/statistics/highlights-archive/highlights-detail/New-FAOSTAT-data-release-Inorganic-fertilizers-(1961-2019)/en (accessed on 5 March 2025).
- Parameswarappa Jayalakshmamma, M.; Na Nagara, V.; Borgaonkar, A.; Sarkar, D.; Obropta, C.; Boufadel, M. Temporal and Spatial Distribution of Microplastics in Green Infrastructures: Rain Gardens. Chemosphere 2024, 362, 142543. [Google Scholar] [CrossRef] [PubMed]
- Sprouse, L.; Kryston, A.; Lebu, S.; Muoghalu, C.; Woods, C.; Manga, M. Septic Systems in North Carolina: A Neglected Half of the State? PLoS Water 2024, 3, e0000304. Available online: https://journals.plos.org/water/article?id=10.1371/journal.pwat.0000304 (accessed on 12 May 2025). [CrossRef]
- Brewton, R.A.; Kreiger, L.B.; Tyre, K.N.; Baladi, D.; Wilking, L.E.; Herren, L.W.; Lapointe, B.E. Septic System–Groundwater–Surface Water Couplings in Waterfront Communities Contribute to Harmful Algal Blooms in Southwest Florida. Sci. Total Environ. 2022, 837, 155319. [Google Scholar] [CrossRef]
- Hitting Us Where It Hurts: The Untold Story of Harmful Algal Blooms. Available online: https://www.fisheries.noaa.gov/west-coast/science-data/hitting-us-where-it-hurts-untold-story-harmful-algal-blooms (accessed on 5 March 2025).
- Alvarez, S.; Brown, C.E.; Garcia Diaz, M.; O’Leary, H.; Solís, D. Non-Linear Impacts of Harmful Algae Blooms on the Coastal Tourism Economy. J. Environ. Manag. 2024, 351, 119811. [Google Scholar] [CrossRef]
- NCCOS Harmful Algal Blooms: NOAA State of the Science Fact Sheet. Available online: https://coastalscience.noaa.gov/news/hab-noaa-fact-sheet/ (accessed on 3 June 2025).
- Neese, A.W. Climate Change Could Cost Ohio Billions. Available online: https://www.axios.com/local/columbus/2022/07/26/climate-change-could-cost-ohio-billions (accessed on 3 June 2025).
- Carias, J.; Vásquez-Lavín, F.; Barrientos, M.; Ponce Oliva, R.D.; Gelcich, S. Economic Valuation of Harmful Algal Blooms (HAB): Methodological Challenges, Policy Implications, and an Empirical Application. J. Environ. Manag. 2024, 365, 121566. [Google Scholar] [CrossRef]
- Milutinović, A.; Sedmak, B.; Horvat-Znidarsic, I.; Suput, D. Renal Injuries Induced by Chronic Intoxication with Microcystins. Cell. Mol. Biol. Lett. 2002, 7, 139–141. [Google Scholar]
- Botha, N.; van de Venter, M.; Downing, T.G.; Shephard, E.G.; Gehringer, M.M. The Effect of Intraperitoneally Administered Microcystin-LR on the Gastrointestinal Tract of Balb/c Mice. Toxicon 2004, 43, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Taylor & Francis: Oxfordshire, UK, 2021; ISBN 978-0-367-53332-8. [Google Scholar]
- Lun, Z.; Hai, Y.; Kun, C. Relationship Between Microcystin in Drinking Water and Colorectal Cancer. Biomed. Environ. Sci. 2002, 15, 166–171. [Google Scholar]
- Lad, A.; Breidenbach, J.D.; Su, R.C.; Murray, J.; Kuang, R.; Mascarenhas, A.; Najjar, J.; Patel, S.; Hegde, P.; Youssef, M.; et al. As We Drink and Breathe: Adverse Health Effects of Microcystins and Other Harmful Algal Bloom Toxins in the Liver, Gut, Lungs and Beyond. Life 2022, 12, 418. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.D.; Colvara, W.A.; Rantin, F.T.; Kalinin, A.L. Microcystin-LR: How It Affects the Cardio-Respiratory Responses to Hypoxia in Nile Tilapia, Oreochromis niloticus. Chemosphere 2011, 84, 154–159. [Google Scholar] [CrossRef]
- Qiu, T.; Xie, P.; Liu, Y.; Li, G.; Xiong, Q.; Hao, L.; Li, H. The Profound Effects of Microcystin on Cardiac Antioxidant Enzymes, Mitochondrial Function and Cardiac Toxicity in Rat. Toxicology 2009, 257, 86–94. [Google Scholar] [CrossRef]
- US EPA. What Are the Effects of HABs. Available online: https://www.epa.gov/habs/what-are-effects-habs (accessed on 5 March 2025).
