
Distribution Patterns and Human Exposure Risks of Microplastics in Dominant Wild Edible Shrimp: A Case Study of Haizhou Bay Marine Ranch

Abstract
1. Introduction
2. Materials and Methods
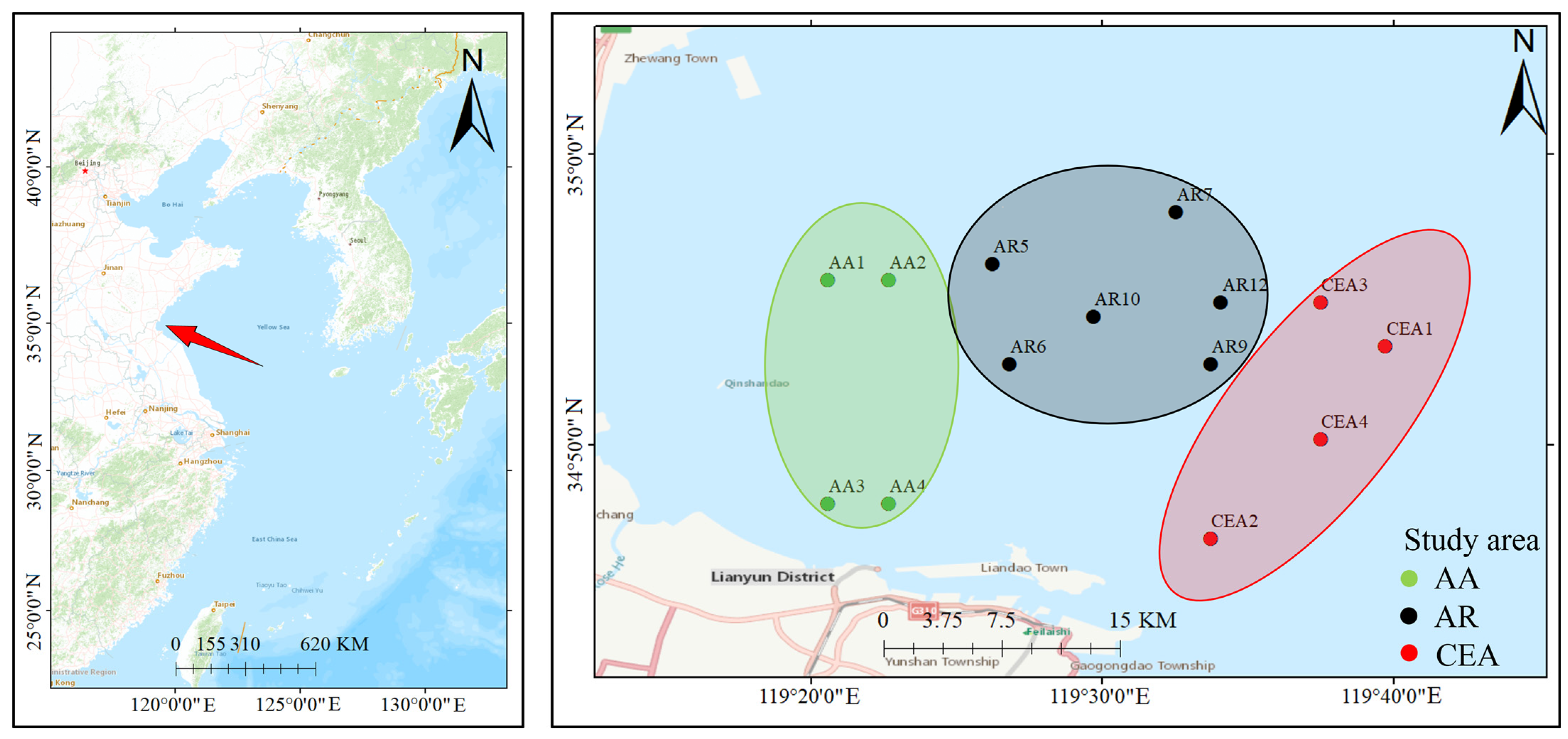
2.1. Survey Area and Sampling Collection
2.2. Sample Pretreatment
2.3. MP Extraction and FT–IR Identification
2.4. Quality Assurance and Control
2.5. Stable Isotope Analysis
2.6. Bioaccumulation Factor Analysis
2.7. Risk Assessment
2.7.1. Polymer Hazard Index (H Score)
2.7.2. Estimated Daily Intake (EDI) and Margin of Exposure (MOE)
2.8. Statistical Analysis
3. Results
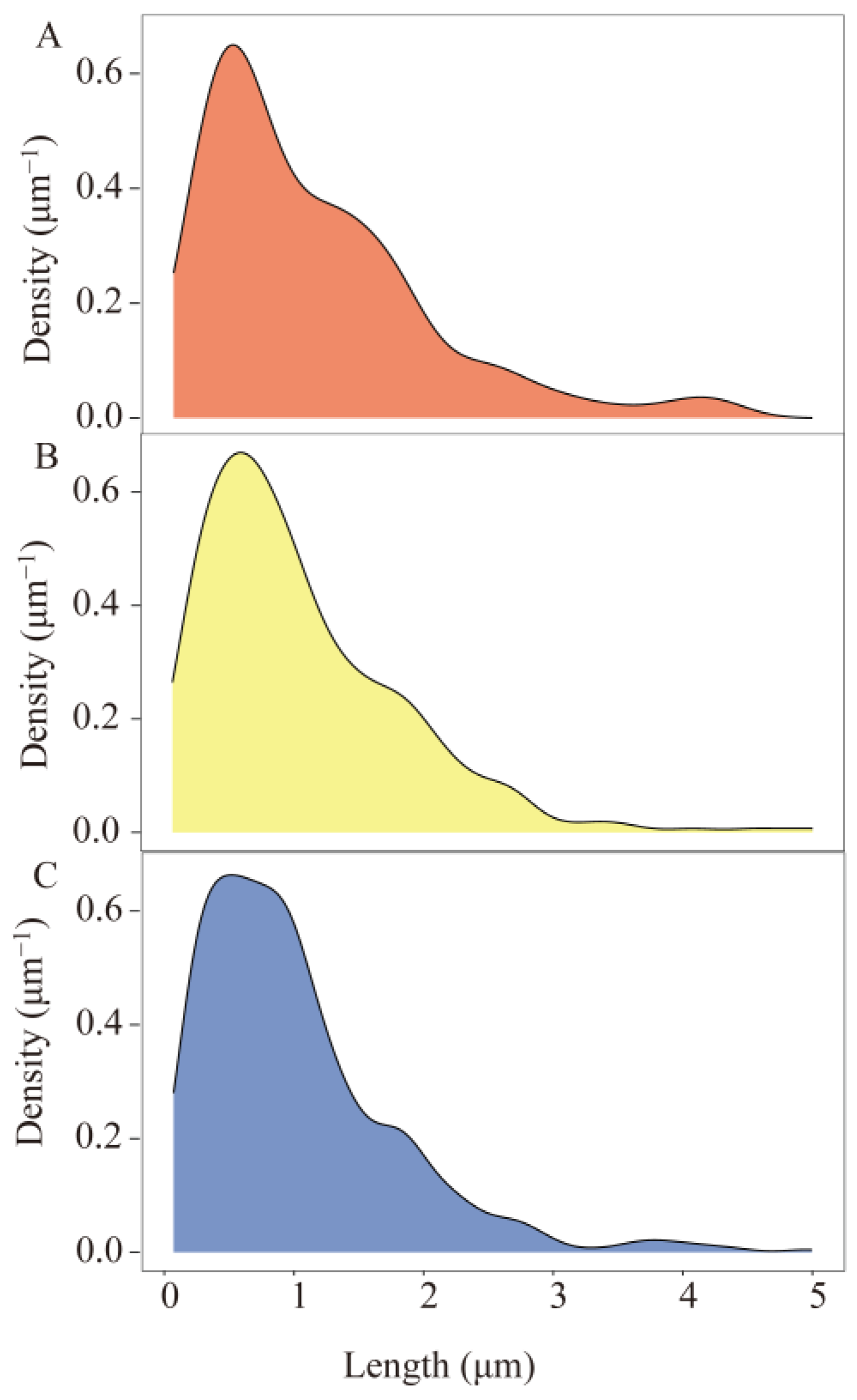
3.1. Morphological Characteristics of MPs in Shrimp
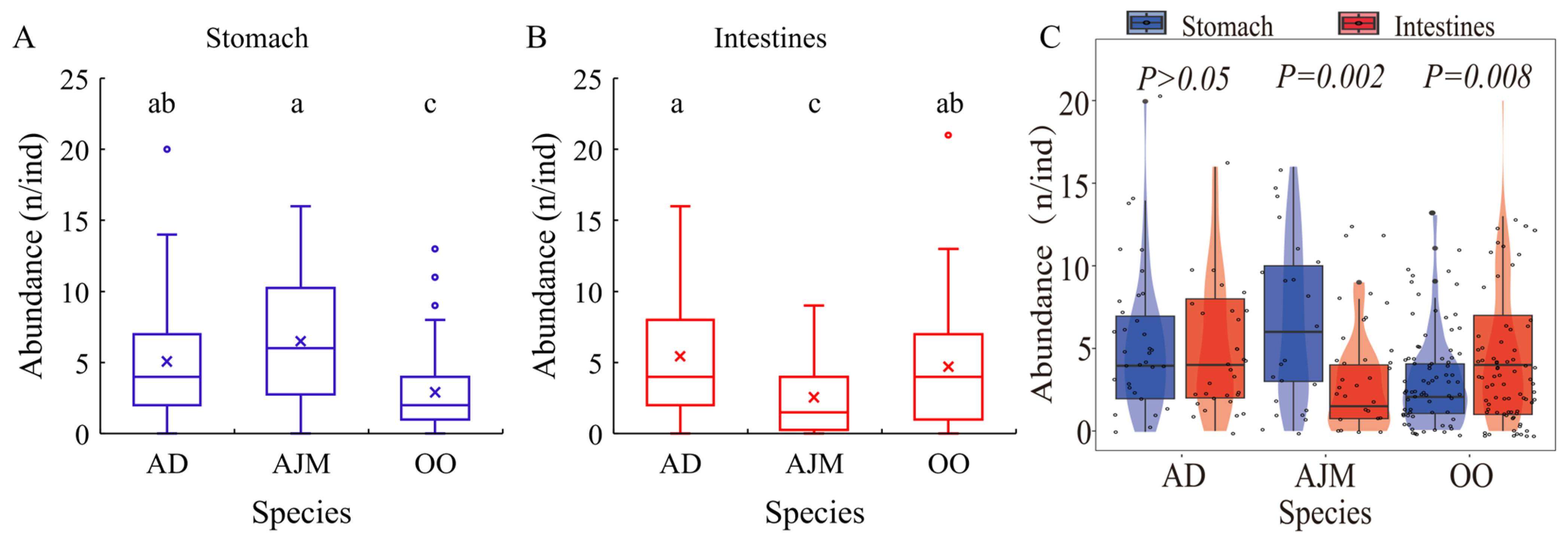
3.2. The Abundance and Distribution of MPs in Dominant Species
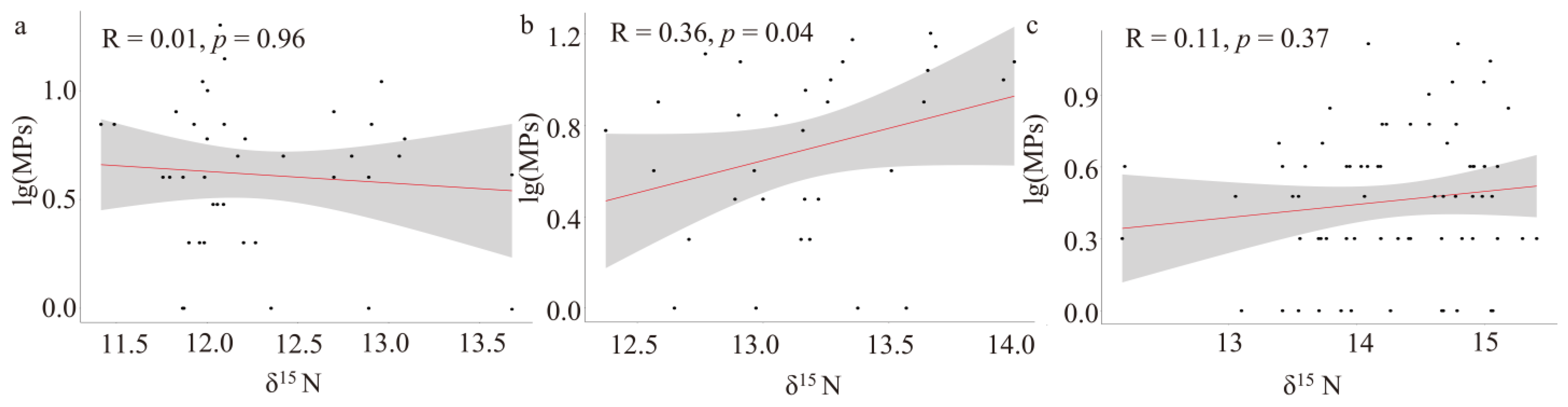
3.3. The Relationship Between the Distribution of MPs and Biological Indicators
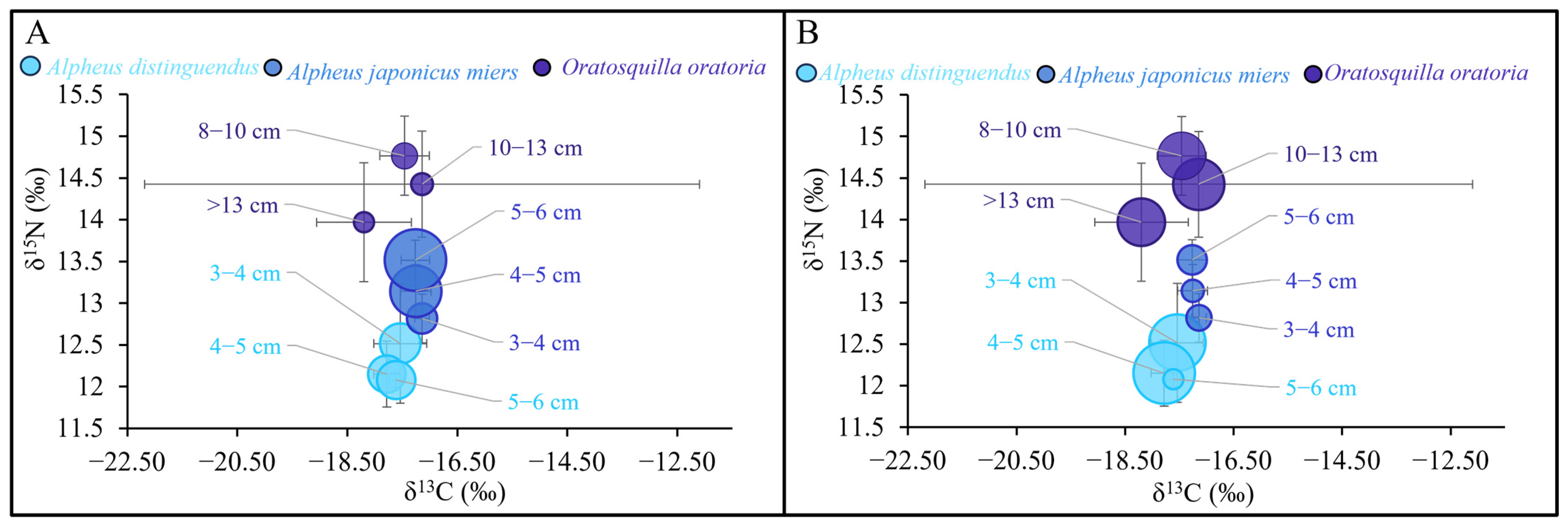
3.3.1. Distribution of MPs in the Gastrointestinal Tracts of Dominant Species in Different Body-Length Groups
3.3.2. The Correlation Between the Distribution of MPs in Dominant Species and Biological Indicators
3.3.3. Changes in MP Abundance in Dominant Species with Nutrient Levels
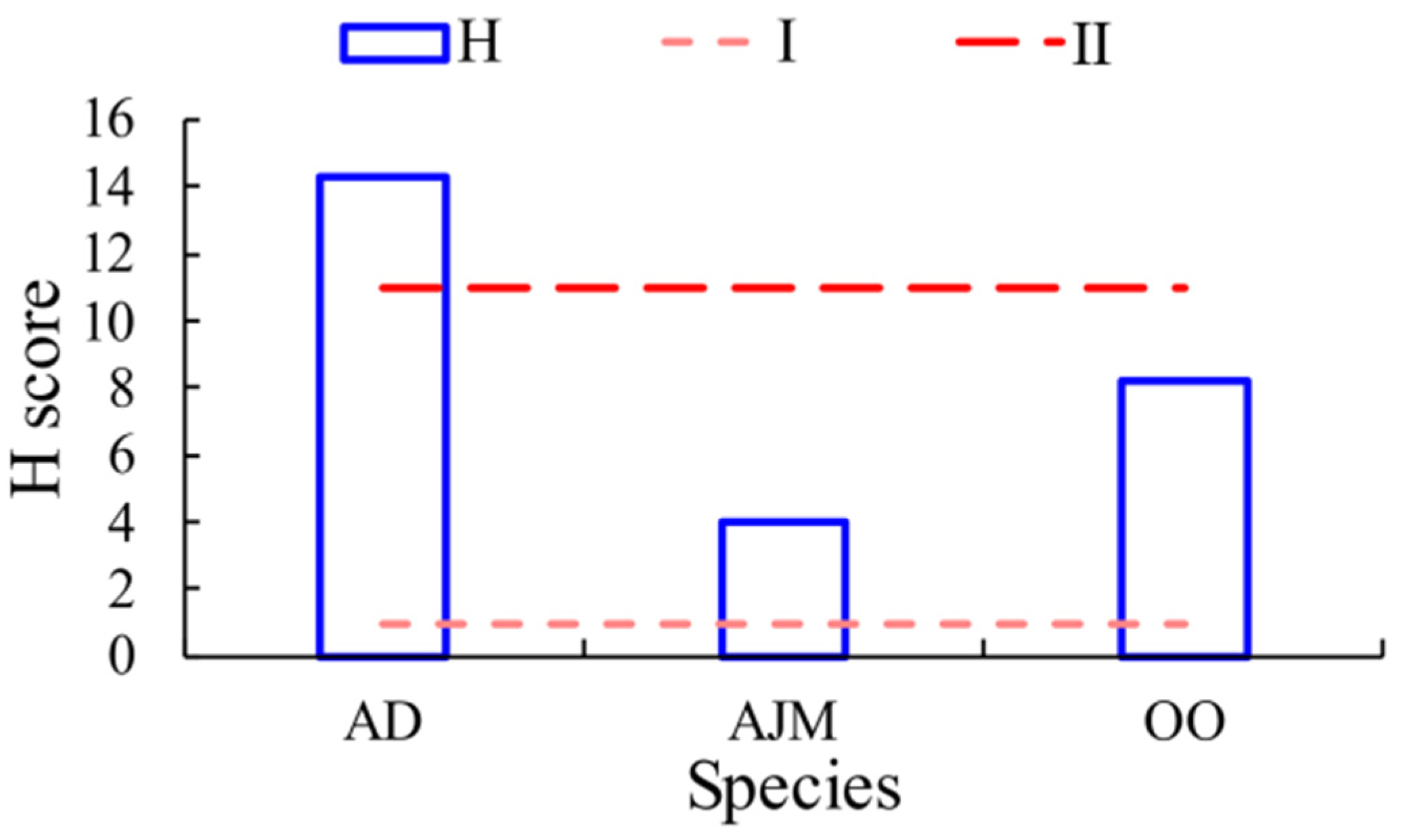
3.4. Health Risk Assessment
3.4.1. Hazard Index of Dominant Species’ MPs (H Score)
3.4.2. EDI and MOE Risk Assessment
4. Discussion
4.1. Composition of MPs in Shrimp Samples
4.2. Analysis of Gastrointestinal MP Pollution Characteristics in Dominant Shrimp Species
4.3. Relationship Between the Distribution of MPs in Dominant Species and Biological Indicators
4.4. Health Risks
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acaroğlu, H.; Güllü, M.; Sivri, N.; Marquez, F.P.G. How can there be an economic transition to a green ecosystem by adapting plastic-to-fuel technologies through renewable energy? Sustain. Energy Technol. Assess. 2024, 64, 103691. [Google Scholar] [CrossRef]
