
Freshwater Salinization Impacts the Interspecific Competition between Microcystis and Scenedesmus
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae and Salinity Adaptation
2.2. Experimental Protocol
2.3. Data Analyses
2.4. Chlorophyll Fluorescence Measurements
3. Results
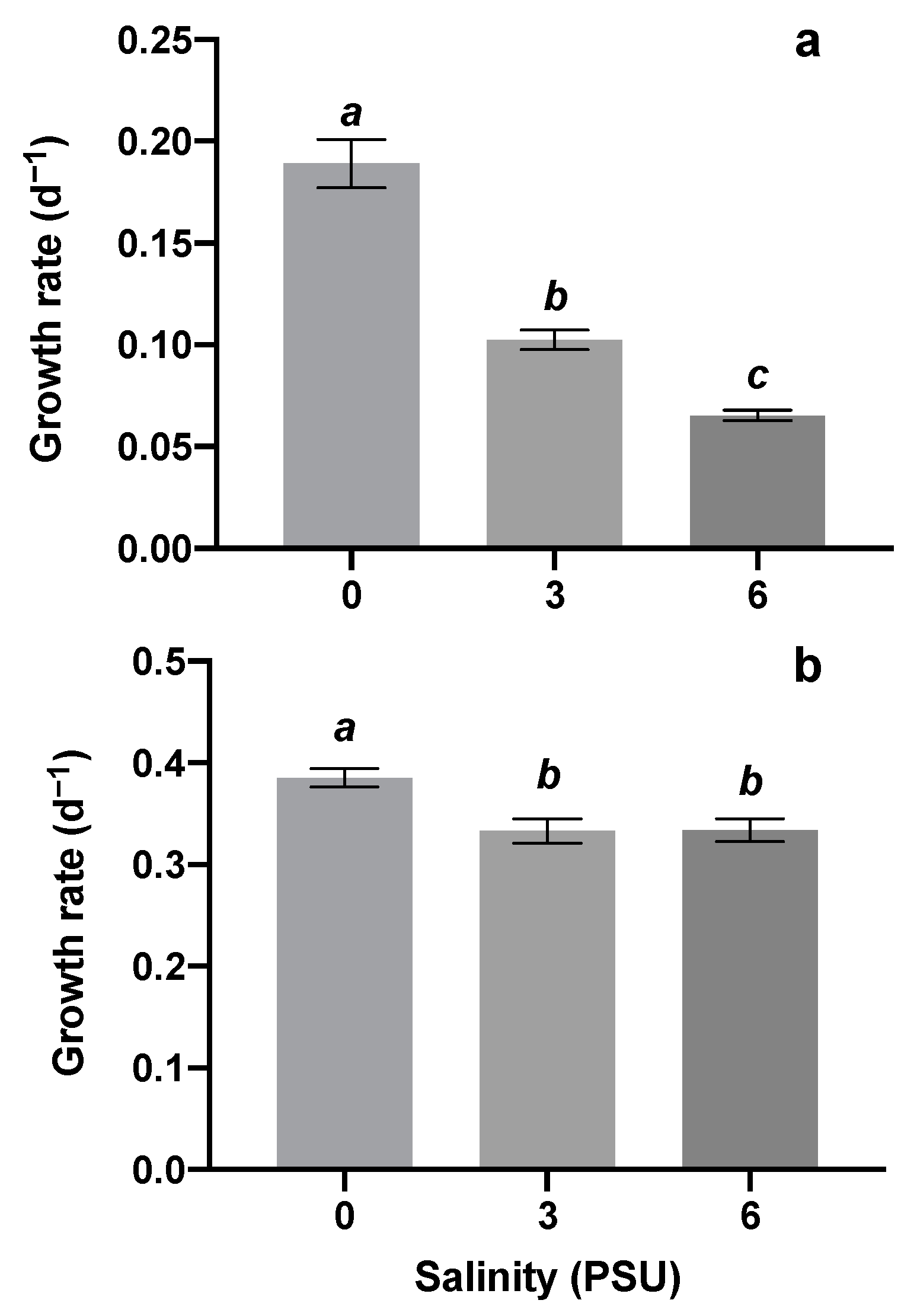
