
Use of Telemetry Data to Quantify Life History Diversity in Migrating Juvenile Chinook Salmon (Oncorhynchus tshawytscha)

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
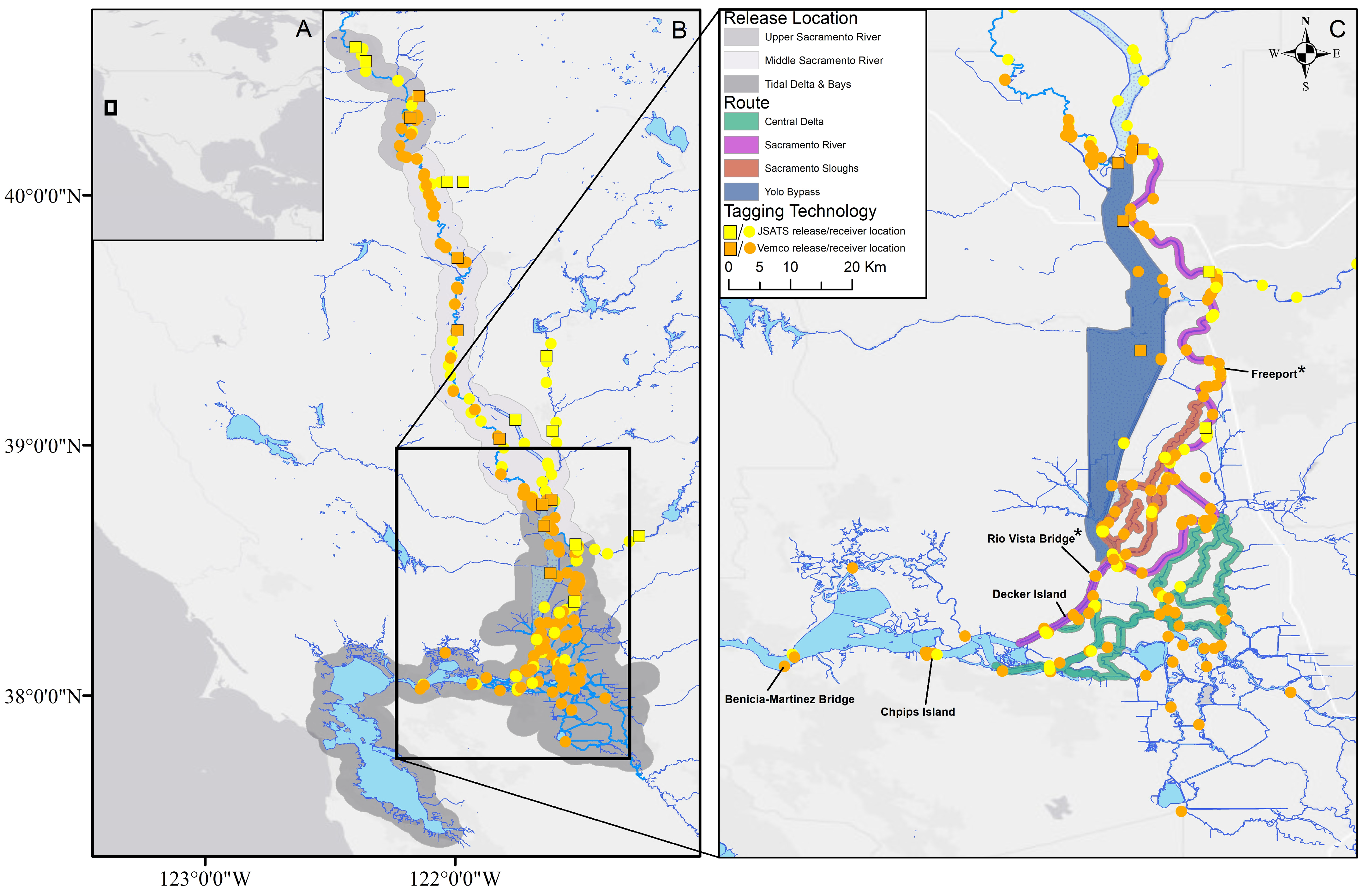
2.1. Study System
2.2. Data Source and Integration
2.3. Outmigration Routing Assignment
2.4. Estimating Travel Time to the Estuary
2.5. Variation in Downstream Movement
3. Results
3.1. Routing
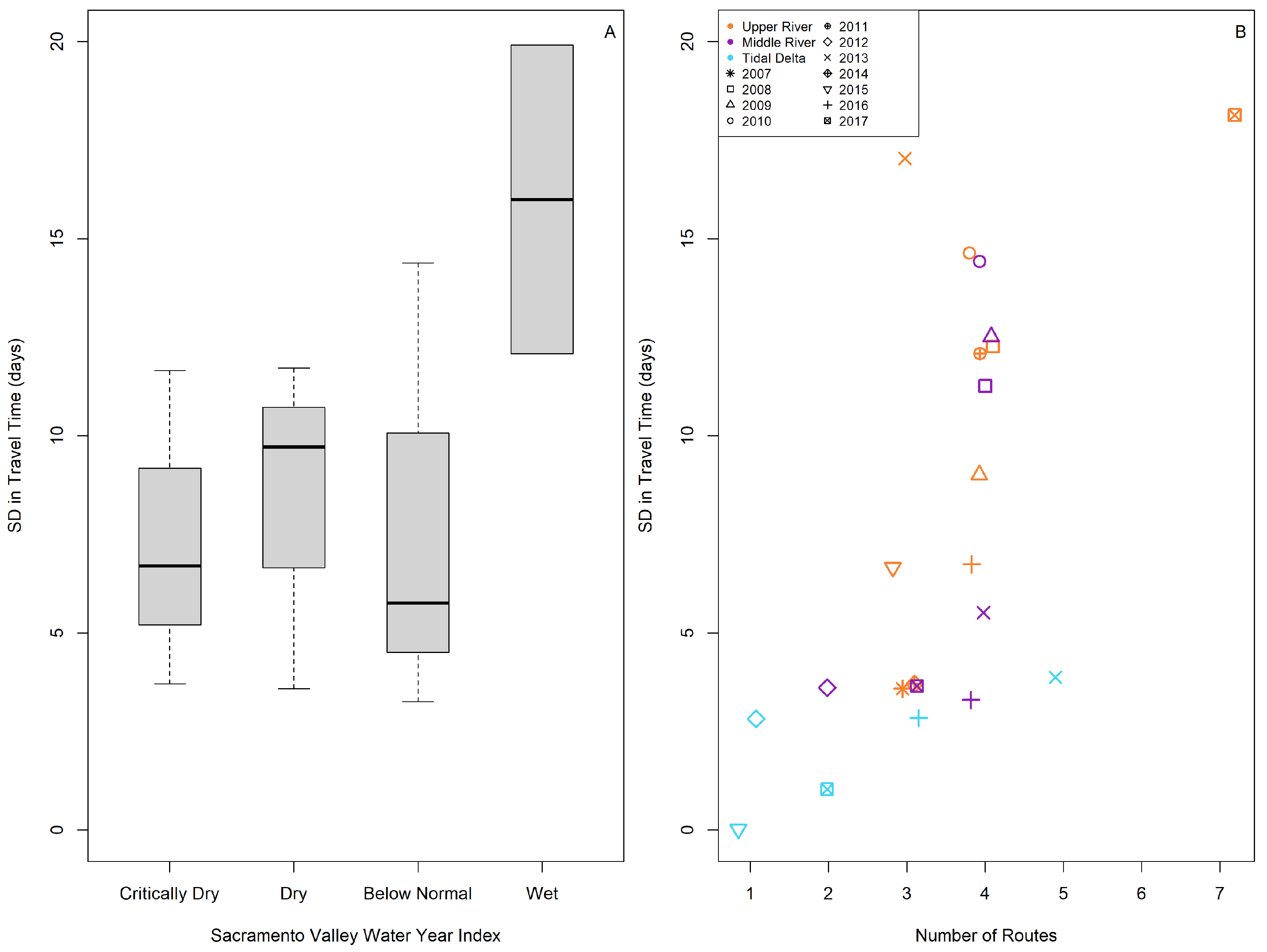
3.2. Travel Time to the Estuary
