



The Impact of Vegetation Types on Soil Hydrological and Mechanical Properties in the Hilly Regions of Southern China: A Comparative Analysis


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
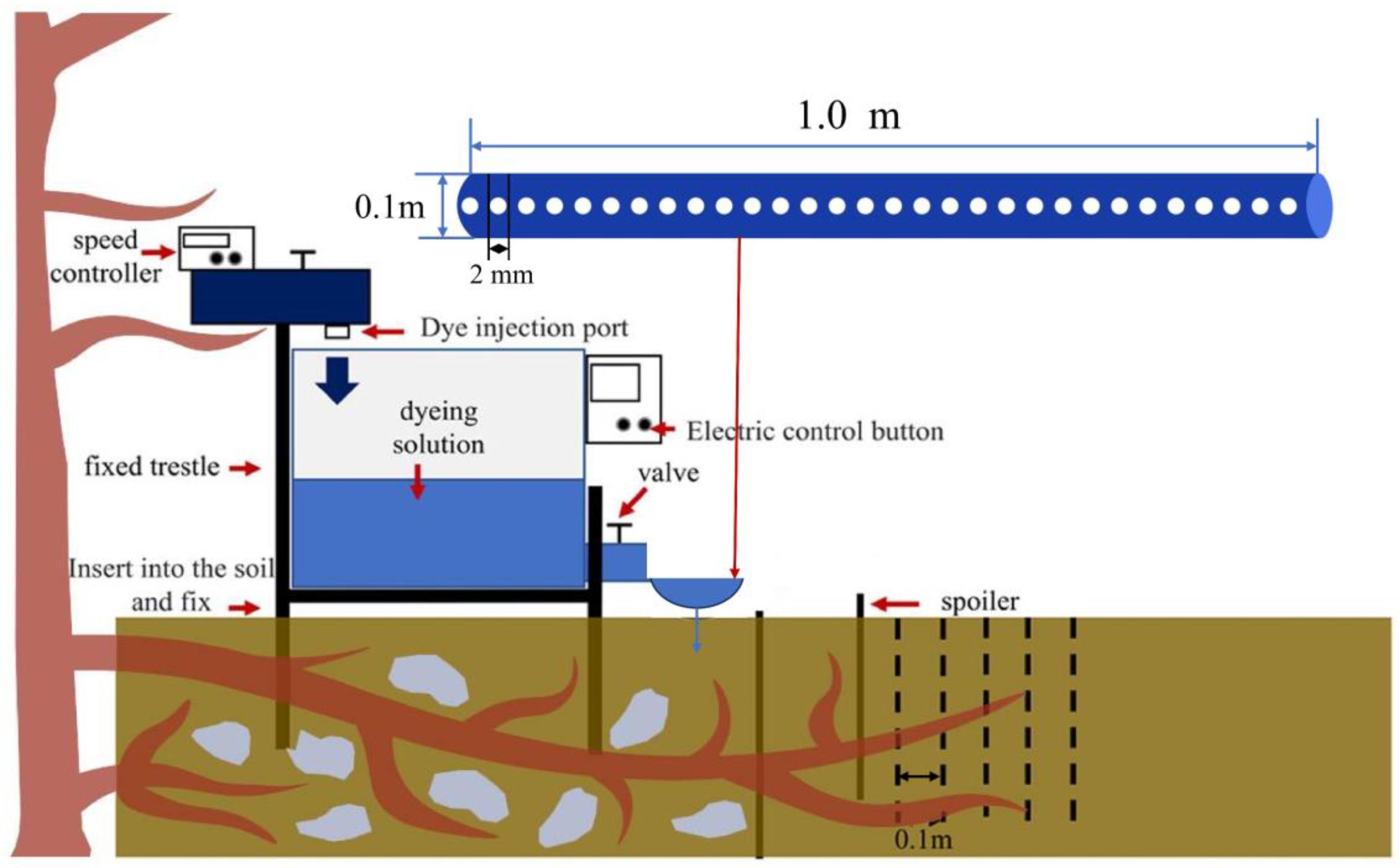
2.2. Experimental Design and Sample Collection
2.3. Laboratory Test
2.4. Data Analysis
3. Results
3.1. Soil Root Distribution
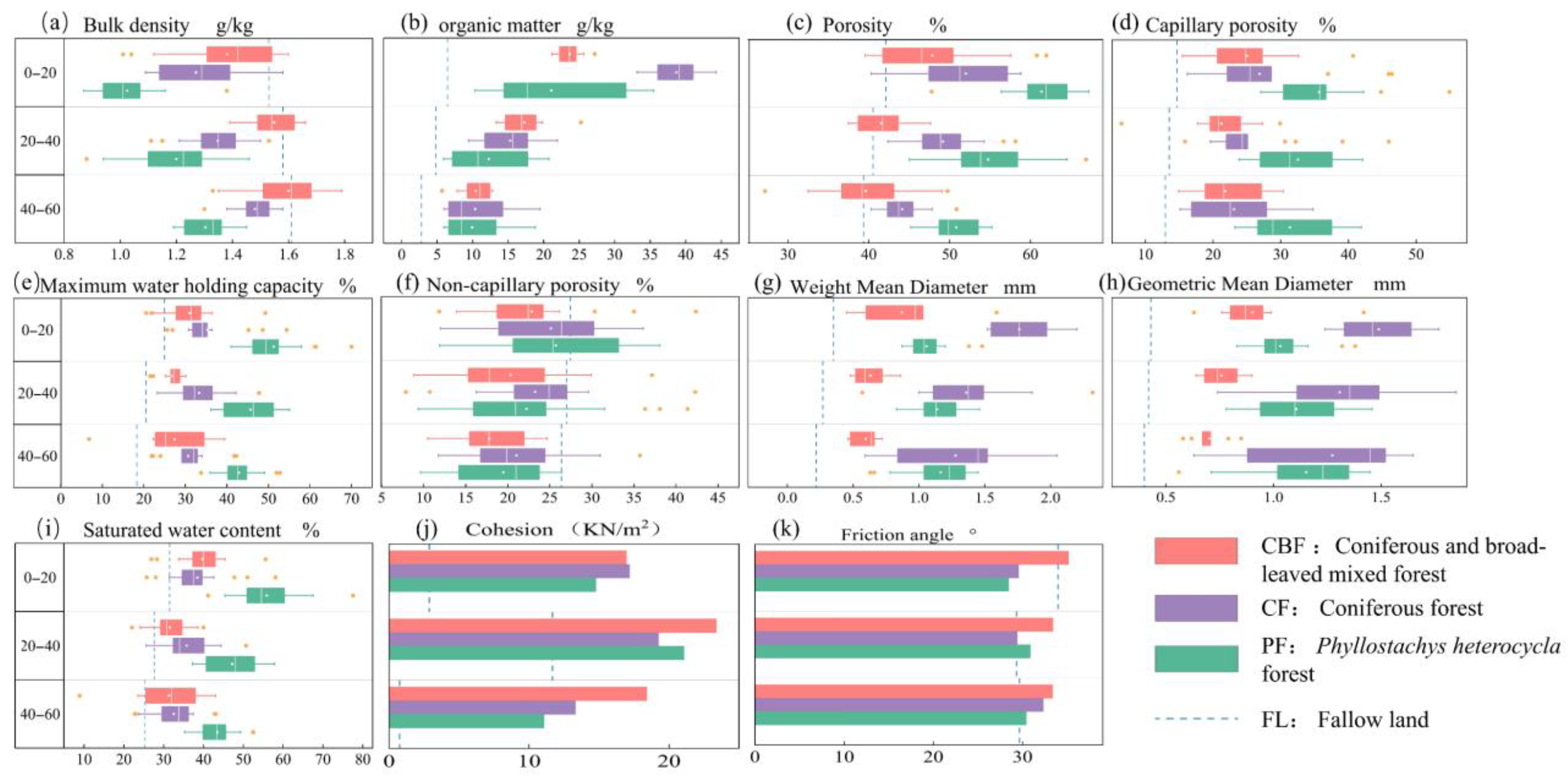
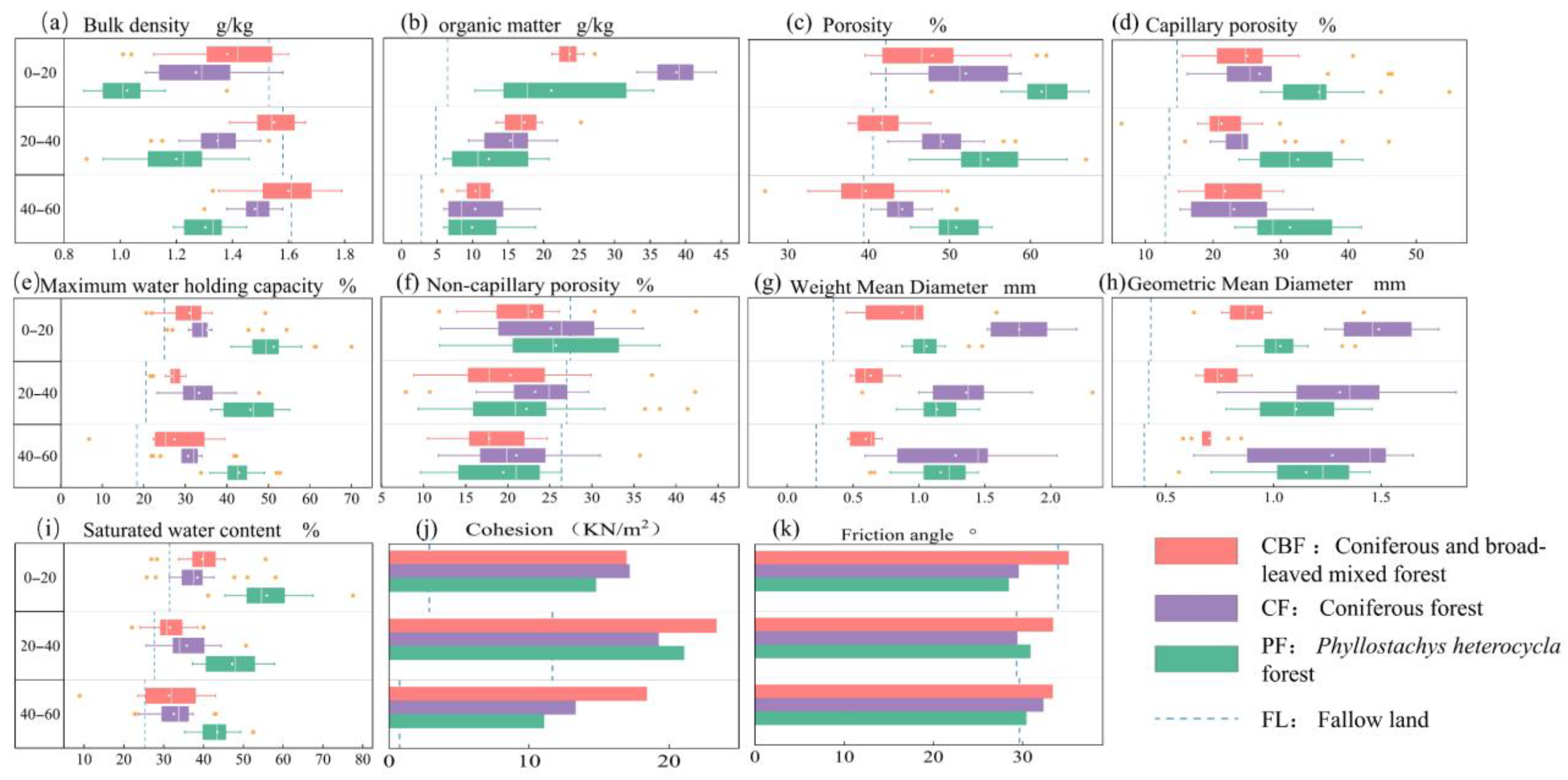
3.2. Soil Physical and Chemical Characteristics
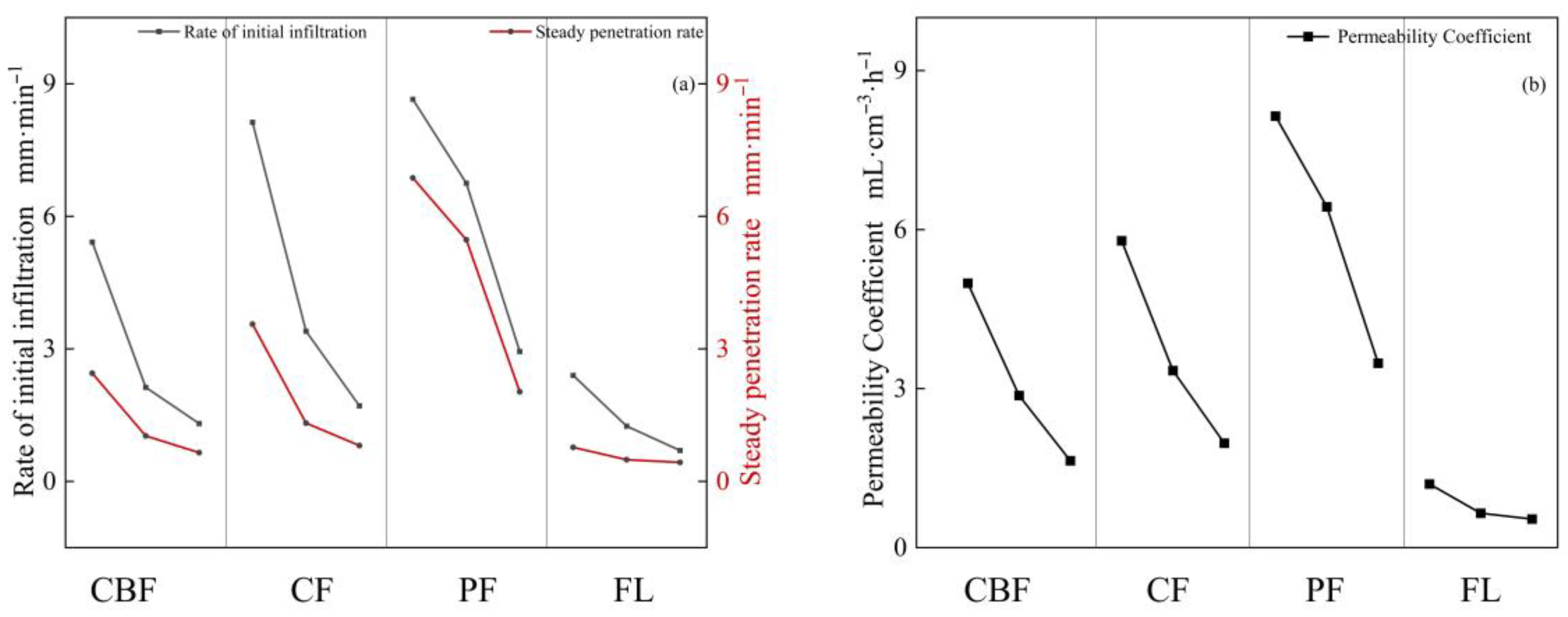
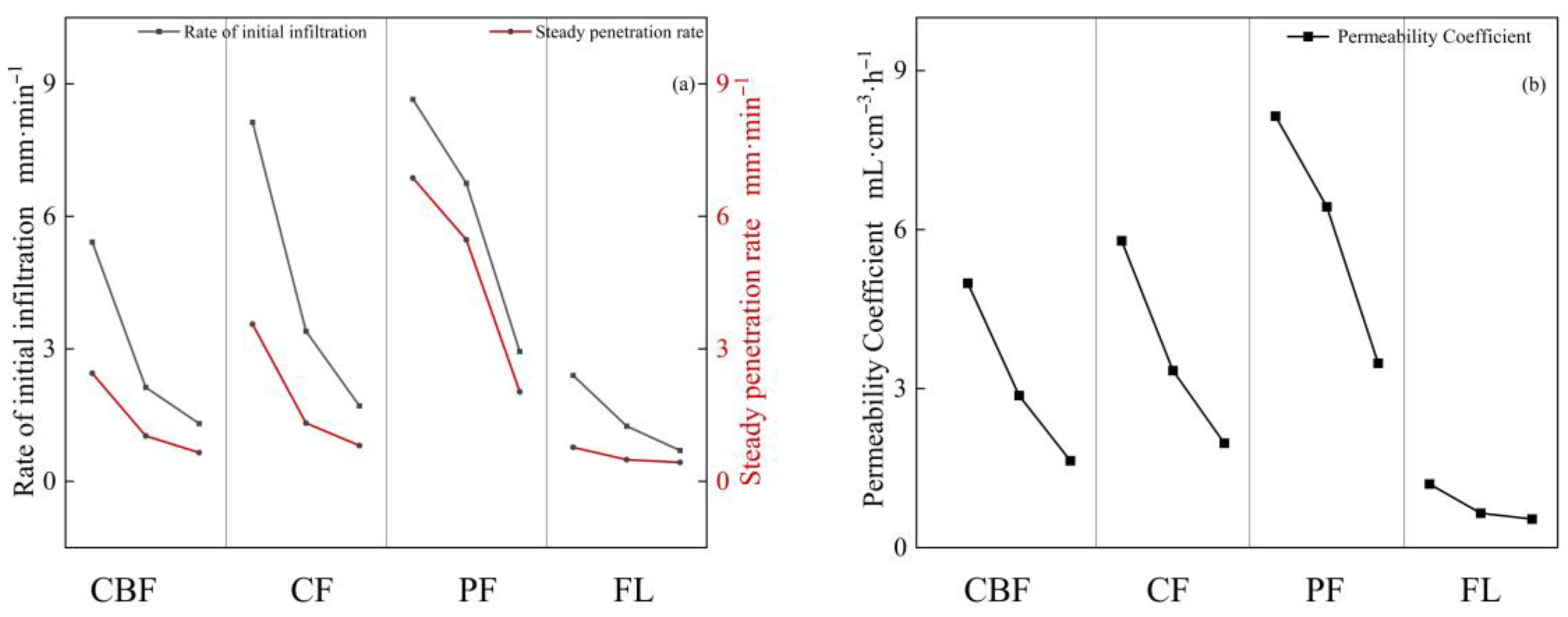
3.3. Characteristics of Soil Infiltration
3.4. Soil Aggregates’ Stability and Shear Strength
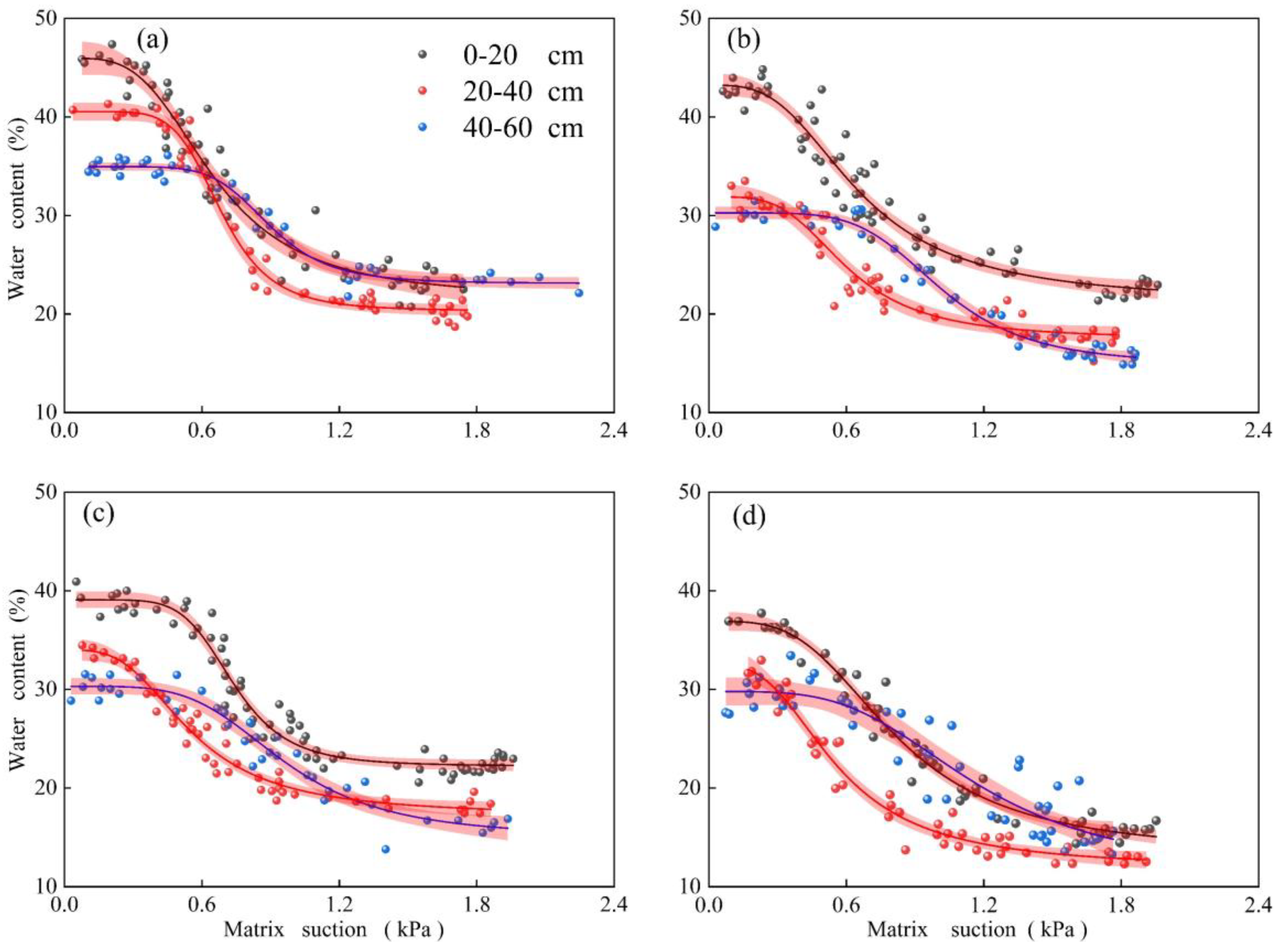
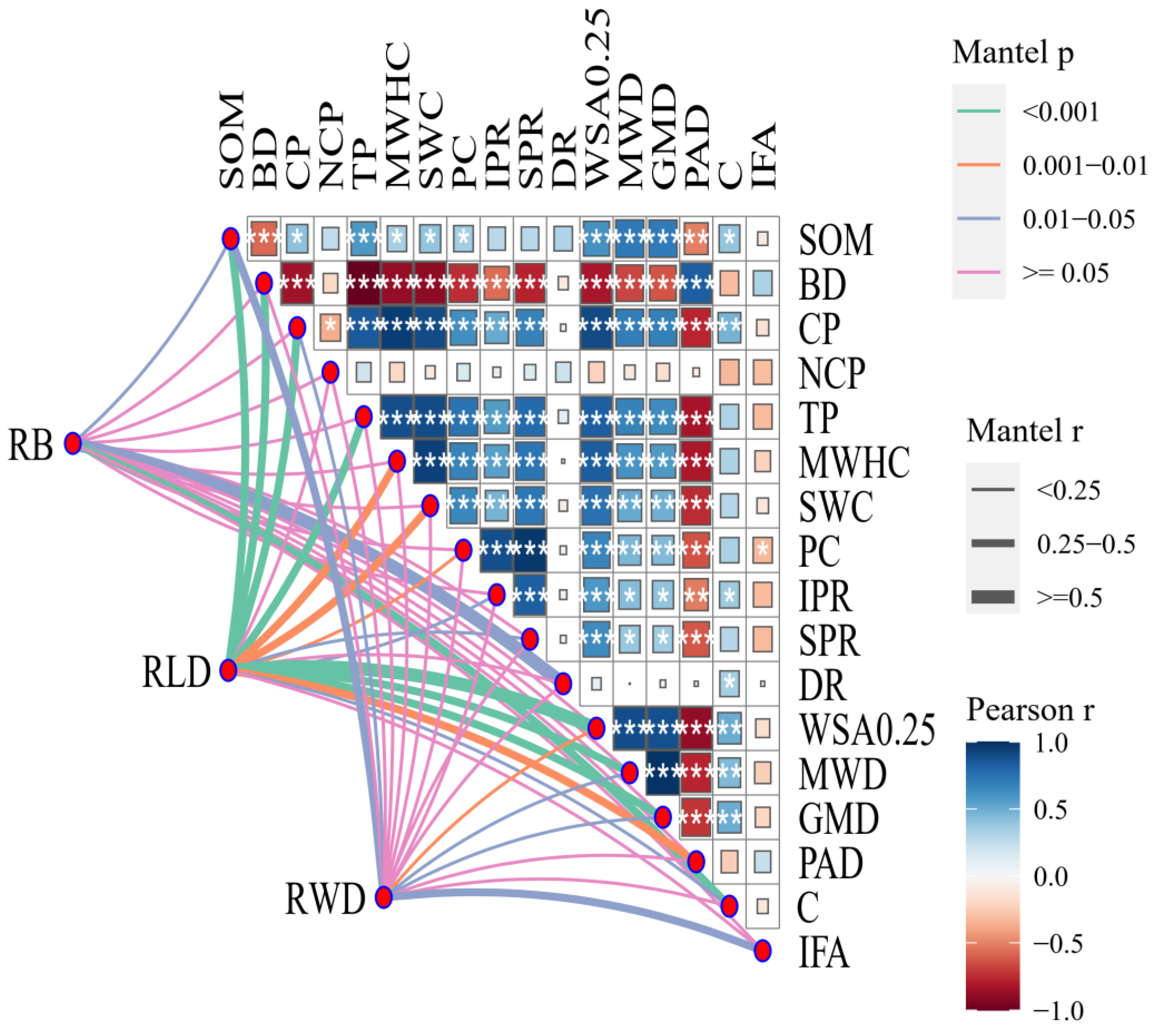
3.5. Analysis of Soil Water Distribution
4. Discussion
4.1. Variation in Soil Aggregate and Shear Strength
4.2. Multiple Effects of Plants on Soil Water Distribution
4.3. Research Limitations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Alvarez, R.; Steinbach, H.S. A review of the effects of tillage systems on some soil physical properties, water content, nitrate availability and crops yield in the Argentine Pampas. Soil Tillage Res. 2009, 104, 1–15. [Google Scholar] [CrossRef]
- Barzegar, A.R.; Hashemi, A.M.; Herbert, S.J.; Asoodar, M.A. Interactive effects of tillage system and soil water content on aggregate size distribution for seedbed preparation in Fluvisols in southwest Iran. Soil Tillage Res. 2004, 78, 45–52. [Google Scholar] [CrossRef]
- Deljouei, A.; Cislaghi, A.; Abdi, E.; Borz, S.A.; Majnounian, B.; Hales, T.C. Implications of hornbeam and beech root systems on slope stability: From field and laboratory measurements to modelling methods. Plant Soil 2023, 483, 547–572. [Google Scholar] [CrossRef]
