
Application of Modern Approaches to Species Delimitation in Freshwater Erpobdellidae (Hirudinea: Arhynchobdellida) with a New Interpretation of Their Genera


Abstract
1. Introduction
2. Materials and Methods
2.1. New Sample Collection and Morphological Analysis
2.2. DNA Extraction, Amplification, and Sequencing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Site | Coordinates | Voucher | Accession Number |
|---|---|---|---|
| Erpobdella octoculata | |||
| LB: Lake Izumrudnoe | 51°23′39″N 104°38′33″E | E275, E276 | MN245559, MN245560 |
| DRb: Lopan River* | 49°59′19″N, 36°13′38″E | KI37, KI38 | MN245561, MN245562 |
| Erpobdella sibirica sp. nov. | |||
| ARb: Angara River | 53°04′31″N, 104°19′05″E | KI34 | MN245518 |
| ARb: Angara River | 52°14′59″N, 104°17′04″E | KI36 | MN245519 |
| ARb: Kudarejka River | 52°51′53″N, 104°27′05″E | KI10 | MN245532 |
| ARb: B Edogon River | 54°12′03″N, 100°05′25″E | E183, E185, E186 | MN245520-MN245522 |
| ARb: Oyok pond | 52°36′20″N, 104°29′02″E | E248, E251 | MN245548, MN245549 |
| LRb: Lake Dalnee | 56°05′28″N, 108°16′07″E | E180, E181 | MN245527, MN245528 |
| LRb: Tukolon River | 55°24′44″N, 107°03′68″E | E176 | MN245554 |
| LB: Shida Bay | 53°03′55″N, 106°47′33″E | KI19, E19 | MN245542, KM220015 |
| LB: Ulirba Bay | 53°04′08″N, 106°48′25″E | KI1, E1, E7, E8 | MN245543, KM220016-KM220018 |
| LB: Khuzhir-Nugo Bay | 53°04′04″N, 106°48′52″E | KI9, E9 | MN245544, KM220019 |
| LB: Kurkut Bay | 53°01′12″N, 106°53′49″E | F1, E101, E17 | MN245539, MN245541, KM220020 |
| LB: Tulkhane Bay | 53°02′48″N, 106°46′36″E | E102 | MN245540 |
| LB: Lake Kurma | 53°11′13″N, 106°58′58″E | E112, E114 | MN245533, MN245534 |
| LB: Lake Kurma | 53°10′57″N, 106°58′41″E | E116 | MN245535 |
| LB: Lake Surhaitornur | 53°16′53″N, 107°09′19″E | E137 | MN245553 |
| LB: Chivyrkuy Bay | 53°46′10″N, 109°00′28″E | KI17 | MN245526 |
| LB: Chivyrkuy Bay | 53°46′05″N, 109°01′22″E | KI2, KI3 | MN245525, MN245523 |
| LB: Chivyrkuy Bay | 53°45′58″N, 109°02′13″E | KI33 | MN245524 |
| LRb: Lena River | 58°07′13″N, 108°46′05″E | KI4, KI5 | MN245536, MN245537 |
| LRb: Lena River | 57°46′26″N, 108°07′43″E | KI6 | MN245538 |
| LRb: Lake Severnoe | 54°19′22″N, 108°22′43″E | 925 | MN245550 |
| LRb: Lake Severnoe | 54°19′15″N, 108°22′40″E | 926 | MN245551 |
| LRb: Lake Severnoe | 54°19′18″N, 108°22′45″E | 927 | MN245552 |
| SRb: Lake Gusinoe | 51°15′57″N, 106°23′54″E | G60 | MN245529 |