- Green, D.S.; Kregting, L.; Boots, B.; Blockley, D.J.; Brickle, P.; da Costa, M.; Crowley, Q. A comparison of sampling methods for seawater microplastics and a first report of the microplastic litter in coastal waters of Ascension and Falkland Islands. Trends Anal. Chem. 2018, 116, 346–359. [Google Scholar] [CrossRef]
- Yin, J.; Li, J.-Y.; Craig, N.J.; Su, L. Microplastic pollution in wild populations of decapod crustaceans: A review. Chemosphere 2022, 291, 132985. [Google Scholar] [CrossRef]
- Lebreton, L.C.M.; van der Zwet, J.; Damsteeg, J.-W.; Slat, B.; Andrady, A.; Reisser, J. River plastic emissions to the world’s oceans. Nat. Commun. 2017, 8, 15611. [Google Scholar] [CrossRef] [PubMed]
- Maione, C. Quantifying plastics waste accumulations on coastal tourism sites in Zanzibar, Tanzania. Mar. Pollut. Bull. 2021, 168, 112418. [Google Scholar] [CrossRef]
- Macfadyen, G.; Huntington, T.; Cappell, R. Abandoned, Lost or Otherwise Discarded Fishing Gear; United Nations Environment Programme: Food and Agriculture Organization of the United Nations: Nairobi, Kenya, 2009. [Google Scholar]
- Syversen, T.; Lilleng, G.; Vollstad, J.; Hanssen, B.J.; Sønvisen, S.A. Oceanic plastic pollution caused by Danish seine fishing in Norway. Mar. Pollut. Bull. 2022, 179, 113711. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.; Allen, S.; Abbasi, S.; Baker, A.; Bergmann, M.; Brahney, J.; Butler, T.; Duce, R.A.; Eckhardt, S.; Evangeliou, N.; et al. Microplastics and nanoplastics in the marine-atmosphere environment. Nat. Rev. Earth Environ. 2022, 3, 393–405. [Google Scholar] [CrossRef]
- Eriksen, M.; Lebreton, L.C.M.; Carson, H.S.; Thiel, M.; Moore, C.J.; Borerro, J.C.; Galgani, F.; Ryan, P.G.; Reisser, J. Plastic Pollution in the World’s Oceans: More than 5 Trillion Plastic Pieces Weighing over 250,000 Tons Afloat at Sea. PLoS ONE 2014, 9, e111913. [Google Scholar] [CrossRef]
