
Fate and Removal of Oxytetracycline and Antibiotic Resistance Genes in Vertical-Flow Constructed Wetland with Different Substrates

 ,
,
Abstract
1. Introduction
2. Materials and Methods
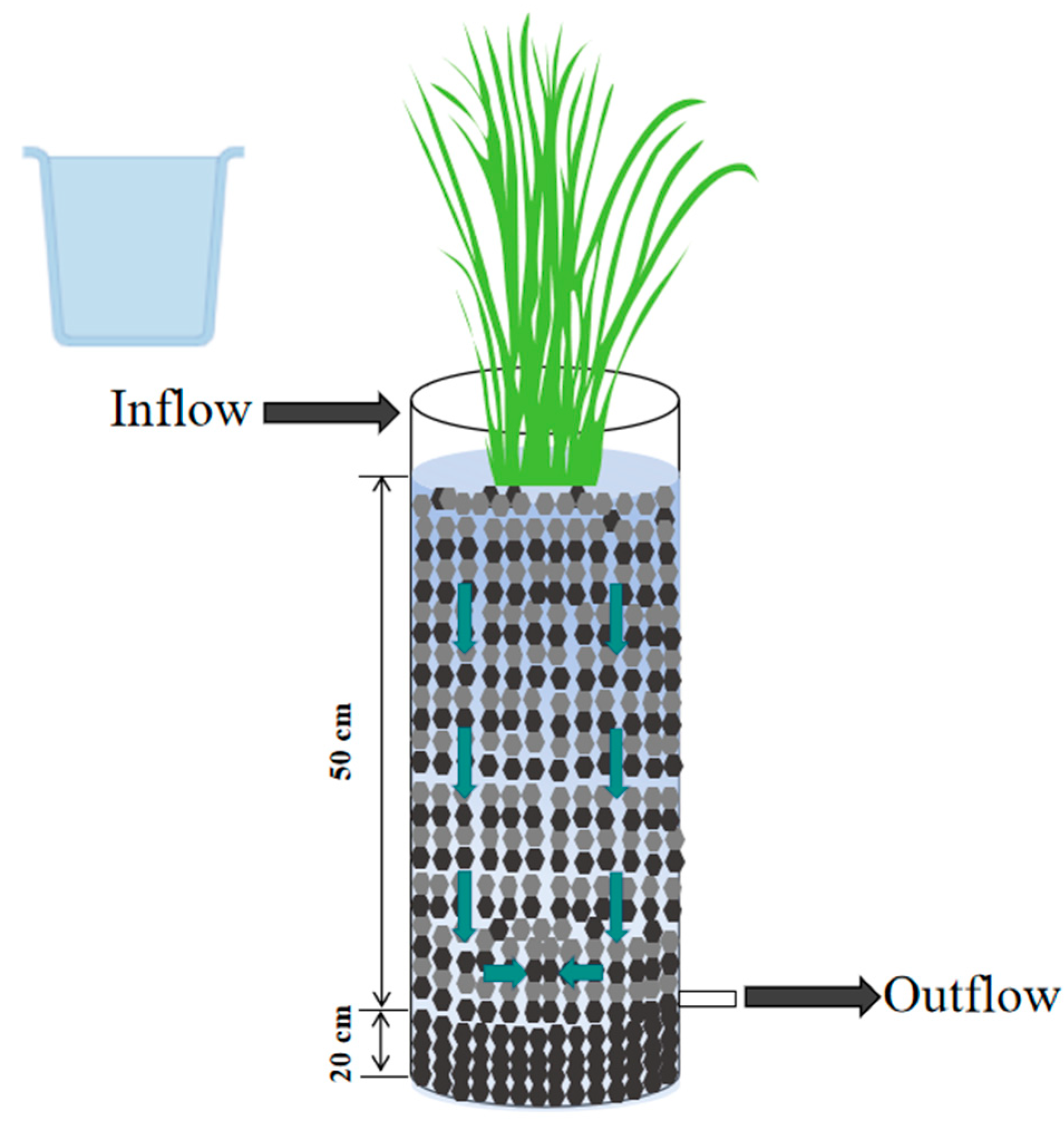
2.1. Constructed Wetland Design and Operation
2.2. Sample Collection
2.3. Detection of Antibiotics
2.4. DNA Extraction and Quantification and PCR
2.5. Microbial Community
2.6. Data Analysis
3. Results and Discussion
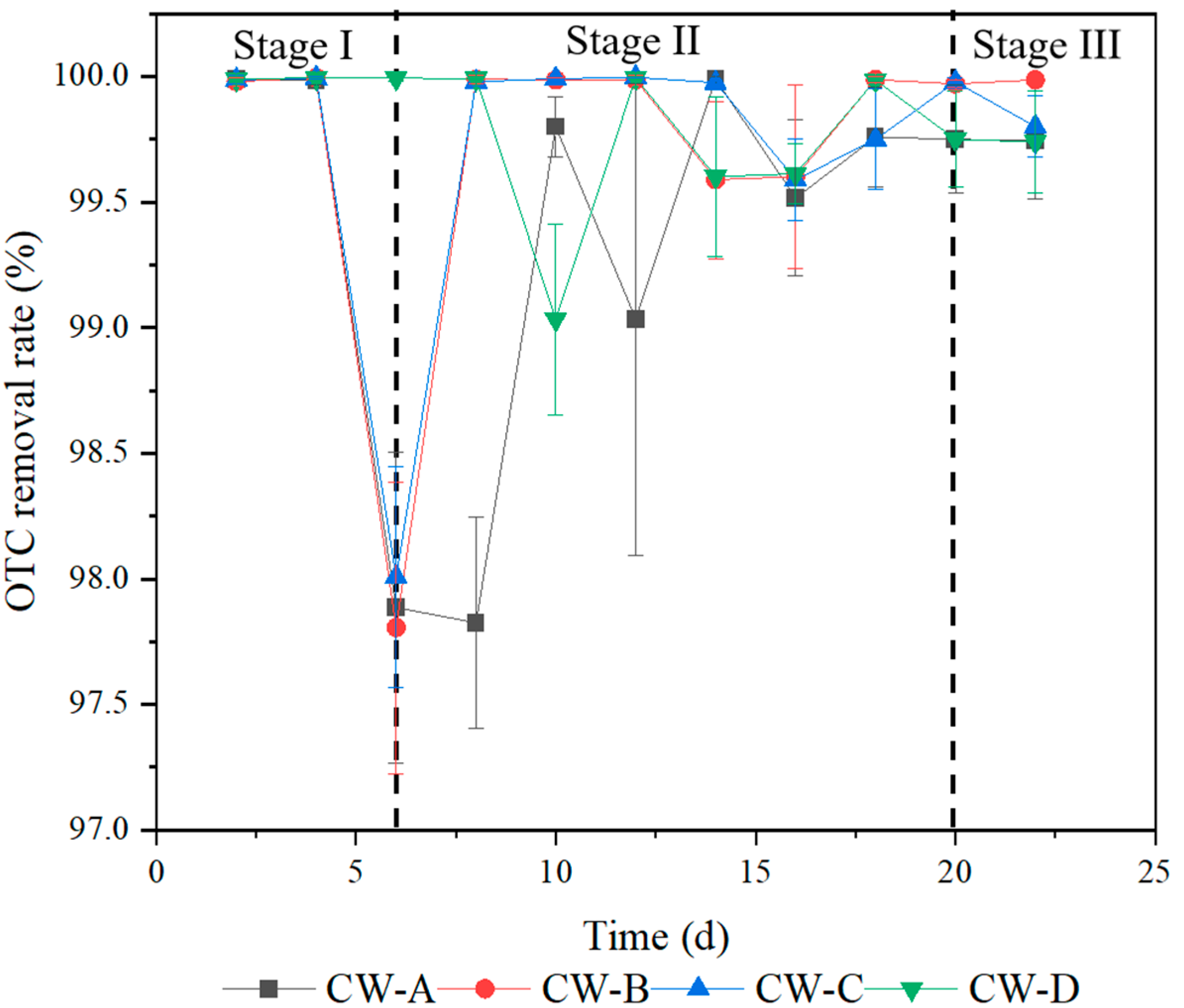
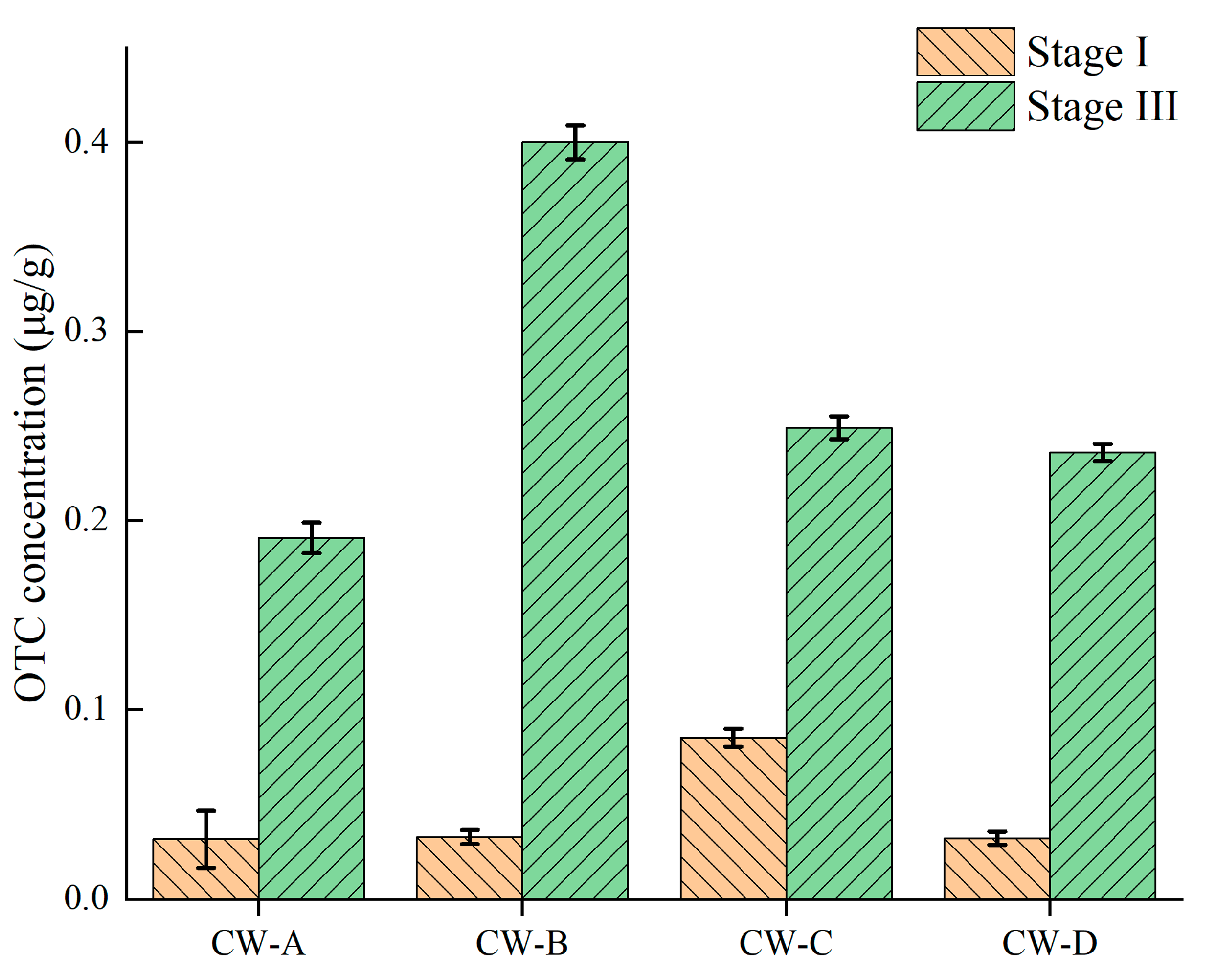
3.1. Accumulation of OTC in Constructed Wetlands
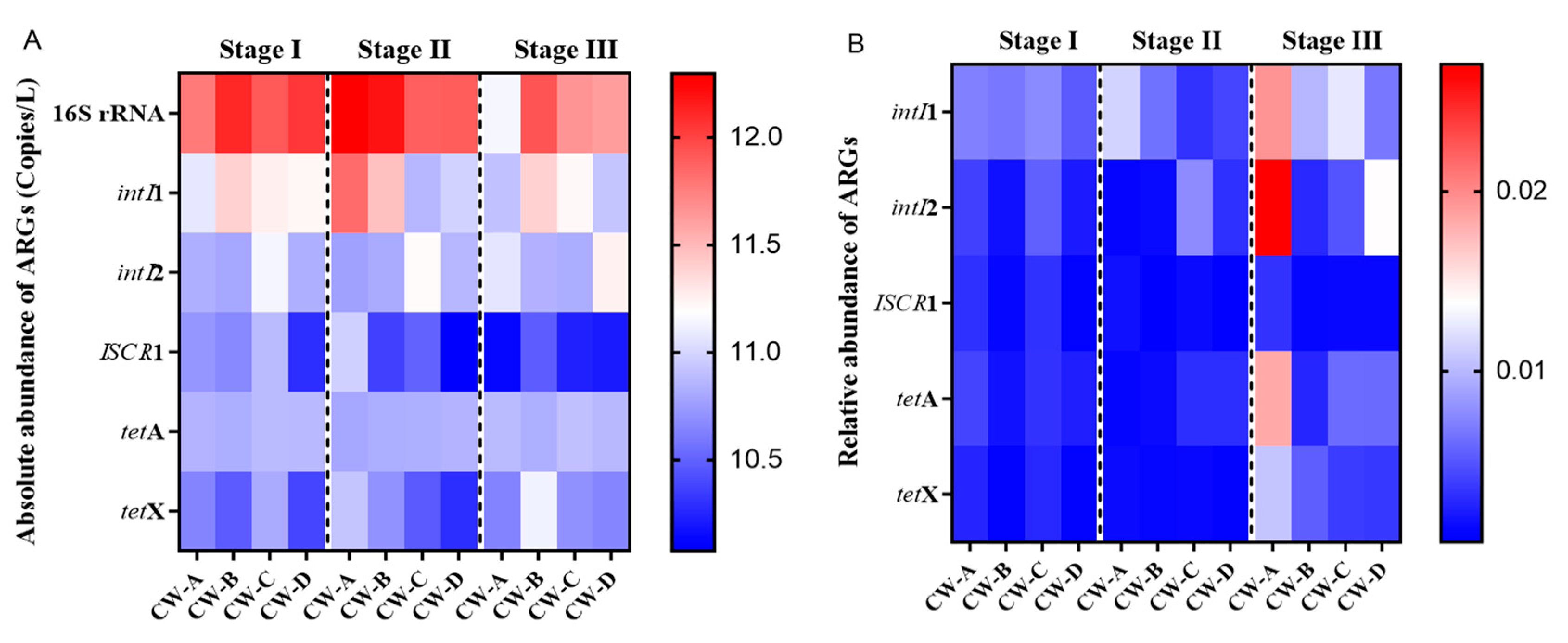
3.2. Abundance of ARGs in Constructed Wetlands
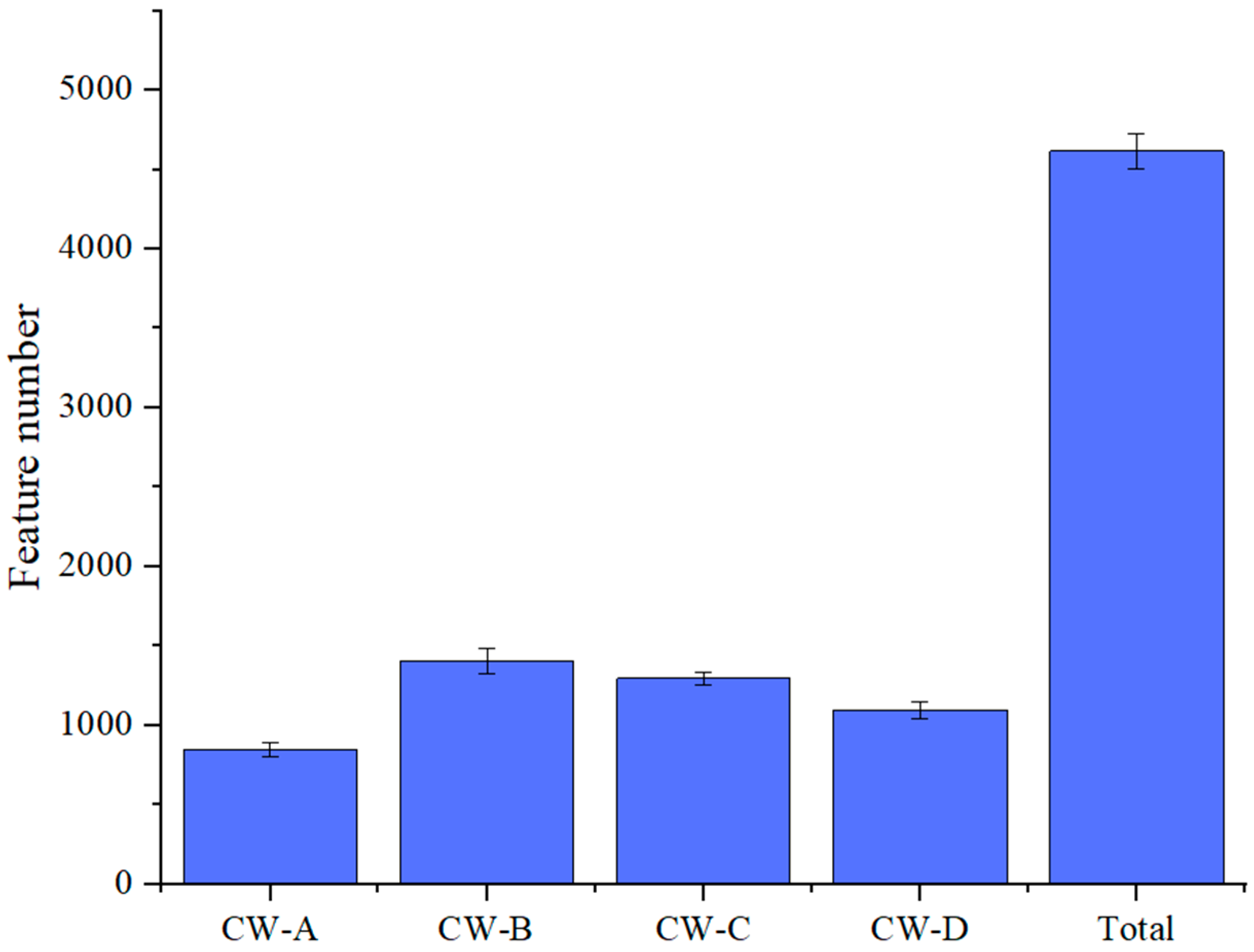
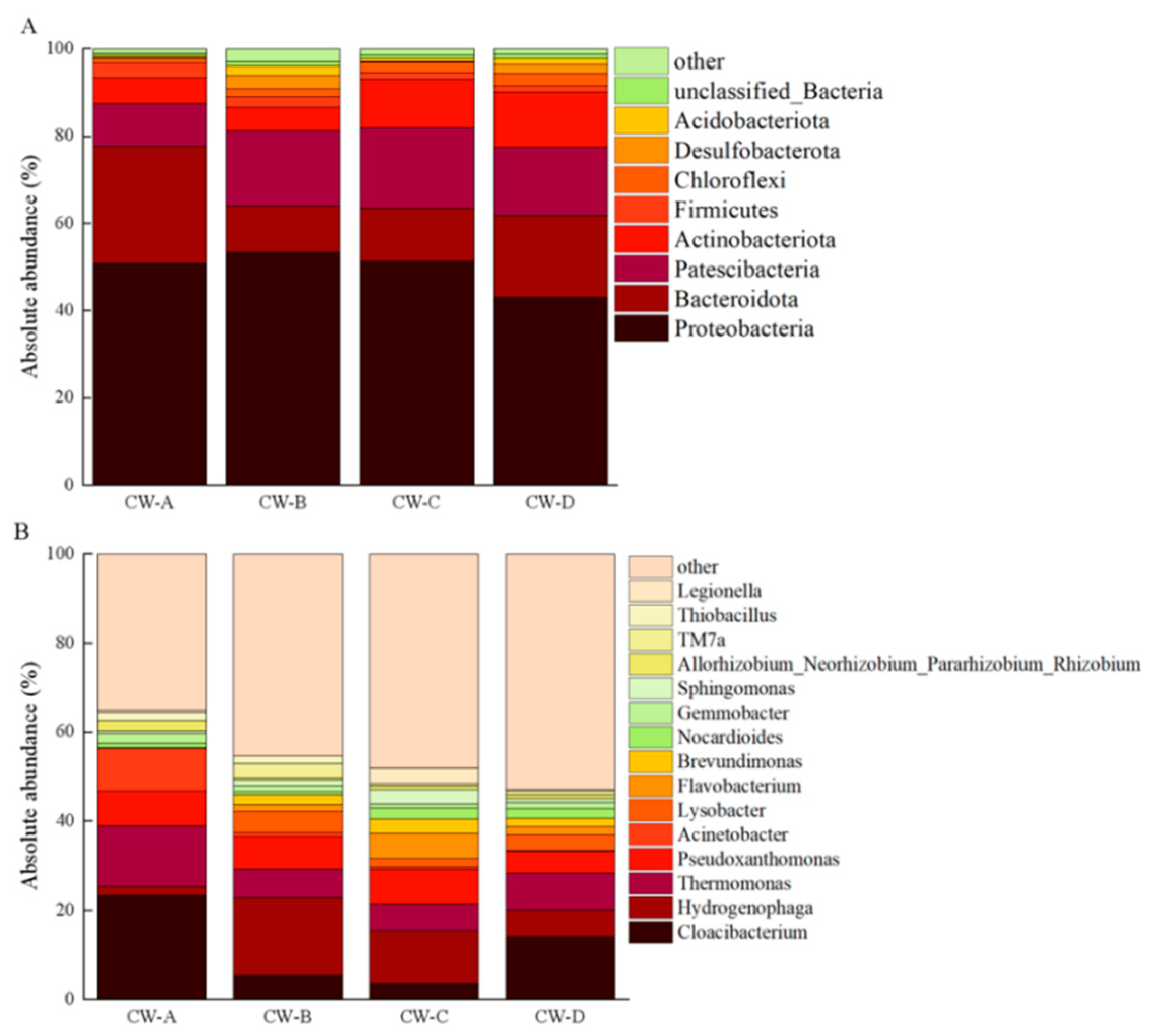
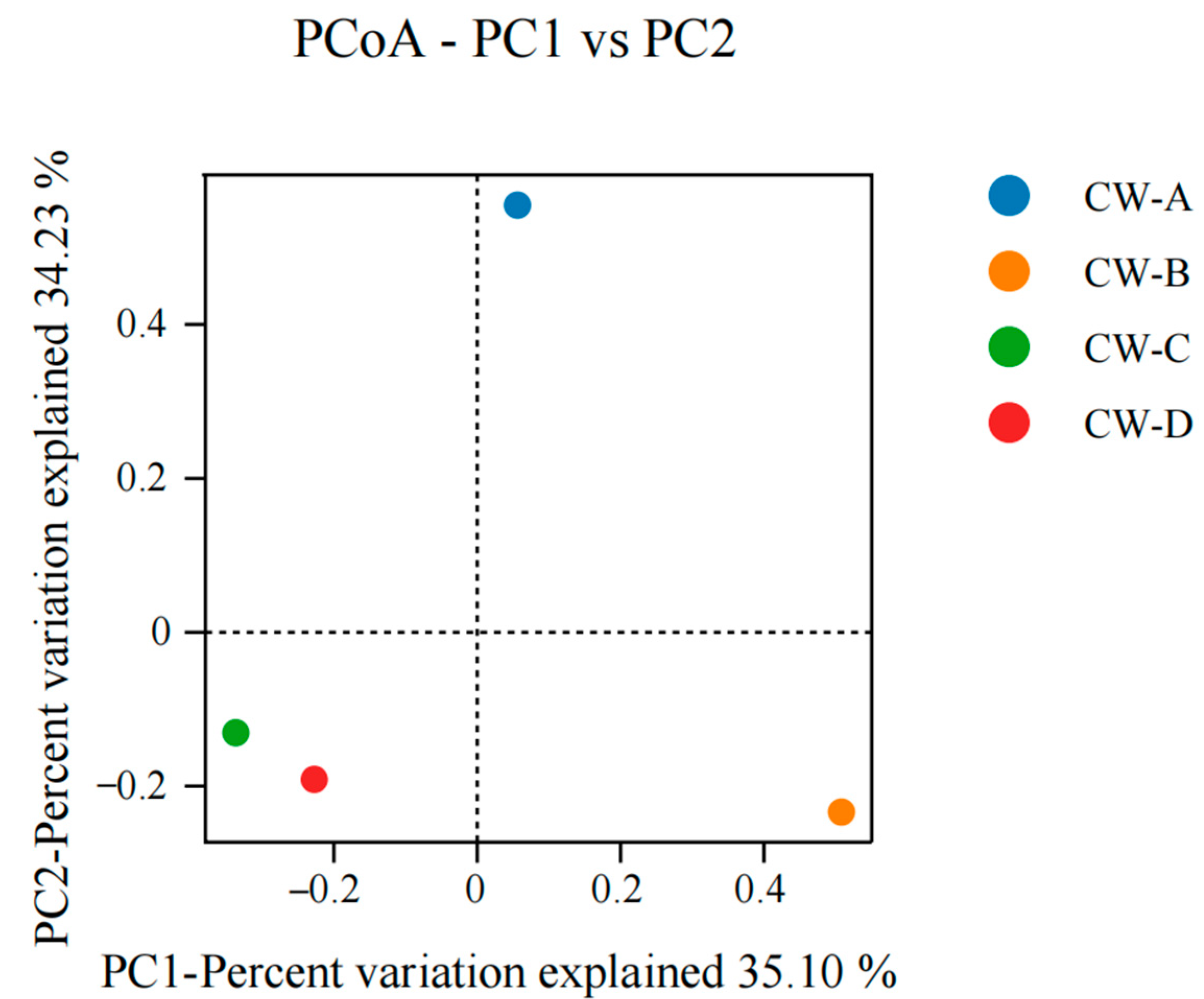
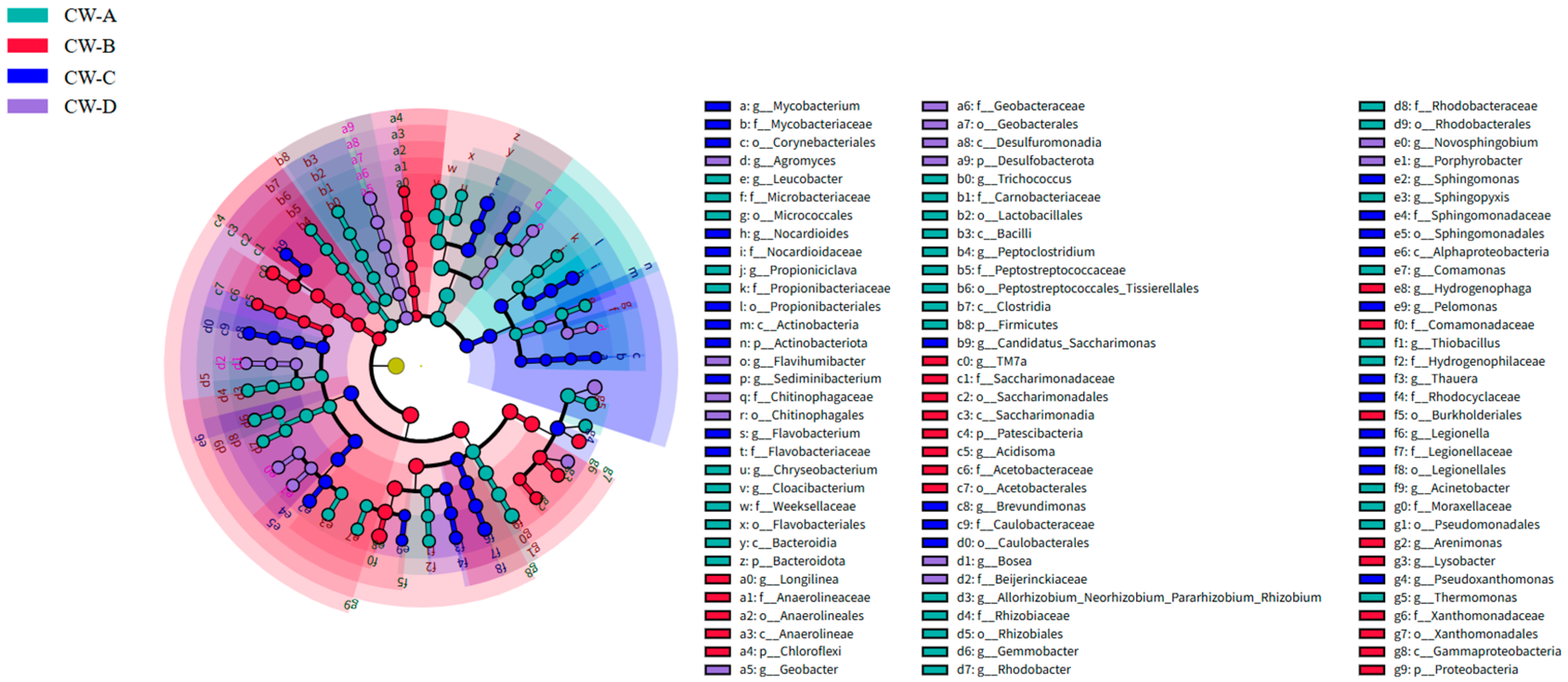
3.3. Analysis of Microbial Community Structure
3.4. Correlation Analysis Among ARGs, MGEs, and Microbial Community
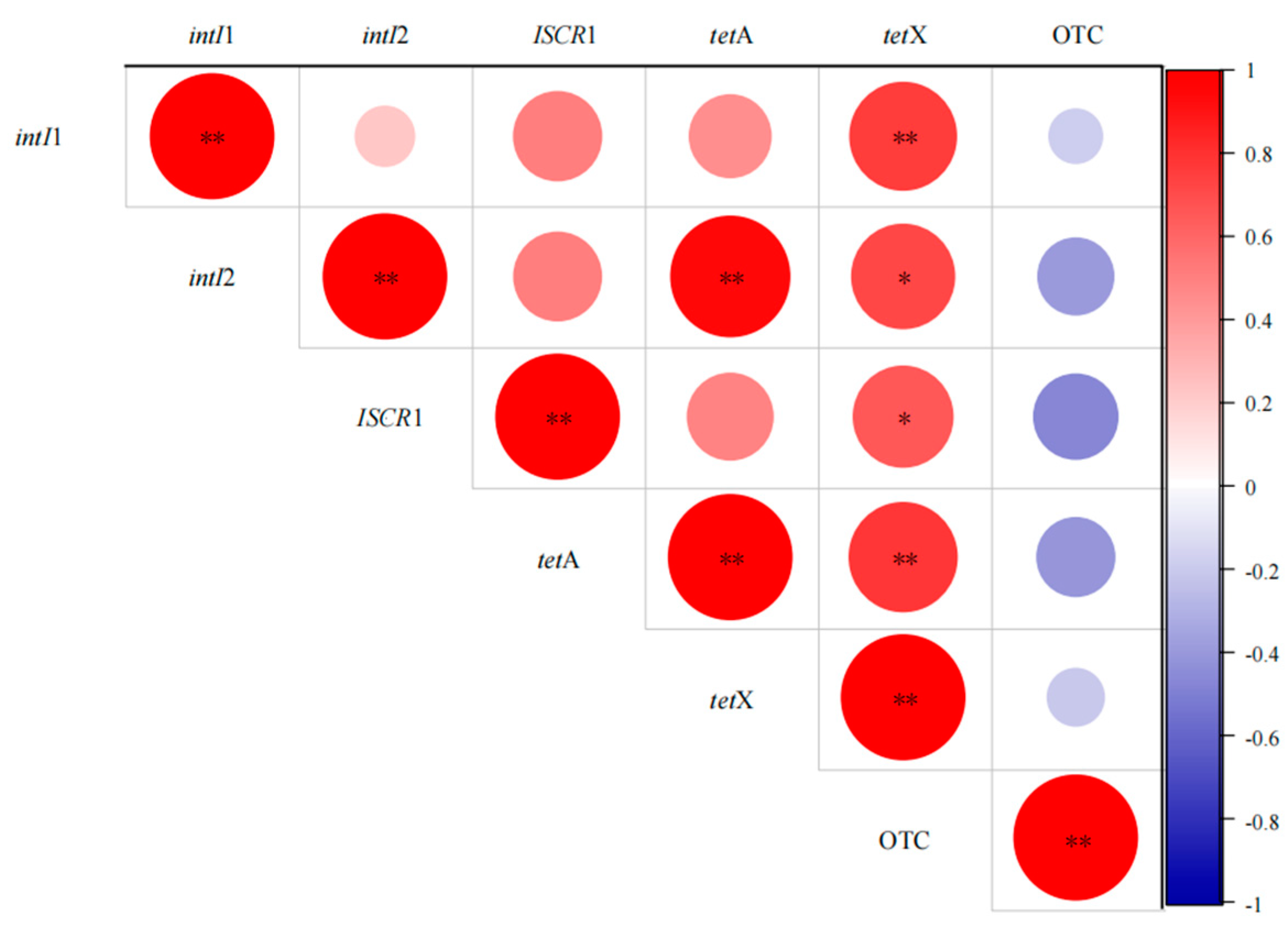
3.4.1. Correlation Analysis of MGEs, Antibiotics, and ARGs
3.4.2. Co-Occurrence Among ARGs, MGEs, and Potential Hosts
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yuan, W.; Zeng, X.; Cao, Y.; Yang, Q.; Riaz, L.; Wang, Q. Distribution of antibiotic resistance genes from human and animal origins to their receiving environments: A regional scale survey of urban settings. Environ. Pollut. 2022, 293, 118512. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.E.; Weatherly, L.; Shane, H.L. Contribution of antimicrobials to the development of allergic disease. Curr. Opin. Immunol. 2019, 60, 91–95. [Google Scholar] [CrossRef]
- Hu, Y.; Jin, L.; Zhao, Y.; Jiang, L.; Yao, S.; Zhou, W.; Lin, K.; Cui, C. Annual trends and health risks of antibiotics and antibiotic resistance genes in a drinking water source in East China. Sci. Total Environ. 2021, 791, 148152. [Google Scholar] [CrossRef]
- de la Fuente-Nunez, C.; Cesaro, A.; Hancock, R.E. Antibiotic failure: Beyond antimicrobial resistance. Drug Resist. Updat. 2023, 71, 101012. [Google Scholar] [CrossRef] [PubMed]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C. Antibiotic resistance genes in treated wastewater and in the receiving water bodies: A pan-European survey of urban settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef]