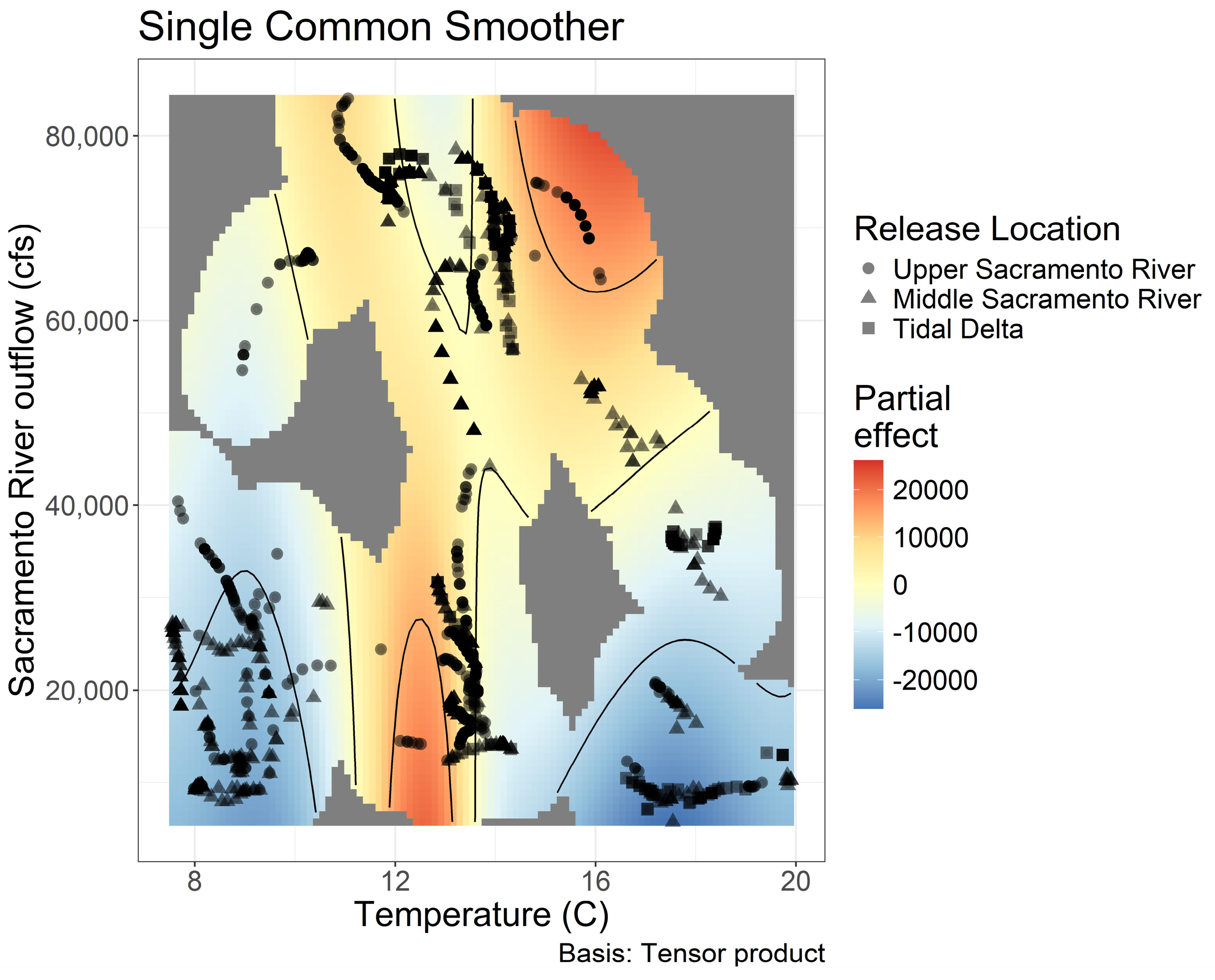
3.3. Variation in Downstream Movement
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cushing, D.H. Plankton Production and Year-Class Strength in Fish Populations: An Update of the Match/Mismatch Hypothesis. Adv. Mar. Biol. 1990, 26, 249–293. [Google Scholar]
- Laurel, B.J.; Hurst, T.P.; Ciannelli, L. An Experimental Examination of Temperature Interactions in the Match—Mismatch Hypothesis. Can. J. Fish. Aquat. Sci. 2011, 68, 51–61. [Google Scholar] [CrossRef]
- Lindley, S.T.; Grimes, C.B.; Mohr, M.S.; Peterson, W.; Stein, J.E.; Anderson, J.T.; Botsford, L.W.; Bottom, D.L.; Busack, C.A.; Collier, T.K.; et al. What Caused the Sacramento River Fall Chinook Stock Collapse? 2009. Available online: https://www.waterboards.ca.gov/waterrights/water_issues/programs/bay_delta/deltaflow/docs/exhibits/nmfs/spprt_docs/nmfs_exh4_lindley_etal_2009.pdf (accessed on 15 January 2020).