- Guan, J.; Qi, K.; Wang, J.; Wang, W.; Wang, Z.; Lu, N.; Qu, J. Microplastics as an emerging anthropogenic vector of trace metals in freshwater: Significance of biofilms and comparison with natural substrates. Water Res. 2020, 184, 116205. [Google Scholar] [CrossRef]
- Kinigopoulou, V.; Pashalidis, I.; Kalderis, D.; Anastopoulos, I. Microplastics as carriers of inorganic and organic contaminants in the environment: A review of recent progress. J. Mol. Liq. 2022, 350, 118580. [Google Scholar] [CrossRef]
- Mei, G.; Zhang, X.; Gu, J.; Fang, Y.; Yang, W. Assessment of Heavy Metals, Polycyclic Aromatic Hydrocarbons, and Perfluorinated Alkyl Substances in two Marine Crustaceans (Oratosquilla oratoria and Portunus trituberculatus) in the Zhoushan Fishing Ground of China East Sea. J. Ocean Univ. China 2021, 20, 1587–1596. [Google Scholar] [CrossRef]
- Xie, Q.; Li, H.-X.; Lin, L.; Li, Z.-L.; Huang, J.-S.; Xu, X.-R. Characteristics of expanded polystyrene microplastics on island beaches in the Pearl River Estuary: Abundance, size, surface texture and their metals-carrying capacity. Ecotoxicology 2021, 30, 1632–1643. [Google Scholar] [CrossRef] [PubMed]
- Farrell, P.; Nelson, K. Trophic level transfer of microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ. Pollut. 2013, 177, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Sharifinia, M.; Bahmanbeigloo, Z.A.; Keshavarzifard, M.; Khanjani, M.H.; Lyons, B.P. Microplastic pollution as a grand challenge in marine research: A closer look at their adverse impacts on the immune and reproductive systems. Ecotoxicol. Environ. Saf. 2020, 204, 111109. [Google Scholar] [CrossRef]
- Wen, J.; Li, T.; Pu, Q.; Li, Y.; Ding, X.; Wang, L.; Li, X. Co-exposure of TMPs and antibiotics in zebrafish: The influence of additives on the risk of hepatotoxicity. Environ. Res. 2025, 275, 121430. [Google Scholar] [CrossRef]