| SRb: Lake Gusinoe | 51°17′31″N, 106°28′42″E | G81 | MN245530 |
| SRb: Lake Gusinoe | 51°17′28″N, 106°28′47″E | G82 | MN245531 |
| SRb: Lake Ulaagchny | 48°22′28″N, 96°10′40″E | 915, 916, 917 | MN245545-MN245547 |
| SRb: Lake Ulaagchny | 48°22′37″N, 96°08′56″E | 918,919, 922,923 | MN245555- MN245558 |
2.3. Bioinformatic Analysis of Molecular Data
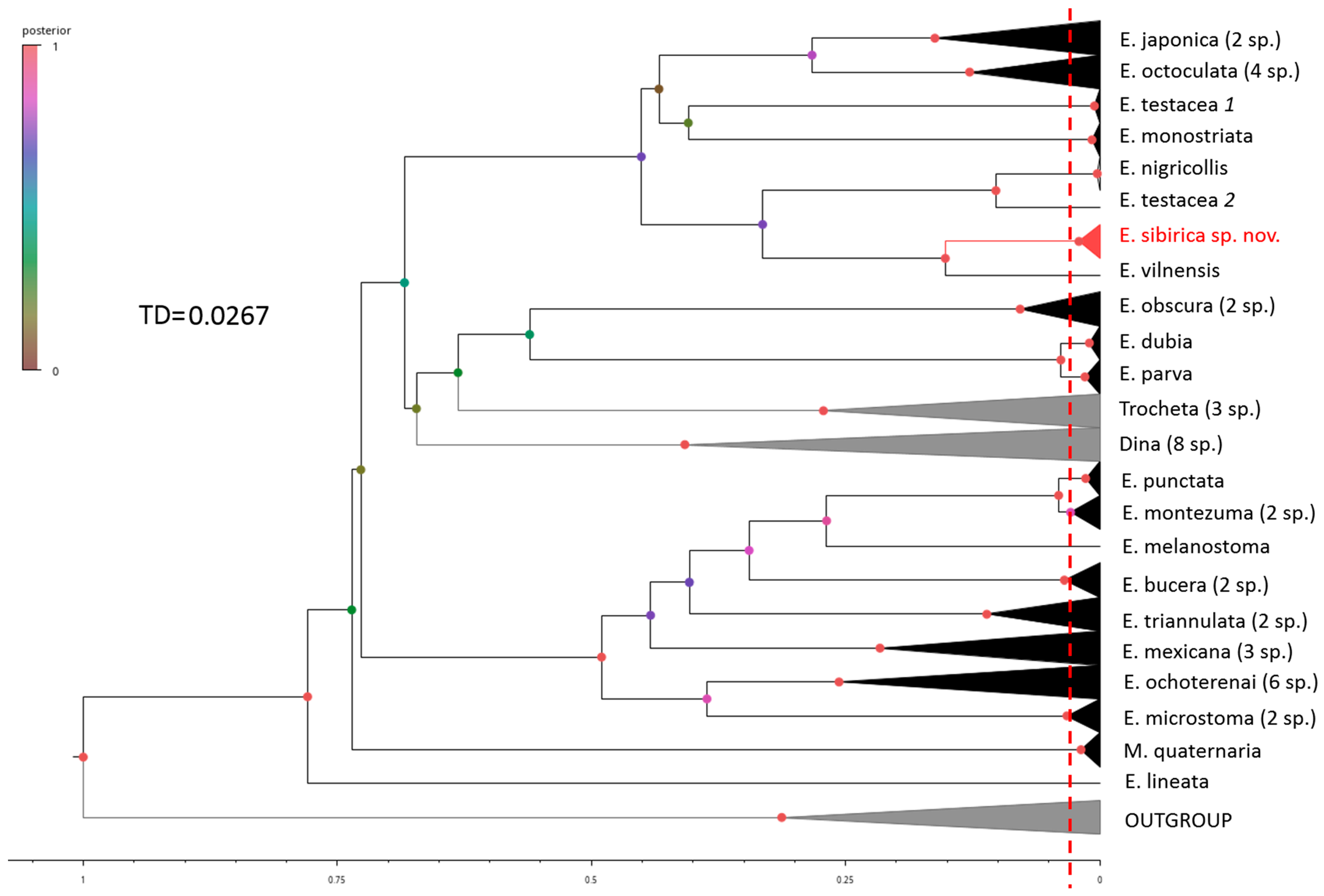
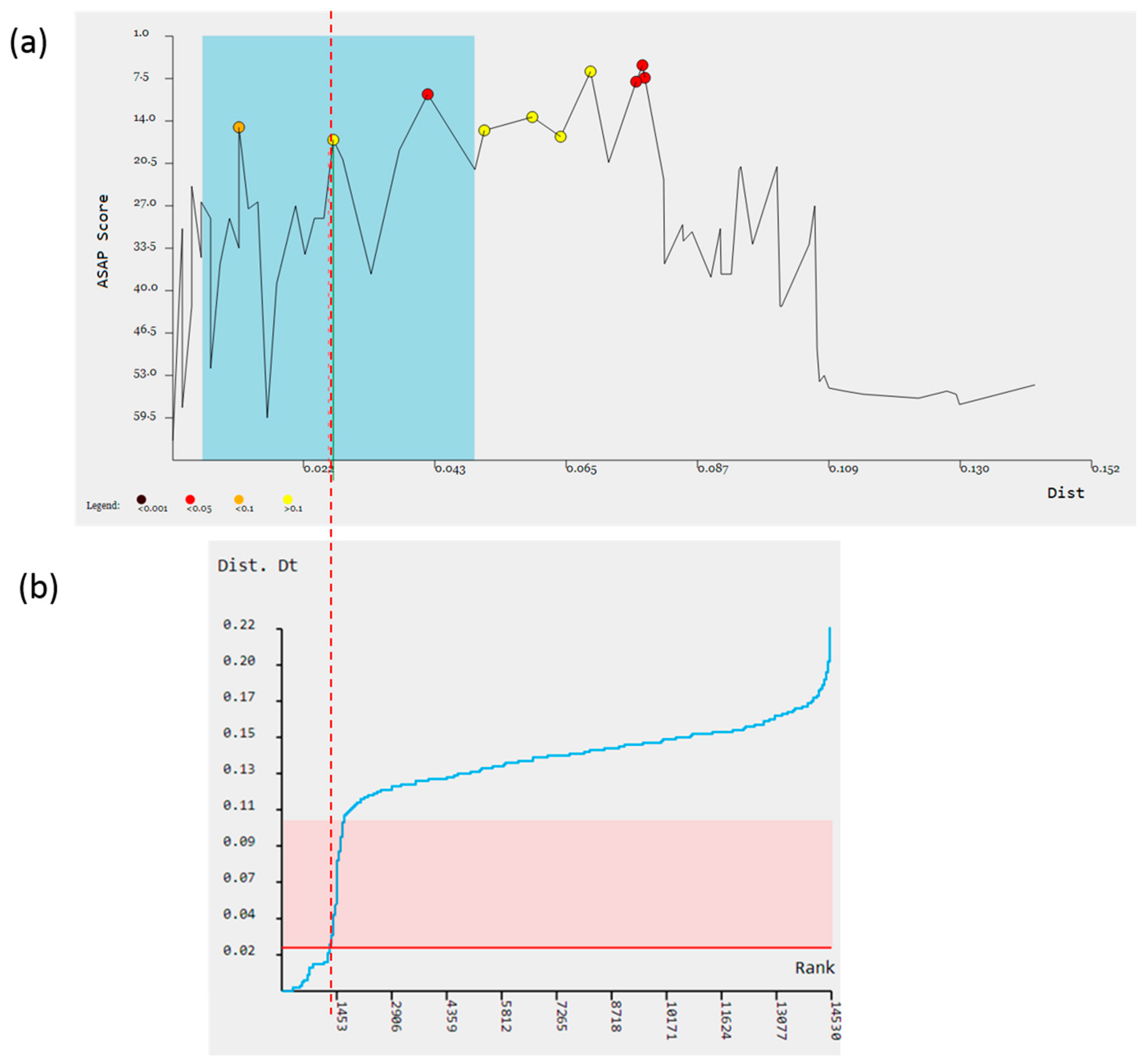
3. Results
4. Discussion
| Region | Morphospecies | Pi within MS | Number of CS | Pi between CS (Average) |
|---|---|---|---|---|
| PA | Dina lineata * | 0.0167 | 1(2) | 0.0196 |
| Erpobdella japonica | 0.0678 | 2 | 0.0973 | |
| Erpobdella octoculata | 0.0555 | 4 | 0.0559–0.0890 (avg. 0.0808) | |
| NA | Erpobdella bucera | 0.0331 | 2 | 0.0331 |
| Erpobdella mexicana | 0.0214 | 3 | 0.0431–0.1149 (avg. 0.0872) | |
| Erpobdella obscura | 0.0188 | 2 | 0.0554 | |
| Erpobdella ochoterenai | 0.0626 | 6 | 0.0612–0.1204 (avg. 0.0972) | |
| Erpobdella triannulata | 0.0492 | 2 | 0.0658 | |
| Erpobdella microstoma * | 0.0206 | 1(2) | 0.0315 | |
| Motobdella Montezuma * | 0.0133 | 1(2) | 0.0265 | |
| Erpobdella dubia/parva ** | 0.0199 | 1(2) | 0.0308 |
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Molecular Description of Erpobdella sibirica sp. nov. (Figure A1)


References
- Valentini, A.; Pompanon, F.; Taberlet, P. DNA barcoding for ecologists. Trends Ecol. Evol. 2009, 24, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Mora, C.; Tittensor, D.; Adl, S.; Simpson, A.; Worm, B. How many species are there on Earth and in the ocean? PLoS Biol. 2011, 9, e1001127. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.; Barnosky, A.; García, A.; Pringle, R.; Todd, P. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.; Cywinska, A.; Ball, S.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. Royal Soc. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.; Stoeckle, M.; Zemlak, T.; Francis, C. Identification of Birds through DNA Barcodes. PLoS Biol. 2004, 2, e312. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.; Hollingsworth, P.; Hajibabaei, M. From writing to reading the encyclopedia of life. Phil. Trans. R. Soc. B 2016, 371, 20150321. [Google Scholar] [CrossRef] [PubMed]
- Wiemers, M.; Fiedler, K. Does the DNA barcoding gap exist?—A case study in blue butterflies (Lepidoptera: Lycaenidae). Front. Zool. 2007, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Freudenstein, J.; Broe, M.; Folk, R.; Sinn, B. Biodiversity and the Species Concept—Lineages are not Enough. Syst. Biol. 2017, 66, 644–656. [Google Scholar] [CrossRef]