- Norris, J.E.; Di Iorio, A.; Stokes, A.; Nicoll, B.C.; Achim, A. Species selection for soil reinforcement and protection. In Slope Stability and Erosion Control: Ecotechnological Solutions; Springer: Berlin/Heidelberg, Germany, 2008; pp. 167–210. [Google Scholar] [CrossRef]
- Demenois, J.; Rey, F.; Stokes, A.; Carriconde, F. Does arbuscular and ectomycorrhizal fungal inoculation improve soil aggregate stability? A case study on three tropical species growing in ultramafic Ferralsols. Pedobiologia 2017, 64, 8–14. [Google Scholar] [CrossRef]
- Osman, N.; Barakbah, S.S. The effect of plant succession on slope stability. Ecol. Eng. 2011, 37, 139–147. [Google Scholar] [CrossRef]
- Faxing, S.; Chongjun, T.; Jichao, Z.; Ronggang, Y.; Taihui, Z.; Dekui, N. Water erosion control of undisturbed soil cores by near s-oil surface factors after 5-year vegetation restoration in red sandstone area from subtropical China. J. Soil Sediment 2023, 23, 1356–1369. [Google Scholar] [CrossRef]
- Karimi, Z.; Abdi, E.; Deljouei, A.; Cislaghi, A.; Shirvany, A.; Schwarz, M.; Hales, T.C. Vegetation-induced soil stabilization in coastal area: An example from a natural mangrove forest. CATENA 2022, 216, 106410. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Jia, H.; Liu, Y.; Zhao, Y.; Shi, C.; Zhang, F.; Wang, K. Assessing the soil moisture effects of planted vegetation on slope stability in shallow land slide-prone areas. J. Soil Sediment 2021, 21, 2551–2565. [Google Scholar] [CrossRef]
- Grose, P. Composted soil conditioner and mulch promote native plant establishment from seed in a constructed seasonal wetland complex. Ecol. Manag. Restor. 2011, 12, 151–154. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Doe. Precipitation Partitioning by Vegetation; Springer International Publishing: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]
