
Development of a Model to Evaluate Water Conservation Function for Various Tree Species

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Forest Model
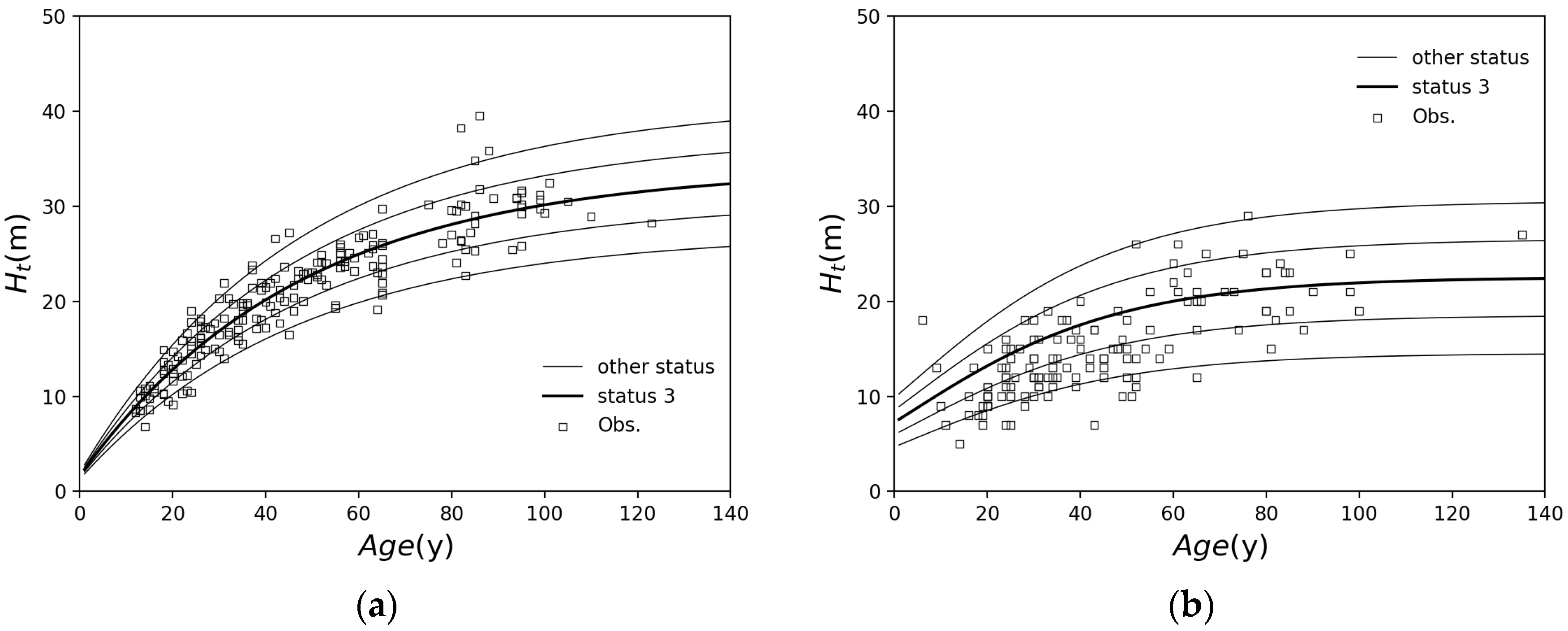
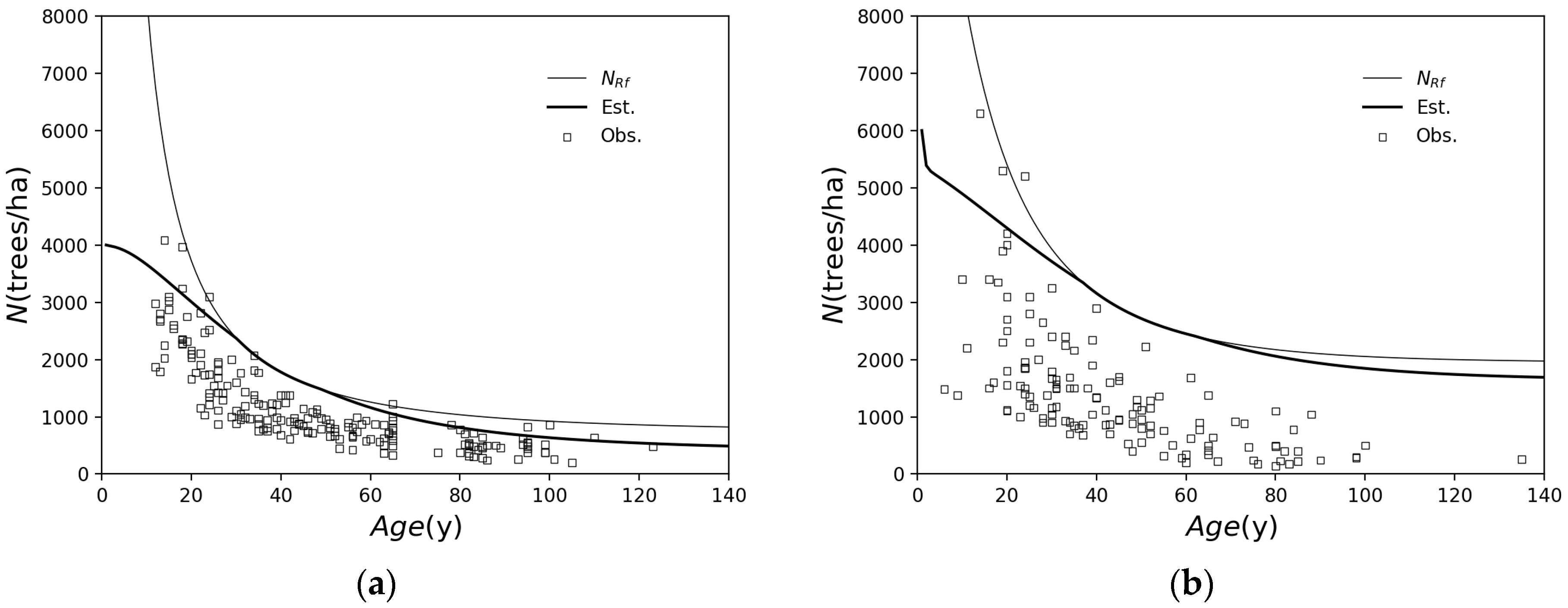
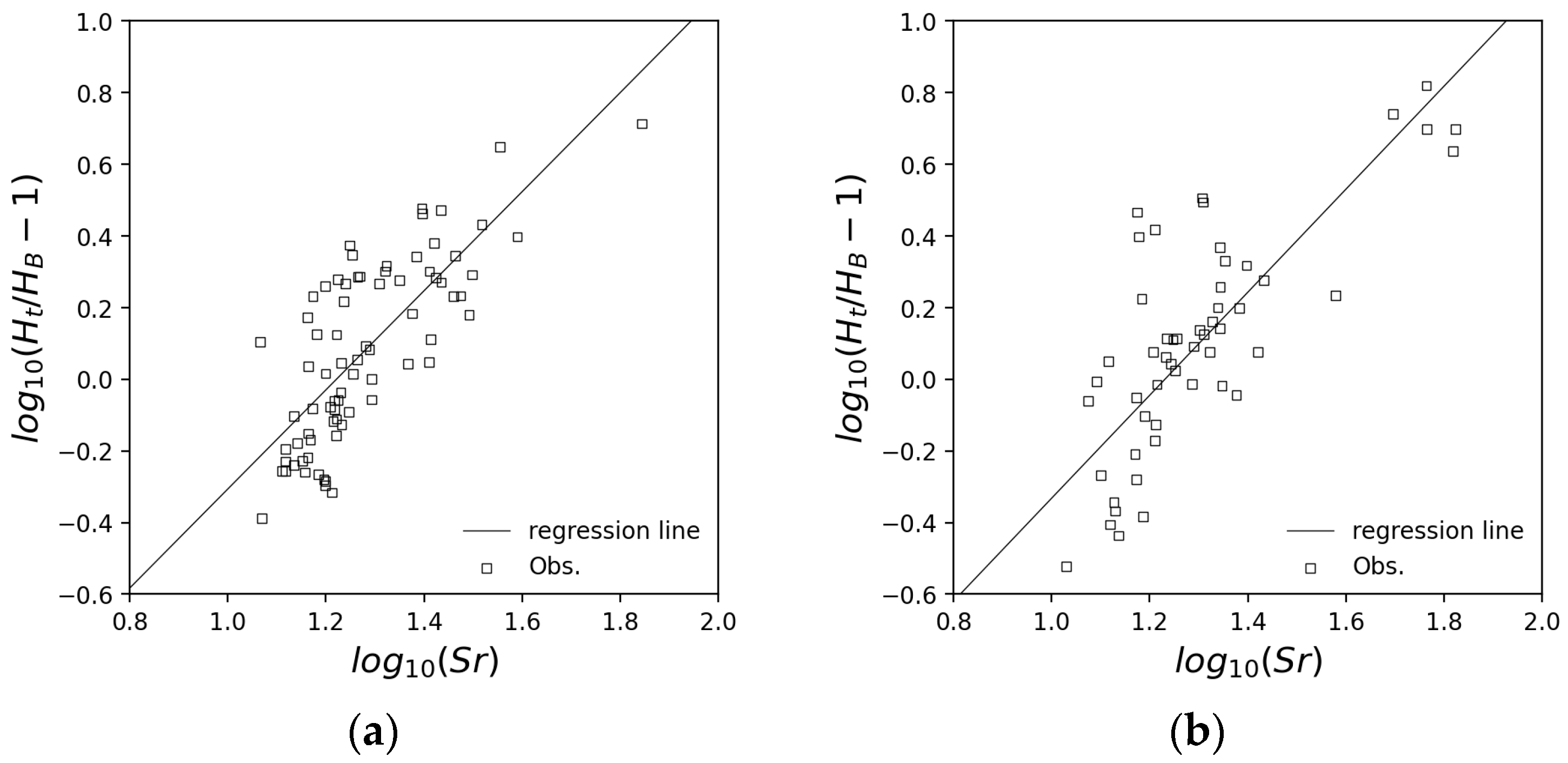
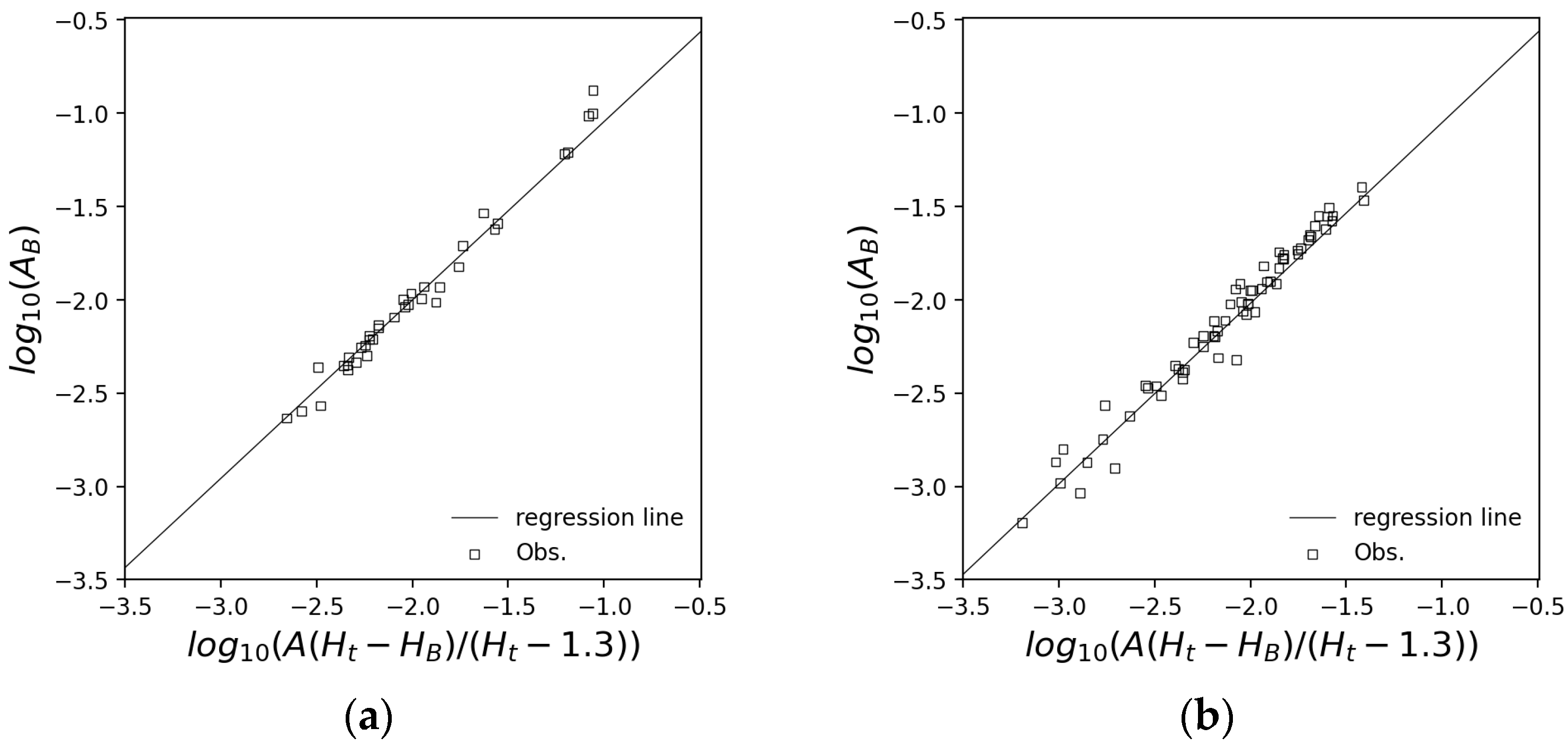
2.1.1. Forest Growth Model
2.1.2. LAI Estimation Model
2.2. Hydrological Model
2.3. Carbon Stock Assessment Model
3. Results and Discussion
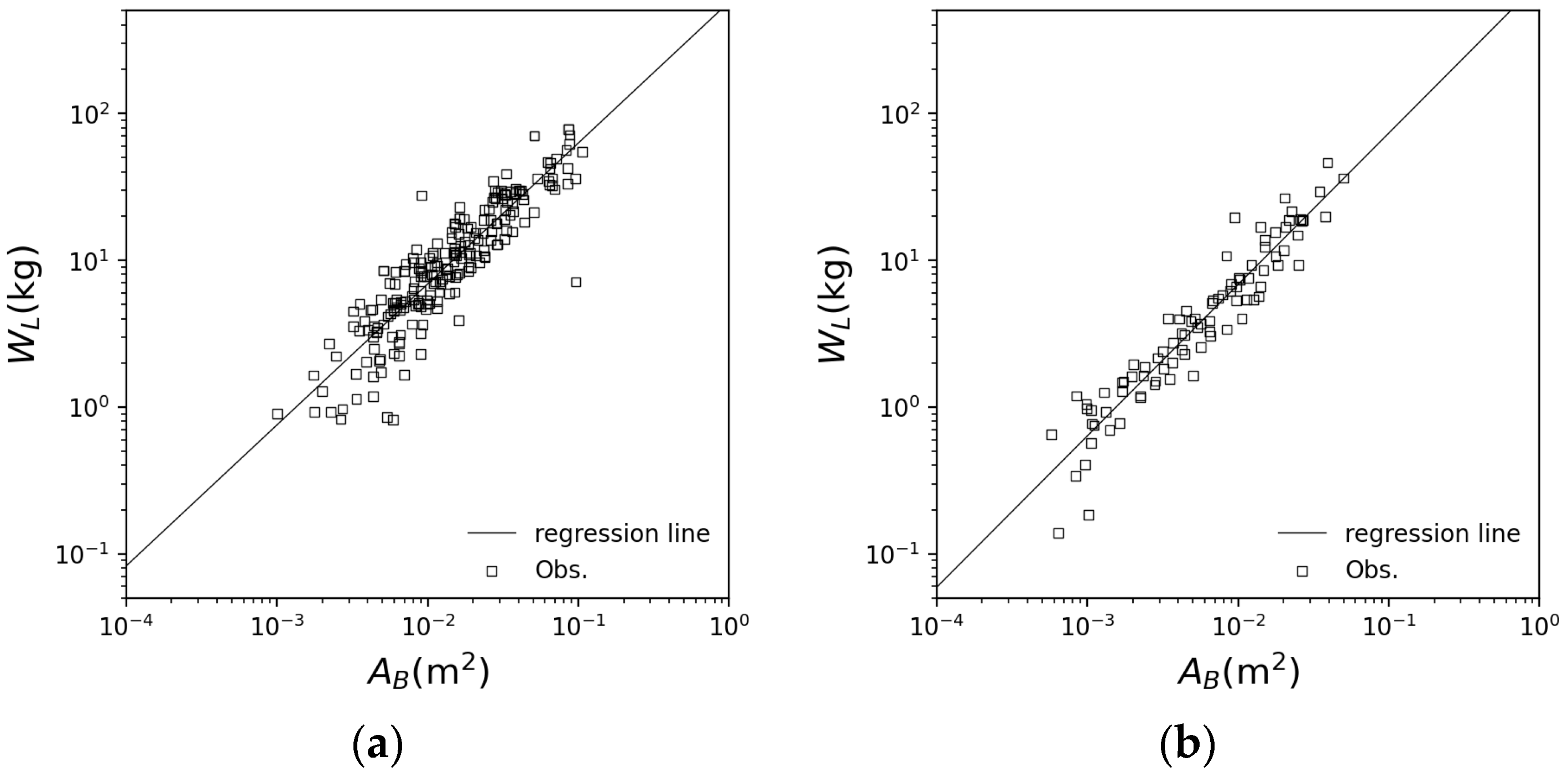
