


Study on Ferritin Gene Expression to Evaluate the Health of White Leg Shrimp (Litopenaeus vannamei) Postlarvae Due to Changes in Water Temperature, Salinity, and pH


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Experimental Animals
2.2. Water Temperature, Salinity, and pH
2.3. Total RNA Extraction
2.4. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
2.5. Statistical Analysis
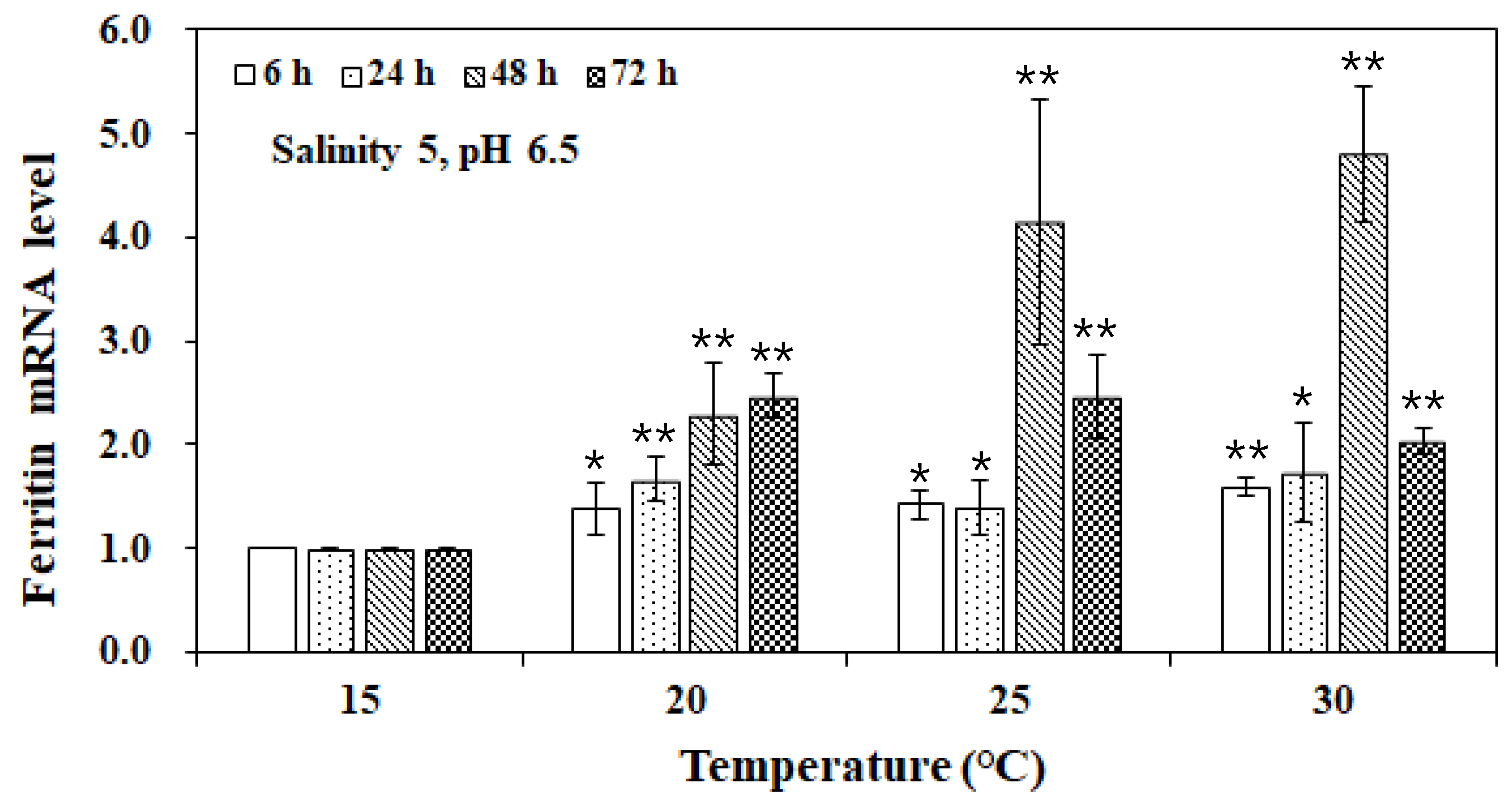
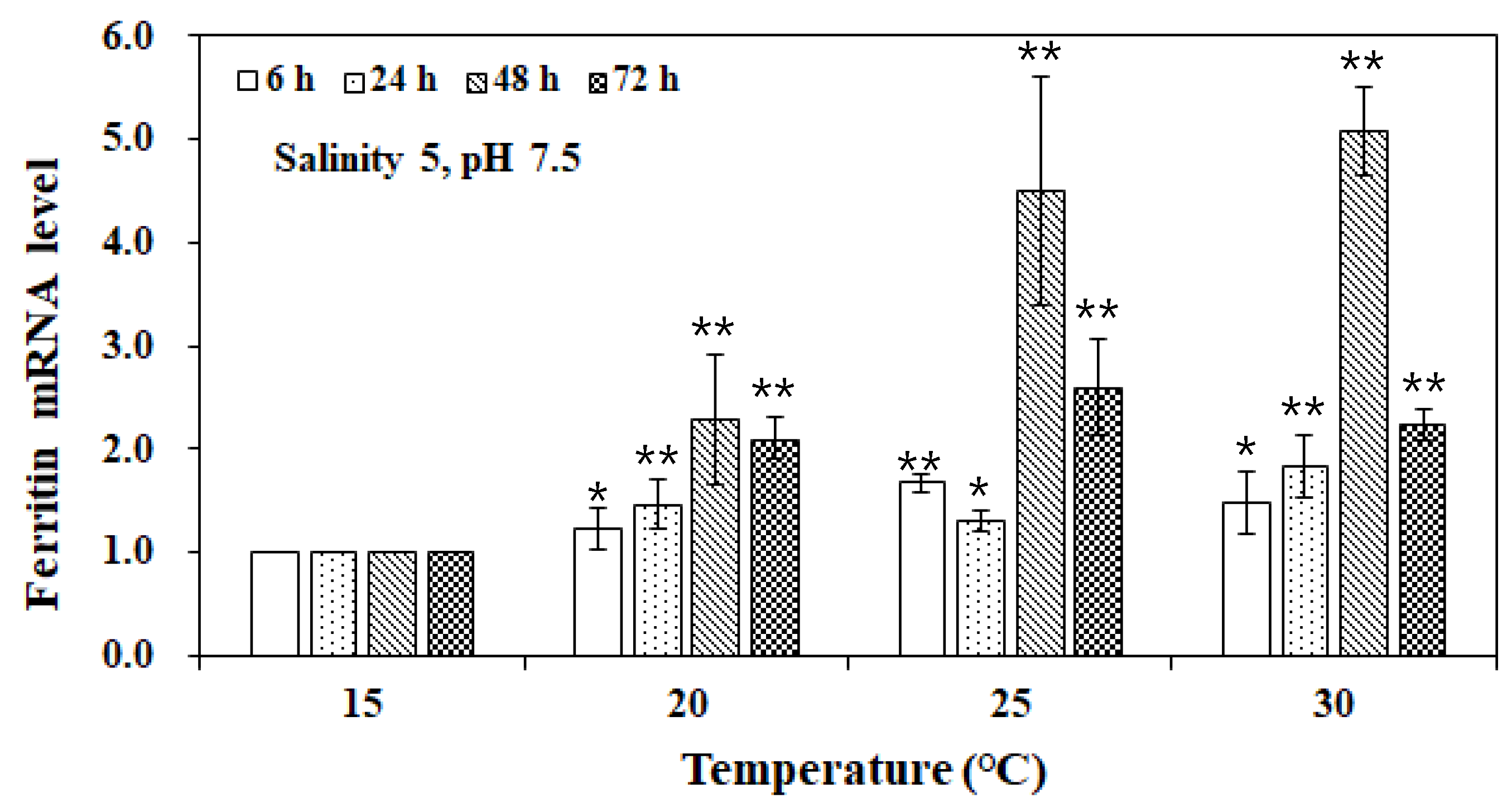
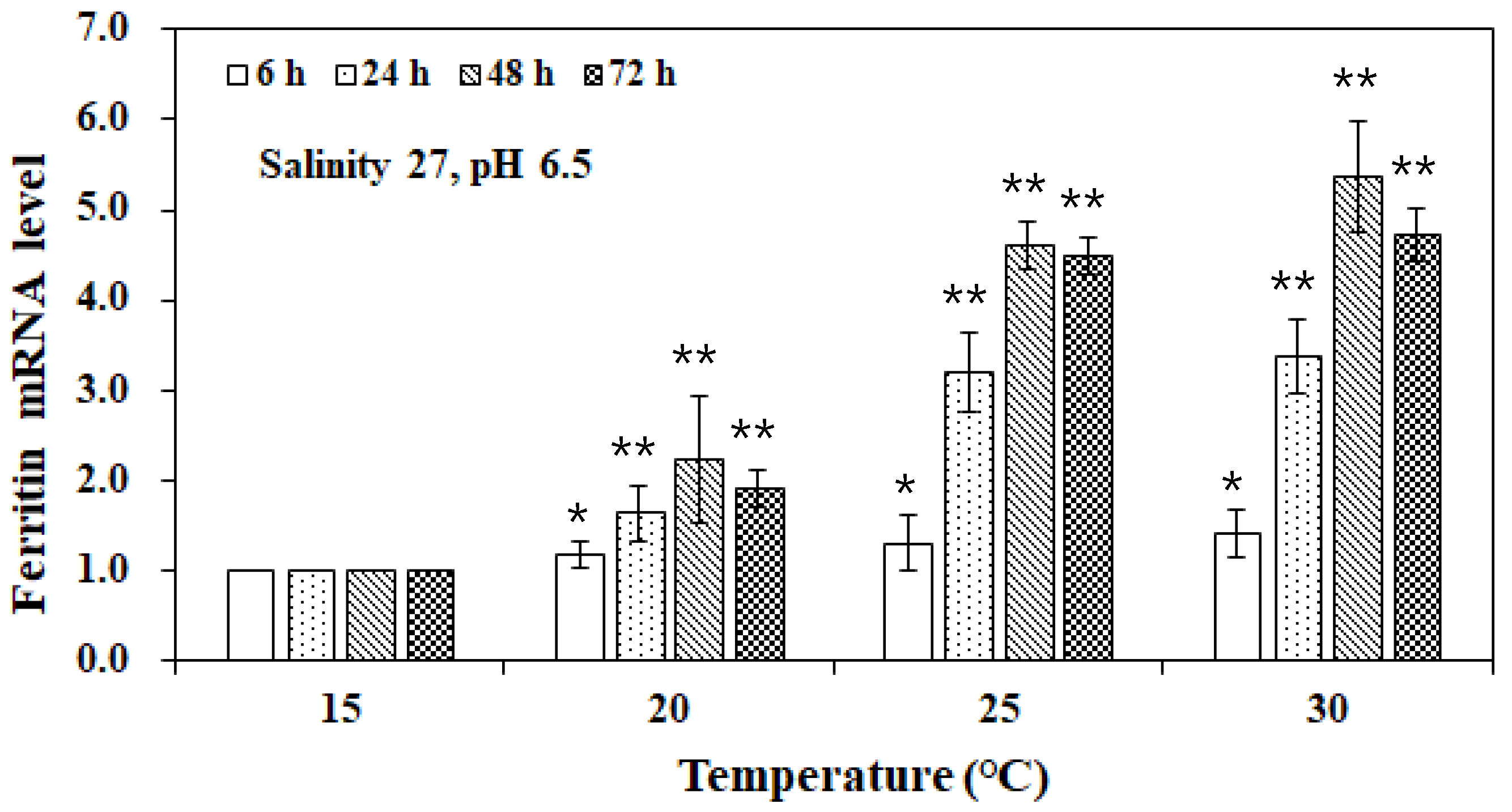
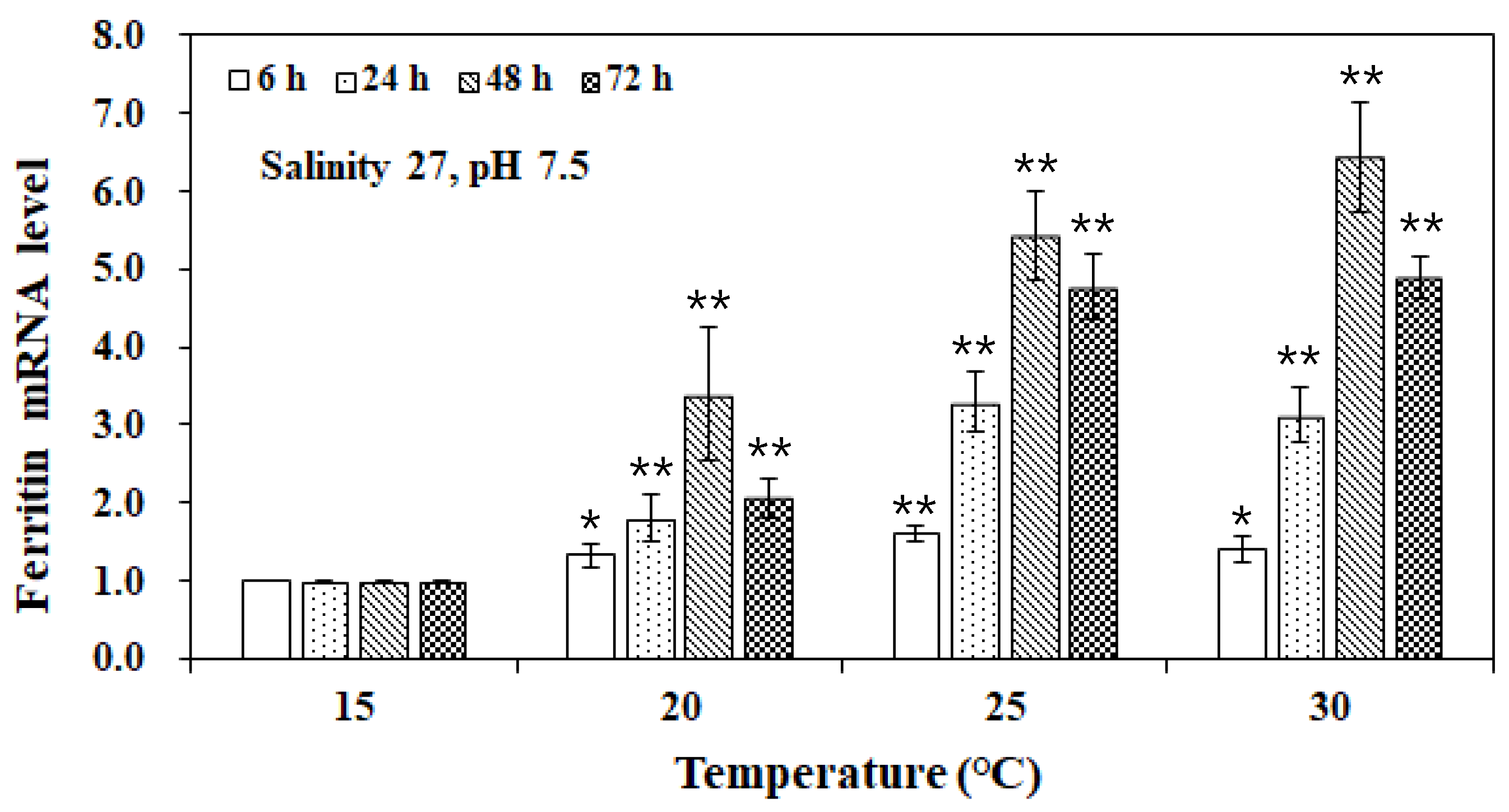
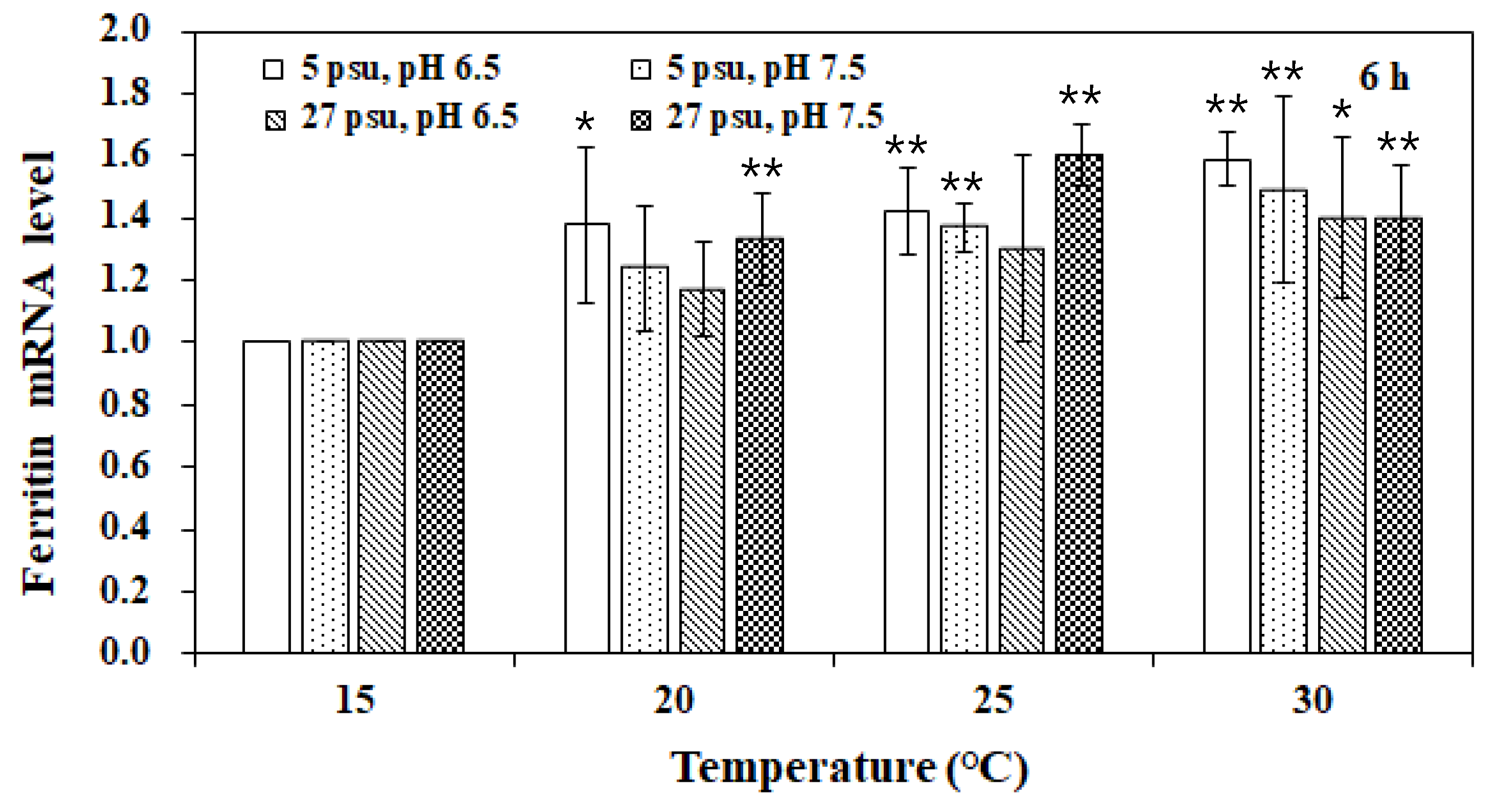
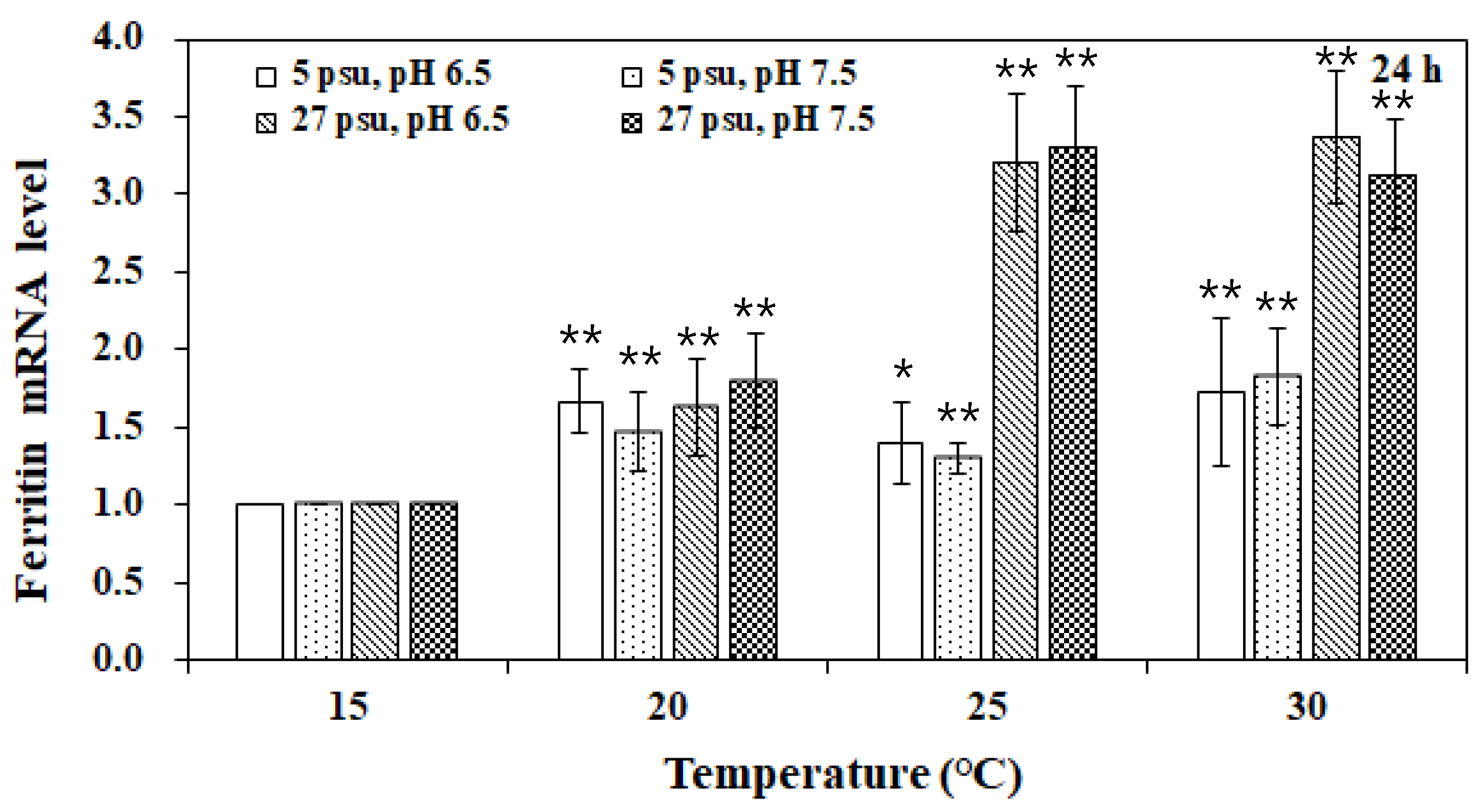
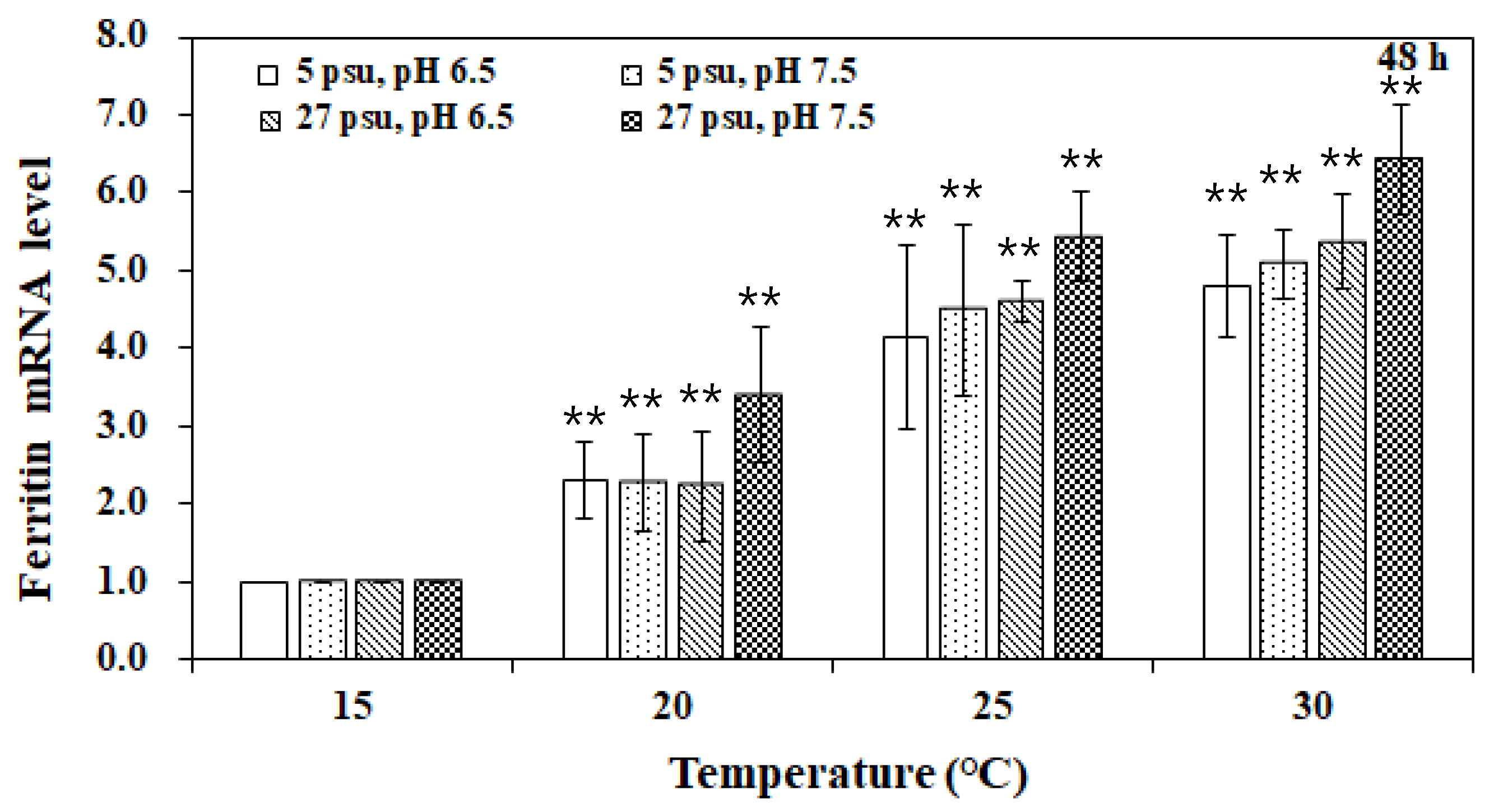
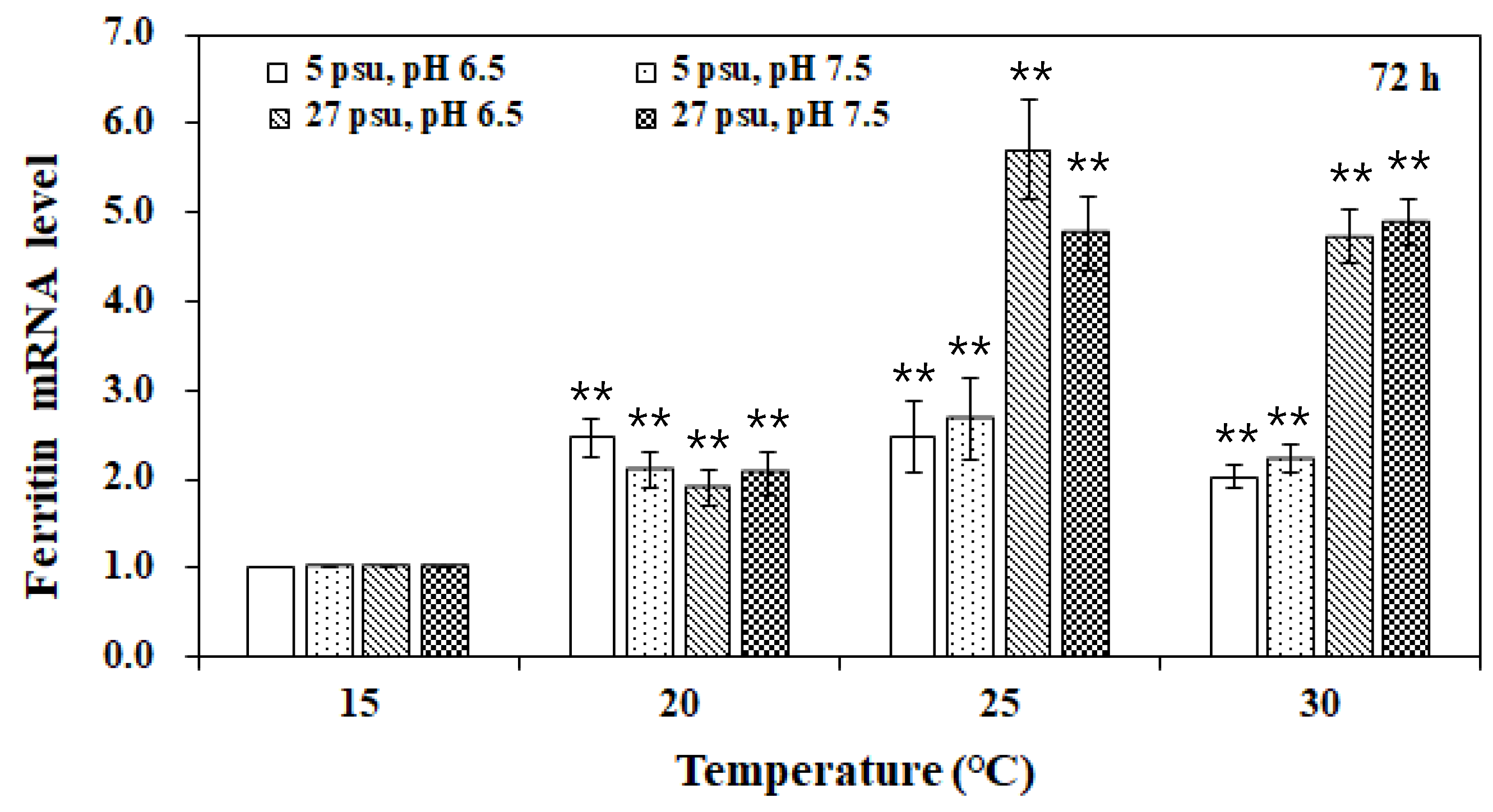
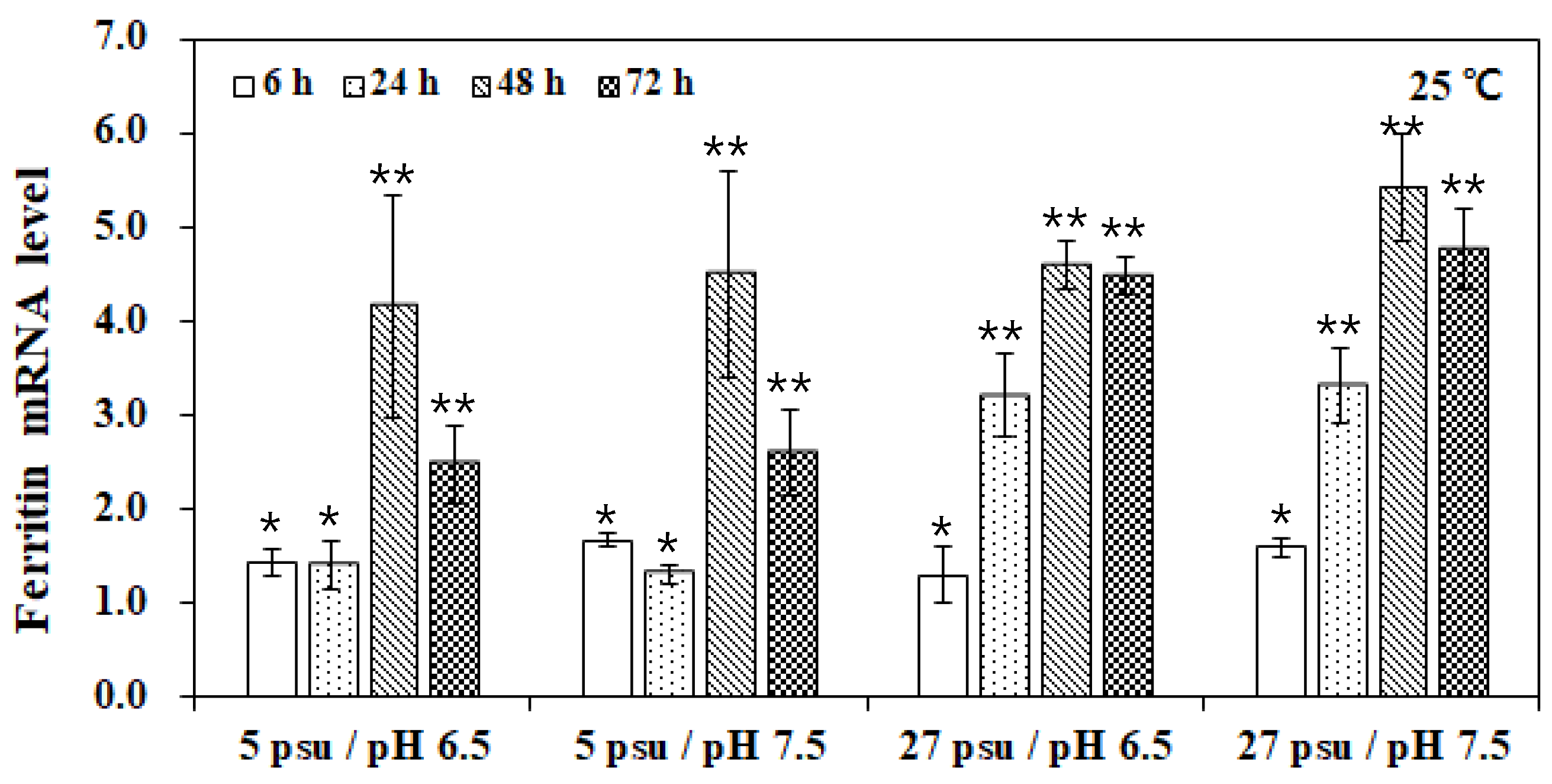
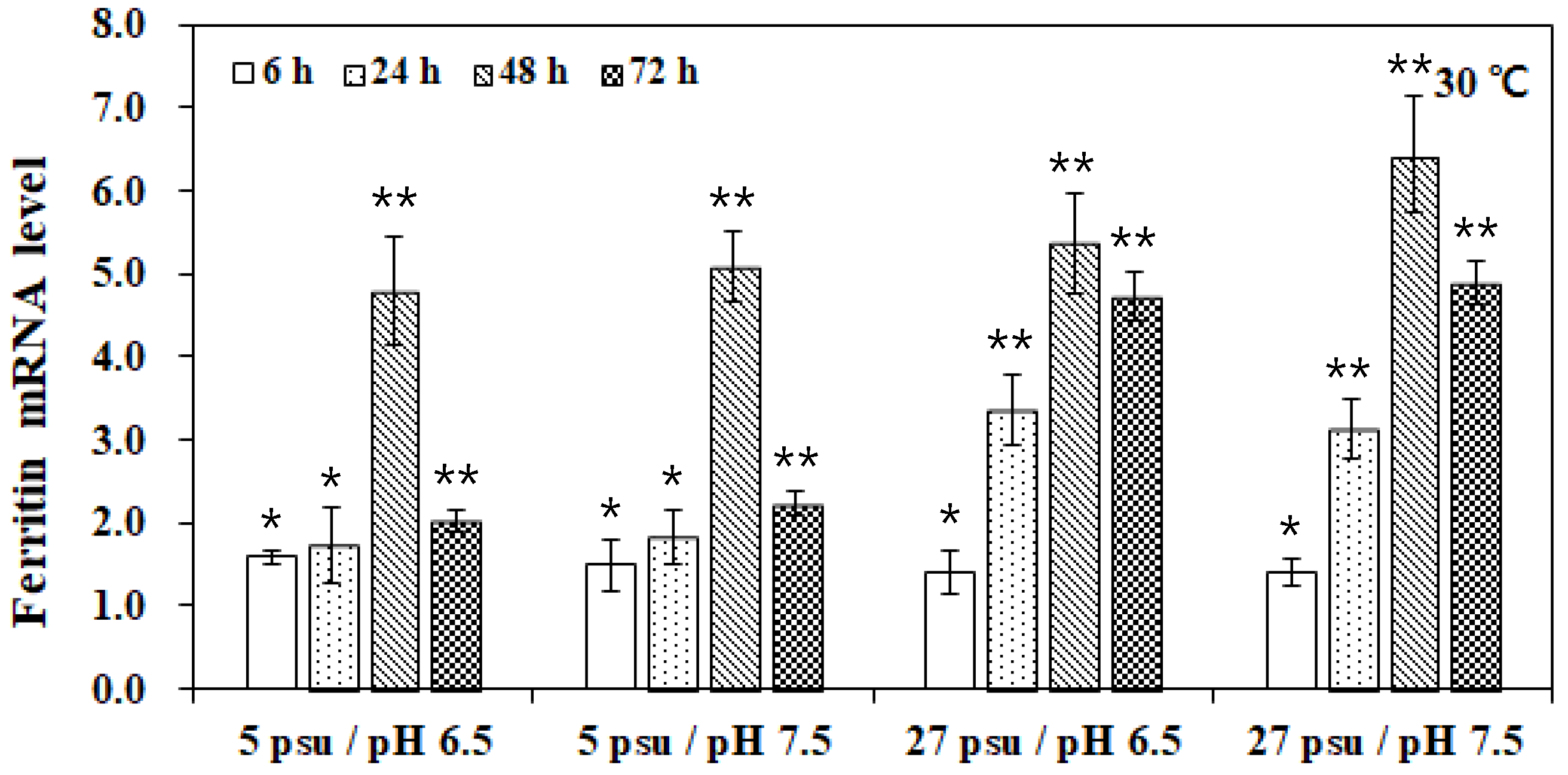
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapple, J.P.; Smerdon, G.R.; Berry, R.J.; Hawkins, A.J.S. Seasonal changes in stress-70 protein levels reflect thermal tolerance in the marine bivalve Mytilus edulis L. J. Exp. Mar. Biol. Ecol. 1998, 229, 53–68. [Google Scholar] [CrossRef]
- Norris, A.L.; Houser, D.S.; Crocker, D.E. Environment and activity affect skin temperature in breeding adult male elephant seals (Mirounga angustirostris). J. Exp. Biol. 2010, 213, 4205–4212. [Google Scholar] [CrossRef]
- Hvas, M.; Folkedal, O.; Imsland, A.; Oppedal, F. Metabolic rates; swimming capabilities; thermal niche and stress response of the lumpfish; Cyclopterus lumpus. Biol. Open. 2018, 7, bio036079. [Google Scholar] [CrossRef]