- Washington State Department of Health Provisional Health Advisory Level for Anatoxin-a and Saxitoxin in Drinking Water. 2024. Available online: https://doh.wa.gov/sites/default/files/2025-01/334-552.pdf (accessed on 5 March 2025).
- Florida Health Charts Saxitoxin Poisoning (Paralytic Shellfish Poisoning). Available online: https://www.flhealthcharts.gov/ChartsDashboards/rdPage.aspx?rdReport=NonVitalIndNoGrpCounts.DataViewer&cid=8615 (accessed on 3 June 2025).
- Kimura, A. Ciguatera Fish Poisoning Fact Sheet. Available online: https://www.cdph.ca.gov/Programs/CID/DCDC/CDPH%20Document%20Library/CiguateraFishPoisoningFactSheet.pdf (accessed on 5 March 2025).
- NYC Health Cigualtera Fish Poisoning—NYC Health. Available online: https://www.nyc.gov/site/doh/health/health-topics/cigualtera-fish-poisoning.page (accessed on 3 June 2025).
- Traylor, J.; Murray, B.P.; Singhal, M. Ciguatera Toxicity. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Heudorf, U.; Neitzert, V.; Spark, J. Particulate Matter and Carbon Dioxide in Classrooms—The Impact of Cleaning and Ventilation. Int. J. Hyg. Environ. Health 2009, 212, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.M.; Bedard, K.; Breining, T.; Cribb, A.E. Disruption of the Endoplasmic Reticulum by Cytotoxins in LLC-PK1 Cells. Toxicol. Lett. 2005, 159, 154–163. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Ouyang, Z.; Su, Z.; Wang, D.; Yu, X. Neuroprotective Effects of Adenosine Isolated from Cordyceps Cicadae against Oxidative and ER Stress Damages Induced by Glutamate in PC12 Cells. Environ. Toxicol. Pharmacol. 2016, 44, 53–61. [Google Scholar] [CrossRef]
- Jiménez-Cárcamo, D.; Garcia, C.; Contreras, H. Toxins of Okadaic Acid-Group Increase Malignant Properties in Cells of Colon Cancer. Available online: https://www.mdpi.com/2072-6651/12/3/179 (accessed on 3 June 2025).
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pásaro, E.; Méndez, J.; Laffon, B. Okadaic Acid: More than a Diarrheic Toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed]
- Landwehr, J. FDA Issues Warning Over Paralytic Shellfish Poisoning After Oyster, Clam Recall—What to Know. Available online: https://www.health.com/fda-shellfish-paralytic-poisoning-recall-8663030 (accessed on 3 June 2025).
- Griffiths, D.; Saker, M. The Palm Island Mystery Disease 20 Years on: A Review of Research on the Cyanotoxin Cylindrospermopsin. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/tox.10103?casa_token=pgVhOOhMR7cAAAAA:L2U6nDjB91iYknda9QA4VUP66J92sKHSDey80O70p7w_Cf43Azzbah8qmZMLmXRiXJw_EaNS0DOJ3sn5DA (accessed on 5 March 2025).