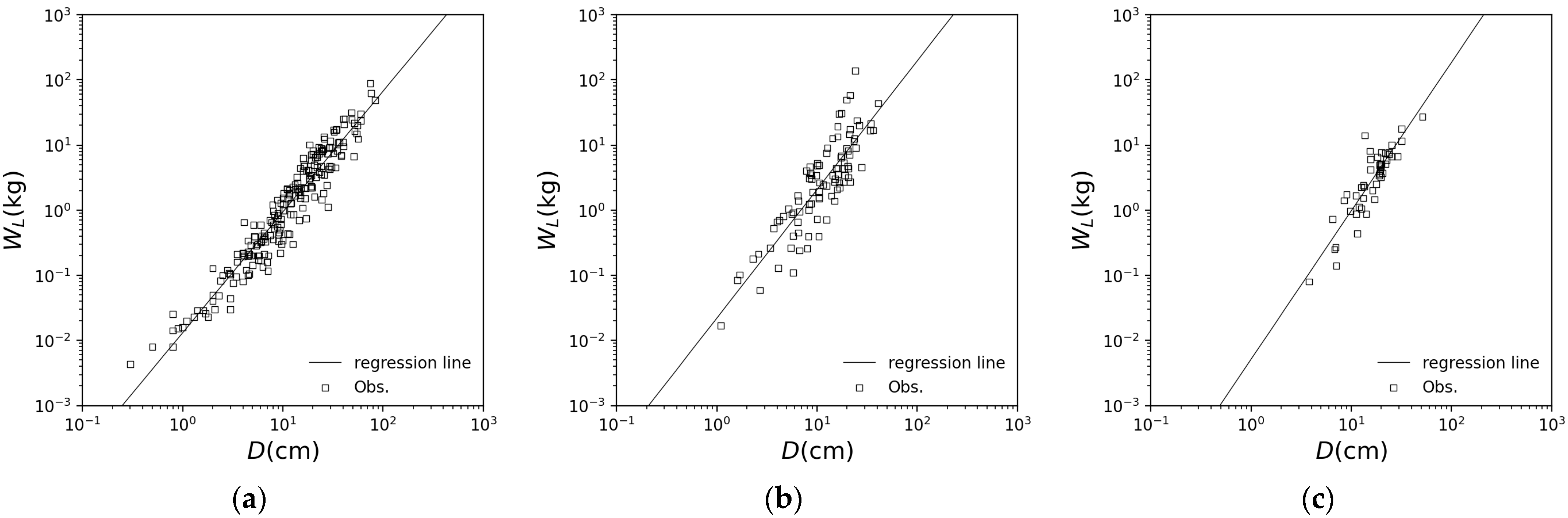
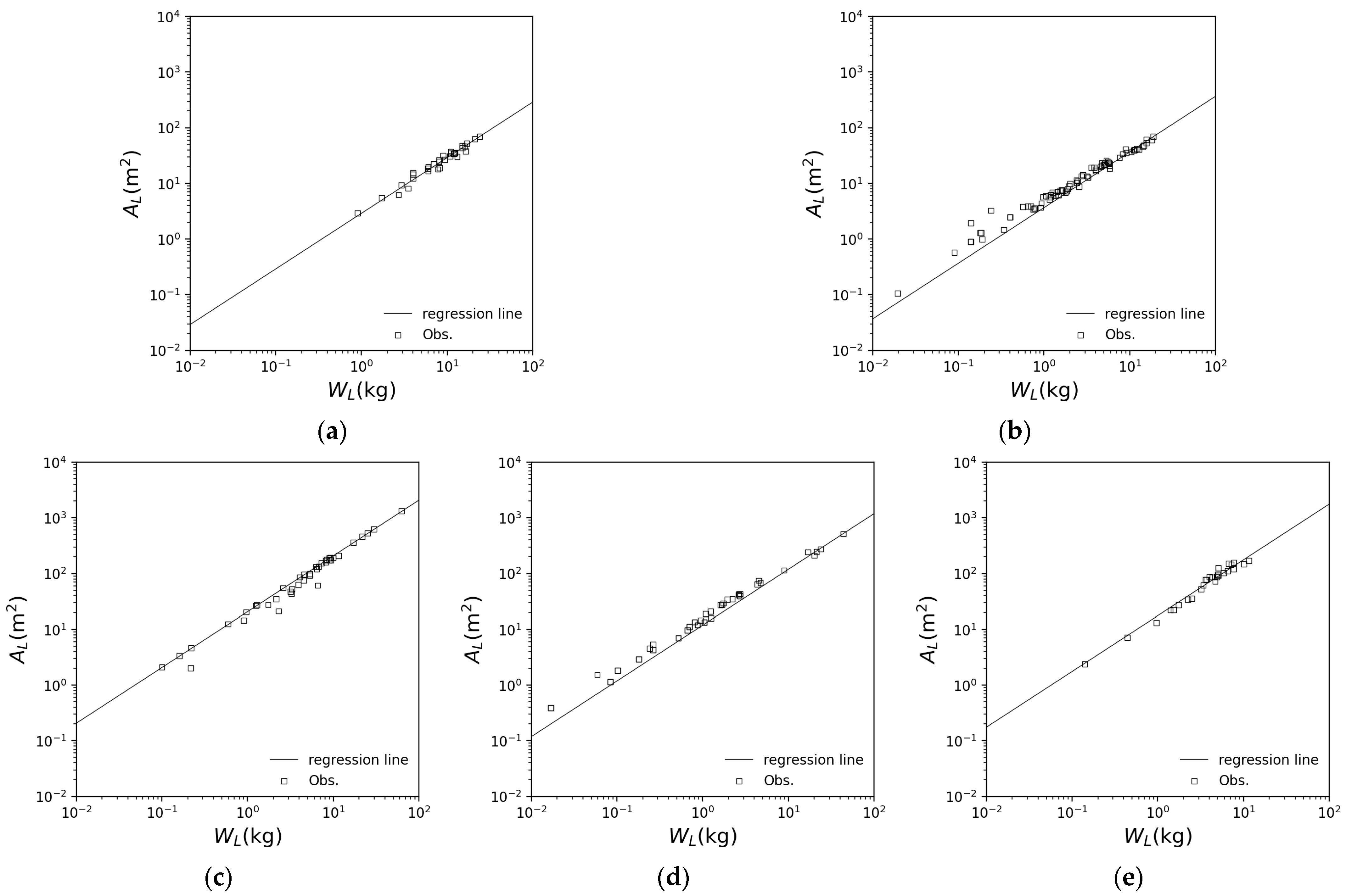
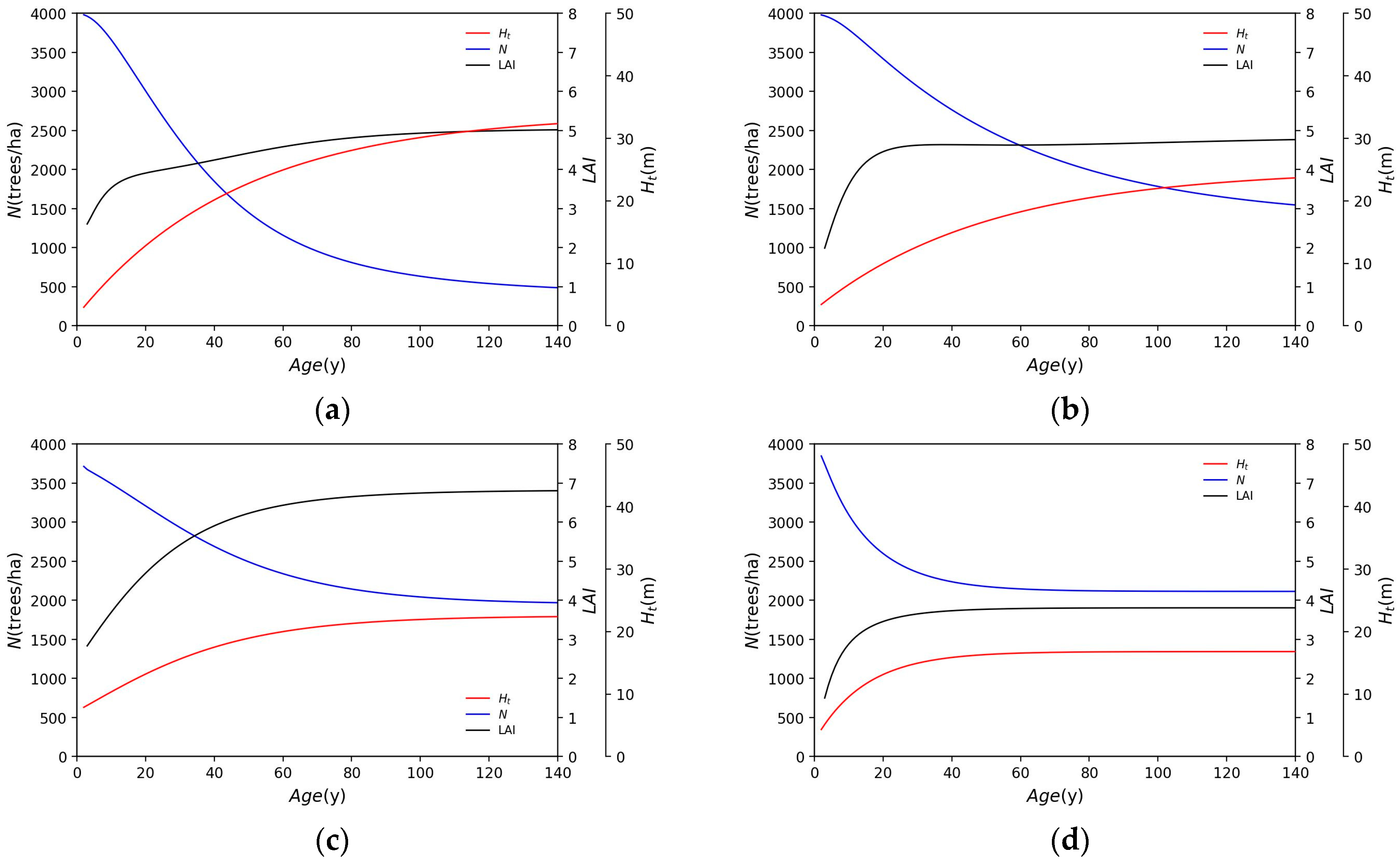
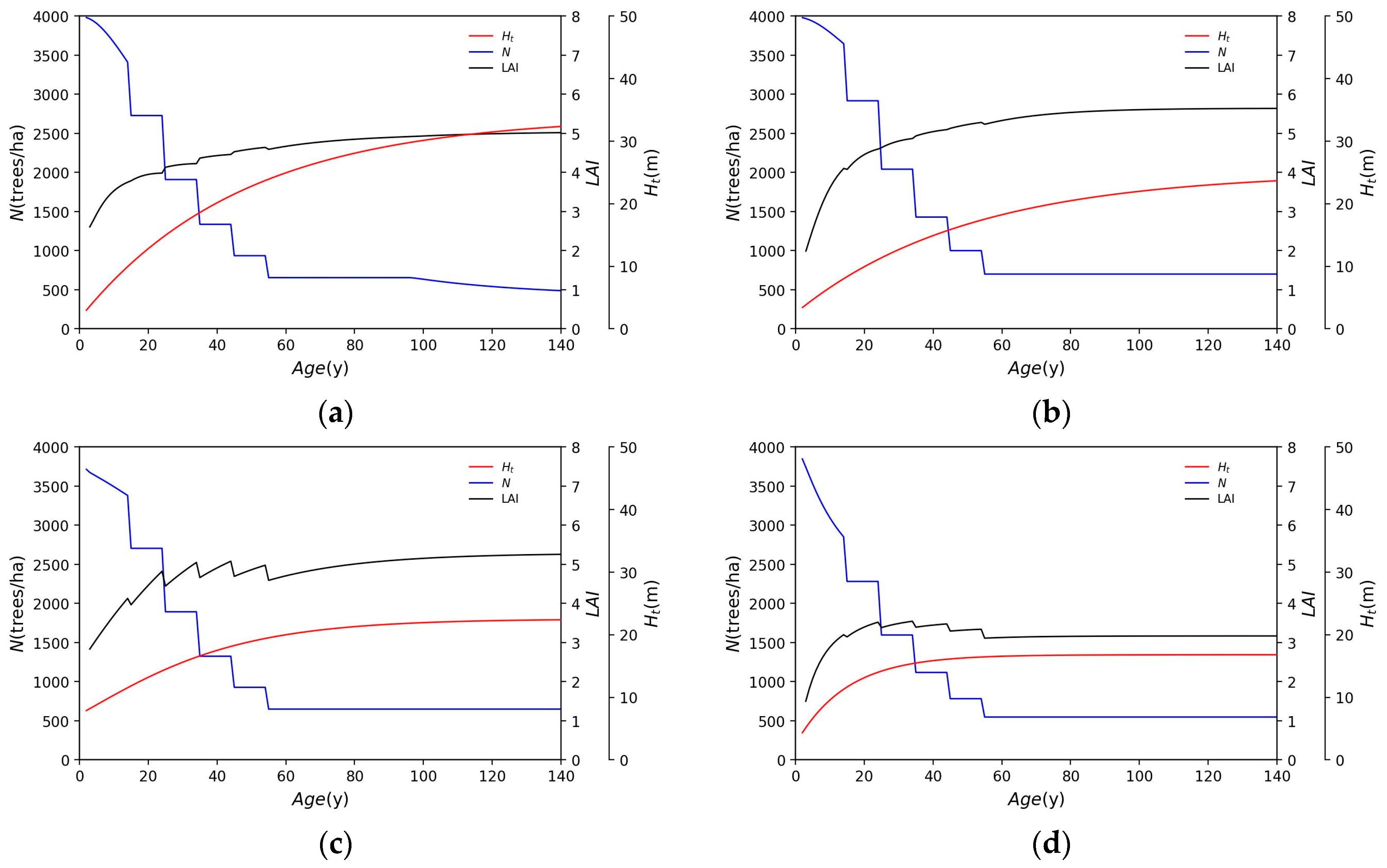
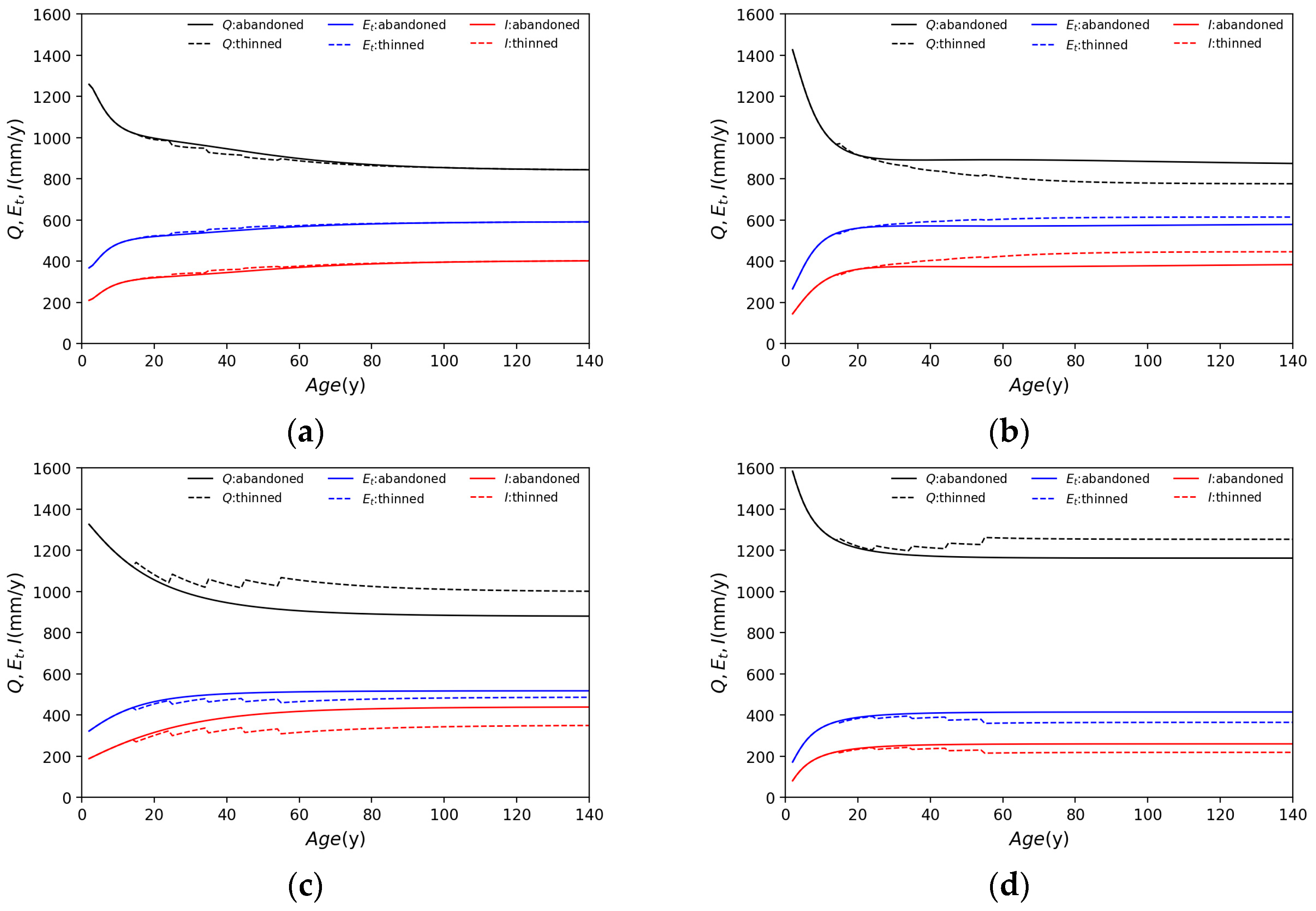
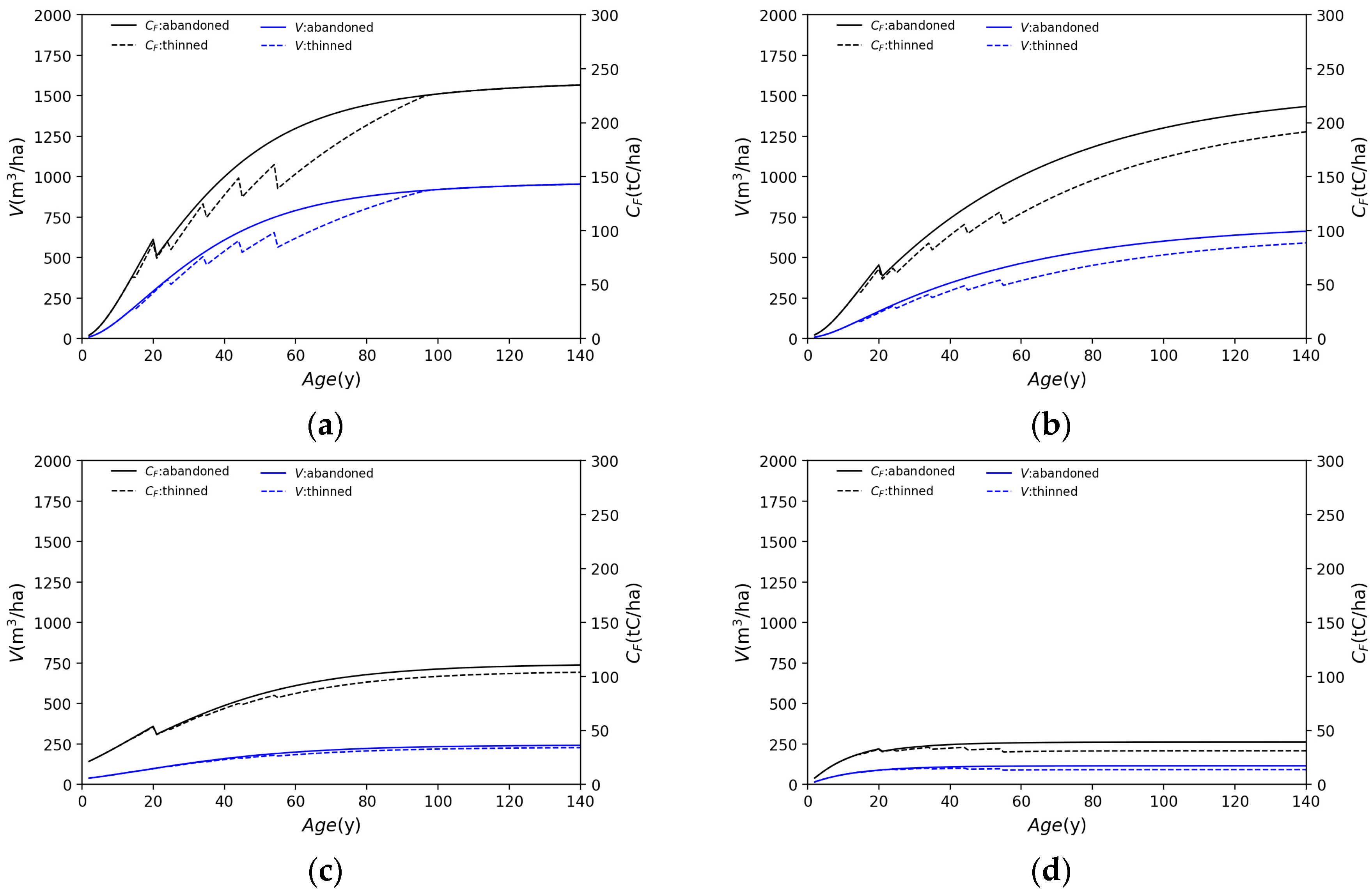
3.1. Application of Each Species
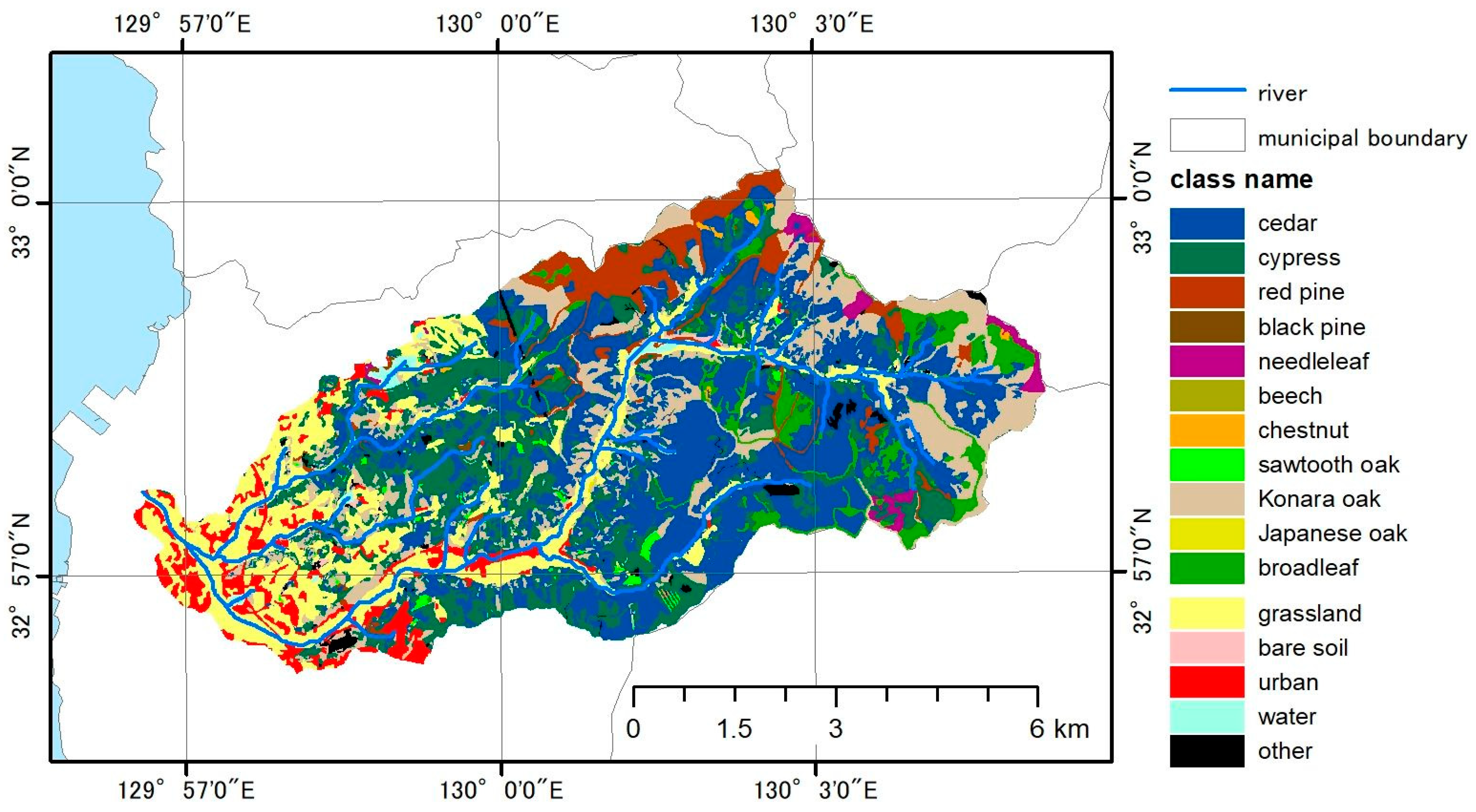
3.2. Basin-Scale Application
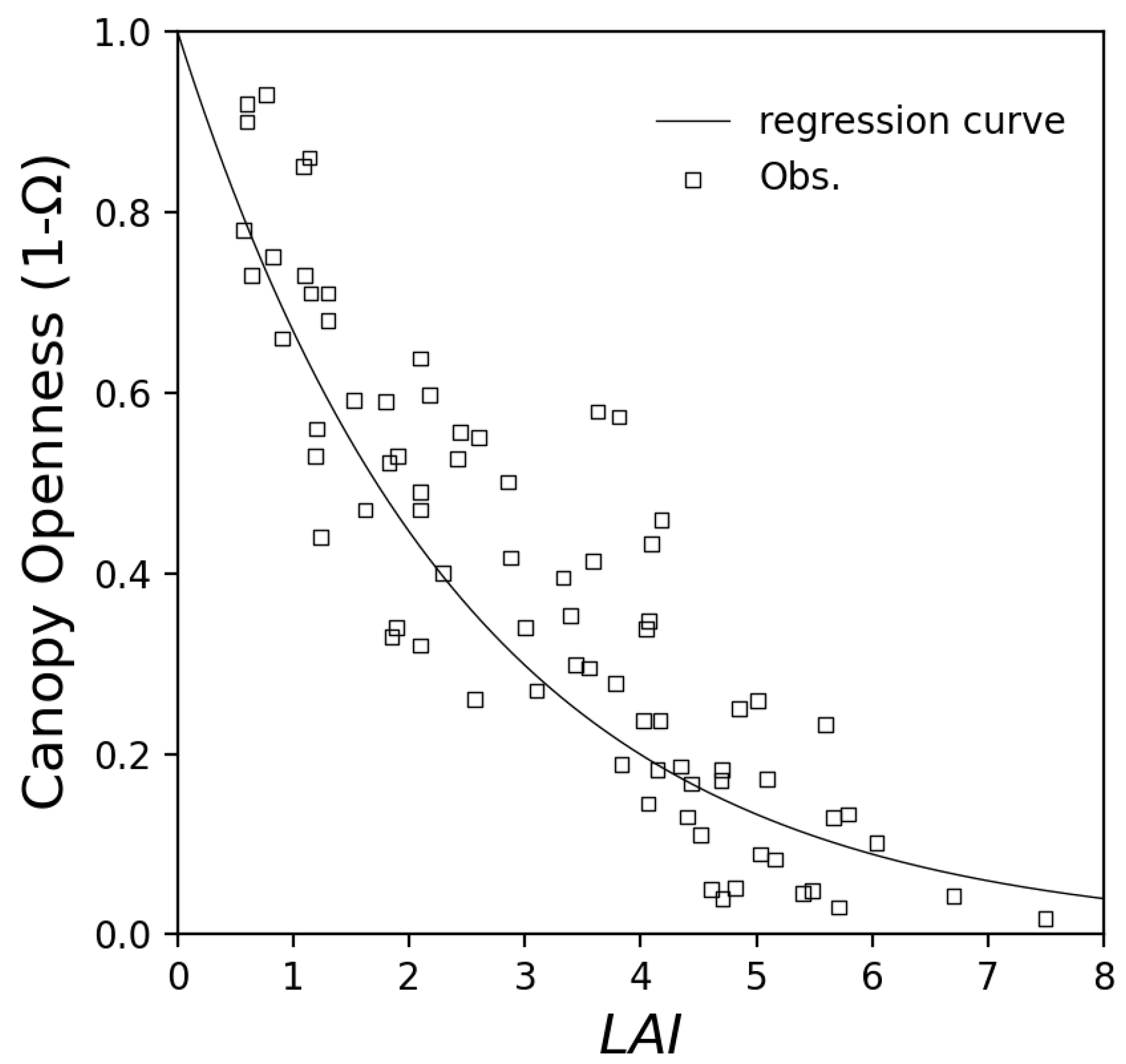
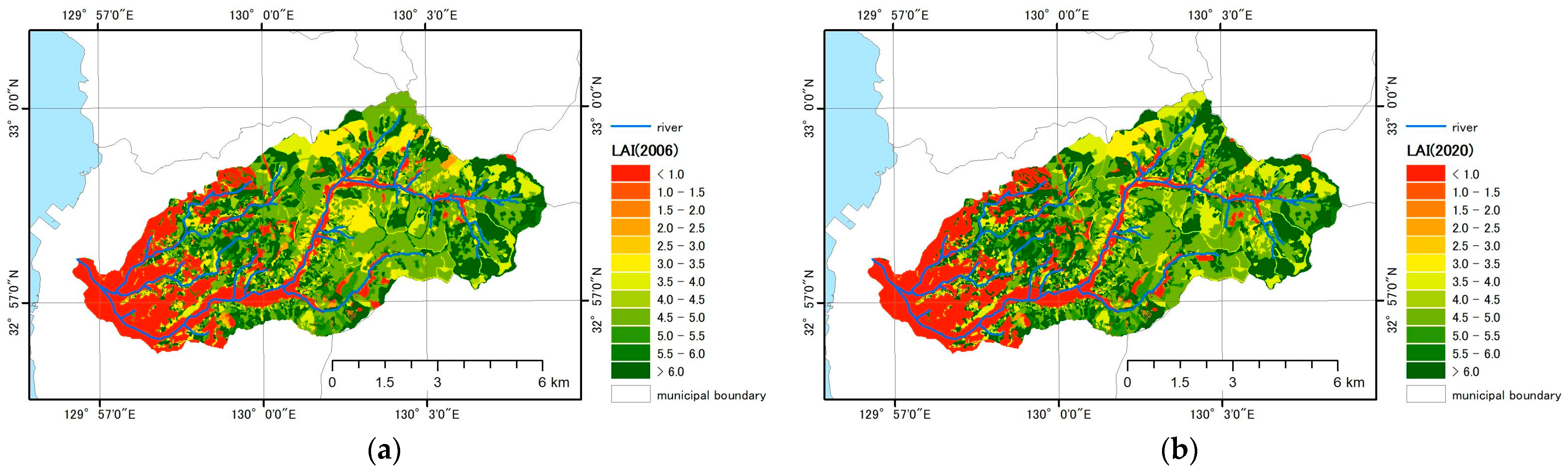
3.2.1. LAI Estimation
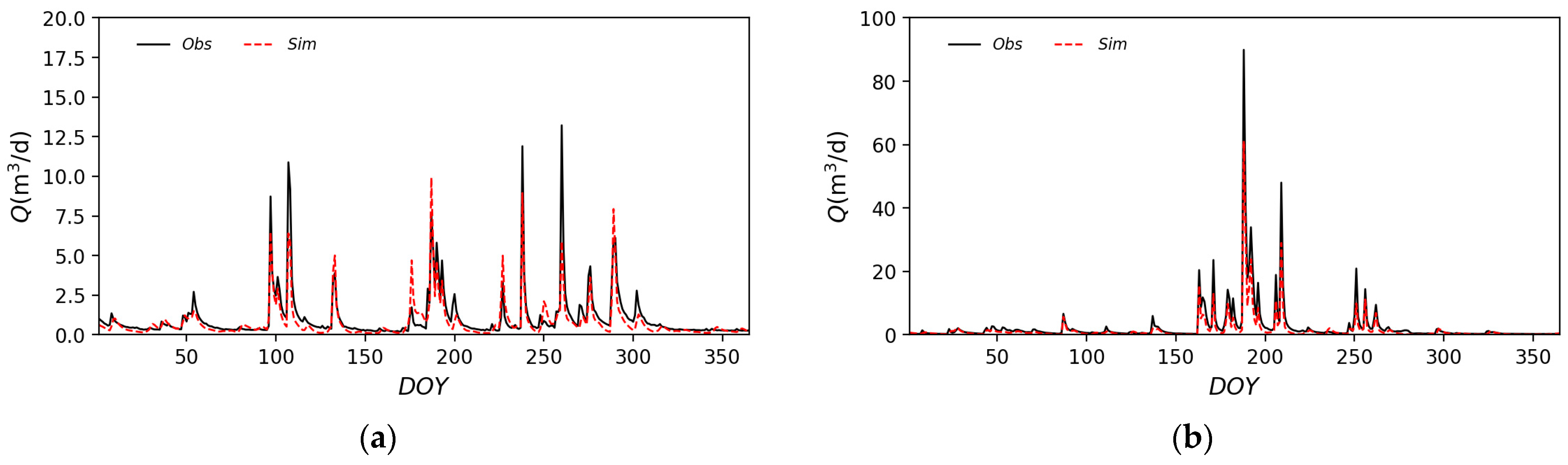