- Sabatino, R.; Cabello-Yeves, P.J.; Eckert, E.M.; Corno, G.; Callieri, C.; Brambilla, D.; Dzhembekova, N.; Moncheva, S.; Di Cesare, A. Antibiotic resistance genes correlate with metal resistances and accumulate in the deep water layers of the Black Sea. Environ. Pollut. 2022, 312, 120033. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Campos, L.C.; Canales, M.; Ciric, L. Drinking water biofiltration: Behaviour of antibiotic resistance genes and the association with bacterial community. Water Res. 2020, 182, 115954. [Google Scholar] [CrossRef]
- Guo, Z.-F.; Boeing, W.J.; Xu, Y.-Y.; Borgomeo, E.; Liu, D.; Zhu, Y.-G. Data-driven discoveries on widespread contamination of freshwater reservoirs by dominant antibiotic resistance genes. Water Res. 2023, 229, 119466. [Google Scholar] [CrossRef]
- Wu, D.-L.; Zhang, M.; He, L.-X.; Zou, H.-Y.; Liu, Y.-S.; Li, B.-B.; Yang, Y.-Y.; Liu, C.; He, L.-Y.; Ying, G.-G. Contamination profile of antibiotic resistance genes in ground water in comparison with surface water. Sci. Total Environ. 2020, 715, 136975. [Google Scholar] [CrossRef]
- Szekeres, E.; Chiriac, C.M.; Baricz, A.; Szőke-Nagy, T.; Lung, I.; Soran, M.-L.; Rudi, K.; Dragos, N.; Coman, C. Investigating antibiotics, antibiotic resistance genes, and microbial contaminants in groundwater in relation to the proximity of urban areas. Environ. Pollut. 2018, 236, 734–744. [Google Scholar] [CrossRef]
- Singh, A.K.; Kaur, R.; Verma, S.; Singh, S. Antimicrobials and antibiotic resistance genes in water bodies: Pollution, risk, and control. Front. Environ. Sci. 2022, 10, 830861. [Google Scholar] [CrossRef]
- Nnadozie, C.F.; Odume, O.N. Freshwater environments as reservoirs of antibiotic resistant bacteria and their role in the dissemination of antibiotic resistance genes. Environ. Pollut. 2019, 254, 113067. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.-Z.; Zou, H.-Y.; Wu, D.-L.; Chen, S.; He, L.-Y.; Zhang, M.; Bai, H.; Ying, G.-G. Swine farming elevated the proliferation of Acinetobacter with the prevalence of antibiotic resistance genes in the groundwater. Environ. Intern. 2020, 136, 105484. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Yang, Z.; Wei, Y.; Miao, J.; Yang, D.; Yin, J.; Li, H.; Chen, T.; Zhou, S.; Li, J. Spatial and temporal analysis of the seasonal dynamics of antibiotic resistance gene occurrence in recreational marine water. Sci. Total Environ. 2023, 893, 164816. [Google Scholar] [CrossRef]
