Abstract
Harmful algal blooms (HABs) are a primary environmental concern, threatening freshwater ecosystems and public health and causing economic damages in the billions of dollars annually. These blooms, predominantly driven by phytoplankton species like cyanobacteria, thrive in nutrient-rich, warm, and low-wind environments. Because of the adverse impacts of HABs, this review examines various control methods, focusing on biological strategies as sustainable solutions. While effective in disrupting algal populations, traditional chemical and physical interventions carry ecological risks and can be resource-intensive. Biological control methods, including biomanipulation and using algicidal microorganisms such as Streptococcus thermophiles, Myxobacteria, and Lopharia spadicea, emerge as eco-friendly alternatives offering long-term benefits. Additionally, barley and rice straw application has demonstrated efficacy in curbing HAB growth. These biological approaches work by inhibiting algal proliferation, disrupting cellular structures, and fostering algal cell aggregation. Despite their advantages over conventional methods, biological controls face challenges, including intricate ecological interactions. This article delves into the latest biological techniques aimed at eradicating HABs, intending to diminish their frequency and reduce toxin levels in aquatic environments. While most research to date has been confined to laboratory settings, scaling these methods to field applications presents hurdles due to the variability and complexity of natural ecosystems. The review underscores the need for further research and development in this critical area of environmental science.
1. Introduction
Harmful algal blooms (HABs) are an intensive recurring issue in lakes and along coastlines. They harm aquatic life by utilizing dissolved oxygen, lasting from a few days to months. These blooms are dangerous and comprise a lot of biomass and toxins [1]. Ecologists are exploring a variety of methods to solve this problem. The high biomass content of HABs blocks light in the water, preventing submerged plants from developing. When these blooms decompose, they use even more oxygen, which can deteriorate the state of aquatic life [2]. HABs also result in significant economic distress, leading to multimillion-dollar losses on a global scale [3]. They release toxins that are capable of poisoning about 2000 people every year [4]. There are several methods for controlling HABs; however, they can be costly, pollute the environment, or be challenging to utilize in fields [5]. Only a handful of these strategies are effective on a large scale [6]. The toxins from HABs are dangerous as they can enter into the brain and skin. These HABs harm aquatic organisms and pollute the air when they release toxins [7]. Cyanobacteria, often found in these waters, grow well using nitrogen and phosphorus [8]. Fish and bacteria feed on these toxins, often leading to massive death due to insufficient oxygen and too many toxins [9].
Developing new strategies to counteract the detrimental effects of HABs is essential. Tackling the challenge of HABs mitigation necessitates the adoption of a variety of regulatory approaches [10]. Physical interventions encompass techniques such as nutrient load reduction, sonication, UV-C exposure, soil adjustments, hydrodynamic methods, and diverse filtration processes to mitigate HABs [11]. While most of these are environmentally friendly, they are energy-intensive, potentially rendering them economically impractical for large-scale applications [12]. Chemical treatments such as Triosyn, hydrogen peroxide, and copper sulfate have shown efficacy in short-term use. However, it is vital to acknowledge their adverse effects on aquatic ecosystems [13]. Conversely, biotic factors have been shown to positively influence the removal of HABs, playing a pivotal role in their prevention, cessation, and control. These microbial populations, called microbial herbicides, are instrumental in combating HABs [14].
Biological control agents like viruses, protozoa, and bacteria are utilized for targeted applications. It is now widely acknowledged that bacteria are vital in regulating phytoplankton biomass in freshwater environments [15]. Many bacteria associated with these environments possess algicidal properties, playing a crucial role in the decomposition and decay of algal blooms. In contrast, others contribute directly to bloom formation [16]. The impacts of blooms are evident in various forms, notably within marine ecosystems where marine life is at risk due to toxin exposure through ingestion. This highlights the necessity for effective methods to predict and mitigate the impacts of HABs [17]. The biological control of HABs is a standout solution for its economic and environmental benefits, and also avoids secondary pollution [18].
Bacteria, in particular, have demonstrated various strategies for combating HABs. These include the secretion of cyanobacterial substances, engaging in cell-to-cell contact mechanisms, producing acyl-homoserine lactone signals, generating antagonistic volatiles, inhibiting photosynthetic electron transport reactions and the activities of glycolate dehydrogenase and nitrogenase [19]. They also have secondary metabolites, release mucous-like secretions for self-defense, and use entrapment techniques leading to the lysis of cyanobacteria [19].
Viral degradation employs species-specific interactions, cell bursting, and the virus lytic cycle, offering the benefit of targeting specific species. Fungi have been observed to attack HABs directly [20]. Zooplankton contribute through grazing, a process that aids in removing invasive species and benefits both fish and zooplankton. Fish use an ingestion and digestion mechanism alongside grazing, which is advantageous for toxin elimination as they can digest the toxins. Algae, mainly through flocculation, have shown potential for bloom control [21]. Golden algae have been identified as mitigators of Microcystis cells and as toxin degraders. Bioflocculation is notable for its minimal harm to other organisms, making it an ideal approach for removing HABs [22].
A primary concern in this context is the potential for harm that extends beyond the intended target organism, impacting the survival of predatory organisms and the presence of other predators. Significant challenges arise in the large-scale production of microbial agents, compounded by their storage and application issues. This is particularly evident when introducing a single microbial agent into field settings. A principal obstacle in biotic mitigation lies in translating laboratory-proven methods into practical environmental applications [23]. This involves integrating various biological factors with a distinct life cycle into an ecological setting, posing considerable challenges.
This article will explore the ongoing problem of HABs, investigating their adverse effects on aquatic ecosystems and the challenges they present. While various strategies have been proposed to mitigate HABs, our primary focus will be physical, chemical, and, especially, recent advancements in biological methods. These innovative approaches aim to reduce the occurrence of HABs and effectively lower toxin levels in aquatic environments. We will explore the potential of microbial agents, viruses, fungi, zooplankton, fish, and algae in combating HABs, highlighting their ecological significance and potential benefits.
2. Materials and Methods
This review includes data from experimental research completed over the last 50 years, from 1973 to 2023. We conducted a thorough search of the literature through Scopus, including terms such as “biological control”, “algal bloom control”, “HAB mitigation”, and “natural algae predators”. Figure 1 illustrates the prevalent terms found in the studies. It is apparent from the figure that the term “algal bloom” was the most frequently utilized in the articles, appearing over 140 times. Following closely was the term “algae”, mentioned approximately 120 times, and “eutrophication”, cited around 115 times. The frequency gradually decreases for other terms, with words like “temperature”, “physiology”, and “humans” being among the least common, each occurring about 15 times, maintaining a similar pattern of presentation.

Figure 1.
Word map showing the most common keywords.
Remarkably, a significant portion of the studies predominantly concentrated on measures related to biological management and the impact of toxic algal blooms. Figure 2 illustrates the distribution of publications across various journals, with the size of each bar indicating the number of articles published by the respective journals. Notably, journals such as “Phycologia”, “Water Research”, “Water Science and Technology”, “Chemosphere”, “Hydrobiologia”, “Science of the Total Environment”, “Journal of Eukaryotic Microbiology,” and “Marine Pollution Bulletin” contributed significantly to the overall publications. Notably, the journal “Environmental Science and Technology” boasts the highest Impact Factor (IF) among the journals listed, scoring 11.4. Following closely is “Science of the Total Environment” with an (IF) of 9.8, and lastly, “Environmental Pollution” holds an (IF) of 8.9. In general, the Impact Factors (IF) across the listed journals ranged from 3.1 to 11.4. These rankings are based on data from the year 2023.
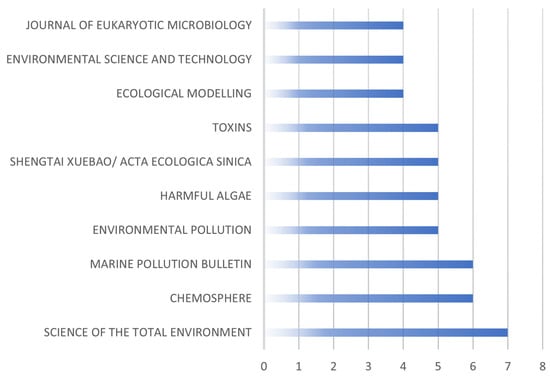
Figure 2.
Journals publishing information on biological control methods of HABs.
The intricate network analysis map for publishing countries in this bibliometric analysis is depicted in Figure 3. Each box in the figure represents the number of publications, while the arcs indicate the total link strength to other publishing countries. A substantial number of publications in this study originated from countries such as the United States, China, and Japan. China emerged as the leading publishing country with 75 publications, followed by the United States with 46, and a total of 19 each for Australia and South Korea. Regarding total link strength and illustrating collaborative efforts between countries, the United States exhibited the highest number of links, followed by China, with Germany ranking third in this aspect.
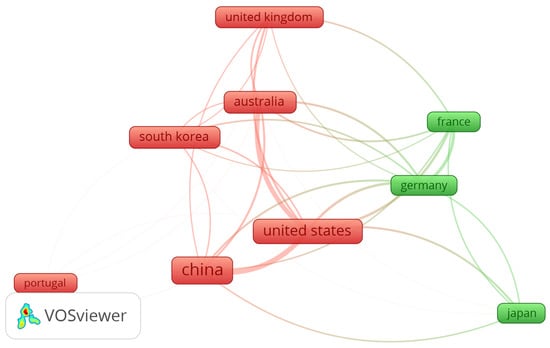
Figure 3.
Map of countries with the most significant publications on the topic.
The gathered data underwent careful classification based on the specific biological strategies employed to manage HABs. Among the prominent methods, using bacteria, zooplankton, and other microorganisms to impede algal growth stands out. Toxins generated by toxic algae species, for example, can be broken down by particular bacterial strains. The primary questions driving this meta-analysis are: Which biological control strategies most effectively prevent HABs? Are there any factors in operation that increase or decrease the effectiveness of the biological controls besides environmental parameters such as pH, light intensity, and temperature? Here, our analysis primarily focused on the evaluation of biological control approaches.
3. Causes of Algal Blooms
Phytoplankton depend on light, carbon dioxide, and nutrients to generate biomass. These elements form the basis of food chains by supplying energy and resources. The growth of phytoplankton depends on the proper mix of light and nutrients. This balance in water ecosystems is vital for life support [24]. Also, blooms occur when predators fail to control rising phytoplankton numbers [25]. This can be attributed to a combination of factors that lead to their spread and specific reasons for the proliferation of different algae species [26]. Chakraborty et al. [27] have highlighted common factors like deteriorating water quality, rising eutrophication, intense aquaculture activities, transport of harmful species through ballast water, and climate change. Algal blooms proliferate under certain conditions. Figure 4 shows the difference between heavy and light bloom conditions. Heavy blooms need many nutrients, warm and calm water, little wind, bright light, and not much salt, along with a higher PH and lower dissolved oxygen. Light blooms happen with less nutrients, cooler water, turbulent water, wind, and salt, followed by a lower PH and a higher dissolved oxygen content [28]. Blooms comprise different tiny water plants like diatoms, dinoflagellates, and cyanobacteria, which can grow in the ocean and freshwater. These tiny plants float to the top of the water and gather together, changing the watercolor from green to red, depending on the type of plant [29].
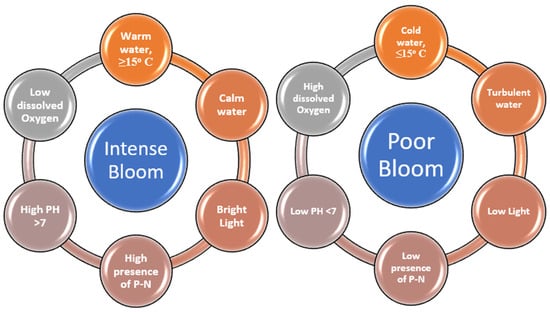
Figure 4.
Difference in parameters between intense and poor blooms.