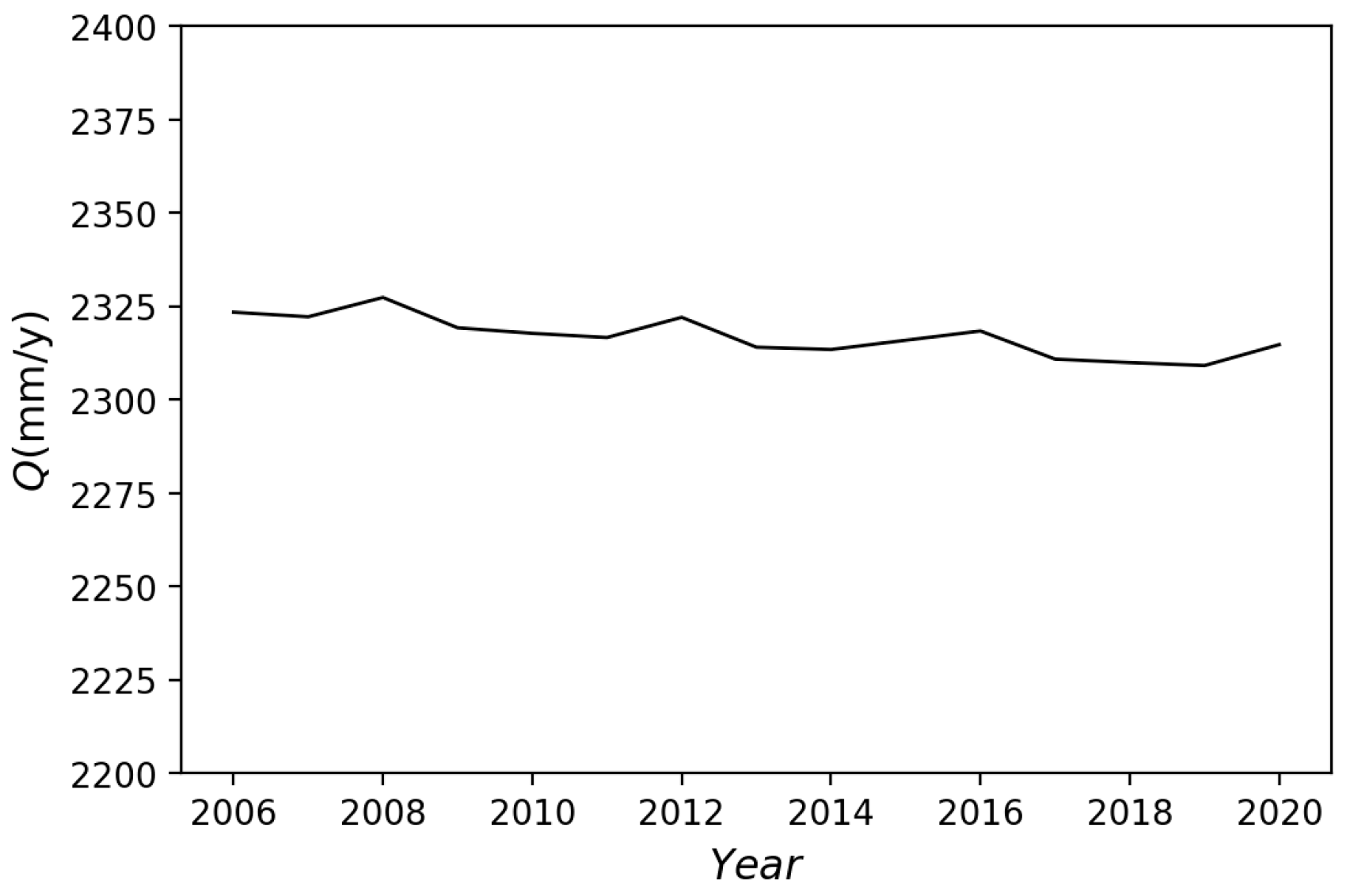
3.2.2. Hydrological Application and Validation
4. Conclusions
- Water supply varies with tree species but generally decreases until about 40 years of age, after which it is near constant.
- Cedar is less affected than other tree species by thinning. The water supply is barely affected by thinning.
- The thinning of cypress trees increases the LAI due to reduced density, and water supply is reduced in thinned forests compared to in abandoned forests, influenced by an increased LAI.
- Although beech forests have a larger LAI than needleleaf forests, water supply is not significantly different.
- Birch has a smaller LAI and a much larger water supply than other species.
- Broadleaf forests are more affected by thinning than needleleaf forests and tend to receive an increased water supply as a result of thinning.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Type L-1: the species that grow well initially but then stop growing, such as beech (Fagus crenata), Mizunara oak (Quercus crispula), Keyaki (Zelkova serrata), a kind of walnut (Juglans), horsechestnuts (Aesculus turbinata), and so on.
- Type L-2: the species with slow initial growth and subsequent growth as well, such as Konara oak (Quercus serrata), cherry blossom (Cerasus jamasakura), chestnuts (Castanea crenata), Honoki (Magnolia obovata), maple kind (Acer), and so on.
- Type L-3: the species with poor initial growth but long-lasting growth afterwards, such as birch (Betula), alder (Alnus japonica), and so on.