- Luan, J.; Wen, L.; Bao, Y.; Bai, H.; Zhao, C.; Zhang, S.; Man, X.; Yin, T.; Feng, X. Systemic toxicity of biodegradable polyglycolic acid microplastics on the gut-liver-brain axis in zebrafish. Sci. Total. Environ. 2024, 954, 176898. [Google Scholar] [CrossRef]
- Hossain, M.S.; Rahman, M.S.; Uddin, M.N.; Sharifuzzaman, S.; Chowdhury, S.R.; Sarker, S.; Chowdhury, M.S.N. Microplastic contamination in Penaeid shrimp from the Northern Bay of Bengal. Chemosphere 2020, 238, 124688. [Google Scholar] [CrossRef]
- Rochman, C.M.; Hoellein, T. The global odyssey of plastic pollution. Science 2020, 368, 1184–1185. [Google Scholar] [CrossRef]
- Graham, E.R.; Thompson, J.T. Deposit- and suspension-feeding sea cucumbers (Echinodermata) ingest plastic fragments. J. Exp. Mar. Biol. Ecol. 2009, 368, 22–29. [Google Scholar] [CrossRef]
- Gao, S.; Yan, K.; Liang, B.; Shu, R.; Wang, N.; Zhang, S. The different ways microplastics from the water column and sediment accumulate in fish in Haizhou Bay. Sci. Total. Environ. 2022, 854, 158575. [Google Scholar] [CrossRef]
- Hara, J.; Frias, J.; Nash, R. Quantification of microplastic ingestion by the decapod crustacean Nephrops norvegicus from Irish waters. Mar. Pollut. Bull. 2020, 152, 110905. [Google Scholar] [CrossRef]
- Moore, R.; Loseto, L.; Noel, M.; Etemadifar, A.; Brewster, J.; MacPhee, S.; Bendell, L.; Ross, P. Microplastics in beluga whales (Delphinapterus leucas) from the Eastern Beaufort Sea. Mar. Pollut. Bull. 2020, 150, 110723. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Liang, J.; Zhu, M.; Zhao, Y.; Zhang, B. Microplastics in seawater and zooplankton from the Yellow Sea. Environ. Pollut. 2018, 242, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Waite, H.R.; Donnelly, M.J.; Walters, L.J. Quantity and types of microplastics in the organic tissues of the eastern oyster Crassostrea virginica and Atlantic mud crab Panopeus herbstii from a Florida estuary. Mar. Pollut. Bull. 2018, 129, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Mercy, F.T.; Alam, A.R. Assessment of microplastic contamination in shrimps from the Bay of Bengal and associated human health risk. Mar. Pollut. Bull. 2024, 201, 116185. [Google Scholar] [CrossRef]