Typically, factors promoting HABs include the rate of introducing species to new areas and local environmental shifts. Such conditions boost the growth of certain species, resulting in HABs. Environmental changes might be minor, and not all factors might shift at once, causing situations where one aspect seems beneficial [30]. Adding a new species to water bodies is not always feasible, as it is contingent on environmental factors and the adaptability of the new species [31]. Human actions are the primary drivers of environmental changes, leading to a rising global trend of HABs. These changes predominantly involve nutrient variations, growth in aquaculture, and major waterway projects [32,33]. Finally, extended stratification periods, rising temperatures, and heightened nutrient input also contribute to a growing prevalence of HABs.
The endeavor to control and handle HABs has been rigorously examined due to their harmful consequences on marine ecosystems. Multiple approaches have been assessed, including chemical, physical, and biological techniques. Figure 5 illustrates a comparative flowchart of these techniques. Each possesses its unique benefits and challenges. This portion elaborates on these methods, highlighting their effectiveness, ecological implications, and practicality in managing HABs.
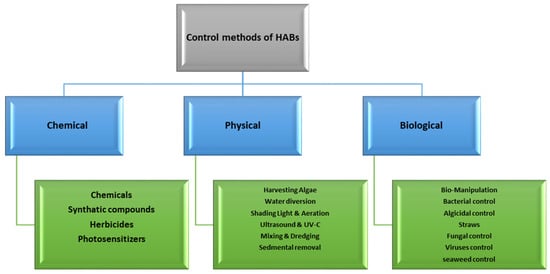
Figure 5.
Flowchart of different control methods of HABs.
4. Methods to Control HABs
4.1. Chemical Methods
Chemical methods are a pivotal approach in the battle against HABs, garnering considerable attention for their effectiveness. These strategies primarily involve the application of metals such as copper, iron, and aluminum, recognized for their ability to control algal populations [11]. The core of their efficacy lies in disrupting algal cells, targeting cell membranes, and interfering with essential cellular functions. In addition to these metals, herbicides like atrazine and simazine have been rigorously explored for their capacity to curb algal growth and spread [34]. Photosensitizers, particularly hydrogen peroxide, have also been investigated for their potential to suppress algal proliferation [35].
However, it is crucial to acknowledge and confront the inherent limitations of chemical methods. While offering practical and often cost-effective means to manage HABs, these interventions demand meticulous oversight to mitigate ecological disturbances and related risks [11]. Striking an optimal balance between the advantages of diminishing algal populations and the potential hazards of toxin release is vital, highlighting the necessity for continuous research. Such ongoing investigations are crucial to enhancing our understanding and refining our approaches to addressing the complex challenges posed by HABs [36].
4.2. Physical Methods
Various strategies are classified as physical methods in addressing HABs. These cover techniques include limiting nutrient influx, UV-C exposure, soil condition modifications, hydrodynamic actions, and multiple filtration processes [37]. A practical step in reducing nutrient inputs involves curbing nitrogen and phosphorus levels, as limiting these essential nutrients can hinder the growth of HABs. UV-C exposure offers a method where ultraviolet light disrupts algal cells, rendering them less potent [38,39]. While ultrasonication’s large-scale algal control effectiveness remains debated, ultrasound emerges as a promising tool. It is compact, straightforward, and deployable in the field, capable of addressing algae over a considerable area, and its energy demands are moderate. Notably, ultrasonography, unlike chemical methods, is residue-free. Nevertheless, even with supportive evidence for ultrasound’s effectiveness in algal control, comprehensive full-scale field tests are scarce, and there are limited data on its commercialization [11]. Soil alteration techniques aim to change the aquatic ecosystem, thus inhibiting algal proliferation. Hydrodynamic processes, in contrast, aim to disperse algal clusters, breaking their unity. Filtration methods work by mechanically extracting algal cells from water. These varied physical tactics provide an environmentally friendly solution with a minimized contamination risk [40]. Dredging proves advantageous in eliminating polluted sediment, yet its expense renders it an impractical approach for managing blooms in most systems [41]. A research review by Faith A. Kibuye et al. summarized that aeration boosted phytoplankton richness, as seen by a more uniform distribution of accessible species, according to long-term monitoring. In addition, there was a shift from cyanophyte dominance to a greater prevalence of cryptophytes [41]. For numerous years, artificial mixing has been employed as a strategy to curb the proliferation of cyanobacteria in eutrophic lakes and reservoirs. This method generally leads to a rise in the oxygen levels within the water [42]. The disruption of the water column and sediment caused by mixing can result in the resuspension of nutrients, leading nutrient-rich pore-water to spread throughout all depths. This nutrient dispersion throughout the water column may enhance algal development, impeding efficient cyanobacterial management [43]. Nonetheless, the limitations of these methods must be acknowledged. Many require significant energy, potentially reducing efficiency and cost-effectiveness, especially in large-scale algal bloom scenarios. The financial implications of their deployment also warrant consideration [43,44,45].
4.3. Challenges, Monitoring, and Considerations
Though the suggested strategies offer potential remedies for curbing HABs, understanding that each method has intrinsic challenges and constraints is crucial. Chemical techniques, while cost-effective and robust, pose environmental hazards. While eco-friendly, physical processes might demand significant energy and monetary investments. Biological processes, with their ecological advantages, could serve as viable alternatives to these conventional approaches. A detailed juxtaposition of physical, chemical, and natural methods, along with their potential impacts, is provided in Table 1. Moreover, the new analysis focuses on the various tactics nations and commercial companies use to monitor and regulate HABs in coastal waterways [46], which in turn are complementary to appropriate ways to meet the goals of the control methods, as monitoring HABs will allow for the responsible entities to act early and manage cases before they spread widely. Remote biosensors [36], atomic force microscopy [47], matrix-assisted laser desorption/ionization-time of flight (MALDI-TOF) mass spectrometry (MS), the enzyme-linked immunosorbent test (ELISA) [48], and satellite remote sensing [49] are some examples of these approaches. These techniques have been demonstrated to be helpful in various circumstances.

Table 1.
Comparative analysis of methods for combating HABs.
4.4. Biological Management of HABs
Biological techniques are drawing interest due to their environmentally friendly and cost-efficient nature compared to chemical and physical methods. These approaches harness the inherent abilities of diverse microorganisms, such as macrophytes, microalgae, macroalgae, bacteria, viruses, actinomycetes, and pathogens, to regulate HABs. For example, biomanipulation introduces specific biological entities that can outcompete or feed on detrimental algal types [45]. Macrophytes and macroalgae can take up surplus nutrients, thereby curbing algal proliferation. Additionally, several strategies employ bacteria and virus species as tools to destabilize algal communities. Notably, Streptomyces neyagawaensis emits particular allelopathic compounds that can stifle the expansion of existing algal cells [59]. Beyond these, recent research has tested crop straws like rice and barley for HABs control [57]. The appeal of biological techniques stems from their eco-compatibility and potential for sustained efficacy. Nevertheless, their use might be constrained by challenges such as grasping intricate ecological dynamics and the possibility of unforeseen repercussions [58]. The present review focuses on the different biotic organisms isolated and used to remove HABs and their mode of action.
4.4.1. Biomanipulation
Biomanipulation is a method of bioengineering aimed at mitigating algal overgrowth by modifying the dynamics within an ecosystem. This technique strategically imposes ecological stress on phytoplankton populations to curb the proliferation of unwanted algal forms [60]. Additionally, it adjusts the predatory impact of fish on zooplankton populations, promoting the prevalence of larger zooplankton species within the ecological hierarchy. Two primary methods are employed to regulate fish populations conducive to biomanipulation: eradicating zooplanktivorous fish species and introducing Piscivorous species, such as silver carp, into the aquatic environment [61].
Recent advancements in the field have highlighted the potential of algal phytoremediation systems, such as algal turf scrubbers, to effectively sequester nutrients from eutrophic surface waters. These systems help manage nutrient levels and carbon sequestration, benefiting ecosystem management efforts [62]. Furthermore, studies have pointed out the imbalance in global nutrient cycles, with more excellent phosphorus retention over nitrogen in lakes, which could exacerbate the challenge of controlling algal blooms [63].
The baseline effort for effective fish capture is categorized by a substantial reduction of 75–80% over several years or a consistent removal rate of 200 kg per hectare over three years. A notable study has introduced a mathematical model specifically for temperate, shallow lakes to estimate the necessary annual fish capture using the formula:
Required annual catch (kg per hectare) = 6.9 × TP0.52, where TP denotes the total phosphorus concentration in milligrams per liter [64]. The study further suggests that the annual catch requirements for deeper lakes may be less stringent than those for their shallower counterparts. While biomanipulation is considered a viable alternative to nutrient restriction methods, its effectiveness is contingent upon various factors, including the dynamics of fish populations, the resilience of the ecosystem, and effective nutrient management practices. Ensuring the enduring success of biomanipulation demands a holistic approach to ecosystem management that extends beyond mere trophic-level adjustments [61].
Furthermore, research by Gallardo-Rodríguez et al. [40] and colleagues has indicated that the efficacy of biomanipulation is notably enhanced when it is part of a broader strategy that includes comprehensive nutrient reduction. An illustrative case is Lake Vesijärvi in Finland, where implementing nutrient management, coupled with the targeted fishing of Planktivorous coregonid fish, led to a marked decrease in phytoplankton biomass, thereby serving the public interest [65]. Acknowledging that biomanipulation is often advocated as a cost-efficient substitute for conventional eutrophication management practices is critical [66]. Nonetheless, various studies have highlighted this approach’s intricate and sometimes unpredictable outcomes, with large grazers sometimes only offering a temporary decrease in algal biomass. Recent research underscores the importance of a season-based pollutant management strategy, which is cost-effective in controlling algal blooms in response to future climate changes [67].
Longevity of Biomanipulation
The influence of the persistence of biomanipulation on aquatic ecosystems, particularly its role in mitigating HABs and bolstering water quality, is subject to a complex interplay of factors. The resilience of the ecosystem, the influx of nutrients, the dynamics between predators and prey, and climatic conditions all contribute to the enduring and immediate outcomes of biomanipulation efforts [68]. For the longevity of these effects, proactive measures such as continuous monitoring and the capacity to adapt to changing conditions are critical [43]. Furthermore, adopting sustainability initiatives, like enacting policies to curtail nutrient pollution, is vital for securing the long-term advantages of biomanipulation. In essence, the sustained success of biomanipulation depends on ecological, environmental, and managerial considerations, necessitating a comprehensive approach to ecosystem stewardship [68].
Biomanipulation is predominantly applied in Europe’s small and shallow lakes, where it is advised to significantly curtail the population of certain fish species within one to two years to affirm its lasting effect. It promotes the development of juvenile fish capable of evolving into piscivores upon reaching maturity [69]. A study conducted on a 14.8 square kilometer section of a Swedish lake serves as a testament to the impressive results achievable through biomanipulation [67]. Here, Ekvall et al. observed that the extraction of cyprinid fish led to reduced predation on zooplankton communities, which sparked a 55% surge in the population of Daphnia sp. [70]. This increase in Daphnia numbers subsequently drove a considerable decline in the biomass of toxin-producing cyanobacteria. This marked improvement culminated in repeated treatments initiated a decade following the first biomanipulation effort. Despite this, the impact of the initial biomanipulation began to diminish over five years [71].
Moreover, the long-term impact of biomanipulation has been linked with external phosphorus load. Internal nutrient management has also been shown to sustain the effects of biomanipulation for up to seven years, as delineated by Jůza et al. [72].
Determinants of Biomanipulation Efficacy
The success of biomanipulation as a treatment strategy is governed by an intricate array of factors. Achieving an optimal equilibrium between herbivorous species’ feeding rates and cyanobacteria’s reproductive rates is essential for effectively mitigating HABs [68]. The objective is for biomanipulation to exert a uniform effect on the diverse characteristics of cyanobacteria [40]. However, should cyanobacteria’s reproductive rate surpass herbivores’ grazing capabilities, the system may need to be revised. Studies have demonstrated that strategically removing non-predatory fish and introducing Piscivorous species can significantly amplify the grazing pressure exerted by zooplankton [73]. This intensification in grazing curtails cyanobacterial proliferation and coincides with a decrease in nutrient recycling, thereby exerting a dual effect on controlling cyanobacterial growth in shallow lakes. De Backer et al. [74] have further corroborated the pivotal role of nutrient management in suppressing cyanobacterial blooms.