- Poniedziałek, B.; Rzymski, P.; Kokociński, M. Cylindrospermopsin: Water-Linked Potential Threat to Human Health in Europe. Environ. Toxicol. Pharmacol. 2012, 34, 651–660. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Marañón, E. Cell Size as a Key Determinant of Phytoplankton Metabolism and Community Structure. Annu. Rev. Mar. Sci. 2015, 7, 241–264. [Google Scholar] [CrossRef]
- Trainer, V.L.; Moore, S.K.; Hallegraeff, G.; Kudela, R.M.; Clement, A.; Mardones, J.I.; Cochlan, W.P. Pelagic Harmful Algal Blooms and Climate Change: Lessons from Nature’s Experiments with Extremes. Harmful Algae 2020, 91, 101591. [Google Scholar] [CrossRef]
- Segura, A.M.; Piccini, C.; Nogueira, L.; Alcántara, I.; Calliari, D.; Kruk, C. Increased Sampled Volume Improves Microcystis Aeruginosa Complex (MAC) Colonies Detection and Prediction Using Random Forests. Ecol. Indic. 2017, 79, 347–354. [Google Scholar] [CrossRef]
- Gholizadeh, M.H.; Melesse, A.M.; Reddi, L. A Comprehensive Review on Water Quality Parameters Estimation Using Remote Sensing Techniques. Sensors 2016, 16, 1298. [Google Scholar] [CrossRef] [PubMed]
- Sòria-Perpinyà, X.; Vicente, E.; Urrego, P.; Pereira-Sandoval, M.; Ruíz-Verdú, A.; Delegido, J.; Soria, J.M.; Moreno, J. Remote Sensing of Cyanobacterial Blooms in a Hypertrophic Lagoon (Albufera of València, Eastern Iberian Peninsula) Using Multitemporal Sentinel-2 Images. Sci. Total Environ. 2020, 698, 134305. [Google Scholar] [CrossRef]
- Astuti, W.; Govindaraju, R. Characterization and Modeling of Harmful Algal Blooms: A Review. J. Hydraul. Eng. 2025, 151, 03125001. [Google Scholar] [CrossRef]
- Xu, S.; Lyu, P.; Zheng, X.; Yang, H.; Xia, B.; Li, H.; Zhang, H.; Ma, S. Monitoring and Control Methods of Harmful Algal Blooms in Chinese Freshwater System: A Review. Environ. Sci. Pollut. Res. 2022, 29, 56908–56927. [Google Scholar] [CrossRef]
- Busch, J.A.; Andree, K.B.; Diogène, J.; Fernández-Tejedor, M.; Toebe, K.; John, U.; Krock, B.; Tillmann, U.; Cembella, A.D. Toxigenic Algae and Associated Phycotoxins in Two Coastal Embayments in the Ebro Delta (NW Mediterranean). Harmful Algae 2016, 55, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Luerssen, R.M.; Thomas, A.C.; Hurst, J. Relationships between Satellite-Measured Thermal Features and Alexandrium-Imposed Toxicity in the Gulf of Maine. Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2656–2673. [Google Scholar] [CrossRef]
- Stumpf, R.P.; Culver, M.E.; Tester, P.A.; Tomlinson, M.; Kirkpatrick, G.J.; Pederson, B.A.; Truby, E.; Ransibrahmanakul, V.; Soracco, M. Monitoring Karenia Brevis Blooms in the Gulf of Mexico Using Satellite Ocean Color Imagery and Other Data. Harmful Algae 2003, 2, 147–160. [Google Scholar] [CrossRef]
- Anderson, D.M. Approaches to Monitoring, Control and Management of Harmful Algal Blooms (HABs). Ocean Coast. Manag. 2009, 52, 342–347. [Google Scholar] [CrossRef]
- Brooks, C. A Breakthrough in Harmful Algal Bloom Monitoring with FlowCam. Available online: https://www.fluidimaging.com/blog/flow-imaging-microscopy-breakthrough-harmful-algal-bloom-monitoring-with-flowcam (accessed on 12 March 2025).
- Monitoring Harmful Algal Blooms Using Drones. Available online: https://www.unh.edu/unhtoday/monitoring-harmful-algal-blooms-using-drones (accessed on 12 March 2025).