- Devriese, L.I.; van der Meulen, M.D.; Maes, T.; Bekaert, K.; Paul-Pont, I.; Frère, L.; Robbens, J.; Vethaak, A.D. Microplastic contamination in brown shrimp (Crangon crangon, Linnaeus 1758) from coastal waters of the Southern North Sea and Channel area. Mar. Pollut. Bull. 2015, 98, 179–187. [Google Scholar] [CrossRef]
- Walkinshaw, C.; Lindeque, P.K.; Thompson, R.; Tolhurst, T.; Cole, M. Microplastics and seafood: Lower trophic organisms at highest risk of contamination. Ecotoxicol. Environ. Saf. 2020, 190, 110066. [Google Scholar] [CrossRef]
- Carreras-Colom, E.; Constenla, M.; Soler-Membrives, A.; Cartes, J.E.; Baeza, M.; Padrós, F.; Carrassón, M. Spatial occurrence and effects of microplastic ingestion on the deep-water shrimp Aristeus antennatus. Mar. Pollut. Bull. 2018, 133, 44–52. [Google Scholar] [CrossRef]
- Curren, E.; Leaw, C.P.; Lim, P.T.; Leong, S.C.Y. Evidence of Marine Microplastics in Commercially Harvested Seafood. Front. Bioeng. Biotechnol. 2020, 8, 562760. [Google Scholar] [CrossRef]
- Liang, B.; Gao, S.; Wang, Z.; Shu, R.; Wang, N.; Tan, W.; Gao, C.; Zhang, S. Spatial distribution characteristics of microplastics in the seawater column and sediments of the artificial reef area and adjacent water in Haizhou Bay. Sci. Total. Environ. 2023, 900, 166236. [Google Scholar] [CrossRef]
- McGregor, S.; Strydom, N.A. Feeding ecology and microplastic ingestion in Chelon richardsonii (Mugilidae) associated with surf diatom Anaulus australis accumulations in a warm temperate South African surf zone. Mar. Pollut. Bull. 2020, 158, 111430. [Google Scholar] [CrossRef]
- RB/T 061-2021; Technical Specifications for Euthanasia in Animal Experiments. The Standardization Administration of the People’s Republic of China: Sichuan, China, 2021.