- Estensmo, E.; Maurice, S.; Morgado, L.; Martin-Sanchez, P.; Skrede, I.; Kauserud, H. The influence of intraspecific sequence variation during DNA metabarcoding: A case study of eleven fungal species. Mol. Ecol. Resour. 2021, 21, 1141–1148. [Google Scholar] [CrossRef]
- Hebert, P.; Gregory, T. The promise of DNA Barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef]
- Burns, J.; Janzen, D.; Hajibabaei, M.; Hallwachs, W.; Hebert, P. DNA barcodes and cryptic species of skipper butterflies in the genus Perichares in Area de Conservación Guanacaste, Costa Rica. Proc. Natl. Acad. Sci. USA 2008, 105, 6350–6355. [Google Scholar] [CrossRef] [PubMed]
- Bely, A.; Weisblat, D. Lessons from leeches: A call for DNA barcoding in the lab. Evol. Dev. 2006, 8, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Koperski, P.; Milanowski, R.; Krzyk, A. Searching for cryptic species in Erpobdella ocloculata (L.) (Hirudinea: Clitellata): Discordance between the results of genetic analysis and cross-breeding experiments. Contrib. Zool. 2011, 80, 85–94. [Google Scholar] [CrossRef]
- Oceguera-Figueroa, A.; Phillips, A.; Pacheco-Chaves, B.; Reeves, W.; Siddall, M. Phylogeny of macrophagous leeches (Hirudinea, Clitellata) based on molecular data and evaluation of the barcoding locus. Zool. Scr. 2011, 40, 194–203. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Mandzyak, N. Molecular phylogeny of Siberian Glossiphoniidae (Hirudinea). Mol. Biol. 2014, 48, 452–455. [Google Scholar] [CrossRef]
- Utevsky, S.; Dubov, P.; Prokin, A. First Russian record of Erpobdella monostriata: DNA barcoding and geographical distribution. Spixiana 2015, 38, 161–168. [Google Scholar]
- de Waard, J.; Ratnasingham, S.; Zakharov, E.; Borisenko, A.; Steinke, D.; Tefler, A.; Perez, A.; Sones, J.; Young, M.; Levesque-Beaudin, V.; et al. A reference library for Canadian invertebrates with 1.5 million barcodes, voucher specimens, and DNA samples. Sci. Data 2019, 6, 308. [Google Scholar] [CrossRef] [PubMed]
- Kaygorodova, I.; Bolbat, N.; Bolbat, A. Species delimitation through DNA barcoding of freshwater leeches of the Glossiphonia genus (Hirudinea: Glossiphoniidae) from Eastern Siberia, Russia. J. Zool. Syst. Evol. 2020, 58, 1437–1446. [Google Scholar] [CrossRef]