- Arias, F.; Zambrano, M.; Galagarza, E.; Broce, K. Mapping Harmful Algae Blooms: The Potential of Hyperspectral Imaging Technologies. Remote Sens. 2025, 17, 608. [Google Scholar] [CrossRef]
- Gupta, A.; Hantush, M.M.; Govindaraju, R.S. Sub-Monthly Time Scale Forecasting of Harmful Algal Blooms Intensity in Lake Erie Using Remote Sensing and Machine Learning. Sci. Total Environ. 2023, 900, 165781. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Yang, J.; Li, Y.; Gao, L. Harmful Algal Bloom Warning Based on Machine Learning in Maritime Site Monitoring. Knowl.-Based Syst. 2022, 245, 108569. [Google Scholar] [CrossRef]
- Cruz, R.C.; Costa, P.R.; Vinga, S.; Krippahl, L.; Lopes, M.B. A Review of Recent Machine Learning Advances for Forecasting Harmful Algal Blooms and Shellfish Contamination. J. Mar. Sci. Eng. 2021, 9, 283. [Google Scholar] [CrossRef]
- Nagkoulis, N.; Vasiloudis, G.; Moumtzidou, A.; Gialampoukidis, I.; Vrochidis, S.; Kompatsiaris, I. Forecasting Lakes’ Chlorophyll Concentrations Using Satellite Images and Generative Adversarial Networks. Water Resour. Res. 2024, 60, e2024WR037138. [Google Scholar] [CrossRef]
- Park, J.; Patel, K.; Lee, W.H. Recent Advances in Algal Bloom Detection and Prediction Technology Using Machine Learning. Sci. Total Environ. 2024, 938, 173546. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Noga, E.J.; Hobbs, C.H.; Glasgow, H.B. New “phantom” Dinoflagellate Is the Causative Agent of Major Estuarine Fish Kills. Nature 1992, 358, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Glasgow, H.B., Jr.; Burkholder, J.M.; Schmechel, D.E.; Tester, P.A.; Rublee, P.A. Insidious Effects of a Toxic Estuarine Dino Flagellate on Fish Survival and Human Health. J. Toxicol. Environ. Health 1995, 46, 501–522. [Google Scholar] [CrossRef]
- Grattan, L.M.; Oldach, D.; Perl, T.M.; Lowitt, M.H.; Matuszak, D.L.; Dickson, C.; Parrott, C.; Shoemaker, R.C.; Kauffman, C.L.; Wasserman, M.P.; et al. Learning and Memory Difficulties after Environmental Exposure to Waterways Containing Toxin-Producing Pfiesteria or Pfiesteria-like Dinoflagellates. Lancet 1998, 352, 532–539. [Google Scholar] [CrossRef]
- Haselow, D.T.; Brown, E.; Tracy, J.K.; Magnien, R.; Grattan, L.M.; Morris, J.G., Jr.; Oldach, D.W. Gastrointestinal and Respiratory Tract Symptoms Following Brief Environmental Exposure to Aerosols during a Pfiesteria-Related Fish Kill. J. Toxicol. Environ. Health—Part A 2001, 63, 553–564. [Google Scholar] [CrossRef]
- Hudnell, H.K. The State of U.S. Freshwater Harmful Algal Blooms Assessments, Policy and Legislation. Toxicon 2010, 55, 1024–1034. [Google Scholar] [CrossRef]
- Shoemaker, R.C. Residential and Recreational Acquisition of Possible Estuary-Associated Syndrome: A New Approach to Successful Diagnosis and Treatment. Environ. Health Perspect. 2001, 109, 791–796. [Google Scholar] [CrossRef] [PubMed][Green Version]
- United States Environmental Protection Agency. Control Measures for Cyanobacterial HABs in Surface Water. Available online: https://www.epa.gov/habs/control-measures-cyanobacterial-habs-surface-water (accessed on 13 March 2025).
- Jacobi, A.L.; Thuneibat, M.; Vigar, M.K.; Rutt, C.; Andújar, A.; Roberts, V.A. Public Awareness and Concern about Harmful Algal Blooms—United States, 2020. J. Water Health 2024, 22, 1337–1346. [Google Scholar] [CrossRef]
- Zhang, P.; Zhu, S.; Xiong, C.; Yan, B.; Wang, Z.; Li, K.; Olivier, I.; Wang, H. Flocculation of Chlorella Vulgaris–Induced Algal Blooms: Critical Conditions and Mechanisms. Environ. Sci. Pollut. Res. 2022, 29, 78809–78820. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).