- Ball, B.C.; Bingham, I.; Rees, R.M.; Watson, C.A.; Litterick, A. The role of crop rotations in determining soil structure and crop growth conditions. Can. J. Soil Sci. 2005, 85, 557–577. [Google Scholar] [CrossRef]
- Adams, H.D.; Luce, C.H.; Breshears, D.D.; Allen, C.D.; Weiler, M.; Hale, V.C.; Smith, A.M.S.; Huxman, T.E. Ecohydrological consequences of drought- and infestation- triggered tree die-off: Insights and hypotheses. Ecohydrology 2012, 5, 145–159. [Google Scholar] [CrossRef]
- Bengough, A.G.; Croser, C.; Pritchard, J. A biophysical analysis of root growth under mechanical stress. Plant Soil 1997, 189, 107–116. [Google Scholar] [CrossRef]
- Dexter, A.R. Compression of soil around roots. Plant Soil 1987, 97, 401–406. [Google Scholar] [CrossRef]
- Zhou, B.Z.; Zhang, S.G.; Fu, M.Y. Minirhizotron, a new technique for plant root system research: Its invention, development and application. Chin. J. Ecol. 2007, 26, 253. [Google Scholar]
- Grayston, S.J.; Vaughan, D.; Jones, D. Rhizosphere carbon flow in trees, in comparison with annual plants: The importance of root exudation and its impact on microbial activity and nutrient availability. Appl. Soil Ecol. 1997, 5, 29–56. [Google Scholar] [CrossRef]
- Traore, O.; Groleau-Renaud, V.; Plantureux, S.; Tubeileh, A.; Boeuf-Tremblay, V. Effect of root mucilage and modelled root exudates on soil structure. Eur. J. Soil Sci. 2000, 51, 575–581. [Google Scholar] [CrossRef]
- Fu, Z.; Hu, W.; Beare, M.; Thomas, S.; Carrick, S.; Dando, J.; Langer, S.; Muller, K.; Baird, D.; Lilburne, L. Land use effects on soil hydraulic properties and the contribution of soil organic carbon. J. Hydrol. 2021, 602, 13. [Google Scholar] [CrossRef]