- Kazanjian, P.H. Efforts to regulate antibiotic misuse in hospitals: A history. Infect. Control Hosp. Epidemiol. 2022, 43, 1119–1122. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, L.; Zhang, T.; Jin, L.; Han, Q.; Zhang, D.; Lin, K.; Cui, C. Occurrence and removal of sulfonamide antibiotics and antibiotic resistance genes in conventional and advanced drinking water treatment processes. J. Hazard. Mater. 2018, 360, 364–372. [Google Scholar] [CrossRef]
- Obayiuwana, A.; Ogunjobi, A.; Ibekwe, A. Prevalence of antibiotic resistance genes in pharmaceutical wastewaters. Water 2021, 13, 1731. [Google Scholar] [CrossRef]
- Lien, L.T.Q.; Lan, P.T.; Chuc, N.T.K.; Hoa, N.Q.; Nhung, P.H.; Thoa, N.T.M.; Diwan, V.; Tamhankar, A.J.; Stålsby Lundborg, C. Antibiotic resistance and antibiotic resistance genes in Escherichia coli isolates from hospital wastewater in Vietnam. Int. J. Environ. Res. Public Health 2017, 14, 699. [Google Scholar] [CrossRef]
- Hassoun-Kheir, N.; Stabholz, Y.; Kreft, J.-U.; De La Cruz, R.; Romalde, J.L.; Nesme, J.; Sørensen, S.J.; Smets, B.F.; Graham, D.; Paul, M. Comparison of antibiotic-resistant bacteria and antibiotic resistance genes abundance in hospital and community wastewater: A systematic review. Sci. Total Environ. 2020, 743, 140804. [Google Scholar] [CrossRef]
- Zhao, R.; Yu, K.; Zhang, J.; Zhang, G.; Huang, J.; Ma, L.; Deng, C.; Li, X.; Li, B. Deciphering the mobility and bacterial hosts of antibiotic resistance genes under antibiotic selection pressure by metagenomic assembly and binning approaches. Water Res. 2020, 186, 116318. [Google Scholar] [CrossRef]
- Zhang, Y.; Pei, M.; Zhang, B.; He, Y.; Zhong, Y. Changes of antibiotic resistance genes and bacterial communities in the advanced biological wastewater treatment system under low selective pressure of tetracycline. Water Res. 2021, 207, 117834. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wu, Y.; Zheng, H.; Li, H.; Zheng, Y.; Nan, J.; Ma, J.; Nagarajan, D.; Chang, J.-S. Antibiotics degradation by advanced oxidation process (AOPs): Recent advances in ecotoxicity and antibiotic-resistance genes induction of degradation products. Chemosphere 2023, 311, 136977. [Google Scholar] [CrossRef]
- Liu, L.; Xin, Y.; Huang, X.; Liu, C. Response of antibiotic resistance genes in constructed wetlands during treatment of livestock wastewater with different exogenous inducers: Antibiotic and antibiotic-resistant bacteria. Bioresour. Technol. 2020, 314, 123779. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chu, L.; Wojnárovits, L.; Takács, E. Occurrence and fate of antibiotics, antibiotic resistant genes (ARGs) and antibiotic resistant bacteria (ARB) in municipal wastewater treatment plant: An overview. Sci. Total Environ. 2020, 744, 140997. [Google Scholar] [CrossRef]