- Pavlosky, K.K.; Yamaguchi, Y.; Lerner, D.T.; Seale, A.P. The effects of transfer from steady-state to tidally-changing salinities on plasma and branchial osmoregulatory variables in adult Mozambique tilapia. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 227, 134–145. [Google Scholar] [CrossRef]
- Sampaio, E.; Lopes, A.R.; Francisco, S.; Paula, J.R.; Pimentel, M.; Maulvault, A.L.; Repolho, T.; Grilo, T.F.; Pousão-Ferreira, P.; Marques, A.; et al. Ocean acidification dampens physiological stress response to warming and contamination in a commercially-important fish (Argyrosomus regius). Sci. Total Environ. 2018, 618, 388–398. [Google Scholar] [CrossRef]
- Newell, R.C.; Kofoed, L.H. Adjustment of the components of energy balance in the gastropod Crepidula fornicate in response to thermal acclimation. Mar. Biol. 1977, 44, 275–286. [Google Scholar] [CrossRef]
- Logue, J.; Tiku, P.; Cossins, A.R. Heat injury and resistance adaptation in fish. J. Therm. Biol. 1995, 20, 191–197. [Google Scholar] [CrossRef]
- Loomis, S.H.; Ansell, A.D.; Gibson, R.N.; Barnes, M. Freezing tolerance of marine invertebrates. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 337–350. [Google Scholar]
- Somero, G.N. Adaptation of enzymes to temperature: Searching for basic “strategies”. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 321–333. [Google Scholar] [CrossRef]