- Jiang, X.J.; Liu, W.; Chen, C.; Liu, J.; Yuan, Z.; Jin, B.; Yu, X. Effects of three morphometric features of roots on soil water flow behavior in three sites in China. Geoderma 2018, 320, 161–171. [Google Scholar] [CrossRef]
- Gong, Y.W.; Yu, H.J.; Tian, P.; Guo, W.Z.; Chen, L.; Shen, D.T. Field experiments on quantifying the contributions of Coreopsis canopies and roots to controlling runoff and erosion on steep loess slopes. J. Mt. Sci. 2023, 20, 1402–1423. [Google Scholar] [CrossRef]
- Farahnak, M.; Mitsuyasu, K.; Hishi, T.; Katayama, A.; Chiwa, M.; Jeong, S.; Kume, A. Relationship between very fine root distribution and soil water content in pre-and post-harvest areas of two coniferous tree species. Forests 2020, 11, 1227. [Google Scholar] [CrossRef]
- Materechera, S.A.; Alston, A.M.; Kirby, J.M.; Dexter, A.R. Field evaluation of laboratory techniques for predicting the ability of roots to penetrate strong soil and of the influence of roots on water sorptivity. Plant Soil 1993, 149, 149–158. [Google Scholar] [CrossRef]
- Wu, G.; Yang, Z.; Cui, Z.; Liu, Y.; Fang, N.; Shi, Z. Mixed artificial grasslands with more roots improved mine soil infiltration capacity. J. Hydrol. 2016, 535, 54–60. [Google Scholar] [CrossRef]
- Jiang, X.J.; Chen, C.; Zhu, X.; Zakari, S.; Singh, A.K.; Zhang, W.; Zeng, H.; Yuan, Z.; He, C.; Yu, S.; et al. Use of dye infiltration experiments and HYDRUS-3D to interpret preferential flow in soil in a rubber-based agroforestry systems in Xishuangbanna, China. CATENA 2019, 178, 120–131. [Google Scholar] [CrossRef]
- Czarnes, S.; Hiller, S.; Dexter, A.R.; Hallett, P.D.; Bartoli, F. Root: Soil adhesion in the maize rhizosphere: The rheological approach. Plant Soil 1999, 211, 69–86. [Google Scholar] [CrossRef]
- Nciizah, A.D.; Wakindiki, I.I.C. Physical indicators of soil erosion, aggregate stability and erodibility. Arch. Agron. Soil Sci. 2015, 61, 827–842. [Google Scholar] [CrossRef]