- GB/T 12763.6-2007; Specifications for Oceanographic Survey—Part 6: Marine Biological Survey. People’s Republic of China Standardization Administration of China: Beijing, China, 2007.
- Parvin, F.; Jannat, S.; Tareq, S.M. Abundance, characteristics and variation of microplastics in different freshwater fish species from Bangladesh. Sci. Total. Environ. 2021, 784, 147137. [Google Scholar] [CrossRef]
- Shu, R.; Hu, W.; Gao, S.; Zhang, S.; Li, Z.; Liang, B.; Yu, W. Transfer pattern of microplastics at an individual level: A case study of two typical Sciaenidae fish in coastal waters. Sci. Total. Environ. 2023, 901, 165570. [Google Scholar] [CrossRef] [PubMed]
- De Witte, B.; Devriese, L.; Bekaert, K.; Hoffman, S.; Vandermeersch, G.; Cooreman, K.; Robbens, J. Quality assessment of the blue mussel (Mytilus edulis): Comparison between commercial and wild types. Mar. Pollut. Bull. 2014, 85, 146–155. [Google Scholar] [CrossRef]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhou, G.; Lu, J.; Shen, C.; Dong, Z.; Yin, S.; Li, F. Spatio-vertical distribution of riverine microplastics: Impact of the textile industry. Environ. Res. 2022, 211, 112789. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Chen, L.; Zhang, K.; Cao, Y.; Ma, Y.; Chau, H.S.; Tao, D.; Wu, C.; Li, C.; Lam, P.K. Microplastic occurrence in the northern South China Sea, A case for Pre and Post cyclone analysis. Chemosphere 2022, 296, 133980. [Google Scholar] [CrossRef]
- Xu, L.; Liu, J.; Ma, X.Y.D.; Li, Z.; He, C.; Lu, X. Facile anchoring mussel adhesive mimic tentacles on biodegradable polymer cargo carriers via self-assembly for microplastic-free cosmetics. J. Colloid Interface Sci. 2022, 612, 13–22. [Google Scholar] [CrossRef]
- Sweeting, C.J.; Polunin, N.V.C.; Jennings, S. Effects of chemical lipid extraction and arithmetic lipid correction on stable isotope ratios of fish tissues. Rapid Commun. Mass Spectrom. 2006, 20, 595–601. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Lotze, H.K.; Tittensor, D.P.; Bryndum-Buchholz, A.; Eddy, T.D.; Cheung, W.W.L.; Galbraith, E.D.; Barange, M.; Barrier, N.; Bianchi, D.; Blanchard, J.; et al. Global ensemble projections reveal trophic amplification of ocean biomass declines with climate change. Proc. Natl. Acad. Sci. USA 2019, 116, 12907–12912. [Google Scholar] [CrossRef] [PubMed]
- Lithner, D.; Larsson, Å.; Dave, G. Environmental and health hazard ranking and assessment of plastic polymers based on chemical composition. Sci. Total Environ. 2011, 409, 3309–3324. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Gao, S.; Zhang, S.; Gao, C. Distribution characteristics and ecological risk assessment of microplastics in intertidal sediments near coastal water. Mar. Environ. Res. 2024, 195, 106353. [Google Scholar] [CrossRef]
- Ferrante, M.; Pietro, Z.; Allegui, C.; Maria, F.; Antonio, C.; Pulvirenti, E.; Favara, C.; Chiara, C.; Grasso, A.; Omayma, M.; et al. Microplastics in fillets of Mediterranean seafood. A risk assessment study. Environ. Res. 2022, 204, 112247. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Nag, R.; Cummins, E. Human health concerns regarding microplastics in the aquatic environment—From marine to food systems. Sci. Total. Environ. 2022, 823, 153730. [Google Scholar] [CrossRef]
- Ra, W.-J.; Yoo, H.J.; Kim, Y.-H.; Yun, T.; Soh, B.; Cho, S.Y.; Joo, Y.; Lee, K.-W. Heavy metal concentration according to shrimp species and organ specificity: Monitoring and human risk assessment. Mar. Pollut. Bull. 2023, 197, 115761. [Google Scholar] [CrossRef]
- Zhang, K.; Zheng, S.; Liang, J.; Zhao, Y.; Li, Q.; Zhu, M.; Dai, S.; Sun, X. Microplastic load of benthic fauna in Jiaozhou Bay, China. Environ. Pollut. 2023, 320, 121073. [Google Scholar] [CrossRef]
- Kim, J.; Maruthupandy, M.; An, K.S.; Lee, K.H.; Jeon, S.; Kim, J.-S.; Cho, W.-S. Acute and subacute repeated oral toxicity study of fragmented microplastics in Sprague-Dawley rats. Ecotoxicol. Environ. Saf. 2021, 228, 112964. [Google Scholar] [CrossRef]
- Keshavarzifard, M.; Vazirzadeh, A.; Sharifinia, M. Occurrence and characterization of microplastics in white shrimp, Metapenaeus affinis, living in a habitat highly affected by anthropogenic pressures, northwest Persian Gulf. Mar. Pollut. Bull. 2021, 169, 112581. [Google Scholar] [CrossRef]