- Caissie, D. The thermal regime of rivers: A review. Freshwater Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Crossin, G.T.; Hinch, S.G.; Cooke, S.J.; Welsh, D.W.; Patterson, D.A.; Jones, S.R.M.; Lotto, A.G.; Leggatt, R.A.; Mathes, M.T.; Shrimpton, J.M.; et al. Exposure to high temperature influences the behaviour; physiology; and survival of sockeye salmon during spawning migration. Can. J. Zool. 2008, 86, 127–140. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Hu, M.; Lu, W. Physiological energetics of the thick shell mussel Mytilus coruscus exposed to seawater acidification and thermal stress. Sci. Total Environ. 2015, 514, 261–272. [Google Scholar] [CrossRef]
- Quinn, N.L.; McGowan, C.R.; Cooper, G.A.; Koop, B.F.; Davidson, W.S. Identification of genes associated with heat tolerance in Arctic charr exposed to acute thermal stress. Physiol. Genom. 2011, 43, 685–696. [Google Scholar] [CrossRef]
- Burt, J.M.; Hinch, S.G.; Patterson, D.A. Parental identity influences progeny responses to incubation thermal stress in sockeye salmon Onchorhynchus nerka. J. Fish Biol. 2012, 80, 444–462. [Google Scholar] [CrossRef]
- Hevrøy, E.M.; Waagbo, R.; Torstensen, B.E.; Takle, H.; Stubhaug, I.; Jorgensen, S.M.; Torgersen, T.; Tvenning, L.; Susort, S.; Breck, O.; et al. Ghrelin is involved in voluntary anorexia in Atlantic salmon raised at elevated sea temperatures. Gen. Comp. Endocrinol. 2012, 175, 118–134. [Google Scholar] [CrossRef]
- Morita, K.; Nakashima, A. Temperature seasonality during fry out-migration influences the survival of hatchery-reared chum salmon Oncorhynchus keta. J. Fish Biol. 2015, 87, 1111–1117. [Google Scholar] [CrossRef]