- Genç, N.; Dogan, E.C. Adsorption kinetics of the antibiotic ciprofloxacin on bentonite, activated carbon, zeolite, and pumice. Desalin. Water Treat. 2015, 53, 785–793. [Google Scholar] [CrossRef]
- Ping, Q.; Yan, T.; Wang, L.; Li, Y.; Lin, Y. Insight into using a novel ultraviolet/peracetic acid combination disinfection process to simultaneously remove antibiotics and antibiotic resistance genes in wastewater: Mechanism and comparison with conventional processes. Water Res. 2022, 210, 118019. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, L.; Gu, J.-D.; Deng, S.; Huang, C.; Luo, L. The factors controlling antibiotic resistance genes in different treatment processes of mainstream full-scale wastewater treatment plants. Sci. Total Environ. 2023, 900, 165815. [Google Scholar] [CrossRef]
- Chen, J.; Wei, X.D.; Liu, Y.S.; Ying, G.G.; Liu, S.S.; He, L.Y.; Su, H.C.; Hu, L.X.; Chen, F.R.; Yang, Y.Q. Removal of antibiotics and antibiotic resistance genes from domestic sewage by constructed wetlands: Optimization of wetland substrates and hydraulic loading. Sci. Total Environ. 2016, 565, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Cui, Y.; Li, A.; Zou, X.; Ma, C.; Chen, Z. Antibiotics and antibiotic resistance genes from wastewater treated in constructed wetlands. Ecol. Eng. 2022, 177, 106548. [Google Scholar] [CrossRef]
- Parde, D.; Patwa, A.; Shukla, A.; Vijay, R.; Killedar, D.J.; Kumar, R. A review of constructed wetland on type, treatment and technology of wastewater. Environ. Technol. Innov. 2021, 21, 101261. [Google Scholar] [CrossRef]
- Huang, X.; Zheng, J.; Liu, C.; Liu, L.; Liu, Y.; Fan, H. Removal of antibiotics and resistance genes from swine wastewater using vertical flow constructed wetlands: Effect of hydraulic flow direction and substrate type. Chem. Eng. J. 2017, 308, 692–699. [Google Scholar] [CrossRef]
- Song, H.-L.; Zhang, S.; Guo, J.; Yang, Y.-L.; Zhang, L.-M.; Li, H.; Yang, X.-L.; Liu, X. Vertical up-flow constructed wetlands exhibited efficient antibiotic removal but induced antibiotic resistance genes in effluent. Chemosphere 2018, 203, 434–441. [Google Scholar] [CrossRef]
- Yuan, W.; Zhang, Y.; Riaz, L.; Yang, Q.; Du, B.; Wang, R. Multiple antibiotic resistance and DNA methylation in Enterobacteriaceae isolates from different environments. J. Hazard. Mater. 2021, 402, 123822. [Google Scholar] [CrossRef]