- Ghorbani, M.; Amirahmadi, E.; Neugschwandtner, R.W.; Konvalina, P.; Kopecky, M.; Moudry, J.; Perna, K.; Murindangabo, Y.T. The impact of pyrolysis temperature on biochar properties and its effects on soil hydrological properties. Sustainability 2022, 14, 14722. [Google Scholar] [CrossRef]
- Wei, H.; Deng, Y.; Huang, J.; He, L.; Tang, Q.; Xiao, Y. A quantitative study of the influence of soil organic carbon and pore characteristics on the stability of aggregates of the karst peak-cluster depression area in Southwest China. J. Soil Sediment. 2023, 23, 312–330. [Google Scholar] [CrossRef]
- Zuazo, V.H.D.; Pleguezuelo, C.R.R. Soil-erosion and runoff prevention by plant covers. A review. Agron. Sustain. Dev. 2008, 28, 785–811. [Google Scholar] [CrossRef]
- Novotný, J.; Klimeš, J. Grain size distribution of soils within the Cordillera Blanca, Peru: An indicator of basic mechanical properties for slope stability evaluation. J. Mt. Sci. 2014, 11, 563–577. [Google Scholar] [CrossRef]
- Alderfer, R.B. Influence of seasonal and cultural conditions on aggregation of Hagerstown soil. Soil Sci. 1950, 69, 93–204. [Google Scholar] [CrossRef]
- He, Y.F.; Xie, H.L.; Peng, C.Z. Analyzing the behavioural mechanism of farmland abandonment in the hilly mountainous areas in China from the perspective of farming household diversity. Land Use Policy 2020, 99, 104826. [Google Scholar] [CrossRef]
- Xu, H.; Song, Y.; Tian, Y. Simulation of land-use pattern evolution in hilly mountainous areas of North China: A case study in Jincheng. Land Use Policy 2022, 112, 105826. [Google Scholar] [CrossRef]
- ISO N. 11465; Soil quality- Determination of pH. International Organization for Standardization (ISO): Geneva, Switzerland, 1994.