- Woodson, L.E.; Wells, B.K.; Weber, P.K.; Macfarlane, R.B.; Whitman, G.E.; Johnson, R.C. Size, Growth, and Origin-Dependent Mortality of Juvenile Chinook Salmon Oncorhynchus tshawytscha during Early Ocean Residence. Mar. Ecol. Prog. Ser. 2013, 487, 163–175. [Google Scholar] [CrossRef]
- Budy, P.; Thiede, G.P.; Bouwes, N.; Petrosky, C.E.; Schaller, H. Evidence Linking Delayed Mortality of Snake River Salmon to Their Earlier Hydrosystem Experience. N. Am. J. Fish. Manag. 2002, 22, 35–51. [Google Scholar] [CrossRef]
- Crozier, L.G.; Bowerman, T.E.; Burke, B.J.; Keefer, M.L.; Caudill, C.C. High-stakes steeplechase: A behavior-based model to predict individual travel times through diverse migration segments. Ecosphere 2017, 8, e01965. [Google Scholar] [CrossRef]
- Bottom, D.L.; Jones, K.K.; Cornwell, T.J.; Gray, A.; Simenstad, C.A. Patterns of Chinook Salmon Migration and Residency in the Salmon River Estuary (Oregon). Estuar. Coast. Shelf Sci. 2005, 64, 79–93. [Google Scholar] [CrossRef]
- Satterthwaite, W.H.; Carlson, S.M.; Allen-Moran, S.D.; Vincenzi, S.; Bograd, S.J.; Wells, B.K. Match-Mismatch Dynamics and the Relationship between Ocean-Entry Timing and Relative Ocean Recoveries of Central Valley Fall Run Chinook Salmon. Mar. Ecol. Prog. Ser. 2014, 511, 237–248. [Google Scholar] [CrossRef]
- Schreck, C.B.; Moyle, P.B. (Eds.) Methods for Fish Biology; American Fisheries Society: Bethesda, MD, USA, 2002. [Google Scholar]
- Greene, C.M.; Hall, J.E.; Guilbault, K.R.; Quinn, T.P. Improved Viability of Populations with Diverse Life-History Portfolios. Biol. Lett. 2010, 6, 382–386. [Google Scholar] [CrossRef]
- Moore, J.W.; Yeakel, J.D.; Peard, D.; Lough, J.; Beere, M. Life-History Diversity and Its Importance to Population Stability and Persistence of a Migratory Fish: Steelhead in Two Large North American Watersheds. J. Anim. Ecol. 2014, 83, 1035–1046. [Google Scholar] [CrossRef]
- Schindler, D.E.; Hilborn, R.; Chasco, B.; Boatright, C.P.; Quinn, T.P.; Rogers, L.A.; Webster, M.S. Population Diversity and the Portfolio Effect in an Exploited Species. Nature 2010, 465, 609–612. [Google Scholar] [CrossRef]
- National Marine Fisheries Service. Biological and Conference Opinion on the Long-Term Operations of the Central Valley Project and State Water Project; National Marine Fisheries Service: Silver Spring, MD, USA, 2009.
- Rand, P.S.; Goslin, M.; Gross, M.R.; Irvine, J.R.; Augerot, X.; Mchugh, P.A.; Bugaev, V.F. Global Assessment of Extinction Risk to Populations of Sockeye Salmon Oncorhynchus nerka. PLoS ONE 2012, 7, e34065. [Google Scholar] [CrossRef] [PubMed]
- Satterthwaite, W.H.; Carlson, S.M. Weakening Portfolio Effect Strength in a Hatchery-Supplemented Chinook Salmon Population Complex. Can. J. Fish. Aquat. Sci. 2015, 72, 1860–1875. [Google Scholar] [CrossRef]
- Larsen, D.A.; Harstad, D.L.; Fuhrman, A.E.; Knudsen, C.M.; Schroder, S.L.; Bosch, W.J.; Galbreath, P.F.; Fast, D.E.; Beckman, B.R. Maintaining a Wild Phenotype in a Conservation Hatchery Program for Chinook Salmon: The Effect of Managed Breeding on Early Male Maturation. PLoS ONE 2019, 14, e0216168. [Google Scholar] [CrossRef]
- Hedrick, P.W.; Hedgecock, D.; Hamelberg, S. Effective Population Size in Winter-Run Chinook Salmon. Conserv. Biol. 1995, 9, 615–624. [Google Scholar] [CrossRef]
- Utter, F.; Milner, G.; Stahl, G.; Teel, D. Genetic Population Structure of Chinook Salmon, Oncorhynchus tshawytscha, in the Pacific Northwest. Fish. Bull. 1989, 87, 239–264. [Google Scholar]
- Thompson, T.Q.; Bellinger, M.R.; O’Rourke, S.M.; Prince, D.J.; Stevenson, A.E.; Rodrigues, A.T.; Sloat, M.R.; Speller, C.F.; Yang, D.Y.; Butler, V.L.; et al. Anthropogenic Habitat Alteration Leads to Rapid Loss of Adaptive Variation and Restoration Potential in Wild Salmon Populations. Proc. Natl. Acad. Sci. USA 2019, 116, 177–186. [Google Scholar] [CrossRef]
- Narum, S.R.; Genova, A.D.; Micheletti, S.J.; Maass, A. Genomic Variation Underlying Complex Life-History Traits Revealed by Genome Sequencing in Chinook Salmon. Proc. R. Soc. B 2018, 285, 20180935. [Google Scholar] [CrossRef]