- Riaz, L.; Wang, Q.; Yang, Q.; Li, X.; Yuan, W. Potential of industrial composting and anaerobic digestion for the removal of antibiotics, antibiotic resistance genes and heavy metals from chicken manure. Sci. Total Environ. 2020, 718, 137414. [Google Scholar] [CrossRef]
- Yuan, W.; Liu, Y.; Liu, R.; Li, L.; Deng, P.; Fu, S.; Riaz, L.; Lu, J.; Li, G.; Yang, Z. Unveiling the overlooked threat: Antibiotic resistance in groundwater near an abandoned sulfuric acid plant in Xingyang, China. Environ. Geochem. Health 2024, 46, 309. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, X.; Xiang, Y.; Wang, P.; Zhang, J.; Zhang, F.; Wei, J.; Luo, L.; Lei, M.; Tang, L. Modification of biochar derived from sawdust and its application in removal of tetracycline and copper from aqueous solution: Adsorption mechanism and modelling. Bioresour. Technol. 2017, 245, 266–273. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, W.; Ngo, H.H.; Wen, H.; Li, N.; Wu, W. Performance evaluation of powdered activated carbon for removing 28 types of antibiotics from water. Environ. Manag. 2016, 172, 193–200. [Google Scholar] [CrossRef]
- Belviso, C.; Guerra, G.; Abdolrahimi, M.; Peddis, D.; Maraschi, F.; Cavalcante, F.; Ferretti, M.; Martucci, A.; Sturini, M. Efficiency in Ofloxacin antibiotic water remediation by magnetic zeolites formed combining pure sources and wastes. Processes 2021, 9, 2137. [Google Scholar] [CrossRef]
- Karkman, A.; Do, T.T.; Walsh, F.; Virta, M.P. Antibiotic-resistance genes in waste water. Trends Microbiol. 2018, 26, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Zhao, Y.; Wang, C.; Zhang, H.; Chen, Q.; Zhang, X.; Zhang, L.; Wu, J.; Wu, Z.; Zhou, Q. Removal performance of antibiotics and antibiotic resistance genes in swine wastewater by integrated vertical-flow constructed wetlands with zeolite substrate. Sci. Total Environ. 2020, 721, 137765. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Canales, M.; Zhou, Q.; Karu, K.; Zhou, X.; Su, J.; Campos, L.C.; Ciric, L. Antibiotic resistance genes and the association with bacterial community in biofilms occurring during the drinking water granular activated carbon (GAC) sandwich biofiltration. J. Hazard. Mater. 2023, 460, 132511. [Google Scholar] [CrossRef]