- Tsuji, Y.; Ayaki, H.; Whitman, S.P.; Morrow, C.S.; Torti, S.V.; Torti, F.M. Coordinate transcriptional and translational regulation of ferritin in response to oxidative stress. Mol. Cell. Biol. 2000, 20, 5818–5827. [Google Scholar] [CrossRef]
- Dame, R.F. Ecology of Marine Bivalves: An Ecosystem Approach; CRC Marine Science Series; CRC Press: Boca Raton, FL, USA, 1996; p. 254. [Google Scholar]
- Hammond, L.S. Experimental studies of salinity tolerance; burrowing behavior and pedicle regeneration in Lingula anatina (Brachiopoda; Inarticulata). J. Paleontol. 1983, 57, 1311–1316. [Google Scholar]
- Beckmann, R.P.; Mizzen, L.E.; Welch, W.J. Interaction of HSP70 with newly synthesized proteins: Implications for protein folding and assembly. Science 1990, 248, 850–854. [Google Scholar] [CrossRef]
- Weirich, C.R.; Tomasso, J.R. Confinement and transport induced stress on red drum juveniles: Effect of salinity. Prog. Fish Cult. 1991, 53, 146–149. [Google Scholar] [CrossRef]
- Boeuf, G.; Payan, P. How should salinity influence fish growth? Comp. Biochem. Physiol. Part C 2001, 130, 411–423. [Google Scholar]
- Marshall, W.S. Na+, Cl−, Ca2+ and Zn2+ transport by fish gills: Retrospective review and prospective synthesis. J. Exp. Zool. 2002, 293, 264–283. [Google Scholar] [CrossRef] [PubMed]