- Jovanović, M.; Haring, E.; Sattmann, H.; Grosser, C.; Pesic, V. DNA barcoding for species delimitation of the freshwater leech genus Glossiphonia from the Western Balkan (Hirudinea, Glossiphoniidae). Biodivers. Data J. 2021, 9, e66347. [Google Scholar] [CrossRef]
- Kondratov, I.; Sitnikova, T.; Kaygorodova, I.; Denikina, N.; Annenkov, V.; Khanaev, I.; Kirilchik, S.; Nebesnykh, I.; Dzyuba, E. Amazing discoveries of benthic fauna from the abyssal zone of Lake Baikal. Biology 2021, 10, 972. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Matveenko, E.; Dzyuba, E. Unexpected discovery of an ectoparasitic invasion first detected in the Baikal coregonid fish population. Fishes 2022, 7, 298. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Matveenko, E. Diversity of the Piscicola species (Hirudinea, Piscicolidae) in the Eastern Palaearctic with a description of three new species and notes on their biogeography. Diversity 2023, 15, 98. [Google Scholar] [CrossRef]
- Trontelj, P.; Sket, B.; Dovč, P.; Steinbrück, G. Phylogenetic relationships in European erpobdellid leeches (Hirudinea: Erpobdellidae) inferred from restriction-site data of the 18S ribosomal gene and ITS2 region. J. Zool. Syst. Evol. Res. 1996, 34, 85–93. [Google Scholar] [CrossRef]
- Govedich, F.; Blinn, D.; Keim, P.; Davies, R. Phylogenetic relationships of three genera of Erpobdellidae (Hirudinoidea), with description of a new genus, Motobdella, and species Motobdella sedonensis. Can. J. Zool. 1998, 76, 2164–2171. [Google Scholar] [CrossRef]
- Trontelj, P.; Sket, B. Molecular re-assessment of some phylogenetic, taxonomic and biogeographic relationships between the leech genera Dina and Trocheta (Hirudinea: Erpobdellidae). Hydrobiologia 2000, 438, 227–235. [Google Scholar] [CrossRef]
- Siddall, M. Phylogeny of the leech family Erpobdellidae (Hirudinida: Oligochaeta). Invertebr. Syst. 2002, 16, 1–6. [Google Scholar] [CrossRef]
- Pffeiffer, I.; Brenig, B.; Kutschera, U. Molecular phylogeny of selected predaceous leeches with reference to the evolution of body size and terrestrialism. Theory Biosci. 2005, 124, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Sket, B. Intralacustrine speciation in the genus Dina (Hirudinea, Erpobdellidae) in Lake Ohrid (Yugoslavia). Hydrobiologia 1989, 182, 49–59. [Google Scholar] [CrossRef]
- Lukin, E. Leeches of Fresh and Saline Waters; Nauka: Leningrad, Russia, 1976; 484p. [Google Scholar]