- Johnson, R.C.; Garza, J.C.; Macfarlane, R.B.; Grimes, C.B.; Phillis, C.C.; Koch, P.L.; Weber, P.K.; Carr, M.H. Isotopes and Genes Reveal Freshwater Origins of Chinook Salmon Oncorhynchus tshawytscha Aggregations in California’s Coastal Ocean. Mar. Ecol. Prog. Ser. 2016, 548, 181–196. [Google Scholar] [CrossRef]
- Cordoleani, F.; Phillis, C.C.; Sturrock, A.M.; FitzGerald, A.M.; Malkassian, A.; Whitman, G.E.; Weber, P.K.; Johnson, R.C. Threatened salmon rely on a rare life history strategy in a warming landscape. Nat. Clim. Chang. 2021, 11, 982–988. [Google Scholar] [CrossRef]
- Phillis, C.C.; Sturrock, A.M.; Johnson, R.C.; Weber, P.K. Endangered Winter-Run Chinook Salmon Rely on Diverse Rearing Habitats in a Highly Altered Landscape. Biol. Conserv. 2018, 217, 358–362. [Google Scholar] [CrossRef]
- Sturrock, A.M.; Wikert, J.D.; Heyne, T.; Mesick, C.; Hubbard, A.E.; Hinkelman, T.M.; Weber, P.K.; Whitman, G.E.; Glessner, J.J.; Johnson, R.C. Reconstructing the Migratory Behavior and Long-Term Survivorship of Juvenile Chinook Salmon under Contrasting Hydrologic Regimes. PLoS ONE 2015, 10, e0122380. [Google Scholar] [CrossRef] [PubMed]
- Adams, N.S.; Beeman, J.W.; Eiler, J.H. (Eds.) Telemetry Techniques: A User Guide for Fisheries Research; American Fisheries Society: Bethesda, MD, USA, 2012. [Google Scholar]
- McMichael, G.A.; Eppard, M.B.; Carlson, T.J.; Carter, J.A.; Ebberts, B.D.; Brown, R.S.; Weiland, M.; Ploskey, G.R.; Harnish, R.A.; Deng, Z.D. The Juvenile Salmon Acoustic Telemetry System: A New Tool. Fisheries 2010, 35, 9–22. [Google Scholar] [CrossRef]
- Sturrock, A.M.; Satterthwaite, W.H.; Yoshida, K.M.; Huber, E.R.; Sturrock, H.J.W.; Nusslé, S.; Carlson, S.M. Eight Decades of Hatchery Salmon Releases in the California Central Valley: Factors Influencing Straying and Resilience. Fisheries 2019, 44, 433–444. [Google Scholar] [CrossRef]
- Yoshiyama, R.M.; Moyle, P.B.; Gerstung, E.R.; Fisher, F.W. Chinook Salmon in the California Central Valley: An Assessment. Fisheries 2000, 25, 6–20. [Google Scholar] [CrossRef]
- Whipple, A.; Grossinger, R.; Rankin, D.; Stanford, B.; Askevold, R. Sacramento-San Joaquin Delta Historical Ecology Investigation: Exploring Pattern and Process; San Franciso Estuary Institute: Richmond, CA, USA, 2012; Available online: https://www.sfei.org/documents/sacramento-san-joaquin-delta-historical-ecology-investigation-exploring-pattern-and-proces (accessed on 15 January 2020).
- Yoshiyama, R.M.; Gerstung, E.R.; Fisher, F.W.; Moyle, P.B. Historical and Present Distribution of Chinook Salmon in the Central Valley Drainage of California. Fish Bull. 2001, 179, 71–176. [Google Scholar]
- Huber, E.R.; Carlson, S.M. Temporal Trends in Hatchery Releases of Fall-Run Chinook Salmon in California’s Central Valley. San Fr. Estuary Watershed Sci. 2015, 13, 1–23. [Google Scholar]
- Michel, C.J.; Ammann, A.J.; Lindley, S.T.; Sandstrom, P.T.; Chapman, E.D.; Thomas, M.J.; Singer, G.P.; Klimley, A.P.; Macfarlane, R.B. Chinook Salmon Outmigration Survival in Wet and Dry Years in California’s Sacramento River. Can. J. Fish. Aquat. Sci. 2015, 72, 1749–1759. [Google Scholar] [CrossRef]
- Perry, R.W.; Pope, A.C.; Romine, J.G.; Brandes, P.L.; Burau, J.R.; Blake, A.R.; Ammann, A.J.; Michel, C.J. Flow-Mediated Effects on Travel Time, Routing, and Survival of Juvenile Chinook Salmon in a Spatially Complex, Tidally Forced River Delta. Can. J. Fish. Aquat. Sci. 2018, 75, 1886–1901. [Google Scholar] [CrossRef]
- Perry, R.W.; Skalski, J.R.; Brandes, P.L.; Sandstrom, P.T.; Klimley, A.P.; Ammann, A.; MacFarlane, B. Estimating Survival and Migration Route Probabilities of Juvenile Chinook Salmon in the Sacramento-San Joaquin River Delta. N. Am. J. Fish. Manag. 2010, 30, 142–156. [Google Scholar] [CrossRef]
- Di Liberto, T. Very Wet 2017 Water Year Ends in California. Science & Information for a Climate-Smart Nation. 2017. Available online: https://www.climate.gov/news-features/featured-images/very-wet-2017-water-year-ends-california (accessed on 15 January 2020).