Environmental Conditions and Weather
The interplay of climate and weather patterns significantly shapes the efficacy and longevity of biomanipulation strategies. Variations in climate temperatures can profoundly influence the layering of water bodies, which, in turn, dictates the spatial arrangement of plankton and fish populations [75]. In scenarios where warmer temperatures induce extended periods of water stratification, conditions may become more conducive to the proliferation of HABs [76]. Moreover, the patterns of precipitation, ranging from intense rainfall to prolonged droughts, play a critical role in modulating the influx of nutrients into aquatic systems, which can either bolster or undermine biomanipulation efforts [77]. The occurrence of storm events is another vital factor, with the potential to significantly alter water quality and the dynamics of nutrient distribution, thereby affecting the success of biomanipulation interventions.
Additionally, wind patterns are instrumental in mixing aquatic environments, which can lead to shifts in species distribution [78]. The overarching challenge of climate change introduces additional layers of complexity, underscoring the need for biomanipulation strategies that are robust and adaptable. Consequently, integrating climate and weather variables is indispensable for a comprehensive assessment of the enduring impact of biomanipulation techniques [79].
4.4.2. The Role of Submerged Macrophytes in Eutrophication Control
Submerged macrophytes, the foundational autotrophs of lacustrine systems, play a pivotal role in mitigating eutrophication through their interactions with living and non-living elements of shallow aquatic environments [80]. These plants are formidable competitors for nutrients, often outcompeting phytoplankton for these vital resources. Macrophytes are especially crucial in ecosystems burdened with high nutrient loads, as they can effectively regulate nutrient availability [81]. The functions of submerged macrophytes are multifaceted. Primarily, they sequester nutrients from the sediment and the overlying water, thereby imposing a state of nutrient scarcity that can suppress cyanobacterial growth. In addition, macrophytes provide refuge for zooplankton populations, which serve as herbivores that graze on phytoplankton, thereby indirectly curtailing the expansion of cyanobacteria and promoting a zooplankton-dominated system [82]. Lastly, macrophytes synthesize and release allelochemicals—distinctive secondary metabolites with the capacity to inhibit phytoplankton proliferation—thus further restraining the growth of cyanobacteria [82].
The strategic deployment of macrophytes in the management of cyanobacterial populations demands a comprehensive understanding of several factors: the necessity for sufficient coverage of macrophytes, the persistence and efficacy of allelochemicals within the aquatic milieu, and the susceptibility of specific cyanobacterial species to these biochemical inhibitors [83].
Utilization of Natural and Engineered Macrophytes
Aquatic plants that thrive beneath the water’s surface, featuring fully submerged foliage and stems, serve as excellent natural macrophytes for extracting excess nutrients from nutrient-rich lakes. Beds of eelgrass, for instance, are not only highly productive but also support a wide array of marine life. They are instrumental in absorbing nutrients from the water, thereby preventing the overgrowth of phytoplankton in eutrophic coastal zones by reducing nutrient availability [84]. Additionally, these plants offer a habitat for a variety of aquatic organisms. Similarly, with its elongated, tape-like leaves, Vallisneria excels in nutrient absorption, particularly within freshwater systems [84]. Conversely, Elodea species are distinguished by their rapid growth and substantial nutrient uptake capabilities, which benefit nutrient removal [79].
Beyond natural plant solutions, engineered macrophytes also play a role in eutrophication management. Floating treatment wetlands (FTWs) are deployed effectively in highly and moderately nutrient-enriched waters to curb the progression of algal blooms and the associated shifts in water chemistry, such as significant pH level increases [85].
The Impact of Macrophyte Density on Algal Suppression
The presence of macrophyte canopies plays a pivotal role in mitigating eutrophication. Notable declines in the levels of nutrients, water cloudiness, and chlorophyll-a have been documented in areas where Ceratophyllum demersum L. establishes a 20–50% coverage [86]. Additional studies have corroborated the beneficial link between the abundance of macrophytes and the clarity of the water body. For example, a macrophyte cover exceeding 25% of a lake’s surface is associated with reduced algal biomass [87]. Furthermore, increased macrophyte coverage within lakes correlates with improved conditions, such as greater water transparency [88].
Allelopathic Dynamics in Aquatic Ecosystems
Macrophytes contribute to aquatic ecosystems through allelopathic interactions by sustainably releasing bioactive compounds that suppress phytoplankton biomass. Research by Declerck et al. [80] highlighted that Elodea nuttallii exerted a significant allelopathic influence on microalgal populations, with the suppression enduring beyond 50 days. Similarly, Švanys et al. [81] observed that in the Curonian Lagoon Myriophyllum spicatum’s presence negatively impacted microalgal growth under nutrient-rich conditions.
Further studies have demonstrated that Stratiotes aloides can markedly reduce microalgal proliferation, with in situ experiments showing a 60% decrease in cyanophyte presence and a 73% reduction in overall phytoplankton [82]. A field study in Lake Krumme Laake, Berlin, attributed a decline in microalgal biomass to the allelopathic influence of Myriophyllum verticillatum, reinforcing macrophytes’ significant role in regulating algal populations.
Zhu et al. [82] conducted an analysis using high-performance liquid chromatography (HPLC) to pinpoint phenolic compounds in macrophytes that were effective against Chlorella sp. and Anabaena sp., both of which are known contributors to harmful algal blooms.
Notably, many field studies focusing on these allelopathic effects are situated in shaded and still water environments, which are more prone to harmful algal bloom occurrences [88].
4.4.3. Utilizing Straw to Combat Harmful Algal Blooms
The emergence of cyanobacterial blooms poses significant ecological and public health challenges by releasing toxins detrimental to marine life and humans and compromising water quality [70]. The strategic application of organic matter, such as barley and rice straw, has been recognized as an innovative method to manage these cyanobacterial outbreaks. Notably, barley straw has been singled out for its effectiveness in mitigating the growth of cyanobacteria [40].
Mechanism of Action
The proposed mechanism behind barley straw’s success in curbing cyanobacterial growth involves the release of allelopathic substances during its aerobic breakdown [84]. Allelopathy describes the biochemical interactions between plant species, where the compounds produced by one can affect the growth of another. In the presence of oxygen, barley straw decomposes to produce chemicals believed to suppress cyanobacteria’s expansion [83].
The disintegration of straw in water bodies can increase the oxygen demand. Maintaining oxygen-rich conditions is crucial to ensure the method’s efficacy. In scenarios where dissolved oxygen (DO) levels are low, the anti-algal properties of the straw may be compromised. However, ponds with aeration systems have shown more promising results [83]. These aeration systems are vital in preserving oxygen levels to continue the algae-inhibitory effects during straw decomposition [40].
For example, studies targeting the control of Anabaena flosaquae have demonstrated that barley straw can significantly curtail algal proliferation. Gallardo-Rodríguez et al. [40] observed a notable reduction in DO levels when straw decayed without sufficient aeration, underscoring the importance of proper oxygenation to balance effective algae management and acceptable oxygen concentrations in water bodies.
Over the last ten years, research by Gallardo-Rodríguez et al. [40] has suggested that compounds such as hydrogen peroxide (H2O2) may be produced during the photooxidation of substances like lignin and quinone, which are byproducts of barley straw decomposition, as depicted in Figure 6. The exact processes remain elusive, necessitating further investigation to elucidate the underlying mechanisms fully.

Figure 6.
Photooxidation of lignin and inhibition of microalgae.
Barley Straw
In an empirical investigation within the UK, researchers dispersed 50 g per cubic meter of barley straws into a water reservoir to test the straw mechanism theory. The outcome was profound, with a 90% decline in algal presence noted, a stark contrast to prior years without the addition of barley straws [89]. This marked reduction in algal biomass underscores the efficacy of barley straws in curbing cyanobacteria overgrowth.
After the barley straw deployment, water samples were analyzed chemically, revealing a spectrum of organic compounds downstream. Notably, several of these compounds are recognized for their lethal effects on algae [90]. The detection of these substances post-straw decomposition offers a plausible rationale for the reduction in algal numbers. Further research indicated that an extract of barley could thwart the growth of Microcystis aeruginosa, evidenced by the chlorophyll-a concentration decreasing by a factor of ten relative to the control, hinting at the decomposed barley straw extract’s capacity to restrain algal proliferation [91].
An additional study lent credence to these observations. Here, 25 g per cubic meter of barley straws were introduced to a reservoir. Within a mere 12 days, algal cell counts plummeted by 90% [91]. The study also documented the swift emergence of phenolic compounds, notorious for their algicidal properties. This emergence of bioactive compounds adds weight to the theory that allelopathic interactions are instrumental in modulating cyanobacterial communities [92].
The exploration of chopped wheat straw’s effect on algal bloom dynamics within aquatic ecosystems was the focus of research by Mariraj Mohan [93]. In a departure from anticipated outcomes, introducing chopped wheat straw did not suppress algal blooms. On the contrary, its presence was associated with an uptick in HABs proliferation. This was quantified by measuring chlorophyll-a levels, a reliable measure of algal biomass. Chlorophyll levels were recorded at 400 micrograms per liter in environments devoid of wheat straw. However, these levels surged to 520 mg/L following the introduction of wheat straw [93].
Additionally, a separate investigation reported that straw bales placed in a loch had no observable effect on algal concentrations throughout the study. Despite initial assumptions that straw bales might help curb algal blooms, the evidence suggested that their deployment did not lead to a marked decrease in algal presence [94].
Rice Straw and Ragi Straw
Mariraj Mohan’s research has shed light on the suppressive effects of rice and ragi straw on algal proliferation in water bodies [88]. These straws, through the collective action of their inherent compounds, have shown a notable capacity to impede algae growth. In particular, rice straw, when decomposed at a 2 g/L concentration, was markedly effective, reducing chlorophyll levels in treatment tanks to about 29.32 mg/m³ over ten weeks—a stark contrast to the 46.73 mg/m³ observed in control tanks. This finding suggests that the breakdown of these straws releases substances that prevent algae from spreading rather than removing existing colonies [93].
Further exploration into the effects of rice straw extracts on aquatic microorganisms revealed exciting dynamics. Over a two-week study, Chlorella sp. and Anabaena sp., harvested from the Abbassa fishpond, responded differently to the straw extracts. The extracts significantly curtailed Anabaena’s growth, with the most profound inhibition—over 95%—occurring at extract concentrations of 1, 5, and 10 mg/L. Intriguingly, these same extracts seemed to promote the growth of Chlorella, highlighting a selective inhibitory effect [95]. A detailed description of straw utilization is provided in Table 2.
Table 2.
Straw utilization against HABs.
4.4.4. Bacterial Use for Controlling HABs
Bacteria exhibit an array of dynamic defenses against HABs. They initiate their countermeasures by swiftly diminishing algal cell populations through the intriguing process of alga-bacterium bioflocculation. This natural phenomenon involves the aggregation of algal cells, precipitating a marked decrease in their numbers [96]. Additionally, these microorganisms serve as natural overseers, adeptly curbing the rampant spread of deleterious algae. By deploying algicidal bacteria, they leverage a lysis mechanism to dismantle their algal counterparts, methodically mitigating potential harm. In a final display of their ecological prowess, bacteria form synergistic clusters known as microbial aggregates, culminating in developing periphytons and biofilms [96]. This collective strategy underscores their collaborative strength and critical role in preserving the delicate balance of aquatic habitats and protecting them from the destabilizing effects of HABs [97].
Bioflocculation
Bacterial bioflocculation is a pivotal method for eradicating algal blooms, relying on the collective settling of algal cells. This process is orchestrated by bacteria, which compel algal cells to emit specific extracellular polymeric substances (EPS). These substances significantly boost the clumping of algal cells, causing them to settle, a process illustrated in Figure 7.