- Mason-Jones, K.; Breidenbach, A.; Dyckmans, J.; Banfield, C.C.; Dippold, M.A. Intracellular carbon storage by microorganisms is an overlooked pathway of biomass growth. Nat. Commun. 2023, 14, 2240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Wu, X.; Gao, Y.; Fan, S.; Zhou, H.; Zhang, X. Diversity shifts in the root microbiome of cucumber under different plant cultivation substrates. Front. Microbiol. 2022, 13, 878409. [Google Scholar] [CrossRef]
- Arroyo, P.; Ansola, G.; de Miera, L.E.S. Effects of substrate, vegetation and flow on arsenic and zinc removal efficiency and microbial diversity in constructed wetlands. Ecol. Eng. 2013, 51, 95–103. [Google Scholar] [CrossRef]
- Abou-Kandil, A.; Shibli, A.; Azaizeh, H.; Wolff, D.; Wick, A.; Jadoun, J. Fate and removal of bacteria and antibiotic resistance genes in horizontal subsurface constructed wetlands: Effect of mixed vegetation and substrate type. Sci. Total Environ. 2021, 759, 144193. [Google Scholar] [CrossRef]
- Suyamud, B.; Lohwacharin, J.; Yang, Y.; Sharma, V.K. Antibiotic resistant bacteria and genes in shrimp aquaculture water: Identification and removal by ferrate (VI). J. Hazard. Mater. 2021, 420, 126572. [Google Scholar] [CrossRef]
- Mortezaei, Y.; Demirer, G.N.; Williams, M.R. Fate of intracellular and extracellular antibiotic resistance genes in sewage sludge by full-scale anaerobic digestion. Sci. Total Environ. 2024, 951, 175760. [Google Scholar] [CrossRef]
- Wang, Y.; Chu, L.; Ma, J.; Chi, G.; Lu, C.; Chen, X. Effects of multiple antibiotics residues in broiler manure on composting process. Sci. Total Environ. 2022, 817, 152808. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Z.; Xu, H.; Wang, L.; Liu, R.; Jia, X. Fate of antibiotic resistance genes and bacteria in a coupled water-processing system with wastewater treatment plants and constructed wetlands in coastal eco-industrial parks. Ecotoxicol. Environ. Safet. 2023, 252, 114606. [Google Scholar] [CrossRef] [PubMed]
- De Mandal, S.; Mathipi, V.; Muthukumaran, R.B.; Gurusubramanian, G.; Lalnunmawii, E.; Kumar, N.S. Amplicon sequencing and imputed metagenomic analysis of waste soil and sediment microbiome reveals unique bacterial communities and their functional attributes. Environ. Monit. Assess. 2019, 191, 778. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Wu, X. Microbial degradation of tetracycline in the aquatic environment: A review. Crit. Rev. Biotechnol. 2020, 40, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, Z.; Wu, B.; Xie, H.; Liu, G. Antibiotics and antibiotic resistance genes removal in biological aerated filter. Bioresour. Technol. 2024, 395, 130392. [Google Scholar] [CrossRef]