- Nespoulous, J.; Merino-Martín, L.; Monnier, Y. Tropical forest structure and understorey determine subsurface flow through biopores formed by plant roots. Catena 2019, 181, 104061. [Google Scholar] [CrossRef]
- Cao, S.; Zhou, Y.; Zhou, Y.; Zhou, X.; Zhou, W. Soil organic carbon and soil aggregate stability as sociated with aggregate fractions in a chronosequence of citrus or chards plantations. J. Environ. Manag. 2021, 293, 8. [Google Scholar] [CrossRef]
- Wei, J.; Shi, B.L.; Li, J.L.; Li, S.S.; He, X.B. Shear strength of purple soil bunds under different soil water contents and dry densities: A case study in the Three Gorges Reservoir Area, China. Catena 2018, 166, 124–133. [Google Scholar] [CrossRef]
- Zhang, G.L.; Gong, Z.T. Soil Survey Laboratory Methods; China Science Publishing & Media Ltd.: Beijing, China, 2012. [Google Scholar]
- Verbist, K.; Torfs, S.; Cornelis, W.M. Comparison of single-and double-ring infiltrometer methods on stony soils. Vadose Zone J. 2010, 9, 462–475. [Google Scholar] [CrossRef]
- Elliott, E.T. Aggregate Structure and Carbon, Nitrogen, and Phosphorus in Native and Cultivated Soils. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Zhu, G.; Shangguan, Z.; Deng, L. Variations in soil aggregate stability due to land use changes from agricultural land on the Loess Plateau, China. CATENA 2021, 200, 105181. [Google Scholar] [CrossRef]
- Defossez, P.; Veylon, G.; Yang, M.; Bonnefond, J.M.; Garrigou, D.; Trichet, P.; Danjon, F. Impact of soil water content on the overturning resistance of young Pinus Pinaster in sandy soil. For. Ecol. Manag. 2021, 480, 118614. [Google Scholar] [CrossRef]
- Van Genuchten, M.T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- Kent, M.G.; Schiavon, S.; Jakubiec, J.A. A dimensionality reduction method to select the most representative daylight illuminance distributions. J. Build. Perform. Simul. 2020, 13, 122–135. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, Y.H. Theoretical analysis of illumination in PCA-based vision systems. Pattern Recognit. 1999, 32, 547–564. [Google Scholar] [CrossRef]
- Zhao, Q.G.; Huang, G.Q.; Ma, Y.Q. The problems in red soil ecosystem in southern of China and its countermeasures. Acta Ecol. Sin. China 2013, 33, 7615–7622. [Google Scholar]
- Fattet, M.; Fu, Y.; Ghestem, M.; Ma, W.; Foulonneau, M.; Nespoulous, J.; Le Bissonnais, Y.; Stokes, A. Effects of vegetation type on soil resistance to erosion: Relationship between aggregate stability and shear strength. Catena 2011, 87, 60–69. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, J.; Liu, J.; Yu, M.; Geng, W.; Wang, P.; Wu, Z. Relationships between shear strength parameters and microstructure of alkaline-contaminated red clay. Environ. Sci. Pollut. Res. 2020, 27, 33848–33862. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhong, X.; Lin, J.; Zhao, D.; Jiang, F.; Wang, M.; Ge, H.; Huang, Y. Effects of fractal dimension and water content on the shear strength of red soil in the hilly granitic region of southern China. Geomorphology 2020, 360, 1. [Google Scholar] [CrossRef]
- Chong-Feng, B.U.; Gale, W.J.; Qiang-Guo, C.A.I.; Shu-Fang, W.U. Process and mechanism for the development of physical crusts in three typical Chinese soils. Pedosphere 2013, 23, 321–332. [Google Scholar] [CrossRef]
- Liu, J.; Shi, B.; Jiang, H.; Huang, H.; Wang, G.; Kamai, T. Research on the stabilization treatment of clay slope topsoil by organic polymer soil stabilizer. Eng. Geol. 2011, 117, 114–120. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, X.; Yu, D.M.; Zhu, H.L.; Li, G.R. Influence of the roots of mixed-planting species on the shear strength of saline loess soil. J. Mt. Sci. 2021, 18, 806–818. [Google Scholar] [CrossRef]
- Duchicela, J.; Sullivan, T.S.; Bontti, E.; Bever, J.D. Soil aggregate stability increase is strongly related to fungal community succession along an abandoned agricultural field chronosequence in the Bolivian Altiplano. J. Appl. Ecol. 2013, 50, 1266–1273. [Google Scholar] [CrossRef]
- Zhang, Y.; Niu, J.; Zhang, M.; Xiao, Z.; Zhu, W. Interaction between plant roots and soil water flow in response to preferential flow paths in northern China. Land Degrad. Dev. 2017, 28, 648–663. [Google Scholar] [CrossRef]
- Luna, L.; Miralles, I.; Andrenelli, M.C.; Gispert, M.; Pellegrini, S.; Vignozzi, N.; Sole-Benet, A. Restoration techniques affect soil organic carbon, glomalin and aggregate stability in degraded soils of a semiarid Mediterranean region. Catena 2016, 143, 256–264. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Jiang, W.; Li, Z.; Xie, H.; Ouyang, K.; Yuan, H.; Duan, L. Land use change impacts on red slate soil aggregates and associated organic carbon in diverse soil layers in subtropical China. Sci. Total Environ. 2023, 856, 11. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Stone, L.F. Physical, Chemical, and Biological Changes in the Rhizosphere and Nutrient Availability. J. Plant Nutr. 2006, 29, 1327–1356. [Google Scholar] [CrossRef]
- Larsbo, M.; Koestel, J.; Kotterer, T.; Jarvis, N. Preferential Transport in Macropores is Reduced by Soil Organic Carbon. Vadose Zone J. 2016, 15, 7. [Google Scholar] [CrossRef]
- Angers, D.A. Changes in soil aggregation and organic-carbon under corn and alfalfa. Soil Sci. Soc. Am. J. 1992, 56, 1244–1249. [Google Scholar] [CrossRef]
- Pohl, M.; Alig, D.; Koerner, C.; Rixen, C. Higher plant diversity enhances soil stability in disturbed alpine ecosystems. Plant Soil 2009, 324, 91–102. [Google Scholar] [CrossRef]
- Tan, H.; Chen, F.; Chen, J.; Gao, Y. Direct shear tests of shear strength of soils reinforced by geomats and plant roots. Geotext. Geomembr. 2019, 47, 780–791. [Google Scholar] [CrossRef]
- Ali, F.H.; Osman, N. Shear strength of a soil containing vegetation roots. Soils Found. 2008, 48, 587–596. [Google Scholar] [CrossRef]
- John, B.; Pandey, H.N.; Tripathi, R.S. Vertical distribution and seasonal changes of fine and coarse root mass in Pinus kesiya Royle Ex. Gordon forest of three different ages. Acta Oecol. 2001, 22, 293–300. [Google Scholar] [CrossRef]