- Wang, T.; Tong, C.; Wu, F.; Jiang, S.; Zhang, S. Distribution characteristics of microplastics and corresponding feeding habits of the dominant shrimps in the rivers of Chongming Island. Sci. Total. Environ. 2023, 888, 164041. [Google Scholar] [CrossRef]
- Barrows, A.; Cathey, S.; Petersen, C. Marine environment microfiber contamination: Global patterns and the diversity of microparticle origins. Environ. Pollut. 2018, 237, 275–284. [Google Scholar] [CrossRef]
- Daniel, D.B.; Ashraf, P.M.; Thomas, S.N. Abundance, characteristics and seasonal variation of microplastics in Indian white shrimps (Fenneropenaeus indicus) from coastal waters off Cochin, Kerala, India. Sci. Total. Environ. 2020, 737, 139839. [Google Scholar] [CrossRef] [PubMed]
- Fernández Severini, M.D.; Buzzi, N.S.; Forero López, A.D.; Colombo, C.V.; Chatelain Sartor, G.L.; Rimondino, G.N.; Truchet, D.M. Chemical composition and abundance of microplastics in the muscle of commercial shrimp Pleoticus muelleri at an impacted coastal environment (Southwestern Atlantic). Mar. Pollut. Bull. 2020, 161, 111700. [Google Scholar] [CrossRef]
- Gao, S.; Li, Z.; Wang, N.; Lu, Y.; Zhang, S. Microplastics in different tissues of caught fish in the artificial reef area and adjacent waters of Haizhou Bay. Mar. Pollut. Bull. 2021, 174, 113112. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, N.; Gong, S.; Gao, S. The patterns of trophic transfer of microplastic ingestion by fish in the artificial reef area and adjacent waters of Haizhou Bay. Mar. Pollut. Bull. 2022, 177, 113565. [Google Scholar] [CrossRef]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R. Accumulation of Microplastic on Shorelines Woldwide: Sources and Sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.Y.; Zhao, S.Y.; Peng, G.Y.; Gao, L.; Li, D.J.; Li, D.J. Occurrence, characteristics of microplastic during urban sewage treatment process. Zhongguo Huanjing Kexue 2018, 38, 1734. [Google Scholar] [CrossRef]
- Pol, W.; Mierzyńska, K.; Włodarczyk, T.; Hauschild, T.; Zieliński, P. No trophy for the trophy? How lake trophy impacts bacterial assemblages of biofilm on microplastic. Ecohydrol. Hydrobiol. 2023, 23, 602–613. [Google Scholar] [CrossRef]
- Gurjar, U.R.; Xavier, M.; Nayak, B.B.; Ramteke, K.; Deshmukhe, G.; Jaiswar, A.K.; Shukla, S.P. Microplastics in shrimps: A study from the trawling grounds of north eastern part of Arabian Sea. Environ. Sci. Pollut. Res. 2021, 28, 48494–48504. [Google Scholar] [CrossRef]
- Wang, J.; Wang, M.; Ru, S.; Liu, X. High levels of microplastic pollution in the sediments and benthic organisms of the South Yellow Sea, China. Sci. Total. Environ. 2018, 651, 1661–1669. [Google Scholar] [CrossRef]
- Welden, N.A.; Cowie, P.R. Environment and gut morphology influence microplastic retention in langoustine, Nephrops norvegicus. Environ. Pollut. 2016, 214, 859–865. [Google Scholar] [CrossRef]
- Cau, A.; Avio, C.G.; Dessì, C.; Moccia, D.; Pusceddu, A.; Regoli, F.; Cannas, R.; Follesa, M.C. Benthic Crustacean Digestion Can Modulate the Environmental Fate of Microplastics in the Deep Sea. Environ. Sci. Technol. 2020, 54, 4886–4892. [Google Scholar] [CrossRef]
- Welden, N.A.; Cowie, P.R. Long-term microplastic retention causes reduced body condition in the langoustine, Nephrops norvegicus. Environ. Pollut. 2016, 218, 895–900. [Google Scholar] [CrossRef]
- Cole, M.; Coppock, R.; Lindeque, P.K.; Altin, D.; Reed, S.; Pond, D.W.; Sørensen, L.; Galloway, T.S.; Booth, A.M. Effects of Nylon Microplastic on Feeding, Lipid Accumulation, and Moulting in a Coldwater Copepod. Environ. Sci. Technol. 2019, 53, 7075–7082. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Z.; Li, M.; Jiang, Q.; Wu, D.; Huang, Y.; Jiao, Y.; Zhang, M.; Zhao, Y. Effects of nanoplastics on antioxidant and immune enzyme activities and related gene expression in juvenile Macrobrachium nipponense. J. Hazard. Mater. 2020, 398, 122990. [Google Scholar] [CrossRef] [PubMed]