- Matthews, Y.T.G.; Fairweather, P.G. Effect of lowered salinity on the survival, condition and reburial of Soletellina alba (Lamarch, 1818) (Bivalvia: Psammobiidae). Aust. Ecol. 2004, 29, 250–257. [Google Scholar] [CrossRef]
- Konijn, A.M.; Glickstein, H.; Vaisman, B.; Meyron-Holtz, E.G.; Slotki, I.N.; Cabantchik, Z.I. The cellular labile iron pool and intracellular ferritin in K562 cells. Blood 1999, 94, 2128–2134. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.Z.; Theil, E.C. Identification and characterization of the iron regulatory element in the ferritin gene of a plant (soybean). J. Biol. Chem. 2000, 275, 17488–17493. [Google Scholar] [CrossRef] [PubMed]
- Kakhlon, O.; Gruenbaum, Y.; Cabantchik, Z.I. Repression of the heavy ferritin chain increases the labile iron pool of human K562 cells. Biochem. J. 2001, 356, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Briat, J.F.; Ravet, K.; Arnaud, N.; Duc, C.; Boucherez, J.; Touraine, B.; Cellier, F.; Gaymard, F. New insights into ferritin synthesis and function highlight a link between iron homeostasis and oxidative stress in plants. Ann. Bot. 2009, 105, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.M.; Arosio, P. The ferritins: Molecular properties, iron storage function and cellular regulation. Biochim. Biophys. Acta (BBA)-Bioenerg. 1996, 1275, 161–203. [Google Scholar] [CrossRef]
- Wu, Y.; Ming, T.; Huo, C.; Qiu, X.; Su, C.; Lu, C.; Zhou, J.; Li, Y.; Su, X. Crystallographic characterization of a marine invertebrate ferritin from the sea cucumber Apostichopus japonicas. FEBS Open Bio 2022, 12, 664–674. [Google Scholar] [CrossRef]