- Schmidt, K.M.; Roering, J.J.; Stock, J.D.; Dietrich, W.E.; Montgomery, D.R.; Schaub, T. The variability of root cohesion as an influence on shallow landslide susceptibility in the Oregon Coast Range. Can. Geotech. J. 2001, 38, 995–1024. [Google Scholar] [CrossRef]
- Kemper, W.D.; Rosenau, R.C.; Dexter, A.R. Cohesion development in disrupted soils as affected by clay and organic-matter content and temperature. Soil Sci. Soc. Am. J. 1987, 51, 860–867. [Google Scholar] [CrossRef]
- Wang, J.; Feng, S.; Ni, S.; Wen, H.; Cai, C.; Guo, Z. Soil detachment by overland flow on hillslopes with permanent gullies in the Granite area of southeast China. Catena 2019, 183, 9. [Google Scholar] [CrossRef]
- Sun, Y.; Li, H.; Cheng, Z.; Dong, J.; Wang, Y. Experimental and Numerical Simulation Study on Mechanical Properties of Shallow Slope Root-soil Composite in Qinghai Area. KSCE J. Civ. Eng. 2023, 27, 2834–2852. [Google Scholar] [CrossRef]
- Belnap, J. The potential roles of biological soil crusts in dryland hydrologic cycles. Hydrol. Process. 2006, 20, 3159–3178. [Google Scholar] [CrossRef]
- Skorobogatov, A.; He, J.; Chu, A.; Valeo, C.; van Duin, B. The impact of media, plants and their interactions on bioretention performance: A review. Sci. Total Environ. 2020, 715, 14. [Google Scholar] [CrossRef]
- Quesada, C.A.; Lloyd, J.; Schwarz, M.; Baker, T.R.; Phillips, O.L.; Patino, S.; Czimczik, C.; Hodnett, M.G.; Herrera, R.; Arneth, A.; et al. Regional and large-scale patterns in Amazon forest structure and function are mediated by variations in soil physical and chemical properties. Biogeosci. Discuss. 2009, 6, 3993–4057. [Google Scholar] [CrossRef]
- Cosby, B.J.; Hornberger, G.M.; Clapp, R.B.; Ginn, T.R. A statistical exploration of the relationships of soil-moisture characteristics to the physical-properties of soils. Water Resour. Res. 1984, 20, 682–690. [Google Scholar] [CrossRef]
- Alaoui, A.; Lipiec, J.; Gerke, H.H. A review of the changes in the soil pore system due to soil deformation: A hydrodynamic perspective. Soil Tillage Res. 2011, 115, 1–15. [Google Scholar] [CrossRef]
- Pulido, M.; Schnabel, S.; Contador, J.F.L.; Lozano-Parra, J.; Gomez-Gutierrez, A.; Brevik, E.C.; Cerda, A. Reduction of the frequency of herbaceous roots as an effect of soil compaction induced by heavy grazing in rangelands of SW Spain. Catena 2017, 158, 381–389. [Google Scholar] [CrossRef]
- Yi, J.; Qiu, W.; Hu, W.; Zhang, H.; Liu, M.; Zhang, D.; Wu, T.; Tian, P.; Jiang, Y. Effects of cultivation history in paddy rice on vertical water flows and related soil properties. Soil Tillage Res. 2020, 200, 12. [Google Scholar] [CrossRef]
- Shah, A.N.; Tanveer, M.; Shahzad, B.; Yang, G.; Fahad, S.; Ali, S.; Bukhari, M.A.; Tung, S.A.; Hafeez, A.; Souliyanonh, B. Soil compaction effects on soil health and crop productivity: An overview. Environ. Sci. Pollut. Res. 2017, 24, 10056–10067. [Google Scholar] [CrossRef] [PubMed]
- Kaixi, X.; Beena, A.; Binod, T.; Yanxiang, H. Effect of long duration rainstorm on stability of Red-clay slopes. Geoenviron. Disasters 2016, 3, 12. [Google Scholar] [CrossRef]
- Tu, X.B.; Kwong, A.K.L.; Dai, F.C.; Tham, L.G.; Min, H. Field monitoring of rainfall infiltration in a loess slope and analysis of failure mechanism of rainfall-induced landslides. Eng. Geol. 2009, 105, 134–150. [Google Scholar] [CrossRef]
- Wang, Z.; Li, X.; Shi, H.; Li, W.; Yang, W.; Qin, Y. Estimating the water characteristic curve for soil containing residual plastic film based on an improved pore-size distribution. Geoderma 2020, 370, 114341. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, Y.; Hu, J.; Li, Z. Spatiotemporal variability of soil-water characteristic curve model parameters of Lanzhou collapsible loess. Water Supply 2022, 22, 1770–1780. [Google Scholar] [CrossRef]
- Jiang, H.; Niu, F.; Zhang, H.; Wang, E.; Ma, Q. Study on Soil Water Characteristics of Black Soil, Northeast China. J. Civ. Eng. 2014, 1010–1012, 1153–1157. [Google Scholar] [CrossRef]
- Li, D.; Wang, S.J.; Li, X.; Chen, H.K.; Liang, G.C.; Jiang, W.J. Soil-water characteristic curve of sandy clayey purple soil under different overburden pressures. Chin. J. Geotech. 2021, 43, 1950–1956. [Google Scholar] [CrossRef]
- Zhen, W.J.; Wei, J.; Tang, J.L.; He, Y. Soil-water Characteristics and Shear Strength of Root-soil Composites from Purple-soiled Bunds. J. Soil Water Conserv. 2023, 37, 75–82. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, L.; Gong, W.; Yang, X.; Kang, Y. Evaluation of the water conservation function of different forest types in Northeastern China. Sustainability 2019, 11, 4075. [Google Scholar] [CrossRef]
- Gong, C.; Tan, Q.; Liu, G.; Xu, M. Impacts of mixed forests on controlling soil erosion in China. Catena 2022, 213, 106147. [Google Scholar] [CrossRef]
- Han, Z.S.; Vanapalli, K. Stiffness and shear strength of unsaturated soils in relation to soil-water characteristic curve. Géotechnique 2016, 66, 627–647. [Google Scholar] [CrossRef]
- Bai, Z.; Duan, X.; Ding, J.; Liu, G.; Shi, X.; Feng, D.; Han, X. An Experience estimation model of soil available water-holding capacity: A case study of black soil. Chin. Agric. Sci. Bull. 2015, 31, 153–159. [Google Scholar]
- Kay, B.D. Soil structure and organic carbon: A review. In Soil Processes Carbon Cycle; CRC Press: Boca Raton, FL, USA, 2018; pp. 169–197. [Google Scholar]
- Panchal, P.; Preece, C.; Penuelas, J.; Giri, J. Soil carbon sequestration by root exudates. Trends Plant Sci. 2022, 27, 9. [Google Scholar] [CrossRef]
- Boxell, J.; Drohan, P.J. Surface soil physical and hydrological characteristics in Bromus tectorum L. (cheatgrass) versus Artemisia tridentata Nutt. (big sagebrush) habitat. Geoderma 2009, 149, 305–311. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Nair, P.K.R.; Nair, V.D.; Kumar, B.M.; Showalter, J.M. Carbon sequestration in agroforestry systems. Adv. Agron. 2010, 108, 237–307. [Google Scholar] [CrossRef]
- Jiang, X.J.; Liu, S.; Zhang, H. Effects of different management practices on vertical soil water flow patterns in the Loess Plateau. Soil Tillage Res. 2017, 166, 33–42. [Google Scholar] [CrossRef]