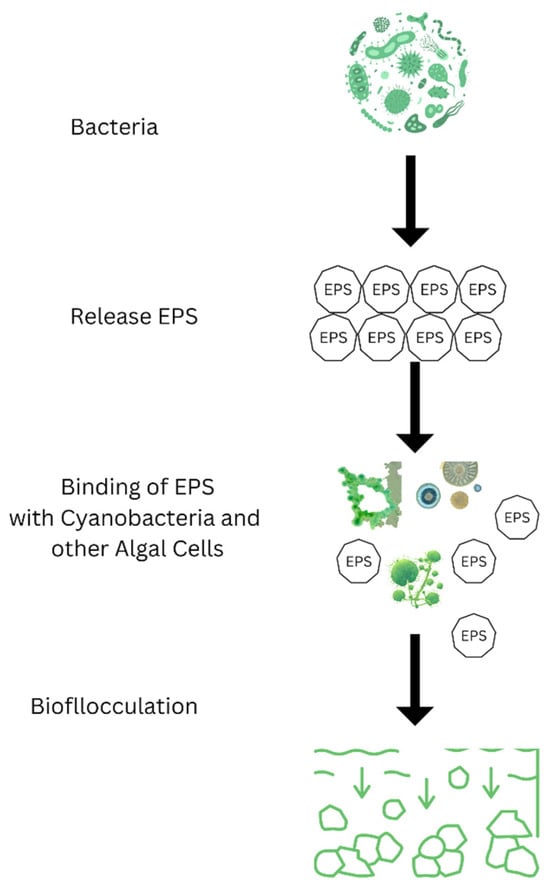
Figure 7.
Schematic diagram of bio-flocculation of algae by bacterial EPS release.
This natural occurrence of bacterial-induced algal clumping is prevalent in aquatic ecosystems, with bacterial species like Porphyrobacter playing a critical role in this aggregation [37]. Consequently, using bacterial bioflocculants has become a focal point in managing HABs.
Bioflocculants, instrumental in HABs mitigation, comprise bacterial secretions such as polysaccharides, proteins, and lipids [98]. Nevertheless, deploying these bioflocculants from pure bacterial cultures is primarily experimental and restricted to laboratories, pilot plants, or localized in situ applications [83].
Inhibition of Harmful Algal Growth
In aquatic ecosystems, the interplay between bacteria and algae is a delicate balance of survival, often hinging on the suppression of HABs [96]. In their quest for dominance, Algicidal bacteria secrete various allelochemicals that give them an edge over algal species. In nutrient-rich, eutrophic lakes, specific bacterial strains stand out for their ability to curb the proliferation of cyanobacteria. Notably, the Bacillus fluxes strain SSZ01 has been identified for its secretion of harmine and norharmane, compounds known for their potent effects against cyanobacteria [97]. Furthermore, a raw bacterial extract containing β-carbolines has been shown to antagonize cyanobacteria by interfering with their cell division [99]. These insights underscore the significant potential of bacteria as a strategic tool for the targeted inhibition and management of HABs.
Algicidal bacteria utilize direct and indirect tactics to eradicate HABs, showcasing their capacity to control these detrimental occurrences [100]. These microorganisms directly engage in physical interactions with algae, leading to their destruction. A notable instance is the Myxobacteria, the first identified algicidal bacteria, which can decisively eliminate unicellular Cladophora through direct contact [101]. Similarly, the Streptococcus thermophilus strain LY03 employs chemotaxis to move toward algal cells and then secretes chitinase enzymes that break down the algal cell walls, causing the cells to lyse and die [102].
Indirectly, bacteria secrete various algicidal substances ranging from peptides, proteins, alkaloids, amino acids, antibiotics, pigments, and fatty acids [98]—Bacillus sp. SY-1, for example, produces Bacillamide, a polypeptide that specifically targets the algal species Cochlodinium polykrikoides [103]. Other noteworthy compounds include phenazine from Pseudomonas aeruginosa strain O-2-2, deinoxanthin from Deinococcus xianganeasis Y35, prodigiosin from Hahella KA22, and palmitoleic acid from Vibrio sp. BS02, all of which have been shown to inhibit various algae, highlighting their potential in HABs management [104]. Moreover, Bacillus bacteria emit Mycosubtilins that disrupt the cytoplasmic membrane of algae, increasing ion permeability and causing the rupture of dinoflagellates [105].
Field studies, however, indicate that bacteria’s algicidal effects can vary significantly from those observed in controlled laboratory experiments. A 2013 study pointed out bacterial cell concentrations ranging from 3.7 × 103 to 1.3 × 106 per ml are necessary to achieve algicidal activity in natural settings [105]. Additionally, the increased presence of these bacteria during the declining phases of algal blooms suggests their role in the natural attenuation of HABs [106,107,108].
Actinomycetes for Inhibiting HABs
The actinomycete Streptomyces neyagawaensis has emerged as a promising agent in the fight against algal blooms. This bacterium is adept at synthesizing allelopathic substances with algicidal effects capable of curbing the expansion of various algae, notably those detrimental to aquatic ecosystems [108].
Intrigued by its potential, researchers have embarked on investigative studies to gauge the effects of S. neyagawaensis on predominant algal species in the Paltang, Juam, Daechung Reservoirs, and the Naktong River [109]. The outcomes of these studies were quite revealing, showing that S. neyagawaensis selectively targets specific algal species within a genus, effectively inhibiting the growth of A. flosaquae and A. cylindrical while leaving A. macrospora and A. affinis unaffected. Additionally, the bacterium’s impact differed even among strains of the same species, highlighting its selective and nuanced approach to algal control [98]. The utilization of bacteria against HABs is summarized in Table 3.
Table 3.
Bacteria utilization against HABs.
4.4.5. Fungi for Algal Control
Algicidal fungi are pivotal in managing algal populations within aquatic ecosystems, thanks to their ability to suppress algal growth. The primary mode of their algicidal action involves the release of bioactive substances that either impede or destroy algae [112]. These fungi-induced allelochemicals can provoke oxidative stress in microalgae, interrupting their growth and disrupting cellular functions [113]. In particular, active metabolites can impair chlorophyll, a vital pigment for photosynthesis, in species such as Heterosigma akashiwo, potentially inhibiting the photosynthetic process [114].
A variety of algicidal fungi have been recognized, including Trametes versicolor F21a, Bjerkandera adusta T1, Lopharia spadicea, Phanerochaete chrysosporium, Trichoderma citrinoviride, and Irpex lacteus T2b. These fungi are known for their production of enzymes and secondary metabolites that can compromise algal cell integrity, obstruct photosynthesis, or trigger oxidative stress [105,106,107,108,109,110,111,112,113,114,115,116,117].
Observations indicate an uptick in algicidal fungi during the decline of algal blooms, suggesting their instrumental role in curbing and potentially reversing harmful algal bloom events by directly targeting overabundant algal growth [118].
Research has detailed the complex interactions between cyanobacterial cells and fungal mycelia, where cyanobacteria, both alive and dead, initially adhere to fungal mycelia before being overwhelmed by the mycelial network [119]. Specifically, the mycelia of P. chrysosporium are known to severely damage cyanobacterial cell membranes, with chlorophyll being a molecule particularly vulnerable to this assault [120].
An investigation by Gao et al. [78] into the algicidal mechanisms of Trametes versicolor F21a, using proteomic analysis, identified several biological processes, including glucan 1,4-α-glucosidase and hydrolase activities, as critical in the eradication of cyanobacterial cells. Additionally, this study implicated various metabolic pathways, such as glycolysis/gluconeogenesis and amino acid synthesis, in the algicidal process [74].
In a comprehensive study, the algicidal capabilities of four Aspergillus fungi species were explored, leading to the identification of eleven potent metabolites, notably five anthraquinones, four steroids, and two terpenoids, with anthraquinones being particularly effective against a range of algae [117].
White-rot fungi (WRF) are adept at degrading diverse organic pollutants, including chlorophenols, polycyclic aromatic hydrocarbons, and pesticides, significantly enhancing water quality [113]. This ability underscores the potential of WRF as an effective agent in combating excessive algal growth [118]. The degradation prowess of WRF is primarily due to their secretion of extracellular enzymes such as lignin peroxidase (LiP), manganese peroxidase (MnP), and laccase, which have a broad range of substrate affinities [119].
Fungi-Based Pelletization
Fungal pelletization emerges as an effective strategy to mitigate algal overabundance. This intricate process entails the symbiotic interaction between fungal and algal cells, leading to the formation and subsequent sedimentation of fungus–alga pellets [120]. Initiated by the swelling of spores and their germination, the process progresses with the expansive growth and branching of fungal hyphae [37]. These hyphae engage with algal cells, culminating in the creation of pellets. The pelletization process is subject to variations influenced by pH levels, salinity, and the rheological properties of the medium [121]. The resulting fungus–alga pellets exhibit diverse morphologies, from spherical to ellipsoidal, entwined with hyphae, and range in size from the microscopic scale to several millimeters [122]. In time, the fungal cells self-digest, leading to the breakdown of the pellets, thus synchronizing their life cycle with that of the fungi.
This innovative fungal pelletization technique is applied in the realms of wastewater treatment and the harvesting of microalgal biomass [123]. A prime example is the utilization of Cunninghamella echinulata NRRL 3655 fungal cells to efficiently collect Chlorella vulgaris UTEX 259 microalgal cells, forming fungus–alga pellets measuring 2 to 10 mm in diameter [124]. Furthermore, fungal bioflocculants, instrumental in microbial aggregation, significantly enhance fungal flocculation processes [37]. These bioflocculants, secreted by various filamentous fungi, are primarily composed of polysaccharides and proteins and have been effectively used in wastewater treatment and microalgal biomass harvesting [124]. Nonetheless, the full potential of these bioflocculants in managing HABs is yet to be realized, as challenges such as adjusting the pH of the bioflocculants and accommodating varying water conditions and species specificity remain to be addressed for practical HABs management [125].
4.4.6. Seaweeds for HABs Control
Seaweed, often called “macro-algae”, is essential in counteracting eutrophication by effectively absorbing surplus nutrients from bodies of water. This nutrient absorption is coupled with photosynthesis, where seaweed utilizes these nutrients and enriches the water with oxygen, a beneficial byproduct. Seaweed is a natural barrier against HABs by taking up excess nitrogen and phosphorus, thus maintaining water clarity, supporting marine life, and fostering ecological balance [125].
Additionally, certain cultivated seaweeds demonstrate exceptional productivity and can assimilate nitrogen, phosphorus, and even carbon dioxide [126]. The cultivation of seaweed on a large scale is being explored, with considerations of practicality, scientific and technological hurdles, ecological preservation, and financial feasibility [127]. In China, for instance, Laminaria japonica Aresch is extensively cultivated. It has been instrumental in mitigating the adverse effects associated with scallop aquaculture. Studies have pinpointed Gracilaria lemaneiformis and Porphyra haitanensis as effective for managing eutrophication along China’s southeastern to southern coasts [128].
Moreover, coastal waters are facing an increased rate of oxygen depletion, mainly due to eutrophication [129]. Research by Gao et al. [130] underscores the importance of seaweed cultivation in boosting oxygen levels in seawater, with an impressive annual oxygen output of 2,532,221 tonnes. Seaweed varieties like Porphyra yezoensis have been noted for their potential to diminish phosphorus and nitrogen concentrations, achieving reductions of 0.049 and 0.008 tons per hectare annually, respectively [131]. Additionally, Xiao et al. [132] have documented the successful nutrient removal from Chinese coastal waters using seaweed, which has led to better aquaculture outcomes by improving water quality and curbing algal blooms. Thus, introducing seaweeds to limit the nutrients available to microalgae emerges as a viable strategy for controlling algal blooms [133].