- Liedtke, T.L.; Beeman, J.W.; Gee, L.P. A Standard Operating Procedure for the Surgical Implantation of Transmitters in Juvenile Salmonids: U.S. Geological Survey Open-File Report 2012–1267. 2012; 50p. Available online: https://pubs.usgs.gov/of/2012/1267/pdf/ofr20121267.pdf (accessed on 15 January 2020).
- Ammann, A.J.; Michel, C.J.; Macfarlane, R.B. The Effects of Surgically Implanted Acoustic Transmitters on Laboratory Growth, Survival and Tag Retention in Hatchery Yearling Chinook Salmon. Environ. Biol. Fishes 2013, 96, 135–143. [Google Scholar] [CrossRef]
- Singer, G.P.; Chapman, E.D.; Ammann, A.J.; Klimley, A.P.; Rypel, A.L.; Fangue, N.A. Historic Drought Influences Outmigration Dynamics of Juvenile Fall and Spring-Run Chinook Salmon. Environ. Biol. Fishes 2020, 103, 543–559. [Google Scholar] [CrossRef]
- Johnston, M.E.; Steel, A.E.; Espe, M.; Sommer, T.; Klimley, A.P.; Sandstrom, P.; Smith, D. Survival of Juvenile Chinook Salmon in the Yolo Bypass and the Lower Sacramento River, California. San Fr. Estuary Watershed Sci. 2018, 16, 1–17. [Google Scholar] [CrossRef]
- Liedtke, T.L.; Hurst, W.R. Yolo Bypass Juvenile Salmon Utilization Study 2016—Summary of Acoustically Tagged Juvenile Salmon and Study Fish Releases, Sacramento River, California. 2017. Available online: https://pubs.usgs.gov/ds/1066/ds1066.pdf (accessed on 15 January 2020).
- Pope, A.C.; Perry, R.W.; Harvey, B.N.; Hance, D.J.; Hansel, H.C. Juvenile Chinook Salmon Survival, Travel Time, and Floodplain Use Relative to Riverine Channels in the Sacramento–San Joaquin River Delta. Trans. Am. Fish. Soc. 2021, 150, 38–55. [Google Scholar] [CrossRef]
- Johnson, R.C.; Nmfs, N.; Windell, S.; Brandes, P.L.; Conrad, J.L.; Ferguson, J.; Goertler, P.A.L.; Harvey, B.N.; Heublein, J.; Israel, J.A.; et al. Science Advancements Key to Increasing Management Value of Life Stage Monitoring Networks for Endangered Sacramento River Winter-Run Chinook Salmon in California. San Fr. Estuary Watershed Sci. 2017, 15, 1–41. [Google Scholar] [CrossRef]
- Goertler, P.; Pien, C. Daily water temperature (°C) in the Yolo Bypass and Sacramento River, 1998–2019 ver 1. Environmental Data Initiative. 2022. Available online: https://portal.edirepository.org/nis/mapbrowse?packageid=edi.1178.1 (accessed on 23 May 2023). [CrossRef]
- Clark, J.; Goertler, P. Inundation. Zenodo. 2022. Available online: https://zenodo.org/records/6450272 (accessed on 23 May 2023). [CrossRef]
- Mantua, N.J.; Hare, S.R. The Pacific Decadal Oscillation. J. Oceanogr. 2002, 58, 35–44. [Google Scholar] [CrossRef]
- Pedersen, E.J.; Miller, D.L.; Simpson, G.L.; Ross, N. Hierarchical Generalized Additive Models in Ecology: An Introduction with Mgcv. PeerJ 2019, 7, e6876. [Google Scholar] [CrossRef] [PubMed]
- Wood, S. Fast stable restricted maximum likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2011, 78, 3–36. [Google Scholar] [CrossRef]
- Goertler, P.; Sommer, T.R.; Satterthwaite, W.H.; Schreier, B.M. Seasonal Floodplain-Tidal Slough Complex Supports Size Variation for Juvenile Chinook Salmon (Oncorhynchus tshawytscha). Ecol. Freshw. Fish 2017, 27, 580–593. [Google Scholar] [CrossRef]
- Sommer, T.R.; Nobriga, M.L.; Harrell, W.C.; Batham, W.; Kimmerer, W.J. Floodplain Rearing of Juvenile Chinook Salmon: Evidence of Enhanced Growth and Survival. Can. J. Fish. Aquat. Sci. 2001, 58, 325–333. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A Protocol for Data Exploration to Avoid Common Statistical Problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Perry, R.W.; Brandes, P.L.; Burau, J.R.; Klimley, A.P.; MacFarlane, B.; Michel, C.; Skalski, J.R. Sensitivity of survival to migration routes used by juvenile Chinook salmon to negotiate the Sacramento−San Joaquin River Delta. Environ. Biol. Fishes 2013, 96, 381–392. [Google Scholar] [CrossRef]