| Region | Gifu | Fukuoka | Nagasaki | Kyushu | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | broadleaf forest | ||||||||||
| cedar | cypress | red pine | larch | L-1 | L-2 | L-3 | cedar | cypress | cypress | red pine | |
| Growth curve | Equation (1) | Equation (1) | Equation (3) | Equation (3) | Equation (3) | Equation (1) | Equation (1) | Equation (2) | Equation (3) | Equation (1) | Equation (3) |
| 34.02 | 25.07 | 2.886 | 3.250 | 3.115 | 21.82 | 16.81 | 30.30 | 3.378 | 30.86 | 2.977 | |
| 0.9522 | 0.8986 | 0.8120 | 0.6541 | 0.3634 | 0.7109 | 0.8504 | 1.368 | 0.6353 | 0.9473 | 0.7617 | |
| 0.02120 | 0.01992 | 0.06159 | 0.06772 | 0.0375 | 0.0262 | 0.06793 | 0.04403 | 0.02347 | 0.01388 | 0.05550 | |
| 0.07736 | 0.06897 | 0.07798 | 0.1794 | 0.7017 | 0.08544 | 0.09967 | 0.04980 | 0.05239 | 0.04933 | 0.1326 | |
| −1.417 | −1.242 | −1.313 | −1.519 | −1.662 | −1.015 | −0.8997 | −1.326 | −1.406 | −1.206 | −1.481 | |
| 9612 | 46673 | 9775 | 3222 | 16944 | 7077 | 15909 | 773.5 | 5274 | 8676 | 2110 | |
| −3.021 | −3.853 | −3.149 | −2.638 | −3.569 | −2.773 | −3.251 | −2.275 | −2.881 | −3.262 | −2.872 | |
| −0.6833 | −0.4443 | −0.8869 | 0.6736 | 1.547 | 1.2867 | 1.348 | 2.356 | 0.7746 | 0.4063 | 1.573 | |
| 0.3736 | 0.4698 | 0.3780 | 0.4767 | 0.3768 | 0.3040 | 0.2967 | 0.2615 | 0.3768 | 0.4247 | 0.3726 | |
| 0.2528 | 0.1924 | 0.1551 | −0.0005389 | −0.2242 | −0.09113 | −0.008533 | 0.2612 | 0.1410 | 0.1574 | 0.1082 | |
| −0.1028 | −0.04263 | −0.3429 | −0.2018 | −0.2038 | −0.08979 | 0.4594 | 0.6868 | −0.2465 | 0.04933 | −0.5569 | |
| 0.9709 | 0.9765 | 0.9886 | 0.9693 | 0.9544 | 0.9401 | 0.8915 | 0.9767 | 0.9624 | 0.9916 | 0.9817 | |
| 0.0000 | 0.0000 | 0.0000 | 0.0005188 | 0.02688 | 0.1031 | 0.04779 | −0.03031 | 0.1410 | −0.02918 | 0.04130 | |
| 0.34 | 0.10 | 0.22 | 0.043 | 0.0315 | 0.146 | 0.0626 | 0.291 | 0.393 | 0.162 | 0.115 | |
Appendix B

| Mizunara Oak | Keyaki | Cherry Blossom | Maple | Chestnuts | Honoki | |
|---|---|---|---|---|---|---|
| 1.752 | 1.677 | 2.097 | 2.036 | 1.478 | 2.456 | |
| −1.703 | −1.494 | −2.298 | −2.183 | −1.329 | −2.711 | |
| 0.937 | 0.849 | 0.858 | 0.924 | 0.937 | 0.965 |

| Mizunara Oak | Keyaki | Cherry Blossom | Maple | Chestnuts | Honoki | |
|---|---|---|---|---|---|---|
| 12.09 | 12.50 | 20.42 | 15.29 | 11.96 | 15.59 | |
| 0.997 | 0.990 | 0.986 | 0.867 | 0.966 | 0.993 |
References
- Bagstad, K.J.; Johnson, G.W.; Voigt, B.; Villa, F. Spatial dynamics of ecosystem service flows: A comprehensive approach to quantifying actual services. Ecosyst. Serv. 2013, 4, 117–125. [Google Scholar] [CrossRef]
- Government Public Relations Office, Japan. Public Opinion Survey on Forest and Livelihoods. Available online: https://survey.gov-online.go.jp/r01/r01-sinrin/ (accessed on 5 December 2023). (In Japanese)
- Kohsaka, R.; Uchiyama, Y. Status and Trends in Forest Environment Transfer Tax and Information Interface between Prefectures and Municipalities: Multi-Level Governance of Forest Management in 47 Japanese Prefectures. Sustainability 2022, 14, 1791. [Google Scholar] [CrossRef]
- Thornton, P.E.; Law, B.E.; Gholz, H.L.; Clark, K.L.; Falge, E.; Ellsworth, D.S.; Goldstein, A.H.; Monson, R.K.; Hollinger, D.; Falk, M.; et al. Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agric. For. Meteorol. 2002, 113, 185–222. [Google Scholar] [CrossRef]
- Yamaura, Y.; Yamada, Y.; Matsuura, T.; Tamai, K.; Taki, H.; Sato, T.; Hashimoto, S.; Murakami, W.; Toda, K.; Saito, H.; et al. Modeling impacts of broad-scale plantation forestry on ecosystem services in the past 60 years and for the future. Ecosyst. Serv. 2021, 49, 101271. [Google Scholar] [CrossRef]
- Cheng, X.; Bai, Y.; Zhu, J.; Han, H. Effects of forest thinning on interception and surface runoff in Larix principis-rupprechtii plantation during the growing season. J. Arid Environ. 2020, 181, 104222. [Google Scholar] [CrossRef]
- Chen, L.; Yuan, Z.; Shao, H.; Wang, D.; Mu, X. Effects of Thinning Intensities on Soil Infiltration and Water Storage Capacity in Chinese Pine-Oak Mixed Forest. Sci. World J. 2014, 2014, 268157. [Google Scholar] [CrossRef]
- Komatsu, H. Relationship between Stem Density and Interception Ratio for Coniferous Plantation Forests in Japan. J. Jpn. For. Soc. 2007, 89, 217–220. (In Japanese) [Google Scholar] [CrossRef][Green Version]
- Watanabe, N.; Kojima, T.; Shinoda, S.; Ohashi, K.; Tamagawa, I.; Saitoh, T. Observation and Modeling of Rainfall Interception in Evergreen Forest and Deciduous Forest. J. Jpn. Soc. Civ. Eng. Ser. B1 Hydraul. Eng. 2012, 68, I_1759–I_1764. (In Japanese) [Google Scholar] [CrossRef]
- Komatsu, H.; Shinohara, Y.; Kume, T.; Otsuki, K. Changes in peak flow with decreased forestry practices: Analysis using watershed runoff data. J. Environ. Manag. 2011, 92, 1528–1536. [Google Scholar] [CrossRef]
- Gao, X.; Huang, X.; Chang, S.; Dang, Q.; Wen, R.; Lo, K.; Li, J.; Yan, A. Long-term improvement in water conservation function at Qilian Mountain national Park northwest China. J. Mt. Sci. 2023, 20, 2885–2897. [Google Scholar] [CrossRef]
- Forestry Resources Management Division of Gifu Prefecture. Forest Harvest Table/Forest Density Control Chart for Japanese Cedar Planted Forest; Forestry Resources Management Division of Gifu Prefecture: Gifu, Japan, 1992. (In Japanese)
- Forestry Resources Management Division of Gifu Prefecture. Forest Harvest Table/Forest Density Control Chart for Japanese Cypress Planted Forest; Forestry Resources Management Division of Gifu Prefecture: Gifu, Japan, 1992. (In Japanese)
- Forestry Resources Management Division of Gifu Prefecture. Forest Harvest Table/Forest Density Control Chart for Japanese Redpine Planted Forest; Forestry Resources Management Division of Gifu Prefecture: Gifu, Japan, 1984. (In Japanese)
- Forestry Resources Management Division of Gifu Prefecture. Forest Harvest Table/Forest Density Control Chart for Japanese Larch Planted Forest; Forestry Resources Management Division of Gifu Prefecture: Gifu, Japan, 1985. (In Japanese)
- Forestry Resources Management Division of Gifu Prefecture. Forest Harvest Table/Forest Density Control Chart for Broad Leaf Forest; Forestry Resources Management Division of Gifu Prefecture: Gifu, Japan, 1992. (In Japanese)
- Maeda, H. Management standard for Hinoki (Chamaecyparis obtuse) plantations applied to long-term management in Nagasaki prefectures. Bull. Nagasaki Agric. For. Technol. Dev. Cent. 2012, 3, 54–64. (In Japanese) [Google Scholar]
- Forestry Experiment Station, Kyushu Region Hinoki Forest Harvest Table Adjustment Instructions. Harvest Table Adjustment Operations Research Materials, 19; Kumamoto For. Off., Forest Agency: Kumamoto, Japan, 1957; p. 49. (In Japanese)
- Forestry Experiment Station, Kitakyushu Region Akamatsu Forest Harvest Table Adjustment Instructions. Harvest Table Adjustment Operations Research Materials, 25; Kumamoto For. Off., Forest Agency: Kumamoto, Japan, 1960; p. 46. (In Japanese)
- Narasaki, K.; Maeda, H.; Sasaki, S. Adjustment of Japanese cedar forest density control chart and status index curve for preparation of Fukuoka Prefecture version of system harvest table. Bull. Fukuoka Agric. For. Res. Cent. 2015, 1, 38–43. (In Japanese) [Google Scholar]
- Shinozaki, K.; Yoda, K.; Hozumi, K.; Kira, T. A Quantitative Analysis of Plant Form-The Pipe Model Theory I. Basic Analyses. Ecol. Soc. Jpn. 1964, 14, 97–105. [Google Scholar]
- Ando, T.; Hatiya, K.; Doi, K.; Kataoka, H.; Kato, Y.; Sakaguchi, K. Studies on the system of density control of sugi (Cryptomeria japonica) stand. Bull. Gov. For. Exp. Stn. 1968, 209, 1–76. (In Japanese) [Google Scholar]
- Makungwa, S.D.; Kodani, J. Estimation of standing crop and productivity of an even-aged young Keyaki (Zelkova serrata Makino) plantation in Suzu prefectural Forest. Bull. Ishikawa-Ken For. Exp. Stn. 1998, 29, 6–11. (In Japanese) [Google Scholar]
- Kanazawa, Y.; Kiyono, Y.; Fujimori, T. Crown Development and Stem Growth in Relation to Stand Density in Even-Aged Pure Stands (II) Cear-Length Model of Cryptomeria japonica Stands as a Function of Stand Density and Tree Height. J. Jpn. For. Soc. 1985, 67, 391–397. [Google Scholar]
- Watanabe, H.; Moteki, Y. Growth progress and biomass in 92-years-old plantation of Japanese cedar. Bull. Gifu Prefect. Res. Inst. For. 2007, 36, 7–13. (In Japanese) [Google Scholar]
- Ishii, T.; Nashimoto, M.; Shimogaki, H. Development of forest observation method using remote sensing data -Estimation of leaf area index-. In Annual Research Report; Central Research Institute of Electric Power Industry: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Inagaki, Y.; Nakanishi, A.; Tange, T. A simple method for leaf and branch biomass estimation in Japanese cedar plantations. Trees 2020, 34, 349–356. [Google Scholar] [CrossRef]
- Saito, H.; Kan, M.; Shidei, T. Studies on the effects of thinning from small diameter trees (I). Changes in stand condition before and after thinning. Bull. Kyoto Univ. For. 1966, 38, 50–67. (In Japanese) [Google Scholar]
- Saito, H.; Yamada, I.; Shidei, T. Studies on the effects of thinning from small diameter trees (II). Changes in stand condition after single growing season. Bull. Kyoto Univ. For. 1967, 39, 64–78. (In Japanese) [Google Scholar]
- Saito, H.; Tamai, S.; Ogino, K.; Shidei, T. Studies on the effects of thinning from small diametered trees (III). Changes in stand condition after the second growing season. Bull. Kyoto Univ. For. 1968, 40, 81–92. (In Japanese) [Google Scholar]