4.4.7. Viruses for HABs Control
Cyanophages, a specialized category of viruses, target cyanobacteria, distinguishing them from conventional bacteriophages. These viruses are characterized by their capacity to infect a wide array of cyanobacterial genera and their dependency on light for specific stages of their developmental cycle [133]. The genetic impact of cyanophages on their host cells is a focal point of interest, as some harbor genes that are integral to photosynthesis, potentially altering the photosynthetic efficiency of the cyanobacteria they infect [134]. Cyanophage research has concentrated on saline and freshwater aquatic environments [131]. A notable study has indicated genetic similarities between cyanophages in marine and freshwater systems [135]. Cyanophages such as Ma-LMM01, Ma-LBP, and VLP are known to specifically attack cyanobacteria that produce the toxin microcystin [136].
In the realm of combating HABs in aquatic ecosystems, antiviral treatments targeting algae have been deployed [137]. Cyanophage research is burgeoning, with discoveries continually advancing the field. Cyanophages like SM-1, SM-2, Ma-LBP, and those belonging to the Myoviridae family are known for their targeted action against M. aeruginosa [138]. The Siphoviridae viral family has been identified as an effective agent against Cylindrospermopsis raciborskii, initiating lytic cycles that could be a biological control for HABs [138]. Early 21st-century research has shown that the Cyanostyloviridae genus and S-PM2 are effective against Lyngbya majuscule, while other studies have reported cyanophages combating Synechococcus sp. [139]. SAM-1, in particular, has demonstrated a broader host range through specific species interactions [46].
A notable cyanophage, Ma-LEP, exemplifies cyanophages’ diversity and functional range. It is essential to recognize that certain phages, initially considered specific to M. aeruginosa, were susceptible to Synechococcus strains, highlighting a misclassification in identifying their cyanobacterial targets [140]. When naturally occurring viral communities were introduced into aquatic habitats, there was a dramatic 95% reduction in the host population within six days [141]. However, the population recovered within three weeks, possibly due to resistance development or ecological balance restoration [142].
The presence of cyanophages has been inversely related to cyanobacteria populations, suggesting that strategic, intermittent cyanophage introductions could temporarily suppress algal blooms under optimal conditions [143]. Moreover, cyanophages can modulate the metabolites produced by cyanobacteria, which may result in less detrimental blooms [142].
Conversely, the application of cyanophages for cyanobacterial management is challenging. Introducing cyanophages can lead to phage-resistant cyanobacterial strains. Additionally, the toxins released during the breakdown of blooms present another layer of complexity [143]. This could require the periodic introduction of new phages to combat recurring blooms. Furthermore, the toxins may be broken down by native microbial communities in the environment [111]. The sensitivity of bacteria to cyanophages varies, ranging from 0.1 to 32% as observed in the Lowland Dam Reservoir [133]. In some cases, cyanobacteria infected by phages have shown reduced photosynthesis and CO2-fixation capabilities [144].
4.4.8. Use of Biological Approaches at Lake
Biological strategies for lake restoration are emerging as a viable alternative to traditional physical and chemical methods. While effective in the short term, these conventional approaches, particularly for smaller bodies of water, do not tackle the fundamental problem of eutrophication and are often costly and partially effective. Chemical treatments, lacking a comprehensive treatment system, are prone to creating secondary pollutants and leaving behind residues and, thus, are typically reserved for urgent scenarios [145].
On the other hand, biological techniques provide a sustainable, economically viable option, albeit with a more extended timeframe required to see full results. For example, biomanipulation was employed as the exclusive treatment in Dongen, temporarily improving water quality. Conversely, no improvement was observed in Eindhoven, indicating that persistent phosphorus release from sediments may obstruct recovery efforts there [146].
Moreover, the combined use of bacteria, fungi, and viruses has shown promise in eradicating HABs, as depicted in Figure 8. This synergistic approach was exemplified when restoring the Xuxi River in Wuxi City, China, using bacterial and biological control methods. However, this treatment coincided with a significant increase in algal populations, underscoring the complexities of water pollution management [147].
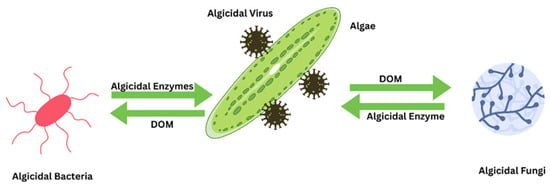
Figure 8.
Synergetic effect of algicidal bacteria, fungi, and viruses against HABs.
The application of this technology was tested in urban rivers like Xuxi and Gankeng to address water pollution. The effectiveness of BT was gauged by monitoring essential water quality parameters such as temperature, chemical oxygen demand (COD), and dissolved oxygen (DO) before and after the intervention [148].
Separately, the deployment of quagga mussels has been studied for their potential to combat algae, especially cyanobacteria, in urban ponds. The results indicated that quagga mussels could significantly reduce phytoplankton biomass, suggesting their utility in controlling algal blooms in urban water bodies [136].
Additionally, using aquatic plants for nutrient absorption present an environmentally friendly and cost-efficient method to counteract eutrophication. An assessment of six macrophyte species under various nitrogen and phosphorus concentrations showed notable nutrient reductions in a pond setting [149]. Moreover, ecological floating beds were found to remove nitrogen and phosphorus from nutrient-rich waters quickly. However, restoring microbial eukaryotic communities did not keep pace with the nutrient removal, likely due to the stabilizing effects of priority and rhizospheric micro-environments.
In a series of in situ experiments over three years on Lake Donghu in China, biomanipulation was tested for its effectiveness in reducing blooms. These studies revealed that an increase in filter-feeding fish populations, such as silver and bighead carp, was instrumental in clearing water blooms from the lake, as documented by Yu et al. [149].
4.5. Future Prospect
This research lays the groundwork for future exploration and practical application by evaluating the strengths and weaknesses of diverse chemical, physical, and biological methods for managing HABs. A critical future direction is the amalgamation of these approaches. Researchers and environmental organizations can craft specialized strategies to tackle specific bloom scenarios and ecological conditions by integrating chemical, physical, and biological methods. Such a comprehensive approach promises enhanced efficiency and effectiveness in controlling HABs. This study emphasizes the importance of understanding each control method’s long-term impacts, ecological threats, and potential unintended consequences. This insight is critical for guiding decision making and refining approaches to lessen adverse environmental effects.
Conducting field tests and validating these methods in real-world scenarios will be essential to future research. Field testing is necessary to confirm these strategies’ real-world efficacy and feasibility, thus narrowing the gap between theoretical research and practical implementation. International cooperation is essential in tackling the global challenge of HABs.
Additionally, the future of HABs management is intrinsically linked to technological advancements. Innovations could result in more accurate and targeted chemical treatments, more energy-efficient physical methods, and sophisticated biological control agents, thereby boosting the effectiveness and sustainability of HABs management strategies; future strategies must be flexible to adapt to evolving environmental conditions. As the demand for scientific research into HABs remains high, further practical application of emerging technologies are essential for effective HABs control.
4.6. Conclusions
Effectively addressing HABs necessitates a thorough and diverse approach involving various biological methods. This analysis underscores the absence of a universal solution, highlighting that each method has strengths and limitations. Successful mitigation in aquatic settings hinges on carefully assessing factors such as the water body’s specific characteristics and the algal bloom’s severity.
Biomanipulation, proven effective in extensive ecosystems, raises uncertainty about its suitability for smaller water bodies with no immediate results. Algicidal bacteria may benefit smaller systems, provided a comprehensive understanding and safe deployment are ensured. However, these methods do not guarantee complete HABs eradication, requiring ongoing applications. Phage therapy utilizing algicidal viruses presents a targeted and promising solution, albeit with the challenge of identifying suitable phages, especially in regions with prevalent algal species.
While not extensively researched, fungal interventions for HABs control exhibit potential in controlled settings or as part of a holistic HABs management plan. Determining viable HABs eradication methods for lakes and ponds depends on meticulously analyzing local conditions and algal types. Complemented by thorough monitoring, a synergistic application of these strategies often proves to be the most effective course of action.
Author Contributions
Conceptualization, H.M.A. and A.A.H.; methodology, H.M.A. and W.H.L.; software, H.M.A.; validation, H.M.A., A.A.H., W.H.L. and M.M.M.; formal analysis, H.M.A.; investigation, H.M.A. and A.A.H.; resources, A.A.H.; data curation, H.M.A.; writing—original draft preparation, H.M.A. and A.A.H.; writing—review and editing, H.M.A., A.A.H., W.H.L., A.A.-A. and M.M.M.; visualization, H.M.A.; supervision, A.A.H.; project administration, A.A.H.; funding acquisition, A.A.H. and A.A.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Water Center and United Arab Emirates University [grant number G00003661].
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a conflict of interest.