- Skalski, J.R.; Townsend, R.; Lady, J.; Giorgi, A.E.; Stevenson, J.R.; McDonald, R.D. Estimating route-specific passage and survival probabilities at a hydroelectric project from smolt radiotelemetry studies. Can. J. Fish. Aquat. Sci. 2002, 59, 1385–1393. [Google Scholar] [CrossRef]
- Gordo, O. Why Are Bird Migration Dates Shifting? A Review of Weather and Climate Effects on Avian Migratory Phenology. Clim. Res. 2007, 35, 37–58. [Google Scholar] [CrossRef]
- Morrice, K.J.; Baptista, A.M.; Burke, B.J. Environmental and behavioral controls on juvenile Chinook salmon migration pathways in the Columbia River estuary. Ecol. Model. 2020, 427, 109003. [Google Scholar] [CrossRef]
- Scheuerell, M.D.; Zabel, R.W.; Sandford, B.P. Relating Juvenile Migration Timing and Survival to Adulthood in Two Species of Threatened Pacific Salmon (Oncorhynchus spp.). J. Appl. Ecol. 2009, 46, 983–990. [Google Scholar] [CrossRef]
- Achord, S.; Zabel, R.W.; Sandford, B.P. Migration Timing, Growth, and Estimated Parr-to-Smolt Survival Rates of Wild Snake River Spring—Summer Chinook Salmon from the Salmon River Basin, Idaho, to the Lower Snake River. Trans. Am. Fish. Soc. 2007, 136, 142–154. [Google Scholar] [CrossRef]
- Goertler, P.; Jones, K.; Cordell, J.; Schreier, B.; Sommer, T. Effects of Extreme Hydrologic Regimes on Juvenile Chinook Salmon Prey Resources and Diet Composition in a Large River Floodplain. Trans. Am. Fish. Soc. 2017, 147, 287–299. [Google Scholar] [CrossRef]
- Dettinger, M.; Udall, B.; Georgakakos, A. Western Water and Climate Change. Ecol. Appl. 2015, 25, 2069–2093. [Google Scholar] [CrossRef]
- Cloern, J.E.; Knowles, N.; Brown, L.R.; Cayan, D.; Dettinger, M.D.; Morgan, T.L.; Schoellhamer, D.H.; Stacey, M.T.; van der Wegen, M.; Wagner, R.W.; et al. Projected Evolution of California’s San Francisco Bay-Delta-River System in a Century of Climate Change. PLoS ONE 2011, 6, e24465. [Google Scholar] [CrossRef]
- Bradshaw, W.E.; Holzapfel, C.M. Evolutionary Response to Rapid Climate Change. Science 2006, 312, 1477–1478. [Google Scholar] [CrossRef]
- Studds, C.E.; Marra, P.P. Linking Fluctuations in Rainfall to Nonbreeding Season Performance in a Long-Distance Migratory Bird, Setophaga ruticilla. Clim. Res. 2007, 35, 115–122. [Google Scholar] [CrossRef]
- Munsch, S.H.; Greene, C.M.; Johnson, R.C.; Satterthwaite, W.H.; Imaki, H.; Brandes, P.L. Warm, Dry Winters Truncate Timing and Size Distribution of Seaward-Migrating Salmon across a Large, Regulated Watershed. Ecol. Appl. 2019, 29, e01880. [Google Scholar] [CrossRef] [PubMed]
- Munsch, S.H.; Greene, C.M.; Mantua, N.J.; Satterthwaite, W.H. One Hundred–seventy Years of Stressors Erode Salmon Fishery Climate Resilience in California’s Warming Landscape. Glob. Chang. Biol. 2022, 28, 2183–2201. [Google Scholar] [CrossRef]
- Carr-Harris, C.N.; Moore, J.W.; Gottesfeld, A.S.; Gordon, J.A.; Shepert, W.M.; Henry, J.D.J., Jr.; Russell, H.J.; Helin, W.N.B.; Doolan, D.J.; Beacham, T.D. Phenological Diversity of Salmon Smolt Migration Timing within a Large Watershed. Trans. Am. Fish. Soc. 2018, 147, 775–790. [Google Scholar] [CrossRef]
- Thorson, J.T.; Scheuerell, M.D.; Buhle, E.R.; Copeland, T. Spatial Variation Buffers Temporal Fluctuations in Early Juvenile Survival for an Endangered Pacific Salmon. J. Anim. Ecol. 2014, 83, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.M.; Satterthwaite, W.H. Weakened Portfolio Effect in a Collapsed Salmon Population Complex. Can. J. Fish. Aquat. Sci. 2011, 68, 1579–1589. [Google Scholar] [CrossRef]