- Ruan, Y.H.; Kuo, C.M.; Lo, C.F.; Lee, M.H.; Lian, J.L.; Hsieh, S.L. Ferritin administration effectively enhances immunity, physiological respnses, and survival of pacific white shrimp (Litopenaeus vannamei) challenged with white spot syndrome virus. Fish Shellfish Immunol. 2010, 28, 542–548. [Google Scholar] [CrossRef]
- Galay, R.L.; Umemiya-Shirafuji, R.; Bacolod, E.T.; Maeda, H.; Kusakisako, K.; Koyama, J.; Tsuji, N.; Mochizuki, M.; Fusisaki, K.; Tanaka, T. Two kinds of ferritin protect ixodid ticks from iron overload and consequent oxidative stress. PLoS ONE 2014, 9, e90661. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Wu, X.; Wu, W.; Dai, C.; Yuan, J. Ferritin protect shrimp Litopenaeus vannamei from WSSV infection by inhibiting virus replication. Fish Shellfish Immunol. 2015, 42, 138–143. [Google Scholar] [CrossRef] [PubMed]
- McGraw, W.J.; Davis, D.A.; Teichert-Coddington, D.; Rouse, D.B. Acclimation of Litopenaeus vannamei postlarvae to low salinity: Influence of Age, Salinity endpoint, and rate of salinity reduction. J. World Aquac. Soc. 2002, 33, 78–84. [Google Scholar] [CrossRef]
- McGraw, W.J.; Scarps, J. Minimum Environmental potassium for survival of Pacific white shrimp Litopenaeus vannamei (Boone) in freshwater. J. Shellfish Res. 2003, 22, 263–267. [Google Scholar]
- McGraw, W.J.; Scarps, J. Mortality of freshwater-acclimated Litopenaeus vannamei associated with acclimation rate, habituation period, and ionic challenge. Aquaculture 2004, 236, 285–296. [Google Scholar] [CrossRef]