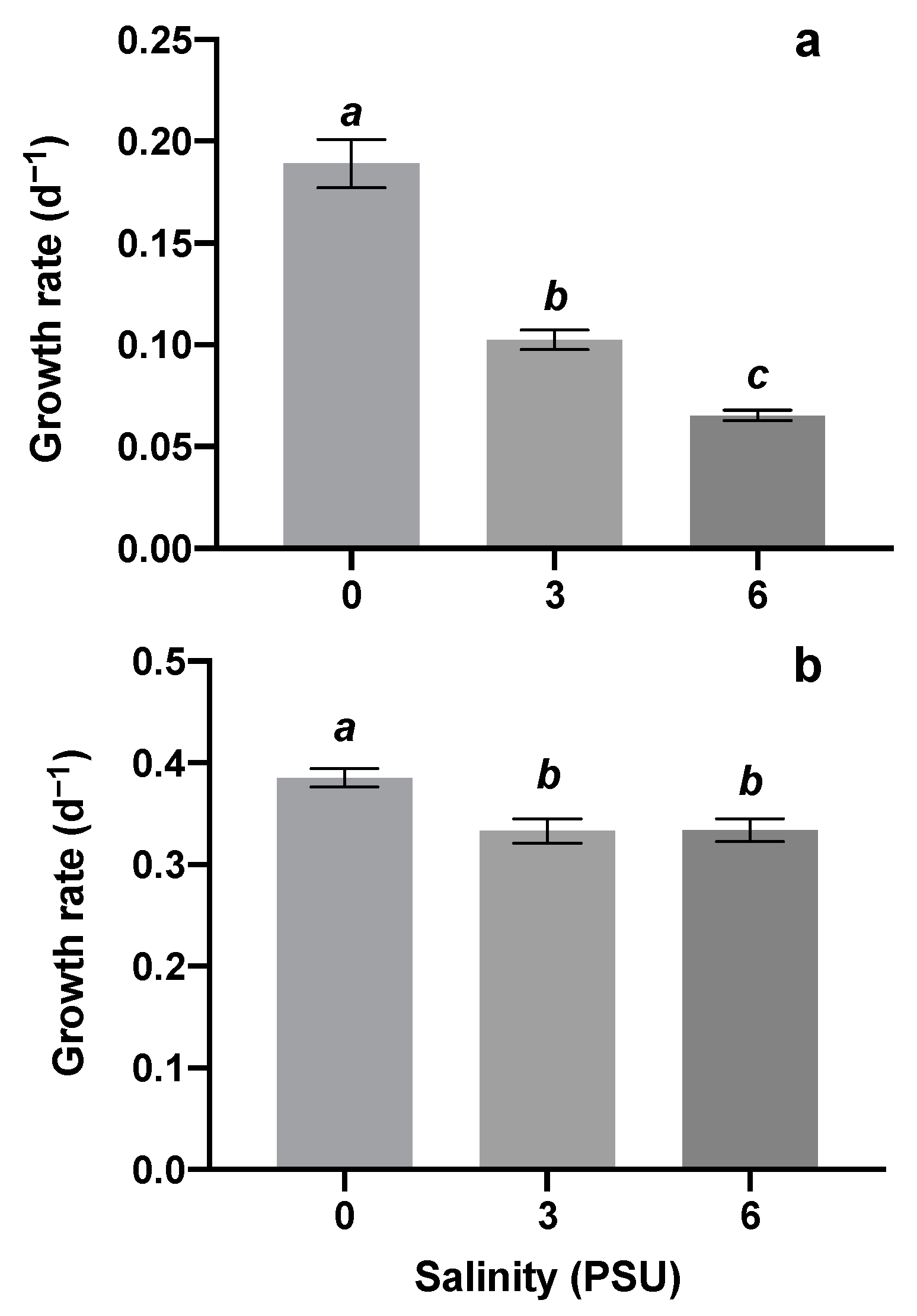
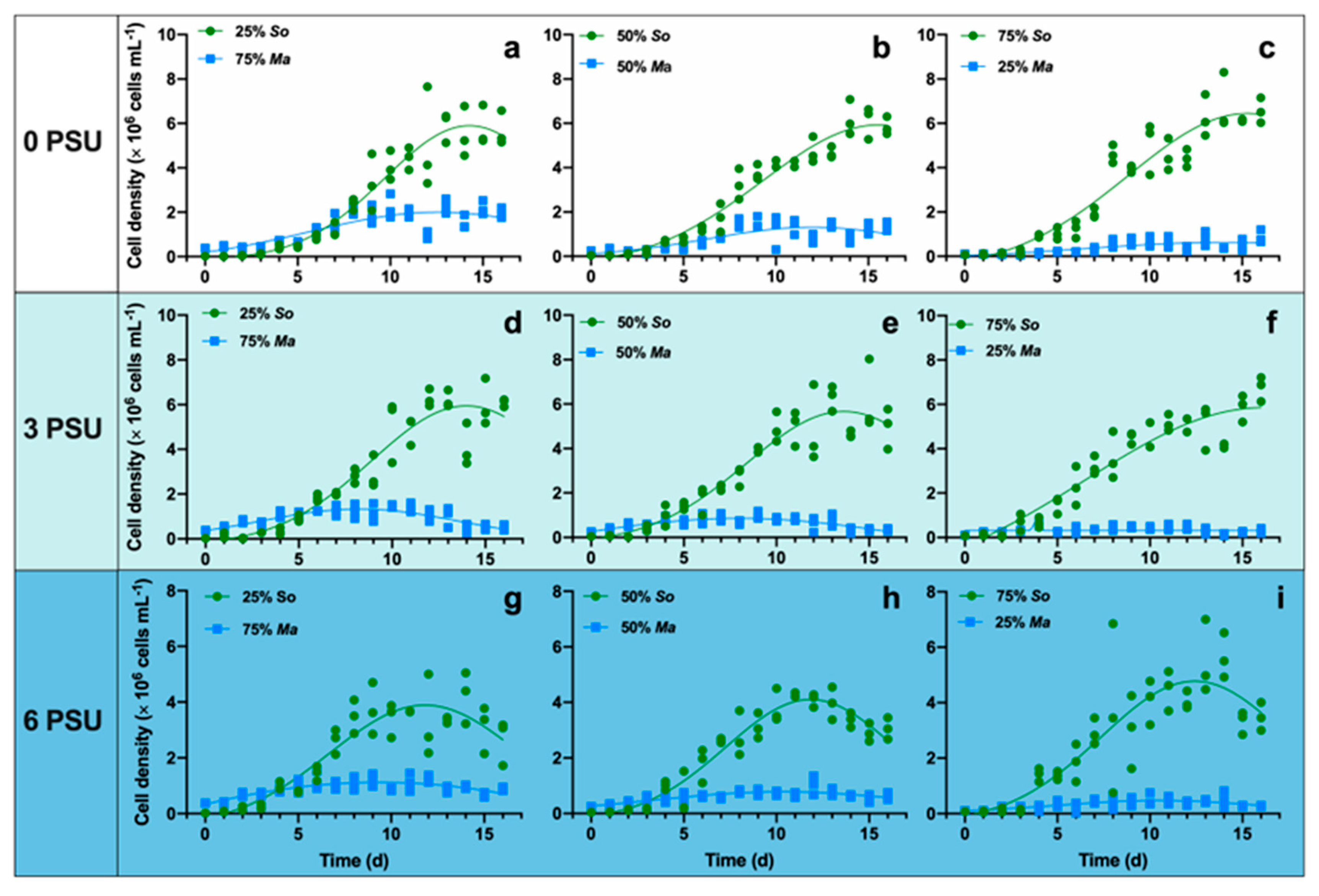
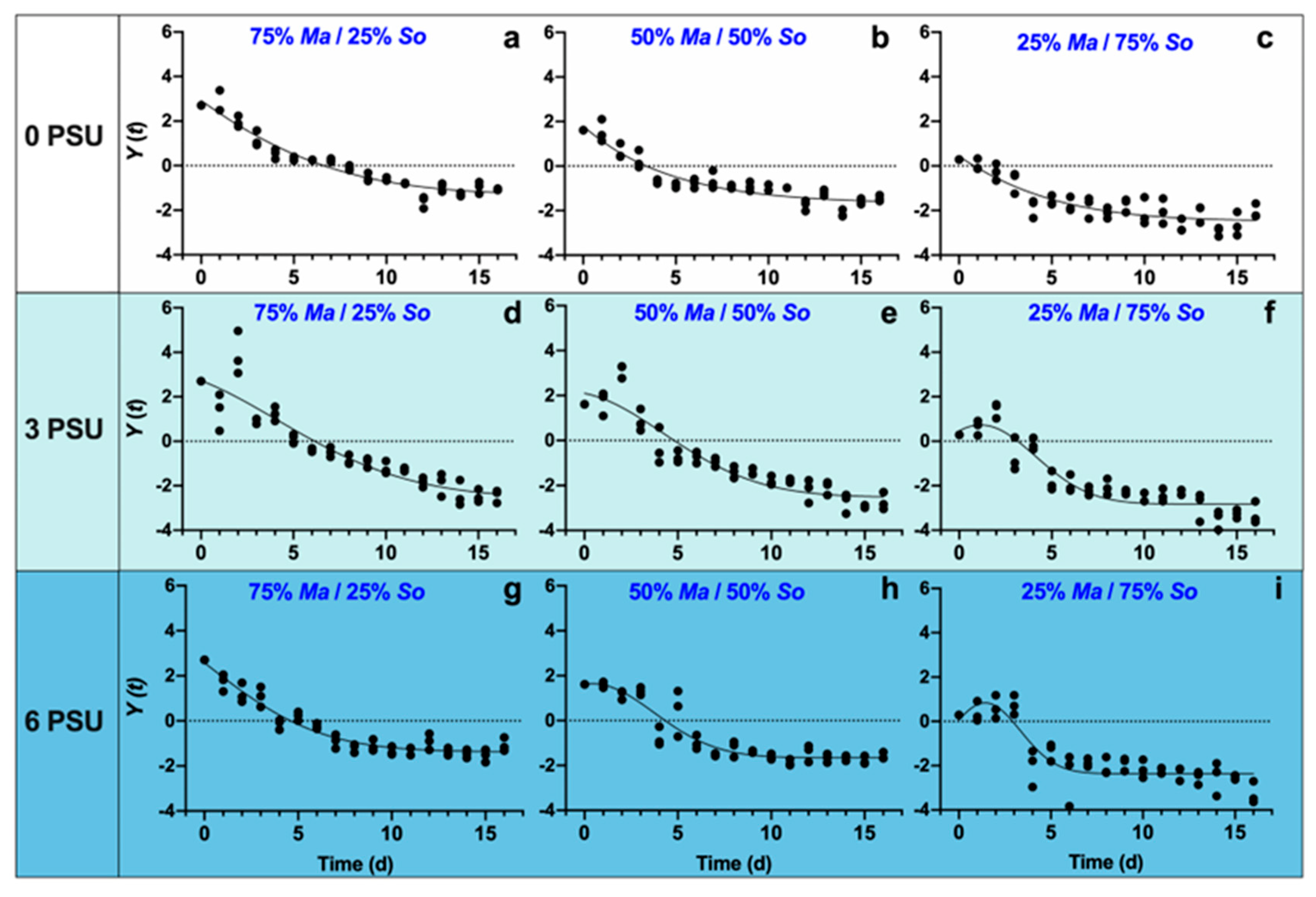