- Guo, N.; Wang, T.; Jin, Y.; Wu, D.; Chen, F.; Cheng, X.; Wang, J.; Feng, L.; Song, H.; Wang, L. Enhanced antibiotic removal in a nitrifying sludge system by ammonia-oxidizing bacteria and heterotrophs. J. Environ. Chem. Eng. 2022, 10, 108585. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, T.; Liu, X.; Zhan, M.; Song, X.; Xu, Y.; Yu, R. Sludge biolysis pretreatment to reduce antibiotic resistance genes (ARGs): Insight into the relationship between potential ARGs hosts and BALOs’ preferred prey. Water Res. 2024, 260, 121949. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Liu, H.; Wang, Y.; Wang, Q.; Zhang, L.; An, F.; Chen, Y. Impacts of cefalexin on nitrite accumulation, antibiotic degradation, and microbial community structure in nitrification systems. J. Hazard. Mater. 2024, 478, 135430. [Google Scholar] [CrossRef]
- Du, W.; Wang, T.; Wang, F.; Li, Z.; Huang, W.; Tai, J.; Fang, S.; Cheng, X.; Cao, J.; Su, Y. Para-chloro-meta-xylenol reshaped the fates of antibiotic resistance genes during sludge fermentation: Insights of cell membrane permeability, bacterial structure and biological pathways. Sci. Total Environ. 2022, 850, 158083. [Google Scholar] [CrossRef]
- Yu, Z.; Liu, Z.; Sun, L.; Dong, C.; Jin, Y.; Hu, B.; Cheng, D. Mobile genetic elements mediate the cross-media transmission of antibiotic resistance genes from pig farms and their risks. Sci. Total Environ. 2024, 926, 172115. [Google Scholar] [CrossRef]
- Jang, H.M.; Lee, J.; Choi, S.; Shin, J.; Kan, E.; Kim, Y.M. Response of antibiotic and heavy metal resistance genes to two different temperature sequences in anaerobic digestion of waste activated sludge. Bioresour. Technol. 2018, 267, 303–310. [Google Scholar] [CrossRef]
- Jiang, L.; Zhai, W.; Wang, J.; Li, G.; Zhou, Z.; Li, B.; Zhuo, H. Antibiotics and antibiotic resistance genes in the water sources of the Wuhan stretch of the Yangtze River: Occurrence, distribution, and ecological risks. Environ. Res. 2023, 239, 117295. [Google Scholar] [CrossRef]
- Newman, M.E. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef]
- Michaelis, C.; Grohmann, E. Horizontal gene transfer of antibiotic resistance genes in biofilms. Antibiotics 2023, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Zhou, G.; Wang, H. Nanoscale zero-valent iron inhibits the horizontal gene transfer of antibiotic resistance genes in chicken manure compost. J. Hazard. Mater. 2022, 422, 126883. [Google Scholar] [CrossRef]