- Roy, L.A.; Davis, D.A.; Saoud, I.P.; Henry, R.P. Effects of varying levels of aqueous potassium and magnesium on survival, growth and respiration of the Pacific white shrimp, Litopenaeus vannamei, reared in low salinity waters. Aquaculture 2007, 262, 461–469. [Google Scholar] [CrossRef]
- Castillo-Soriano, F.A.; Ibarra-Junquera, V.A.; Olivos-Ortiz, F.J.; Barragán-Vázquez, F.J.; Meyer-Willerer, A.O. Influence of water supply chemistry on white shrimp (Litopenaeus vannamei) culture in low-salinity and zero-water exchange ponds. Panam. J. Aquat. Sci. 2010, 5, 376–386. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wickins, J.F.; Lee, D.O.C. Crustacean Farming: Ranching and Culture, 2nd ed.; Blackwell Science: Ames, IA, USA, 2008. [Google Scholar]
- Wyban, J.; Walsh, W.A.; Godin, D.M. Temperature effects on growth, feeding rate and feed 858 conversion of the Pacific white shrimp. Aquaculture 1995, 138, 267–279. [Google Scholar] [CrossRef]
- Brett, J.R. Environmental factors and growth. In Fish Physiology; Hoar, W.S., Randall, D.J., Brett, J.R., Eds.; Academic Press: London, UK, 1979; Volume 8, pp. 599–667. [Google Scholar]
- Lester, L.; Pante, M.J. Penaeid temperature and salinity responses. Dev. Aquacult. Fish. Sci. 1992, 23, 515–534. [Google Scholar]
- FAO. Health Management and Biosecurity Maintenance in White Shrimp (Penaeus vannamei) Hatcheries in Latin America; FAO Fisheries Technical Paper. No. 450; FAO: Rome, Italy, 2003; 58p. [Google Scholar]