- Noguchi, S.; Nik, A.R.; Sammori, T.; Tani, M.; Tsuboyama, Y. Rainfall characteristics of tropical rain forest and temperate forest: Comparison between Bukit Tarek in Peninsular Malaysia and Hitachi Ohta in Japan. Trop. For. Sci 1996, 9, 206–220. [Google Scholar]
- Cui, Z.; Wu, G.; Huang, Z.; Liu, Y. Fine roots determine soil infiltration potential than soil water content in semi-arid grassland soils. J. Hydrol. 2019, 578, 8. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Tang, Z.; Zhang, Y. Effects of the Root System Architecture of Pinus taeda and Phyllostachys edulis on the Index of Hydrological Connectivity in Subtropical Forest Ecosystems. Forests 2022, 13, 2008. [Google Scholar] [CrossRef]
- Hendrickx, J.M.H.; Flury, M. Uniform and preferential flow mechanisms in the vadose zone. conceptual models of flow and transport in the fractured vadose zone. In Conceptual Models of Flow and Transport in the Fractured Vadose Zone; National Academies Press: Washington, DC, USA, 2001; pp. 149–187. [Google Scholar]
- Allison, G.B.; Hughes, M.W. The use of natural tracers as indicators of soil-water movement in a temperate semi-arid region. J. Hydrol. 1983, 60, 157–173. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Tree Age | Tree Height (m) | Slope (°) | Aspect of Slope | Altitude (m) | Canopy Density (%) | Diameter at Breast Height (cm) |
|---|---|---|---|---|---|---|---|
| CBF | 25 | 13 | 13 | southeast | 220 | 80 | 114 |
| CF | 8 | 16 | 13 | southeast | 140 | 87 | 98 |
| PF | 18 | 12 | 12 | southeast | 120 | 75 | 55 |
| FL | 5 | / | 9 | southeast | 100 | / | / |
| Plot | Soil Horizon/cm | Root Biomass/g | SRL | Root Length Density/m·m−3 | Root Weight Density/kg·m−3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤2 mm | 2~10 mm | >1 mm | SUM | ≤2 mm | 2~10 mm | >10 mm | SUM | ||||
| CBF | 0~20 | 235.31 | 0.09 | 125.74 | 548.18 | 17.75 | 691.67 | 4.62 | 85.52 | 27.52 | 117.66 |
| 20~40 | 53.17 | 0.21 | 125.80 | 128.32 | 6.93 | 261.05 | 3.35 | 17.44 | 5.80 | 26.59 | |
| 40~60 | 32.64 | 0.20 | 101.64 | 95.42 | 0.00 | 197.06 | 2.83 | 13.49 | 0.00 | 16.32 | |
| CF | 0~20 | 99.47 | 0.21 | 369.53 | 303.51 | 1.48 | 674.51 | 12.49 | 36.87 | 0.38 | 49.74 |
| 20~40 | 68.47 | 0.20 | 244.06 | 222.06 | 2.37 | 468.49 | 8.25 | 24.92 | 1.06 | 34.24 | |
| 40~60 | 36.53 | 0.29 | 151.59 | 117.12 | 1.09 | 269.80 | 3.84 | 13.83 | 0.59 | 18.26 | |
| PF | 0~20 | 90.55 | 0.28 | 406.46 | 303.40 | 0.55 | 710.41 | 15.81 | 29.39 | 0.07 | 45.27 |
| 20~40 | 79.20 | 0.16 | 169.40 | 318.89 | 0.00 | 488.29 | 6.68 | 32.92 | 0.00 | 39.60 | |
| 40~60 | 46.12 | 0.20 | 153.15 | 138.06 | 0.78 | 291.99 | 6.71 | 16.41 | 0.06 | 23.06 | |
| FL | 0~20 | 235.88 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 20~40 | 120.34 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 40~60 | 45.03 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Plot | Soil Horizon/cm | Particle Size Distribution/(%) | Bulk Density/(g/cm3) | Organic Matter/(g/kg) | Soil Water Content/(%) | Saturated Water Content/(%) | Maximum Water Retention/(%) | Porosity/(%) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Sand 2 mm~50 μm | Silt 50 μm~2 μm | Clay <2 μm | ||||||||
| CBF | 0~20 | 5.46 | 63.27 | 31.27 | 1.38 ± 0.19 | 23.26 ± 1.90 | 12.34 ± 4.89 | 39.74 ± 6.59 | 31.04 ± 6.78 | 47.86 ± 7.21 |
| 20~40 | 6.70 | 65.39 | 27.91 | 1.49 ± 0.84 | 17.07 ± 3.67 | 10.67 ± 3.18 | 34.63 ± 5.44 | 28.68 ± 4.36 | 43.84 ± 3.16 | |
| 40~60 | 5.83 | 64.04 | 30.13 | 1.66 ± 0.96 | 6.37 ± 2.40 | 9.75 ± 3.87 | 28.43 ± 6.75 | 25.52 ± 6.52 | 37.46 ± 3.63 | |
| CF | 0~20 | 2.69 | 50.17 | 47.14 | 1.27 ± 0.14 | 38.78 ± 3.48 | 20.60 ± 8.62 | 38.45 ± 7.78 | 35.30 ± 7.22 | 52.00 ± 5.17 |
| 20~40 | 4.44 | 63.42 | 32.14 | 1.34 ± 0.12 | 14.89 ± 4.19 | 18.78 ± 5.92 | 35.79 ± 6.48 | 33.30 ± 6.45 | 49.17 ± 4.50 | |
| 40~60 | 5.20 | 71.02 | 23.78 | 1.48 ± 0.08 | 10.59 ± 4.70 | 18.21 ± 7.82 | 32.55 ± 6.31 | 30.66 ± 5.92 | 44.13 ± 2.87 | |
| PF | 0~20 | 5.06 | 52.50 | 42.44 | 1.02 ± 0.13 | 21.98 ± 9.68 | 21.46 ± 5.76 | 55.86 ± 8.82 | 51.24 ± 7.47 | 61.37 ± 4.74 |
| 20~40 | 5.67 | 54.49 | 39.84 | 1.20 ± 0.15 | 13.22 ± 5.59 | 17.89 ± 6.61 | 47.24 ± 6.83 | 45.66 ± 6.41 | 54.76 ± 5.50 | |
| 40~60 | 6.44 | 56.44 | 37.12 | 1.30 ± 0.08 | 7.80 ± 4.47 | 16.05 ± 7.16 | 43.48 ± 5.19 | 42.91 ± 5.41 | 50.82 ± 2.92 | |
| FL | 0~20 | 1.55 | 20.54 | 77.92 | 1.53 ± 0.08 | 6.45 ± 2.04 | 11.75 ± 4.21 | 31.80 ± 6.35 | 24.93 ± 4.39 | 42.10 ± 3.07 |
| 20~40 | 1.23 | 19.97 | 78.80 | 1.58 ± 0.04 | 4.81 ± 1.97 | 9.32 ± 3.92 | 27.83 ± 5.87 | 20.95 ± 6.62 | 40.57 ± 1.35 | |
| 40~60 | 0.95 | 20.17 | 78.89 | 1.61 ± 0.04 | 2.79 ± 1.02 | 6.63 ± 3.75 | 25.34 ± 5.71 | 17.90 ± 7.77 | 39.29 ± 1.62 | |
| Factor | Contribution to Soil Dyeing Area |
|---|---|
| Root length density (D ≤ 2 mm) | 6.89% |
| Root length density (SUM) | 17.49% |
| Total porosity | 17.09% |
| Soil organic carbon | 1.15% |
| Mean weight diameter | 0.48% |
| Soil cohesion | 2.24% |
| Root biomass | 51.48% |
| Specific root length | 3.18% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, B.; Wang, D.; Chen, Y.; Jiang, Y.; Hu, F.; Xu, L.; Zhang, J.; Zhu, J. The Impact of Vegetation Types on Soil Hydrological and Mechanical Properties in the Hilly Regions of Southern China: A Comparative Analysis. Water 2024, 16, 350. https://doi.org/10.3390/w16020350
Zheng B, Wang D, Chen Y, Jiang Y, Hu F, Xu L, Zhang J, Zhu J. The Impact of Vegetation Types on Soil Hydrological and Mechanical Properties in the Hilly Regions of Southern China: A Comparative Analysis. Water. 2024; 16(2):350. https://doi.org/10.3390/w16020350
Chicago/Turabian StyleZheng, Bofu, Dan Wang, Yuxin Chen, Yihui Jiang, Fangqing Hu, Liliang Xu, Jihong Zhang, and Jinqi Zhu. 2024. "The Impact of Vegetation Types on Soil Hydrological and Mechanical Properties in the Hilly Regions of Southern China: A Comparative Analysis" Water 16, no. 2: 350. https://doi.org/10.3390/w16020350
APA StyleZheng, B., Wang, D., Chen, Y., Jiang, Y., Hu, F., Xu, L., Zhang, J., & Zhu, J. (2024). The Impact of Vegetation Types on Soil Hydrological and Mechanical Properties in the Hilly Regions of Southern China: A Comparative Analysis. Water, 16(2), 350. https://doi.org/10.3390/w16020350