- Zhang, Z.-X.; Fan, X.-Y.; Li, X.; Gao, Y.-X.; Zhao, J.-R. Effects of combined antibiotics on nitrification, bacteria and antibiotic resistance genes in activated sludge: Insights from legacy effect of antibiotics. J. Environ. Sci. 2023, 131, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, Y.; Li, W.-X.; Qu, J.-H.; Chen, T.; Wang, Y.-P.; Geng, N.-Y. Deciphering the microbial community tolerance mechanism and alteration of antibiotic resistance genes during chloramphenicol wastewater treatment. Int. Biodeterior. Biodegrad. 2023, 178, 105546. [Google Scholar] [CrossRef]
- Stallworth, C.; Steed, L.; Fisher, M.A.; Nolte, F.S. Legionnaires’ disease caused by Legionella londiniensis. J. Clin. Microbiol. 2012, 50, 4178–4179. [Google Scholar] [CrossRef]
- Dowling, J.; Pasculle, A. Bactericidal activity of antibiotics against Legionella micdadei (Pittsburgh pneumonia agent). Antimicrob. Agents Chemother. 1982, 22, 272–276. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Feature | ACE | Chao1 | Simpson | Shannon |
|---|---|---|---|---|---|
| CW-A | 849 | 867.9716 | 853.5922 | 0.9353 | 5.8005 |
| CW-B | 1459 | 1478.7525 | 1462.5455 | 0.9575 | 6.8134 |
| CW-C | 1318 | 1333.5655 | 1320.0413 | 0.9711 | 6.9839 |
| CW-D | 1079 | 1092.5013 | 1081.2857 | 0.9686 | 7.0705 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, W.; Liu, Y.; Shang, Y.; Bai, M.; Li, L.; Li, X.; Deng, P.; Riaz, L.; Guo, Y.; Lu, J. Fate and Removal of Oxytetracycline and Antibiotic Resistance Genes in Vertical-Flow Constructed Wetland with Different Substrates. Water 2025, 17, 1412. https://doi.org/10.3390/w17101412
Yuan W, Liu Y, Shang Y, Bai M, Li L, Li X, Deng P, Riaz L, Guo Y, Lu J. Fate and Removal of Oxytetracycline and Antibiotic Resistance Genes in Vertical-Flow Constructed Wetland with Different Substrates. Water. 2025; 17(10):1412. https://doi.org/10.3390/w17101412
Chicago/Turabian StyleYuan, Wei, Yan Liu, Yijun Shang, Meng Bai, Leicheng Li, Xunan Li, Peiyuan Deng, Luqman Riaz, Yiping Guo, and Jianhong Lu. 2025. "Fate and Removal of Oxytetracycline and Antibiotic Resistance Genes in Vertical-Flow Constructed Wetland with Different Substrates" Water 17, no. 10: 1412. https://doi.org/10.3390/w17101412
APA StyleYuan, W., Liu, Y., Shang, Y., Bai, M., Li, L., Li, X., Deng, P., Riaz, L., Guo, Y., & Lu, J. (2025). Fate and Removal of Oxytetracycline and Antibiotic Resistance Genes in Vertical-Flow Constructed Wetland with Different Substrates. Water, 17(10), 1412. https://doi.org/10.3390/w17101412