- Le Moullac, G.; Haffner, P. Environmental factors affecting immune responses in Crustacea. Aquaculture 2000, 191, 121–131. [Google Scholar] [CrossRef]
- Chen, J.C.; Chin, T.S. Effect of Ammonia at different pH levels on Penaeus monodon postlarvae. Asia Fish. Sci. 1989, 2, 233–238. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.-W.; Lee, J.-W.; Kang, S.-W.; Kang, H.-S. Study on Ferritin Gene Expression to Evaluate the Health of White Leg Shrimp (Litopenaeus vannamei) Postlarvae Due to Changes in Water Temperature, Salinity, and pH. Water 2024, 16, 1477. https://doi.org/10.3390/w16111477
Kim C-W, Lee J-W, Kang S-W, Kang H-S. Study on Ferritin Gene Expression to Evaluate the Health of White Leg Shrimp (Litopenaeus vannamei) Postlarvae Due to Changes in Water Temperature, Salinity, and pH. Water. 2024; 16(11):1477. https://doi.org/10.3390/w16111477
Chicago/Turabian StyleKim, Chul-Won, Ju-Wook Lee, Seung-Won Kang, and Han-Seung Kang. 2024. "Study on Ferritin Gene Expression to Evaluate the Health of White Leg Shrimp (Litopenaeus vannamei) Postlarvae Due to Changes in Water Temperature, Salinity, and pH" Water 16, no. 11: 1477. https://doi.org/10.3390/w16111477
APA StyleKim, C.-W., Lee, J.-W., Kang, S.-W., & Kang, H.-S. (2024). Study on Ferritin Gene Expression to Evaluate the Health of White Leg Shrimp (Litopenaeus vannamei) Postlarvae Due to Changes in Water Temperature, Salinity, and pH. Water, 16(11), 1477. https://doi.org/10.3390/w16111477