- Saito, H.; Shidei, T. Studies on the productivity and its estimation methodology in a young stand of Cryptomeria japonica D. Don. J. Jpn. For. Soc. 1973, 55, 52–62. (In Japanese) [Google Scholar]
- Tadaki, Y.; Kawasaki, Y. Studies on the production structure of forest. IX. Primary productivity of a young Cryptomeria plantation with excessively high stand density. J. Jpn. For. Soc. 1966, 48, 55–61. [Google Scholar]
- Tadaki, Y.; Ogata, N.; Nagatomo, Y. Studies on production structure of forest. XI. Primary productivities of 28-years-old plantations of Cryptomeria of cuttings and of seedlings origin. Bull. Gov. For. Exp. Stn. 1967, 199, 47–65. (In Japanese) [Google Scholar]
- Inagaki, Y.; Miyamoto, K.; Itou, T.; Kitahara, F.; Sakai, H.; Okuda, S.; Noguchi, M.; Mitsuda, Y. Estimation of leaf biomass cypress plantation in Kochi Prefecture. Appl. For. Sci. 2015, 24, 11–18. (In Japanese) [Google Scholar]
- Inagaki, Y.; Fukata, H.; Noguchi, K.; Kuramoto, S.; Nakanishi, A. Recovery of leaf biomass after thinning of hinoki cypress plantations in Kochi Prefecture. Appl. For. Sci. 2018, 27, 1–9. (In Japanese) [Google Scholar]
- Ishii, T. Estimation of Forest Leaf Area Index Using Remote Sensing Data and Its Applications to Predicting Forest Carbon Sequestration and Water Budget. Ph.D. Thesis, Waseda University, Tokyo, Japan, 2007. (In Japanese). [Google Scholar]
- Wakiyama, Y.; Onda, Y.; Nanko, K.; Mizugaki, S.; Kim, Y.; Kitahara, H.; Ono, H. Estimation of temporal variation in splash detachment in two Japanese cypress plantations of contrasting age. Earth Surf. Process. Landf. 2010, 35, 995–1005. [Google Scholar] [CrossRef]
- Nakaya, K.; Wakamatsu, T.; Ikeda, H.; Abe, S.; Toyoda, Y. Development of raindrop kinetic energy model under canopy for the estimation of soil erosion in forest. In Annual Research Report; Central Research Institute of Electric Power Industry: Tokyo, Japan, 2011. (In Japanese) [Google Scholar]
- Takeuchi, I.; Kawasaki, T.; Mori, S. Changes of stem height-to-diameter ratio in hinoki (Chamaecyparis obtuse) young man-made stands. J. Jpn. For. Soc. 1997, 79, 137–142. (In Japanese) [Google Scholar]
- Watanabe, H.; Moteki, Y.; Obora, T. Effects of Thinning on Stand Structure and Diameter Growth in Old and Overcrowded Japanese Cypress (Chamaecyparis obtuse) Stands. J. Jpn. For. Soc. 2015, 97, 182–185. (In Japanese) [Google Scholar] [CrossRef]
- Sumida, A.; Nakai, T.; Yamada, M.; Ono, K.; Uemura, S.; Hara, T. Ground-Based Estimation of Leaf Area Index and Vertical Distribution of Leaf Area Density in a Betula ermanii Forest. Silva Fenn. 2009, 43, 799–816. [Google Scholar] [CrossRef]
- Kawanabe, S.; Ando, M. Studies on regeneration of natural forest on lower limit of cool temperate deciduous broad-leaved forest V -Biomass and growth in natural forest of Cryptomeria japonica-. Bull. Kyoto Univ. For. 1988, 60, 67–76. (In Japanese) [Google Scholar]
- Tange, T.; Murakawa, I. Growth and Biomass of an 87-year-old Manmade Chamaecyparis obtuse Stand. Bull. Univ. Tokyo For. 1990, 82, 103–112. (In Japanese) [Google Scholar]
- Utsugi, H. Effects of Leaf Community Structure on Forest Canopy Photosynthetic Production in a Forest Community: Particularly on the Effect of Leaf Tilt Angle. Ph.D. Thesis, The University of Tokyo, Tokyo, Japan, 2009. (In Japanese). [Google Scholar]
- Daniel, S.F.; Remko, A.D.; Masae, I.I.; Diego, R.B.; Richard, G.F.; Angelica, V.; Masahiro, A.; Makoto, A.; Niels, A.; Michael, J.A.; et al. BAAD: A Biomass and Allometry Data Base for woody plants. Ecology 2015, 96, 1445. [Google Scholar] [CrossRef]
- Tange, T.; Kojima, K. Aboveground biomass data of Anno growth monitoring stands of Cryptomeria japonica in the University Forest in Chiba, The University of Tokyo. Misc. Inf. Univ. Tokyo For. 2010, 49, 1–6. (In Japanese) [Google Scholar]
- Sugimoto, T.; Ishii, H.; Chiba, Y.; Kanazawa, Y. Branch and Foliage Mass and their Vertical Distribution in a 90-year-old Chamaecyparis obtuse Plantation. J. Jpn. For. Soc. 2010, 92, 63–71. (In Japanese) [Google Scholar] [CrossRef]
- Karizumi, N. (Ed.) The Latest Illustrations of Tree Roots; Seibundo Shinkosha Publishing Co., Ltd.: Tokyo, Japan, 2010; ISBN 978-4-416-41005-9. (In Japanese) [Google Scholar]
- Tadaki, Y.; Hatiya, K.; Tochiaki, K. Studies on the Production Structure of Forest (XV) Primary Productivity of Fagus crenata in Plantation. J. Jpn. For. Soc. 1969, 51, 331–339. (In Japanese) [Google Scholar]
- Kotani, J. Estimation of standing crop and productivity of an even-aged young Buna (Fagus crenata) plantation. Bull. Ishikawa-Ken For. Exp. Stn. 2008, 40, 12–16. (In Japanese) [Google Scholar]
- Hoshi, H.; Tatsuhara, S.; Abe, N. Estimation of Leaf Area Index in Natural Deciduous Broad-leaved Forests Using Landsat TM Data. J. Jpn. For. Soc. 2001, 83, 315–321. (In Japanese) [Google Scholar]
- Kai, S. Dry-matter production of a young Konara oak (Quercus serrata Thunb.) plantation. Kyushu J. For. Res. 2011, 64, 130–131. (In Japanese) [Google Scholar]
- Katakura, M.; Koyama, Y. Existing and carbon stocks in larch, red pine, and Konara oak forests, and changes in soil carbon content after logging of red pine forests. Bull. Nagano Prefect. For. Res. Cent. 2007, 22, 33–55. (In Japanese) [Google Scholar]
- Ogasawara, R.; Yamamoto, T.; Arita, T. Biomass and Production of the Konara (Quercus serrata) Secondary Stand. Hardwood Res. 1987, 4, 257–262. (In Japanese) [Google Scholar]
- Hasegawa, M. Productivity of Oak Coppice Forests. J. Toyama For. For. Prod. Res. Cent. 1989, 2, 5–12. (In Japanese) [Google Scholar]
- Tadaki, Y.; Ogata, N.; Nagatomo, Y. The dry matter productivity in several stands of cryptomeria japonica in Kyushu. Bull. Gov. For. Exp. Stn. 1965, 173, 45–66. (In Japanese) [Google Scholar]
- Tsuruta, K.; Nogata, M.; Shinohara, Y.; Komatsu, H.; Otsuki, K. The correction coefficient for leaf area index measurement based on the optical method in a Japanese cedar forest. Bull. Kyusyu Univ. For. 2014, 95, 88–92. (In Japanese) [Google Scholar]
- Ogata, N.; Nagatomo, Y.; Kaminaka, S.; Tutumi, T. An example of analysis of production structure in a mature cypress forest at different growth status. Bull. Kyushu Branch Jpn. For. Soc. 1973, 26, 51–52. (In Japanese) [Google Scholar]
- Yamakura, T.; Saito, H.; Shidei, T. Production and structure of under-ground part of Hinoki (Chamaecyparis obtusa) stand (I) Estimation of root production by means of root analysis. J. Jpn. For. Soc. 1972, 54, 118–125. [Google Scholar]
- Miyamoto, M.; Tanimoto, T.; Ando, T. Analysis of the Growth of Hinoki (Chamaecyparis obtuse) Artificial Forests in Shikoku District. Bull. For. For. Prod. Res. Inst. 1980, 309, 89–107. (In Japanese) [Google Scholar]
- Kakubari, Y. Beech forests in the Naeba Mountains I. Distribution of primary productivity along the altitudinal gradient. In Primary Productivity in Japanese Forests; Shidei, T., Kira, T., Eds.; Japanese Committee for the International Biological Program, University of Tokyo Press, JIBP Synthesis: Tokyo, Japan, 1977; Volume 16, pp. 201–212. [Google Scholar]
- Maruyama, K. Effect of altitude on dry-matter production of primeval Japanese beech forest communities in Naeba Mountains. Mem. Fac. Agric. Niigata Univ. 1971, 9, 85–171. [Google Scholar]
- Okumura, T.; Tanaka, K.; Moriishi, M. Measurement of Transpiration by Oak Trees with the Butt Immersed in the Water. Hardwood Res. 1987, 4, 119–128. (In Japanese) [Google Scholar]
- Nagai, A. Estimation of pan evaporation by Makkink equation. J. Jpn. Soc. Hydrol. Water Resour. 1993, 6, 237–243. (In Japanese) [Google Scholar] [CrossRef]
- Kondo, J. Dependence of evapotranspiration on the precipitation amount and leaf area index for various vegetated surfaces. J. Jpn. Soc. Hydrol. Water Resour. 1998, 11, 679–693. (In Japanese) [Google Scholar] [CrossRef]
- Kondo, J.; Watanabe, T. A guide to study on evaporation from the complex land surface. Tenki 1991, 38, 699–710. (In Japanese) [Google Scholar]
- Kondo, J.; Ishii, M. Estimation of rainfall interception loss from forest canopies and comparison with measurements. J. Jpn. Soc. Hydrol. Water Resour. 1992, 5, 27–34. (In Japanse) [Google Scholar] [CrossRef]
- Kondo, J.; Watanabe, T.; Nakazono, M.; Ishii, M. Estimation of forest rainfall interception. Tenki 1992, 39, 159–167. (In Japanese) [Google Scholar]
- Ebisu, N.; Takase, K.; Otake, N. Construction of forest hydrological tree form model for Japanese cedar and cypress trees. J. Jpn. Soc. Eros. Control Eng. 2015, 68, 25–31. (In Japanese) [Google Scholar]
- Leblanc, S.G.; Chen, J.M.; Fernandes, R.; Deering, D.W.; Conley, A. Methodology comparison for canopy structure parameters extraction from digital hemispherical photography in boreal forests. Agric. For. Meteorol. 2005, 129, 187–207. [Google Scholar] [CrossRef]
- Ena Regional Agriculture and Forestry Office of Gifu Prefecture and Japan Forest Engineering Consultants. Report of the Kiso River Basin Forest Hydrologic Function Study -Futatsumori, Fukuoka, Nakatsugawa City, Gifu Prefecture-; Ena Regional Agriculture and Forestry Office of Gifu Prefecture and Japan Forest Engineering Consultants: Gifu, Japan, 2008. (In Japanese)
- Greenhouse Gas Inventory Office of Japan (Ed.) National Greenhouse Gas Inventory Report of JAPAN; Ministry of the Environment: Tokyo, Japan, 2022; pp. 6-12–6-29. ISSN 2434-5679.