3.1. Response of Algae Growth and Competition to Salinity Stress
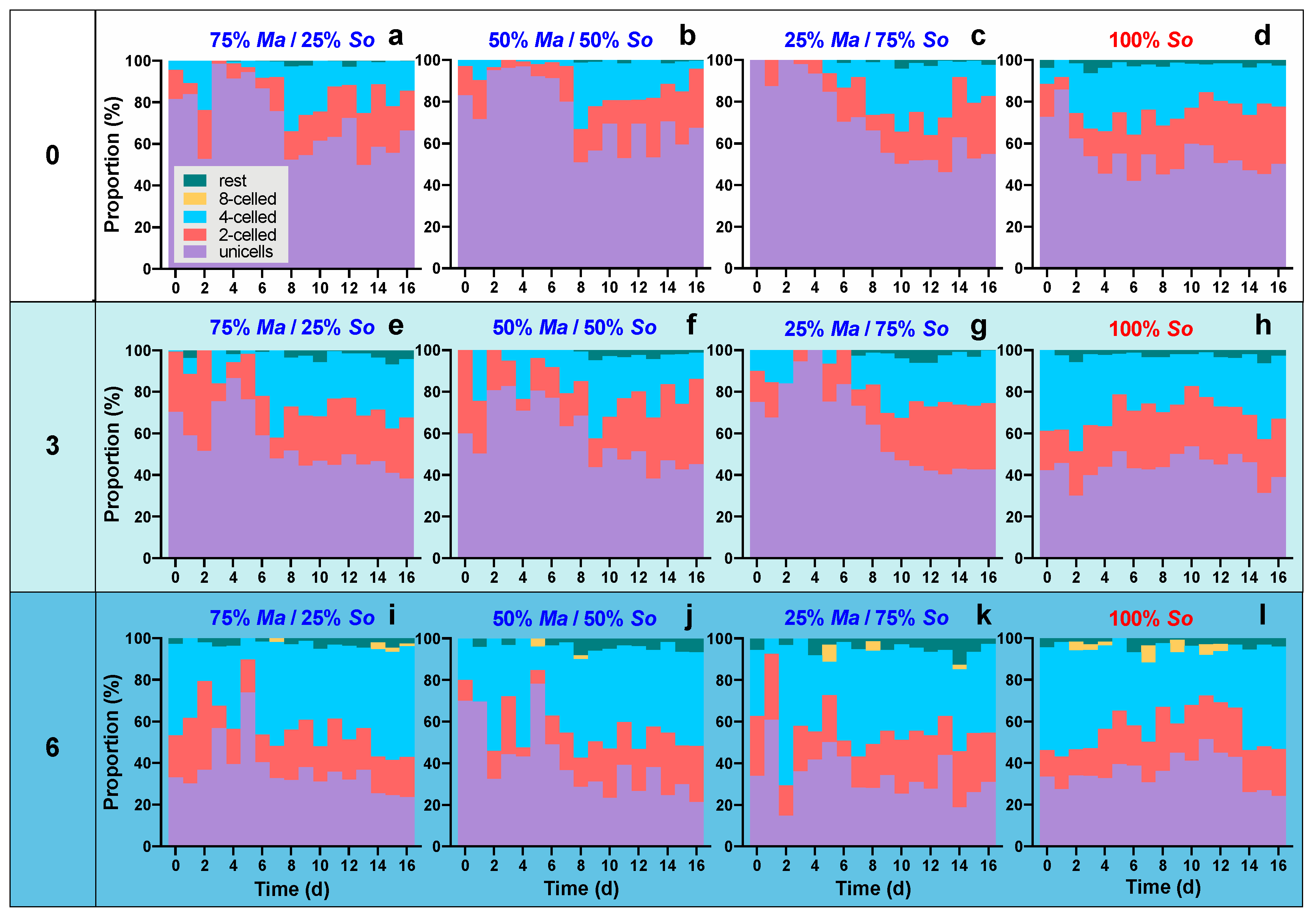