- Nesemann, H.; Neubert, E. Annelida, Clitellata: Branchiobdellida, Acanthobdellea, Hirudinea, Susswasserfauna von Mitteleuropa; Spectrum Akademischer Verlag: Heidelberg, Germany, 1999; 178p. [Google Scholar]
- Hovingh, P. Erpobdella (Dina) parva complex (Annelida: Hirudinea: Arhynchobdellida: Erpobdellidae): Additional description of Erpobdella parva, E. dubia, and E. lahontana and taxonomic revision. Hydrobiologia 2004, 517, 89–105. [Google Scholar] [CrossRef]
- Trajanovski, S.; Albrecht, C.; Schreiber, K.; Schultheiß, R.; Stadler, T.; Benke, M.; Wilke, T. Testing the spatial and temporal framework of speciation in an ancient lake species flock: The leech genus Dina (Hirudinea: Erpobdellidae) in Lake Ohrid. Biogeosciences 2010, 7, 3387–3402. [Google Scholar] [CrossRef]
- Anderson, K.; Braoudakis, G.; Kvist, S. Genetic variation, pseudocryptic diversity, and phylogeny of Erpobdella (Annelida: Hirudinida: Erpobdelliformes), with emphasis on Canadian species. Mol. Phylogenet. Evol. 2020, 143, 106688. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.; Siddall, M. Poly-paraphyly of Hirudinidae: Many lineages of medicinal leeches. BMC Evol. Biol. 2009, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Khomenko, A.; Utevsky, S.; Utevsky, A.; Trontelj, P. Unrecognized diversity of Trocheta species (Hirudinea: Erpobdellidae): Resolving a century-old taxonomic problem in Crimean leeches. Syst. Biodivers. 2020, 18, 129–141. [Google Scholar] [CrossRef]
- Pešić, V.; Grosser, C. Dina serbica, a new species of leeches (Annelida: Hirudinea: Erpobdellidae) from Serbia, based on morphological and molecular evidence. Ecol. Montenegrina 2022, 51, 1–14. [Google Scholar] [CrossRef]
- Grosser, C.; Barjadze, S.; Maghradze, E. Trocheta ariescornuta n. sp. (Annelida, Hirudinida: Erpobdellidae)—A new cavernicolous leech from Motena Cave in Georgia. Ecol. Montenegrina 2021, 44, 32–43. [Google Scholar] [CrossRef]
- Cichocka, J.; Bielecki, A.; Kur, J.; Pikuła, D.; Kilikowska, A.; Biernacka, B. A new leech species (Hirudinida: Erpobdellidae: Erpobdella) from a cave in the West Azerbaijan province of Iran. Zootaxa 2015, 4013, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Liskiewicz, S. Leeches (Hirudinea) of the Kazan province (Russia, Eastern). Proc. Soc. Friends Sci. Vilnius 1925, 2, 2–8. [Google Scholar]