References
- Zhou, J.; Lao, Y.-M.; Song, J.-T.; Jin, H.; Zhu, J.-M.; Cai, Z.-H. Temporal heterogeneity of microbial communities and metabolic activities during a natural algal bloom. Water Res. 2020, 183, 116020. [Google Scholar] [CrossRef] [PubMed]
- Berdalet, E.; Fleming, L.E.; Gowen, R.; Davidson, K.; Hess, P.; Backer, L.C.; Moore, S.K.; Hoagland, P.; Enevoldsen, H. Marine harmful algal blooms, human health and wellbeing: Challenges and opportunities in the 21st century. J. Mar. Biol. Assoc. UK 2016, 96, 61–91. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.; Klaiber, H.A. Bloom and bust: Toxic algae’s impact on nearby property values. Ecol. Econ. 2017, 135, 209–221. [Google Scholar] [CrossRef]
- Zingone, A.; Enevoldsen, H.O. The diversity of harmful algal blooms: A challenge for science and management. Ocean. Coast. Manag. 2000, 43, 725–748. [Google Scholar] [CrossRef]
- Anderson, D.M. Turning back the harmful red tide. Nature 1997, 388, 513–514. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, G.; Zhu, X.; Chen, L.; Cai, Z. A review of the relationship between algae and bacteria in harmful algal blooms. Acta Ecol. Sin. 2014, 34, 269–281. [Google Scholar]
- Kramer, B.J.; Davis, T.W.; Meyer, K.A.; Rosen, B.H.; Goleski, J.A.; Dick, G.J.; Oh, G.; Gobler, C.J. Nitrogen limitation, toxin synthesis potential, and toxicity of cyanobacterial populations in Lake Okeechobee and the St. Lucie River Estuary, Florida, during the 2016 state of emergency event. PLoS ONE 2018, 13, e0196278. [Google Scholar] [CrossRef]
- Plaas, H.E.; Paerl, H.W. Toxic cyanobacteria: A growing threat to water and air quality. Environ. Sci. Technol. 2020, 55, 44–64. [Google Scholar] [CrossRef]
- Hallegraeff, G. Harmful algal blooms: A global overview. Man. Harmful Mar. Microalgae 2003, 33, 1–22. [Google Scholar]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef]
- Balaji-Prasath, B.; Wang, Y.; Su, Y.P.; Hamilton, D.P.; Lin, H.; Zheng, L.; Zhang, Y. Methods to control harmful algal blooms: A review. Environ. Chem. Lett. 2022, 20, 3133–3152. [Google Scholar] [CrossRef]
- Alayande, A.B.; Lim, J.; Kim, J.; Hong, S.; Al-Amoudi, A.S.; Park, B. Fouling control in SWRO desalination during harmful algal blooms: A historical review and future developments. Desalination 2022, 543, 116094. [Google Scholar] [CrossRef]
- Koreivienė, J.; Anne, O.; Kasperovičienė, J.; Burškytė, V. Cyanotoxin management and human health risk mitigation in recreational waters. Environ. Monit. Assess. 2014, 186, 4443–4459. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; He, Y.; Li, H.; Wei, Y.; Zhao, L.; Yang, G.; Chen, X. Using flocculation and subsequent biomanipulation to control microcystis blooms: A laboratory study. Harmful Algae 2020, 99, 101917. [Google Scholar] [CrossRef] [PubMed]
- Demeke, A. Cyanobacteria blooms and biological control methods. Int. J. Fauna Biol. Stud. 2016, 3, 32–38. [Google Scholar]
- Pal, M.; Yesankar, P.J.; Dwivedi, A.; Qureshi, A. Biotic control of harmful algal blooms (HABs): A brief review. J. Environ. Manag. 2020, 268, 110687. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Cunha, M.Â. Assessment of the microbiological quality of recreational waters: Indicators and methods. Euro-Mediterr. J. Environ. Integr. 2017, 2, 25. [Google Scholar] [CrossRef]
- Monsalve, E.R.; Quiroga, E. Farmed shrimp aquaculture in coastal wetlands of Latin America—A review of environmental issues. Mar. Pollut. Bull. 2022, 183, 113956. [Google Scholar] [CrossRef]
- Karlson, B.; Andersen, P.; Arneborg, L.; Cembella, A.; Eikrem, W.; John, U.; West, J.J.; Klemm, K.; Kobos, J.; Lehtinen, S.; et al. Harmful algal blooms and their effects in coastal seas of Northern Europe. Harmful Algae 2021, 102, 101989. [Google Scholar] [CrossRef]
- Jacquet, S.; Miki, T.; Noble, R.; Peduzzi, P.; Wilhelm, S. Viruses in aquatic ecosystems: Important advancements of the last 20 years and prospects for the future in the field of microbial oceanography and limnology. Adv. Oceanogr. Limnol. 2010, 1, 97–141. [Google Scholar] [CrossRef]
- Wojewódka, M.; Hruševar, D. The role of paleolimnology in climate and environment reconstruction and lake restoration in light of research on selected bioindicators. Holist. Approach Environ. 2020, 10, 16–28. [Google Scholar] [CrossRef]
- Gogoi, N.; Paul, B.; Purkayastha, K.D. Eco-bioengineering Tools in Ecohydrological Assessment of Eutrophic Water Bodies. Ecotoxicology 2022, 31, 581–601. [Google Scholar]
- Silverstein, M.R.; Segrè, D.; Bhatnagar, J.M. Environmental microbiome engineering for the mitigation of climate change. Glob. Chang. Biol. 2023, 29, 2050–2066. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.; Xiong, H.; Li, C.; Lu, Z.; Zhang, J.; Zhong, H.; Zhang, W.; Yan, B. Harmful algal blooms and their eco-environmental indication. Chemosphere 2021, 274, 129912. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Church, J.; Son, Y.; Kim, K.-T.; Lee, W.H. Recent advances in ultrasonic treatment: Challenges and field applications for controlling harmful algal blooms (HABs). Ultrason. Sonochemistry 2017, 38, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, S.S.U.H.; Yapa, N.; Karunarathna, S.C.; Suwannarach, N. Perceived intensification in harmful algal blooms is a wave of cumulative threat to the aquatic ecosystems. Biology 2022, 11, 852. [Google Scholar] [CrossRef]
- Chakraborty, S.; Moorthi, S.D.; Karnatak, R.; Feudel, U. Irregular harmful algal blooms triggered by feedback between toxin production and zooplankton feeding. Ecol. Model. 2022, 473, 110120. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Paerl, H.W. Mitigating harmful cyanobacterial blooms in a human-and climatically-impacted world. Life 2014, 4, 988–1012. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef]
- Griffith, A.W.; Gobler, C.J. Harmful algal blooms: A climate change co-stressor in marine and freshwater ecosystems. Harmful Algae 2020, 91, 101590. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.W.; Lazorchak, J.M.; Howard, M.D.; Johnson, M.-V.V.; Morton, S.L.; Perkins, D.A.; Reavie, E.D.; Scott, G.I.; Smith, S.A.; Steevens, J.A. Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems? Environ. Toxicol. Chem. 2016, 35, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Glibert, P.M.; Burford, M.A. Globally changing nutrient loads and harmful algal blooms: Recent advances, new paradigms, and continuing challenges. Oceanography 2017, 30, 58–69. [Google Scholar] [CrossRef]
- Zhan, M.-M.; Liu, P.-R.; Liu, X.-Y.; Hong, Y.; Xie, X. Inactivation and removal technologies for algal-bloom control: Advances and challenges. Curr. Pollut. Rep. 2021, 7, 392–406. [Google Scholar] [CrossRef]
- Anderson, D.M. Prevention, control and mitigation of harmful algal blooms: Multiple approaches to HAB management. Harmful Algae Manag. Mitig. 2004, 123–130. Available online: https://www.researchgate.net/publication/255649174_Prevention_control_and_mitigation_of_harmful_algal_blooms_multiple_approaches_to_HAB_management (accessed on 6 January 2024).
- McPartlin, D.A.; Loftus, J.H.; Crawley, A.S.; Silke, J.; Murphy, C.S.; O’Kennedy, R.J. Biosensors for the monitoring of harmful algal blooms. Curr. Opin. Biotechnol. 2017, 45, 164–169. [Google Scholar] [CrossRef]
- Sun, R.; Sun, P.; Zhang, J.; Esquivel-Elizondo, S.; Wu, Y. Microorganisms-based methods for harmful algal blooms control: A review. Bioresour. Technol. 2018, 248, 12–20. [Google Scholar] [CrossRef]
- Dyble, J.; Bienfang, P.; Dusek, E.; Hitchcock, G.; Holland, F.; Laws, E.; Lerczak, J.; McGillicuddy, D.J., Jr.; Minnett, P.; Moore, S.K.; et al. Environmental controls, oceanography and population dynamics of pathogens and harmful algal blooms: Connecting sources to human exposure. In Environmental Health; Springer: Berlin/Heidelberg, Germany, 2008; Volume 7, pp. 1–13. [Google Scholar]
- Li, S.; Tao, Y.; Zhan, X.-M.; Dao, G.-H.; Hu, H.-Y. UV-C irradiation for harmful algal blooms control: A literature review on effectiveness, mechanisms, influencing factors and facilities. Sci. Total Environ. 2020, 723, 137986. [Google Scholar] [CrossRef]
- Gallardo-Rodríguez, J.J.; Astuya-Villalón, A.; Llanos-Rivera, A.; Avello-Fontalba, V.; Ulloa-Jofré, V. A critical review on control methods for harmful algal blooms. Rev. Aquac. 2019, 11, 661–684. [Google Scholar] [CrossRef]
- Sierp, M.T.; Qin, J.G.; Recknagel, F. Biomanipulation: A review of biological control measures in eutrophic waters and the potential for Murray cod Maccullochella peelii peelii to promote water quality in temperate Australia. Rev. Fish Biol. Fish. 2009, 19, 143–165. [Google Scholar] [CrossRef]
- Visser, P.M.; Ibelings, B.W.; Bormans, M.; Huisman, J. Artificial mixing to control cyanobacterial blooms: A review. Aquat. Ecol. 2016, 50, 423–441. [Google Scholar] [CrossRef]
- Kibuye, F.A.; Zamyadi, A.; Wert, E.C. A critical review on operation and performance of source water control strategies for cyanobacterial blooms: Part II-mechanical and biological control methods. Harmful Algae 2021, 109, 102119. [Google Scholar] [CrossRef] [PubMed]
- Donaghay, P.L.; Osborn, T.R. Toward a theory of biological-physical control of harmful algal bloom dynamics and impacts. Limnol. Oceanogr. 1997, 42 Pt 2, 1283–1296. [Google Scholar] [CrossRef]
- Sanseverino, I.; Conduto, D.; Pozzoli, L.; Dobricic, S.; Lettieri, T. Algal bloom and its economic impact. Eur. Comm. Jt. Res. Cent. Inst. Environ. Sustain. 2016. Available online: https://data.europa.eu/doi/10.2788/660478 (accessed on 6 January 2024).
- Anderson, D.M. Approaches to monitoring, control and management of harmful algal blooms (HABs). Ocean. Coast. Manag. 2009, 52, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Marcuello, C. Present and future opportunities in the use of atomic force microscopy to address the physico-chemical properties of aquatic ecosystems at the nanoscale level. Int. Aquat. Res. 2022, 14, 231–240. [Google Scholar]
- Badagian, N.; Schirmer, M.P.; Parada, A.P.; Gonzalez-Sapienza, G.; Brena, B.M. Determination of Microcystins in Fish Tissue by ELISA and MALDI-TOF MS Using a Highly Specific Single Domain Antibody. Toxins 2023, 15, 84. [Google Scholar] [CrossRef]
- Stumpf, R.; Culver, M.; Tester, P.; Tomlinson, M.; Kirkpatrick, G.; Pederson, B.; Truby, E.; Ransibrahmanakul, V.; Soracco, M. Monitoring Karenia brevis blooms in the Gulf of Mexico using satellite ocean color imagery and other data. Harmful Algae 2003, 2, 147–160. [Google Scholar] [CrossRef]
- Ho, J.C.; Michalak, A.M.; Pahlevan, N. Widespread global increase in intense lake phytoplankton blooms since the 1980s. Nature 2019, 574, 667–670. [Google Scholar] [CrossRef]
- Baek, S.H.; Son, M.; Jung, S.W.; Na, D.H.; Cho, H.; Yamaguchi, M.; Kim, S.W.; Kim, Y.O. Enhanced species-specific chemical control of harmful and non-harmful algal bloom species by the thiazolidinedione derivative TD49. J. Appl. Phycol. 2014, 26, 311–321. [Google Scholar] [CrossRef]
- Liu, B.; Gai, S.; Lan, Y.; Cheng, K.; Yang, F. Metal-based adsorbents for water eutrophication remediation: A review of performances and mechanisms. Environ. Res. 2022, 212, 113353. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Gil-Izquierdo, A.; Pedreño, M.; Montoro-García, S.; Tárraga-Martínez, M.; Iglesias, P.; Ferreres, F.; Barceló, D.; Núñez-Delicado, E.; Gabaldón, J. A sustainable approach by using microalgae to minimize the eutrophication process of Mar Menor lagoon. Sci. Total Environ. 2021, 758, 143613. [Google Scholar] [CrossRef] [PubMed]
- Maris, J. Evaluating Rice Straw as a Substitute for Barley Straw in Inhibiting Algal Growth in Farm Ponds. Bachelor’s Thesis, University of Arkansa, Fayetteville, AR, USA, 2019. [Google Scholar]
- Iredale, R.S.; McDonald, A.T.; Adams, D.G. A series of experiments aimed at clarifying the mode of action of barley straw in cyanobacterial growth control. Water Res. 2012, 46, 6095–6103. [Google Scholar] [CrossRef] [PubMed]
- Eladel, H.; Battah, M.; Dawa, A.; Abd-Elhay, R.; Anees, D. Effect of rice straw extracts on growth of two phytoplankton isolated from a fish pond. J. Appl. Phycol. 2019, 31, 3557–3563. [Google Scholar] [CrossRef]
- Shumway, S.E.; Burkholder, J.M.; Morton, S.L. Harmful Algal Blooms: A Compendium Desk Reference; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Gedalanga, P.; Kotay, S.M.; Sales, C.M.; Butler, C.S.; Goel, R.; Mahendra, S. Novel applications of molecular biological and microscopic tools in environmental engineering. Water Environ. Res. 2013, 85, 917–950. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.; Burkholder, J.; Anderson, D.; Cochlan, W.; Dennison, W.; Dortch, Q.; Gobler, C.; Heil, C.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Wells, M.L.; Trainer, V.L.; Smayda, T.J.; Karlson, B.S.; Trick, C.G.; Kudela, R.M.; Ishikawa, A.; Bernard, S.; Wulff, A.; Anderson, D.M.; et al. Harmful algal blooms and climate change: Learning from the past and present to forecast the future. Harmful Algae 2015, 49, 68–93. [Google Scholar] [CrossRef]
- Theiling, C.H. A Review of Algal Phytoremediation Potential to Sequester Nutrients from Eutrophic Surface Water. 2023. Available online: https://erdc-library.erdc.dren.mil/jspui/handle/11681/47720 (accessed on 6 January 2024).