- Jonsson, B.; Skúlason, S.; Snorrason, S.S.; Sandlund, O.T.; Malmquist, H.J.; Jónasson, P.M.; Cydemo, R.; Lindem, T. Life History Variation of Polymorphic Arctic Charr (Salvelinus alpinus) in Thingvallavatn, Iceland. Can. J. Fish. Aquat. Sci. 1988, 45, 1537–1547. [Google Scholar] [CrossRef]
- Chapman, E.D.; Singer, G.P.; Hayes, S.; Merz, A.J.; Zueg, S.; Hassrick, J.; Null, R.; Kurt, R.; Kindopp, J.; Klimley, A.P. Evaluating the Effects of Release Strategy on Movement and Survival of Fall and Spring Run Chinook Salmon Tagged with JSATS Transmitters in the Sacramento-San Joaquin Delta; Interagency Ecolicigal Program Workshop: Folsom, CA, USA, 2015. [Google Scholar]
- Cordoleani, F.; Notch, J.; McHuron, A.S.; Ammann, A.J.; Michel, C.J. Movement and Survival of Wild Chinook Salmon Smolts from Butte Creek During Their Out-Migration to the Ocean: Comparison of a Dry Year versus a Wet Year. Trans. Am. Fish. Soc. 2018, 147, 171–184. [Google Scholar] [CrossRef]
- Hassrick, J.L.; Ammann, A.J.; Perry, R.W.; John, S.N.; Daniels, M.E. Factors Affecting Spatiotemporal Variation in Survival of Endangered Winter-Run Chinook Salmon Out-Migrating from the Sacramento River. N. Am. J. Fish. Manag. 2022, 42, 375–395. [Google Scholar] [CrossRef]
- Notch, J.J.; McHuron, A.S.; Michel, C.J.; Cordoleani, F.; Johnson, M.; Henderson, M.J.; Ammann, A.J. Outmigration Survival of Wild Chinook Salmon Smolts through the Sacramento River during Historic Drought and High Water Conditions. Environ. Biol. Fishes 2020, 103, 561–576. [Google Scholar] [CrossRef]
- Zeug, S.C.; Null, R.; Brodsky, A.; Johnston, M.; Ammann, A.J. Effect of Release Timing on Apparent Survival of Juvenile Fall Run Chinook Salmon from Coleman National Fish Hatchery. Environ. Biol. Fishes 2020, 103, 411–423. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sacramento Valley Water Year Index | Difference in the Observed Means | Lower Interval | Upper Interval | Adjusted p-Value |
|---|---|---|---|---|
| Dry-Critically Dry | −2.48665 | −5.8542 | 0.8809 | 0.2289 |
| Below Normal-Critically Dry | −10.7529 | −13.2718 | −8.2340 | 0 |
| Wet-Critically Dry | 5.8221 | 3.1579 | 8.4862 | <0.0001 |
| Below Normal-Dry | −8.2663 | −10.9851 | −5.5474 | 0 |
| Wet-Dry | 8.3087 | 5.4548 | 11.1627 | 0 |
| Wet-Below Normal | 16.5750 | 14.8003 | 18.3497 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goertler, P.A.L.; Johnston, M.; Michel, C.J.; Grimes, T.; Singer, G.; Notch, J.; Sommer, T. Use of Telemetry Data to Quantify Life History Diversity in Migrating Juvenile Chinook Salmon (Oncorhynchus tshawytscha). Water 2024, 16, 2529. https://doi.org/10.3390/w16172529
Goertler PAL, Johnston M, Michel CJ, Grimes T, Singer G, Notch J, Sommer T. Use of Telemetry Data to Quantify Life History Diversity in Migrating Juvenile Chinook Salmon (Oncorhynchus tshawytscha). Water. 2024; 16(17):2529. https://doi.org/10.3390/w16172529
Chicago/Turabian StyleGoertler, Pascale Ava Lake, Myfanwy Johnston, Cyril Joseph Michel, Tracy Grimes, Gabriel Singer, Jeremy Notch, and Ted Sommer. 2024. "Use of Telemetry Data to Quantify Life History Diversity in Migrating Juvenile Chinook Salmon (Oncorhynchus tshawytscha)" Water 16, no. 17: 2529. https://doi.org/10.3390/w16172529
APA StyleGoertler, P. A. L., Johnston, M., Michel, C. J., Grimes, T., Singer, G., Notch, J., & Sommer, T. (2024). Use of Telemetry Data to Quantify Life History Diversity in Migrating Juvenile Chinook Salmon (Oncorhynchus tshawytscha). Water, 16(17), 2529. https://doi.org/10.3390/w16172529