- Utevsky, S.; Son, M.; Dyadichko, V.; Kaygorodova, I. New information on the geographical distribution of Erpobdella vilnensis (Liskiewicz, 1925) (Hirudinida, Erpobdellidae) in Ukraine. Lauterbornia 2012, 75, 75–78. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Camacho, C.; Boratyn, G.; Joukov, V.; Vera Alvarez, R.; Madden, T. ElasticBLAST: Accelerating sequence search via cloud computing. BMC Bioinform. 2023, 24, 117. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, S.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Barraclough, T. Delimiting species using single-locus data and the Generalized Mixed Yule Coalescent approach: A revised method and evaluation on simulated data sets. Syst Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Bolbat, A.; Kaygorodova, I.; Bukin, Y.; Fedorova, L.; Sorokovikova, N. Application of bioinformational methods for species delimitation in the genus Erpobdella (Erpobdellidae, Hirudinea). Bull. Irkutsk State Univ. Ser. Biol. Ecol. 2017, 20, 3–13. [Google Scholar]
- Hebert, P.; Ratnasingham, S.; de Waard, J. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B 2003, 270, S96–S99. [Google Scholar] [CrossRef]
- Kekkonen, M.; Hebert, P. DNA barcode-based delineation of putative species: Efficient start for taxonomic workflows. Mol. Ecol. Resour. 2014, 14, 706–715. [Google Scholar] [CrossRef]
- Bugarski-Stanojević, V.; Stamenković, G.; Jojić, V.; Ćosić, N.; Ćirović, D.; Stojković, O.; Veličković, J.; Savić, I. Cryptic diversity of the european blind mole rat Nannospalax leucodon species complex: Implications for Conservation. Animals 2022, 12, 1097. [Google Scholar] [CrossRef]
- Cerca, J.; Meyer, C.; Purschke, G.; Struck, T. Delimitation of cryptic species drastically reduces the geographical ranges of marine interstitial ghost-worms (Stygocapitella; Annelida, Sedentaria). Mol. Phylogenet. Evol. 2020, 143, 106663. [Google Scholar] [CrossRef] [PubMed]
- Jörger, K.; Norenburg, J.; Wilson, N.; Schrödl, M. Barcoding against a paradox? Combined molecular species delineations reveal multiple cryptic lineages in elusive meiofaunal sea slugs. BMC Evol. Biol. 2012, 12, 245. [Google Scholar] [CrossRef] [PubMed]
- Novo, M.; Almodovar, A.; Fernandez, R.; Cosin, D. Cryptic speciation of hormogastrid earthworms revealed by mitochondrial and nuclear data. Mol. Phylogenet. Evol. 2010, 56, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Kon, T.; Yoshino, T.; Mukai, T.; Nishida, M. DNA sequences identify numerous cryptic species of the vertebrate: A lesson from the gobioid fish Schindleria. Mol. Phylogen. Evol. 2007, 44, 53–62. [Google Scholar] [CrossRef]