- Hoagland, P.; Scatasta, S. The economic effects of harmful algal blooms. Ecol. Harmful Algae 2006, 189, 391–402. [Google Scholar]
- Nicholls, K. A limnological basis for a Lake Simcoe phosphorus loading objective. Lake Reserv. Manag. 1997, 13, 189–198. [Google Scholar] [CrossRef]
- Salonen, K.; Sarvala, J.; Horppila, J.; Keto, J.; Malin, I.; Malinen, T.; Niemistö, J.; Ruuhijärvi, J. Development of Lake Vesijärvi through four decades of remediation efforts. Hydrobiologia 2020, 847, 4601–4619. [Google Scholar] [CrossRef]
- Wang, C.; Lu, Y.; Sun, B.; Zhang, M.; Mao, R.; Li, X.; Song, S.; Zhao, J.; Yu, M.; Shi, Y.; et al. Biomanipulation impacts on per-and polyfluoroalkyl substances accumulation and trophic transfer in an eutrophic lake. Environ. Int. 2022, 160, 107057. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, K.H.; Hwang, S.J.; Lee, T.K. Role of algal community stability in harmful algal blooms in river-connected lakes. Microb. Ecol. 2021, 82, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Davidson, T.A.; Liu, Z.; Mazzeo, N.; Trochine, C.; Özkan, K.; Jensen, H.S.; Trolle, D.; et al. Biomanipulation as a restoration tool to combat eutrophication: Recent advances and future challenges. Adv. Ecol. Res. 2012, 47, 411–488. [Google Scholar]
- Ren, L.; Gao, Y.; Hu, Z.; Jiang, X.; Yang, L. The Growth of Vallisneria natans and Its Epiphytic Biofilm in Simulated Nutrient-Rich Flowing Water. Water 2022, 14, 2236. [Google Scholar] [CrossRef]
- Ekvall, M.K.; Urrutia-Cordero, P.; Hansson, L.-A. Linking cascading effects of fish predation and zooplankton grazing to reduced cyanobacterial biomass and toxin levels following biomanipulation. PLoS ONE 2014, 9, e112956. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Cordero, P.; Ekvall, M.K.; Hansson, L.-A. Controlling harmful cyanobacteria: Taxa-specific responses of cyanobacteria to grazing by large-bodied Daphnia in a biomanipulation scenario. PLoS ONE 2016, 11, e0153032. [Google Scholar] [CrossRef]
- Jůza, T.; Duras, J.; Blabolil, P.; Sajdlová, Z.; Hess, J.; Chocholoušková, Z.; Kubečka, J. Recovery of the Velky Bolevecky pond (Plzen, Czech Republic) via biomanipulation–key study for management. Ecol. Eng. 2019, 136, 167–176. [Google Scholar] [CrossRef]
- Triest, L.; Stiers, I.; Van Onsem, S. Biomanipulation as a nature-based solution to reduce cyanobacterial blooms. Aquat. Ecol. 2016, 50, 461–483. [Google Scholar] [CrossRef]
- De Backer, S.; Teissier, S.; Triest, L. Stabilizing the clear-water state in eutrophic ponds after biomanipulation: Submerged vegetation versus fish recolonization. Hydrobiologia 2012, 689, 161–176. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Jacobsen, B.A.; Hansen, R.S.; Søndergaard, M.; Jensen, J.P.; Lauridsen, T.L.; Mazzeo, N. Restoration of shallow lakes by nutrient control and biomanipulation—The successful strategy varies with lake size and climate. Hydrobiologia 2007, 581, 269–285. [Google Scholar] [CrossRef]
- Van Colen, W.; Portilla, K.; Oña, T.; Wyseure, G.; Goethals, P.; Velarde, E.; Muylaert, K. Limnology of the neotropical high elevation shallow lake Yahuarcocha (Ecuador) and challenges for managing eutrophication using biomanipulation. Limnologica 2017, 67, 37–44. [Google Scholar] [CrossRef]
- Sayer, C.D.; Burgess, A.M.Y.; Kari, K.; Davidson, T.A.; Peglar, S.; Yang, H.; Rose, N. Long-term dynamics of submerged macrophytes and algae in a small and shallow, eutrophic lake: Implications for the stability of macrophyte-dominance. Freshw. Biol. 2010, 55, 565–583. [Google Scholar] [CrossRef]
- Gao, H.; Qian, X.; Wu, H.; Li, H.; Pan, H.; Han, C. Combined effects of submerged macrophytes and aquatic animals on the restoration of a eutrophic water body—A case study of Gonghu Bay, Lake Taihu. Ecol. Eng. 2017, 102, 15–23. [Google Scholar] [CrossRef]
- Wang, F.; Nishijima, W.; Uchida, Y.; Umehara, A.; Nakai, S.; Kasamo, K.; Shiraki, Y. Impact of eelgrass bed recovery and expansion on phytoplankton growth through nutrient competition. J. Environ. Manag. 2020, 260, 109898. [Google Scholar] [CrossRef] [PubMed]
- Declerck, S.; Vanderstukken, M.; Pals, A.; Muylaert, K.; Meester, L.D. Plankton biodiversity along a gradient of productivity and its mediation by macrophytes. Ecology 2007, 88, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Švanys, A.; Paškauskas, R.; Hilt, S. Effects of the allelopathically active macrophyte Myriophyllum spicatum on a natural phytoplankton community: A mesocosm study. Hydrobiologia 2014, 737, 57–66. [Google Scholar] [CrossRef]
- Zhu, X.; Dao, G.; Tao, Y.; Zhan, X.; Hu, H. A review on control of harmful algal blooms by plant-derived allelochemicals. J. Hazard. Mater. 2021, 401, 123403. [Google Scholar] [CrossRef]
- Boylan, J.D.; Morris, J.E. Limited effects of barley straw on algae and zooplankton in a midwestern pond. Lake Reserv. Manag. 2003, 19, 265–271. [Google Scholar] [CrossRef][Green Version]
- West, M.; Fenner, N.; Gough, R.; Freeman, C. Evaluation of algal bloom mitigation and nutrient removal in floating constructed wetlands with different macrophyte species. Ecol. Eng. 2017, 108, 581–588. [Google Scholar] [CrossRef]
- Szabó, S.; Peeters, E.; Várbíró, G.; Borics, G.; Lukács, B.A. Phenotypic plasticity as a clue for invasion success of the submerged aquatic plant Elodea nuttallii. Plant Biol. 2019, 21, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Mowe, M.A.; Song, Y.; Sim, D.Z.; Lu, J.; Mitrovic, S.M.; Tan, H.T.; Yeo, D.C. Comparative study of six emergent macrophyte species for controlling cyanobacterial blooms in a tropical reservoir. Ecol. Eng. 2019, 129, 11–21. [Google Scholar] [CrossRef]
- Dai, Y.; Jia, C.; Liang, W.; Hu, S.; Wu, Z. Effects of the submerged macrophyte Ceratophyllum demersum L. on restoration of a eutrophic waterbody and its optimal coverage. Ecol. Eng. 2012, 40, 113–116. [Google Scholar] [CrossRef]
- Haggard, K.G.; Geiger, N.S.; Hayes, P.M.; Milligan, A.J. Suppression of cyanobacterial growth of Aphanizomenon flos-aquae by vascular plant decomposition products in Upper Klamath Lake, Oregon. Lake Reserv. Manag. 2013, 29, 13–22. [Google Scholar] [CrossRef]
- Cooke, G.D.; Welch, E.B.; Peterson, S.; Nichols, S.A. Restoration and Management of Lakes and Reservoirs; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Hongying, H.; Yu, H. Algal-bloom control by allelopathy of aquatic macrophytes-A review. Front. Environ. Sci. Eng. 2008, 2, 421–438. [Google Scholar]
- Ferrier, M.; Sr, B.B.; Terlizzi, D.; Lacouture, R. The effects of barley straw (Hordeum vulgare) on the growth of freshwater algae. Bioresour. Technol. 2005, 96, 1788–1795. [Google Scholar] [CrossRef]
- Hu, J.; Kokoette, E.; Xu, C.; Huang, S.; Tang, T.; Zhang, Y.; Liu, M.; Huang, Y.; Yu, S.; Zhu, J.; et al. Natural Algaecide Sphingosines Identified in Hybrid Straw Decomposition Driven by White-Rot Fungi. Adv. Sci. 2023, 10, 2300569. [Google Scholar] [CrossRef]
- Mohan, S.M. Comparative study of rice straw and ragi straw for the inhibition of algal bloom in fresh water. Int. Res. J. Biol. Sci. 2012, 1, 31–37. [Google Scholar]
- Alam, M.A.; Vandamme, D.; Chun, W.; Zhao, X.; Foubert, I.; Wang, Z.; Muylaert, K.; Yuan, Z. Bioflocculation as an innovative harvesting strategy for microalgae. Rev. Environ. Sci. Bio/Technol. 2016, 15, 573–583. [Google Scholar] [CrossRef]
- Chen, H.; Zhong, C.; Berkhouse, H.; Zhang, Y.; Lv, Y.; Lu, W.; Yang, Y.; Zhou, J. Removal of cadmium by bioflocculant produced by Stenotrophomonas maltophilia using phenol-containing wastewater. Chemosphere 2016, 155, 163–169. [Google Scholar] [CrossRef]
- Powell, R.J.; Hill, R.T. Mechanism of algal aggregation by Bacillus sp. strain RP1137. Appl. Environ. Microbiol. 2014, 80, 4042–4050. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.-F.; Lin, H.; Wang, G.; Lu, L.-L.; Zhao, Y.-H. Preparation of a new-style composite containing a key bioflocculant produced by Pseudomonas aeruginosa ZJU1 and its flocculating effect on harmful algal blooms. J. Hazard. Mater. 2015, 284, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, H.; Fu, L.; An, X.; Zhang, B.; Li, Y.; Chen, Z.; Zheng, W.; Yi, L.; Zheng, T. A novel algicide: Evidence of the effect of a fatty acid compound from the marine bacterium, Vibrio sp. BS02 on the harmful dinoflagellate, Alexandrium tamarense. PLoS ONE 2014, 9, e91201. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huo, M.; Sun, C.; Cui, X.; Zhou, D.; Crittenden, J.C.; Yang, W. Bioresources inner-recycling between bioflocculation of Microcystis aeruginosa and its reutilization as a substrate for bioflocculant production. Sci. Rep. 2017, 7, 43784. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lei, X.; Zhu, H.; Zhang, H.; Guan, C.; Chen, Z.; Zheng, W.; Fu, L.; Zheng, T. Chitinase producing bacteria with direct algicidal activity on marine diatoms. Sci. Rep. 2016, 6, 21984. [Google Scholar] [CrossRef]
- Shilo, M. Lysis of blue-green algae by myxobacter. J. Bacteriol. 1970, 104, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, X.; Zhou, Y.; Zheng, W.; Yu, C.; Zheng, T. Novel insights into the algicidal bacterium DH77-1 killing the toxic dinoflagellate Alexandrium tamarense. Sci. Total Environ. 2014, 482, 116–124. [Google Scholar] [CrossRef]
- Jeong, S.-Y.; Ishida, K.; Ito, Y.; Okada, S.; Murakami, M. Bacillamide, a novel algicide from the marine bacterium, Bacillus sp. SY-1, against the harmful dinoflagellate, Cochlodinium polykrikoides. Tetrahedron Lett. 2003, 44, 8005–8007. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, H.; Lei, X.; Zhang, H.; Guan, C.; Chen, Z.; Zheng, W.; Xu, H.; Tian, Y.; Yu, Z.; et al. The first evidence of deinoxanthin from Deinococcus sp. Y35 with strong algicidal effect on the toxic dinoflagellate Alexandrium tamarense. J. Hazard. Mater. 2015, 290, 87–95. [Google Scholar] [CrossRef]
- Seong, K.A.; Jeong, H.J. Interactions between marine bacteria and red tide organisms in Korean waters. Algae 2013, 28, 297–305. [Google Scholar] [CrossRef]
- Inaba, N.; Watanabe, T.; Sakami, T.; Nishi, H.; Tahara, Y.; Imai, I. Temporal and spatial distribution of algicidal and growth-inhibiting bacteria in the coastal sea of southwest Japan. J. Plankton Res. 2014, 36, 388–397. [Google Scholar] [CrossRef]
- Scherer, P.I.; Millard, A.D.; Miller, A.; Schoen, R.; Raeder, U.; Geist, J.; Zwirglmaier, K. Temporal dynamics of the microbial community composition with a focus on toxic cyanobacteria and toxin presence during harmful algal blooms in two South German lakes. Front. Microbiol. 2017, 8, 2387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhai, C.; Chen, R.; Liu, C.; Xue, Y.; Jiang, J. The dynamics of the water bloom-forming Microcystis aeruginosa and its relationship with biotic and abiotic factors in Lake Taihu, China. Ecol. Eng. 2012, 47, 274–277. [Google Scholar] [CrossRef]
- Gumbo, R.J.; Ross, G.; Cloete, E.T. Biological control of Microcystis dominated harmful algal blooms. Afr. J. Biotechnol. 2008, 7. Available online: https://www.researchgate.net/publication/237465093_Biological_control_of_Microcystis_dominated_harmful_algal_blooms (accessed on 6 January 2024).