- Timilsina, A.; Adhikari, K.; Yadav, A.K.; Joshi, P.; Ramena, G.; Bohara, K. Effects of microplastics and nanoplastics in shrimp: Mechanisms of plastic particle and contaminant distribution and subsequent effects after uptake. Sci. Total. Environ. 2023, 894, 164999. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Zhu, X.; Duan, Y.; Huang, J.; Nan, Y.; Zhang, J. Toxic effects of nitrite and microplastics stress on histology, oxidative stress, and metabolic function in the gills of Pacific white shrimp, Litopenaeus vannamei. Mar. Pollut. Bull. 2023, 187, 114531. [Google Scholar] [CrossRef]
- Castejón, D.; Rotllant, G.; Ribes, E.; Guerao, G. Morphological description of the midgut tract and midgut–hindgut junction in the larvae of the spider crab Maja brachydactyla Balss, 1922 (Malacostraca: Decapoda). Arthropod Struct. Dev. 2022, 70, 101168. [Google Scholar] [CrossRef]
- Welden, N.A.; Cowie, P.R.; Taylor, A.C. Growth and gut morphology of the lobster Nephrops norvegicus. J. Crustac. Biol. 2015, 35, 20–25. [Google Scholar] [CrossRef]
- Nan, B.; Su, L.; Kellar, C.; Craig, N.J.; Keough, M.J.; Pettigrove, V. Identification of microplastics in surface water and Australian freshwater shrimp Paratya australiensis in Victoria, Australia. Environ. Pollut. 2020, 259, 113865. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | A. distinguendus (De Man, 1909) | Alpheus japonicus (Miers, 1884) | Exopalaemon carinicauda (Holthuis, 1950) | Metapenaeus joyneri (Miers, 1879) | O. oratoria | Palaemon graviera (Rathbun, 1914) |
|---|---|---|---|---|---|---|
| Number | 41 | 34 | 13 | 5 | 91 | 5 |
| IRI | 2240.19 | 2624.84 | 334.34 | 162.25 | 13,176.34 | 78.60 |
| Organ | Species | Significance |
|---|---|---|
| Stomach | A. distinguendus | 0.990 |
| A. japonicus | 0.237 | |
| O. oratoria | 0.733 | |
| Gastrointestinal tract | A. distinguendus | 0.598 |
| A. japonicus | 0.799 | |
| O. oratoria | 0.628 |
| Species | 13C | 15N | TLs | 13C/15N | Carapace Length (cm) | Total Length (cm) | Weight (g) |
|---|---|---|---|---|---|---|---|
| A. distinguendus | −0.003 | 0.008 | 0.008 | −0.044 | 0.117 | 0.036 | −0.025 |
| A. japonicus | 0.341 | 0.361 * | 0.361 * | 0.218 | 0.363 * | 0.385 * | 0.352 * |
| O. oratoria | 0.153 | 0.106 | 0.106 | 0.23 | −0.181 | −0.193 | −0.244 * |
| Species | 13C | 15N | TLs | 13C/15N | Carapace Length (cm) | Total Length (cm) | Weight (g) | |
|---|---|---|---|---|---|---|---|---|
| A. distinguendus | Correlation coefficient | 0.314 | −0.396 | −0.396 | −0.092 | −0.274 | −0.106 | 0.107 |
| Sig. (2-tailed) | 0.155 | 0.068 | 0.068 | 0.685 | 0.218 | 0.639 | 0.635 | |
| A. japonicus | Correlation coefficient | −0.063 | 0.242 | 0.242 | 0.282 | 0.047 | 0.144 | 0.027 |
| Sig. (2-tailed) | 0.822 | 0.385 | 0.385 | 0.308 | 0.868 | 0.608 | 0.925 | |
| O. oratoria | Correlation coefficient | −0.166 | 0.127 | 0.127 | −0.036 | 0.102 | 0.055 | 0.076 |
| Sig. (2-tailed) | 0.239 | 0.369 | 0.369 | 0.799 | 0.473 | 0.698 | 0.594 |
| EDI (g/kg BW/d) | MOE | |
|---|---|---|
| Male | 8.17 × 10−8 | 3.06 × 105 |
| Female | 9.64 × 10−8 | 2.59 × 105 |
| p | Shape | Color | Polymer Type | Size |
|---|---|---|---|---|
| Seawater–dominants | 0.794 | 0.751 | 0.627 | 0.889 |
| Sediment–dominants | 0.945 | 0.208 | 0.779 | 0.699 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, C.; Chen, M.; Liang, B.; Cai, E.; Zhang, S.; Gao, S. Distribution Patterns and Human Exposure Risks of Microplastics in Dominant Wild Edible Shrimp: A Case Study of Haizhou Bay Marine Ranch. Water 2025, 17, 1495. https://doi.org/10.3390/w17101495
Gao C, Chen M, Liang B, Cai E, Zhang S, Gao S. Distribution Patterns and Human Exposure Risks of Microplastics in Dominant Wild Edible Shrimp: A Case Study of Haizhou Bay Marine Ranch. Water. 2025; 17(10):1495. https://doi.org/10.3390/w17101495
Chicago/Turabian StyleGao, Chunmei, Minghe Chen, Baogui Liang, En Cai, Shuo Zhang, and Shike Gao. 2025. "Distribution Patterns and Human Exposure Risks of Microplastics in Dominant Wild Edible Shrimp: A Case Study of Haizhou Bay Marine Ranch" Water 17, no. 10: 1495. https://doi.org/10.3390/w17101495
APA StyleGao, C., Chen, M., Liang, B., Cai, E., Zhang, S., & Gao, S. (2025). Distribution Patterns and Human Exposure Risks of Microplastics in Dominant Wild Edible Shrimp: A Case Study of Haizhou Bay Marine Ranch. Water, 17(10), 1495. https://doi.org/10.3390/w17101495