- Trontelj, P.; Fišer, C. Perspectives: Cryptic species diversity should not be trivialized. Syst. Biodivers. 2009, 7, 1–3. [Google Scholar] [CrossRef]
- Poulin, R.; Pérez-Ponce de Leόn, G. Global analysis reveals that cryptic diversity is linked with habitat but not mode of life. J. Evol. Biol. 2017, 30, 641–649. [Google Scholar] [CrossRef]
- Pante, E.; Puillandre, N.; Viricel, A.; Arnaud-Haond, S.; Aurelle, D.; Castelin, M.; Chenuil, A.; Destombe, C.; Forcioli, D.; Valero, M.; et al. Species are hypotheses: Avoid connectivity assessments based on pillars of sand. Mol. Ecol. 2015, 24, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Tessler, M.; Siddall, M.; Oceguera-Figueroa, A. Leeches from Chiapas, Mexico with a new species of Erpobdella (Hirudinida: Erpobdellidae). Am. Mus. Novit. 2018, 3895, 129–134. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Pronin, N. New records of Lake Baikal Leech Fauna: Species Diversity and Spatial Distribution in Chivyrkuy Gulf. Sci. World J. 2013, 2013, 206590. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Mandzyak, N.; Petryaeva, E.; Pronin, N. Genetic diversity of leeches in Lake Gusinoe (Eastern Siberia, Russia). Sci. World J. 2014, 2014, 619127. [Google Scholar] [CrossRef]
- Kaygorodova, I. Annotated checklist of the leech species diversity in the Maloe More Strait of Lake Baikal, Russia. ZooKeys 2015, 545, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Bolbat, A.; Bukin, Y.; Kaygorodova, I. Genome-based taxa delimitation (GBTD): A new approach. Diversity 2022, 14, 948. [Google Scholar] [CrossRef]
- Grosser, C.; Kutschera, U. Feeding behaviour and reproductive biology of the semiaquatic leech Trocheta haskonis (Hirudinea: Erpobdellidae). Lauterbornia 2004, 52, 163–169. [Google Scholar]
- Kutschera, U.; Pfeiffer, I.; Ebermann, E. The European land leech: Biology and DNA-based taxonomy of a rare species that is threatened by climate warming. Naturwissenschaften 2007, 94, 967–974. [Google Scholar] [CrossRef]
- Jueg, U.; Grosser, C. Erpobdellopsis graacki n. gen., n. sp. —A peculiar leech from Spain (Annelida, Hirudinida: Erpobdellidae). Lauterbornia 2017, 84, 69–87. [Google Scholar]
- ICZN. International Code of Zoological Nomenclature, 4th ed.; The International Trust for Zoological Nomenclature: London, UK, 1999; Available online: https://www.iczn.org/the-code/the-code-online/ (accessed on 20 March 2024).