- Vertessy, R.A.; Watson, F.G.R.; O’sullivan, S.K. Factors determining relations between stand age and catchment water balance in mountain ash forests. For. Ecol. Manag. 2001, 143, 13–26. [Google Scholar] [CrossRef]
- Kojima, T. Relationship between forest stand condition and water balance in a forested basin. In River Basin Environment: Evaluation, Management and Conservation; Li, F., Awaya, Y., Kageyama, K., Wei, Y., Eds.; Springer: Singapore, 2022; pp. 231–258. [Google Scholar]
- Biodiversity Center of Japan, Natural Environmental Information GIS, Japan Integrated Biodiversity Information System. Available online: https://www.biodic.go.jp/kiso/fnd_f.html (accessed on 25 December 2023).
- Suzuki, M.; Tange, T.; Suzuki, T.; Suzuki, S. Growth and biomass of manmade Zelkova serrata stands in Tokyo University Forest in Chiba. Bull. Tokyo Univ. For. 1990, 82, 113–129. (In Japanese) [Google Scholar]
- Kumagai, S. Estimation of Surface Area of AKAMATSU Foliage. Rep. Kyushu Univ. For. 1962, 16, 1–8. (In Japanese) [Google Scholar]
- Fellner, H.; Dirnberger, G.F.; Sterba, H. Specific leaf area of European Larch (Larix decidua Mill.). Trees 2016, 30, 1237–1244. [Google Scholar] [CrossRef]
- Satoo, T.; Negisi, K.; Senda, M. Materials for the studies of growth in stands. V. Amount of leaves and growth in plantations of Zelkova serrata applied with crown thinning. Bull. Tokyo Univ. For. 1959, 55, 101–123. (In Japanese) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








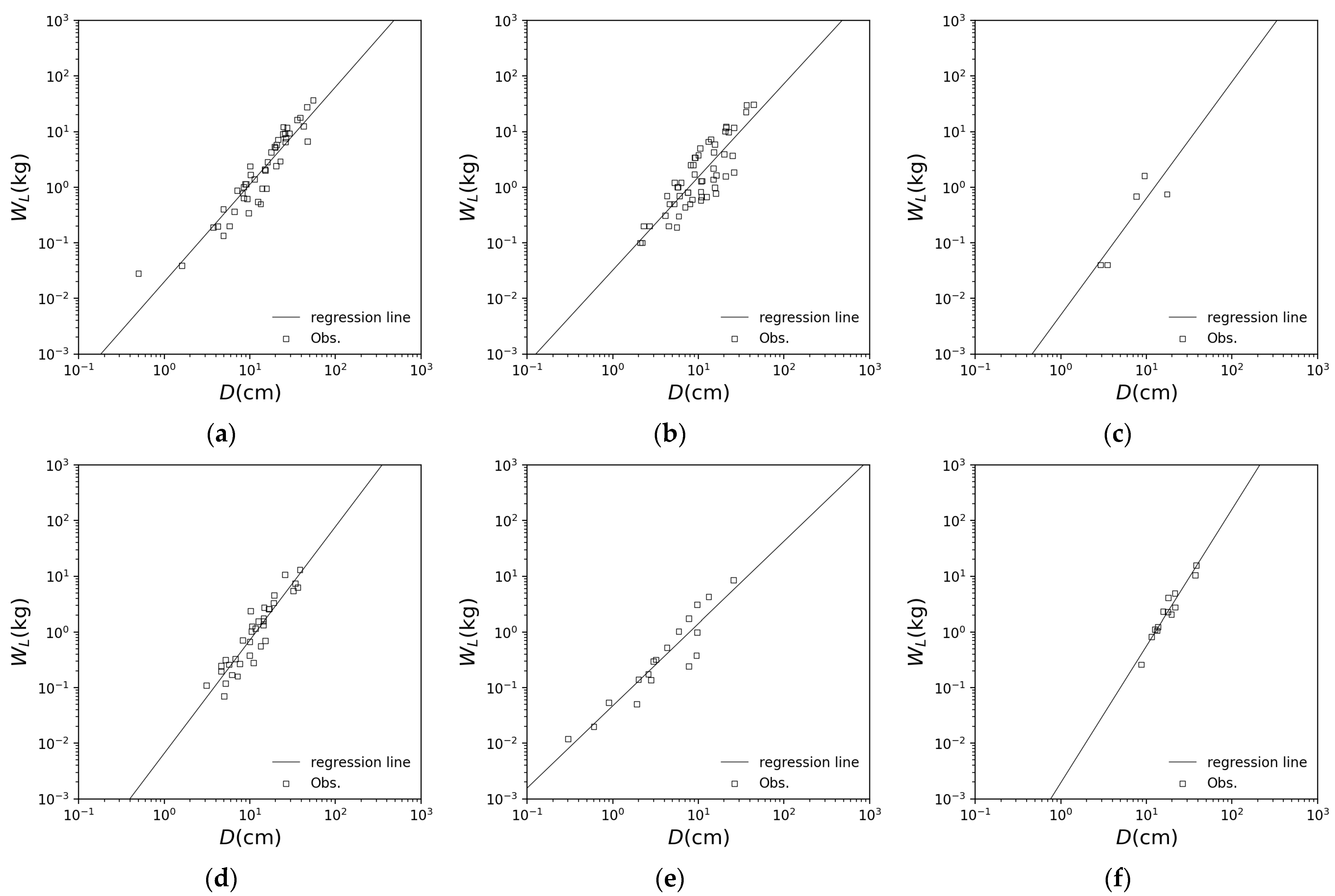
| Beech | Konara Oak | Birch | |
|---|---|---|---|
| 1.854 | 1.974 | 2.278 | |
| −1.883 | −1.659 | −2.290 | |
| 0.958 | 0.858 | 0.889 |
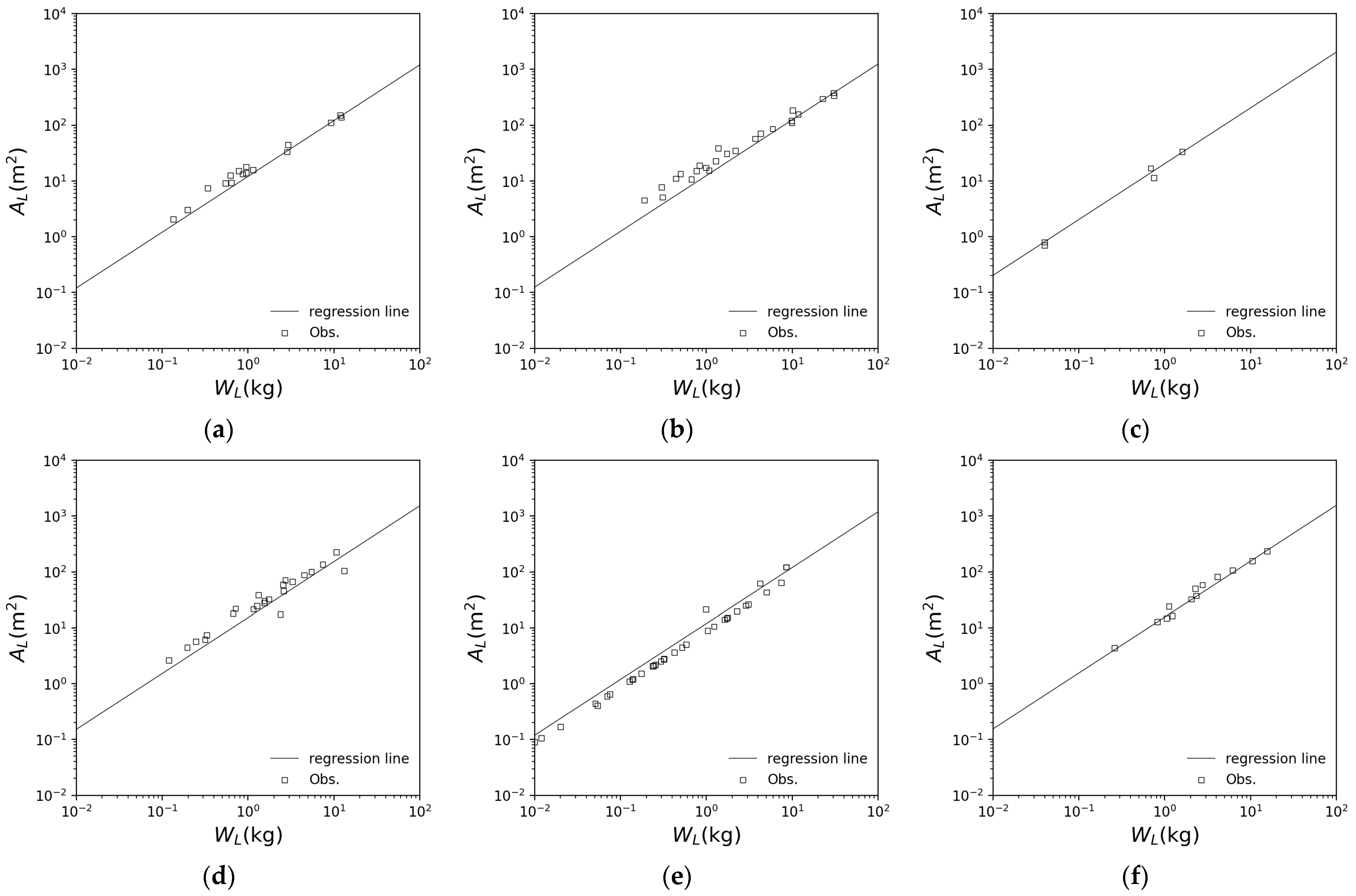
| Cedar | Cypress | Beech | Konara Oak | Birch | |
|---|---|---|---|---|---|
| 2.897 | 3.666 | 20.77 | 11.90 | 17.62 | |
| 0.995 | 0.993 | 0.998 | 0.992 | 0.973 |
| Cedar | Cypress | Beech | Oak | Birch | ||
|---|---|---|---|---|---|---|
| 1.57 | 1.55 | 1.58 | 1.40 | 1.31 | ||
| 1.23 | 1.24 | 1.32 | 1.26 | 1.20 | ||
| 0.25 | 0.26 | 0.26 | 0.26 | 0.26 | ||
| 0.314 | 0.407 | 0.573 | 0.624 | 0.438 | ||
| 0.51 | 0.51 | 0.48 | 0.48 | 0.48 | ||
| 2016 | 2017 | 2018 | 2019 | 2020 | |
|---|---|---|---|---|---|
| NSE | 0.875 | 0.774 | 0.823 | 0.848 | 0.848 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kojima, T.; Shimono, R.; Ota, T.; Hashimoto, H.; Hasegawa, Y. Development of a Model to Evaluate Water Conservation Function for Various Tree Species. Water 2024, 16, 588. https://doi.org/10.3390/w16040588
Kojima T, Shimono R, Ota T, Hashimoto H, Hasegawa Y. Development of a Model to Evaluate Water Conservation Function for Various Tree Species. Water. 2024; 16(4):588. https://doi.org/10.3390/w16040588
Chicago/Turabian StyleKojima, Toshiharu, Ryoma Shimono, Takahiro Ota, Hiroshi Hashimoto, and Yasuhiro Hasegawa. 2024. "Development of a Model to Evaluate Water Conservation Function for Various Tree Species" Water 16, no. 4: 588. https://doi.org/10.3390/w16040588
APA StyleKojima, T., Shimono, R., Ota, T., Hashimoto, H., & Hasegawa, Y. (2024). Development of a Model to Evaluate Water Conservation Function for Various Tree Species. Water, 16(4), 588. https://doi.org/10.3390/w16040588