- Alamri, S.A.; Mohamed, Z.A. Selective inhibition of toxic cyanobacteria by β-carboline-containing bacterium Bacillus flexus isolated from Saudi freshwaters. Saudi J. Biol. Sci. 2013, 20, 357–363. [Google Scholar] [CrossRef]
- Zhang, J.-T.; Yang, F.; Du, K.; Li, W.-F.; Chen, Y.; Jiang, Y.-L.; Li, Q.; Zhou, C.-Z. Structure and assembly pattern of a freshwater short-tailed cyanophage Pam1. Structure 2022, 30, 240–251.e4. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Ma, H.; Ren, S.; Gao, X.; He, X.; Zhu, S.; Deng, R.; Zhang, S. Insights into the mechanism of cyanobacteria removal by the algicidal fungi Bjerkandera adusta and Trametes versicolor. MicrobiologyOpen 2020, 9, e1042. [Google Scholar] [CrossRef]
- Miazek, K.; Remacle, C.; Richel, A.; Goffin, D. Effect of lignocellulose related compounds on microalgae growth and product biosynthesis: A review. Energies 2014, 7, 4446–4481. [Google Scholar] [CrossRef]
- Dai, W.; Chen, X.; Wang, X.; Xu, Z.; Gao, X.; Jiang, C.; Deng, R.; Han, G. The algicidal fungus Trametes versicolor F21a eliminating blue algae via genes encoding degradation enzymes and metabolic pathways revealed by transcriptomic analysis. Front. Microbiol. 2018, 9, 826. [Google Scholar] [CrossRef]
- Wang, Q.; Su, M.; Zhu, W.; Li, X.; Jia, Y.; Guo, P.; Chen, Z.; Jiang, W.; Tian, X. Growth inhibition of Microcystis aeruginosa by white-rot fungus Lopharia spadicea. Water Sci. Technol. 2010, 62, 317–323. [Google Scholar] [CrossRef]
- Zeng, G.; Zhang, M.; Wang, P.; Li, X.; Wu, P.; Sun, D. Genotoxicity effects of Phanerochaete chrysosporium against harmful algal bloom species by micronucleus test and comet assay. Chemosphere 2019, 218, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Zuo, J.; Liu, X.; Ji, N. Algicidal activities of secondary metabolites of marine macroalgal-derived endophytic fungi. J. Oceanol. Limnol. 2019, 37, 112–121. [Google Scholar] [CrossRef]
- Tortella, G.; Durán, N.; Rubilar, O.; Parada, M.; Diez, M. Are white-rot fungi a real biotechnological option for the improvement of environmental health? Crit. Rev. Biotechnol. 2015, 35, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.W. Structure and action mechanism of ligninolytic enzymes. Appl. Biochem. Biotechnol. 2009, 157, 174–209. [Google Scholar] [CrossRef] [PubMed]
- Heitner, C.; Dimmel, D.; Schmidt, J. Lignin and Lignans: Advances in Chemistry; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Grimm, L.; Kelly, S.; Völkerding, I.; Krull, R.; Hempel, D. Influence of mechanical stress and surface interaction on the aggregation of Aspergillus niger conidia. Biotechnol. Bioeng. 2005, 92, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Cheng, Y.; Li, Y.; Wan, Y.; Liu, Y.; Lin, X.; Ruan, R. Novel fungal pelletization-assisted technology for algae harvesting and wastewater treatment. Appl. Biochem. Biotechnol. 2012, 167, 214–228. [Google Scholar] [CrossRef]
- Tyagi, Y.R.; Surampalli, R. Isolation of extra cellular biopolymer producing microorganisms from wastewater sludge for sludge settling and dewatering. In Proceedings of the WEFTEC 2006, Dallas, TX, USA, 21–25 October 2006; Water Environment Federation: Alexandria, VA, USA, 2006; pp. 473–489. [Google Scholar]
- Xie, S.; Sun, S.; Dai, S.Y.; Yuan, J.S. Efficient coagulation of microalgae in cultures with filamentous fungi. Algal Res. 2013, 2, 28–33. [Google Scholar] [CrossRef]
- Aljuboori, A.H.R.; Idris, A.; Abdullah, N.; Mohamad, R. Production and characterization of a bioflocculant produced by Aspergillus flavus. Bioresour. Technol. 2013, 127, 489–493. [Google Scholar] [CrossRef]
- Zheng, Y.; Jin, R.; Zhang, X.; Wang, Q.; Wu, J. The considerable environmental benefits of seaweed aquaculture in China. Stoch. Environ. Res. Risk Assess. 2019, 33, 1203–1221. [Google Scholar] [CrossRef]
- Xu, S.; Yu, Z.; Zhou, Y.; Yue, S.; Liang, J.; Zhang, X. The potential for large-scale kelp aquaculture to counteract marine eutrophication by nutrient removal. Mar. Pollut. Bull. 2023, 187, 114513. [Google Scholar] [CrossRef]
- Fei, X. Solving the coastal eutrophication problem by large scale seaweed cultivation. In Asian Pacific Phycology in the 21st Century: Prospects and Challenges: Proceeding of The Second Asian Pacific Phycological Forum, held in Hong Kong, China, 21–25 June 1999; Springer: Berlin/Heidelberg, Germany, 2004; pp. 145–151. [Google Scholar]
- Claret, M.; Galbraith, E.D.; Palter, J.B.; Bianchi, D.; Fennel, K.; Gilbert, D.; Dunne, J.P. Rapid coastal deoxygenation due to ocean circulation shift in the northwest Atlantic. Nat. Clim. Chang. 2018, 8, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Gao, L.; Jiang, M.; Jian, A.; He, L. The potential of seaweed cultivation to achieve carbon neutrality and mitigate deoxygenation and eutrophication. Environ. Res. Lett. 2021, 17, 014018. [Google Scholar] [CrossRef]
- He, P.; Xu, S.; Zhang, H.; Wen, S.; Dai, Y.; Lin, S.; Yarish, C. Bioremediation efficiency in the removal of dissolved inorganic nutrients by the red seaweed, Porphyra yezoensis, cultivated in the open sea. Water Res. 2008, 42, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Agusti, S.; Lin, F.; Li, K.; Pan, Y.; Yu, Y.; Zheng, Y.; Wu, J.; Duarte, C.M. Nutrient removal from Chinese coastal waters by large-scale seaweed aquaculture. Sci. Rep. 2017, 7, 46613. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Murase, J.; Asakawa, S.; Kimura, M. Novel cyanophage photosynthetic gene psbA in the floodwater of a Japanese rice field. FEMS Microbiol. Ecol. 2009, 70, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.W.; Bullerjahn, G.S.; Eldridge, M.L.; Rinta-Kanto, J.M.; Poorvin, L.; Bourbonniere, R.A. Seasonal hypoxia and the genetic diversity of prokaryote populations in the central basin hypolimnion of Lake Erie: Evidence for abundant cyanobacteria and photosynthesis. J. Great Lakes Res. 2006, 32, 657–671. [Google Scholar] [CrossRef]
- Zhang, D.; He, Y.; Gin, K.Y.-H. Novel freshwater cyanophages provide new insights into evolutionary relationships between freshwater and marine cyanophages. Microbiol. Spectr. 2021, 9, e00593-21. [Google Scholar] [CrossRef]
- Waajen, G.W.; Van Bruggen, N.C.; Pires, L.M.D.; Lengkeek, W.; Lürling, M. Biomanipulation with quagga mussels (Dreissena rostriformis bugensis) to control harmful algal blooms in eutrophic urban ponds. Ecol. Eng. 2016, 90, 141–150. [Google Scholar] [CrossRef]
- Rong, C.; Zhou, K.; Li, S.; Xiao, K.; Xu, Y.; Zhang, R.; Yang, Y.; Zhang, Y. Isolation and characterization of a novel cyanophage encoding multiple auxiliary metabolic genes. Viruses 2022, 14, 887. [Google Scholar] [CrossRef] [PubMed]
- Mankiewicz-Boczek, J.; Jaskulska, A.; Pawełczyk, J.; Gągała, I.; Serwecińska, L.; Dziadek, J. Cyanophages infection of Microcystis bloom in lowland dam reservoir of Sulejów, Poland. Microb. Ecol. 2016, 71, 315–325. [Google Scholar] [CrossRef]
- Mann, N.H. Phages of the marine cyanobacterial picophytoplankton. FEMS Microbiol. Rev. 2003, 27, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Takashima, Y.; Tomaru, Y.; Shirai, Y.; Takao, Y.; Hiroishi, S.; Nagasaki, K. Isolation and characterization of a cyanophage infecting the toxic cyanobacterium Microcystis aeruginosa. Appl. Environ. Microbiol. 2006, 72, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Yoshida-Takashima, Y.; Yoshida, M.; Ogata, H.; Nagasaki, K.; Hiroishi, S.; Yoshida, T. Cyanophage infection in the bloom-forming cyanobacteria Microcystis aeruginosa in surface freshwater. Microbes Environ. 2012, 27, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Engel, B.A.; Reuhs, M.; Simsek, H. Cyanophage technology in removal of cyanobacteria mediated harmful algal blooms: A novel and eco-friendly method. Chemosphere 2023, 315, 137769. [Google Scholar] [CrossRef] [PubMed]
- Coloma, S.; Dienstbier, A.; Bamford, D.; Sivonen, K.; Roine, E.; Hiltunen, T. Newly isolated Nodularia phage influences cyanobacterial community dynamics. Environ. Microbiol. 2017, 19, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Antosiak, A.; Šulčius, S.; Malec, P.; Tokodi, N.; Łobodzińska, A.; Dziga, D. Cyanophage infections reduce photosynthetic activity and expression of CO2 fixation genes in the freshwater bloom-forming cyanobacterium Aphanizomenon flos-aquae. Harmful Algae 2022, 116, 102215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, P.; Zhao, S.; Kang, S.; Wang, P.; Zhou, M.; Lyu, J. Control and remediation methods for eutrophic lakes in the past 30 years. Water Sci. Technol. 2020, 81, 1099–1113. [Google Scholar] [CrossRef]
- Waajen, G.; van Oosterhout, F.; Douglas, G.; Lürling, M. Geo-engineering experiments in two urban ponds to control eutrophication. Water Res. 2016, 97, 69–82. [Google Scholar] [CrossRef]
- Yudianto, D.; Yuebo, X. Numerical modeling and practical experience of Xuxi River’s natural restoration using biological treatment. Water Environ. Res. 2011, 83, 2087–2098. [Google Scholar] [CrossRef]
- Hashim, S.; Yuebo, X.; Saifullah, M.; Jan, R.N.; Muhetaer, A. Integrated evaluation of urban water bodies for pollution abatement based on fuzzy multicriteria decision approach. BioMed Res. Int. 2015, 2015, 327280. [Google Scholar] [CrossRef]
- Yu, S.; Miao, C.; Song, H.; Huang, Y.; Chen, W.; He, X. Efficiency of nitrogen and phosphorus removal by six macrophytes from eutrophic water. Int. J. Phytoremediation 2019, 21, 643–651. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).