| Description | Max Score | Query Cover | Identity | Length, bp | Accession |
|---|---|---|---|---|---|
| Erpobdella vilnensis | 843 | 87% | 93.76% | 563 | HM246551 |
| Erpobdella vilnensis | 874 | 92% | 93.39% | 592 | HM246585 |
| Erpobdella vilnensis | 852 | 91% | 92.98% | 584 | DQ009663 |
| Erpobdella sp. | 832 | 89% | 92.84% | 575 | HM246537 |
| Erpobdella vilnensis | 900 | 100% | 92.03% | 659 | KP300763 |
| Erpobdella japonica | 815 | 99% | 89.67% | 14725 | NC_036150 |
| Erpobdella sp. | 809 | 99% | 89.62% | 675 | KM095091 |
| Erpobdella japonica | 800 | 99% | 89.43% | 650 | AF116026 |
| Erpobdella obscura | 798 | 99% | 89.20% | 658 | JQ821638 |
| Erpobdella octoculata | 765 | 97% | 88.68% | 651 | AF003274 |
| Method | Threshold Distance (TD) | Total Number of Subsets | Number of Species in Cryptic Groups | |||
|---|---|---|---|---|---|---|
| D. lineata | E. dubia/parva | E. microstoma | M. montezuma | |||
| Morphology | visual | 29 | 1 | 2 | 1 | 1 |
| p-distance | 0.0247 | 47 | 1 | 1 | 1 | 1 |
| ASAP | 0.0262 | 48 | 2 | 1 | 2 | 1 |
| BI+GMYC | 0.0268 | 50 | 2 | 2 | 2 | 2 |
| bPTP | 0.0308 | 49 | 2 | 1 | 2 | 2 |
| ON BALANCE | 47 | 1 | 1 | 2 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaygorodova, I. Application of Modern Approaches to Species Delimitation in Freshwater Erpobdellidae (Hirudinea: Arhynchobdellida) with a New Interpretation of Their Genera. Water 2024, 16, 1030. https://doi.org/10.3390/w16071030
Kaygorodova I. Application of Modern Approaches to Species Delimitation in Freshwater Erpobdellidae (Hirudinea: Arhynchobdellida) with a New Interpretation of Their Genera. Water. 2024; 16(7):1030. https://doi.org/10.3390/w16071030
Chicago/Turabian StyleKaygorodova, Irina. 2024. "Application of Modern Approaches to Species Delimitation in Freshwater Erpobdellidae (Hirudinea: Arhynchobdellida) with a New Interpretation of Their Genera" Water 16, no. 7: 1030. https://doi.org/10.3390/w16071030
APA StyleKaygorodova, I. (2024). Application of Modern Approaches to Species Delimitation in Freshwater Erpobdellidae (Hirudinea: Arhynchobdellida) with a New Interpretation of Their Genera. Water, 16(7), 1030. https://doi.org/10.3390/w16071030