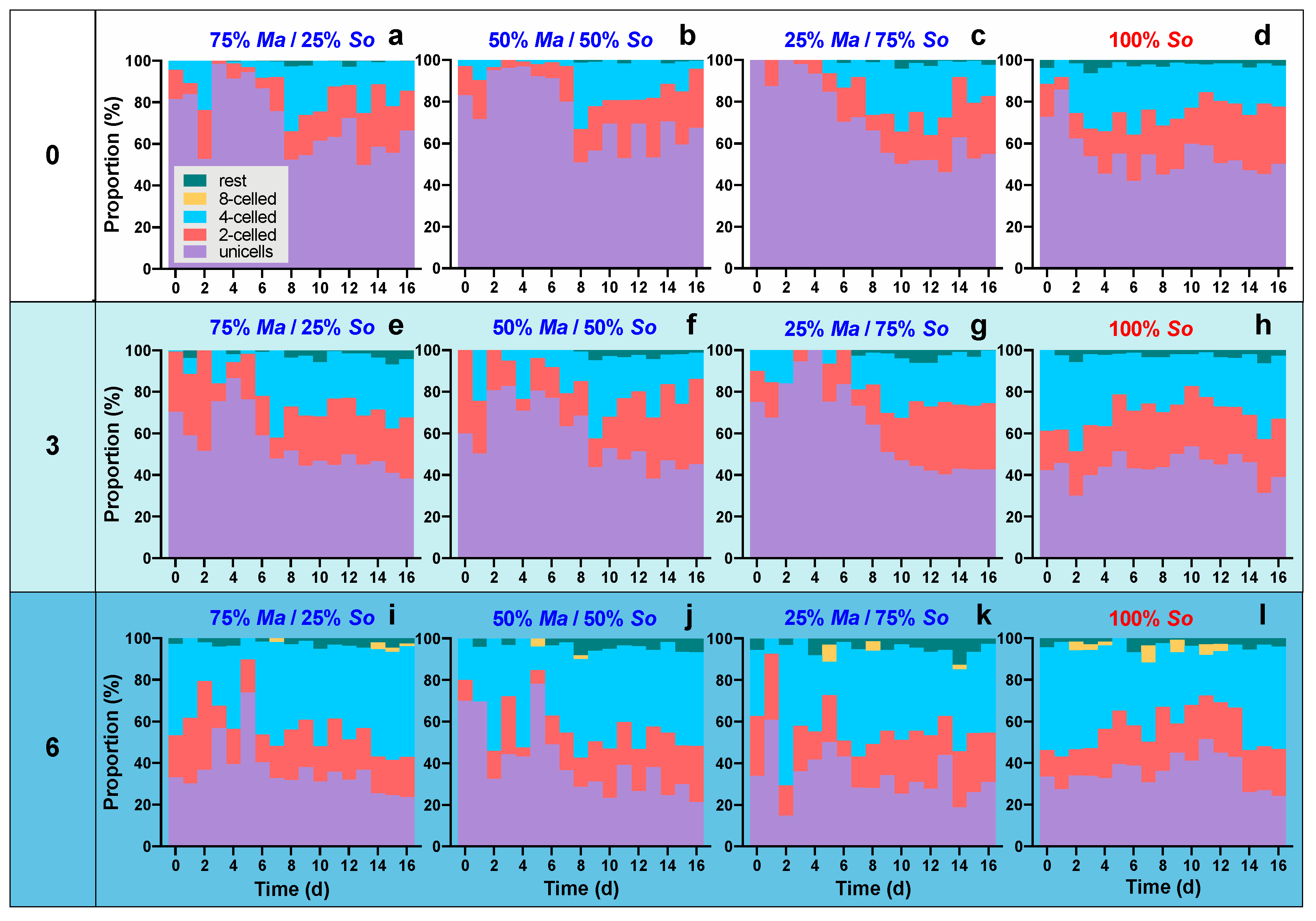
3.2. Morphological Variations of S. obliquus
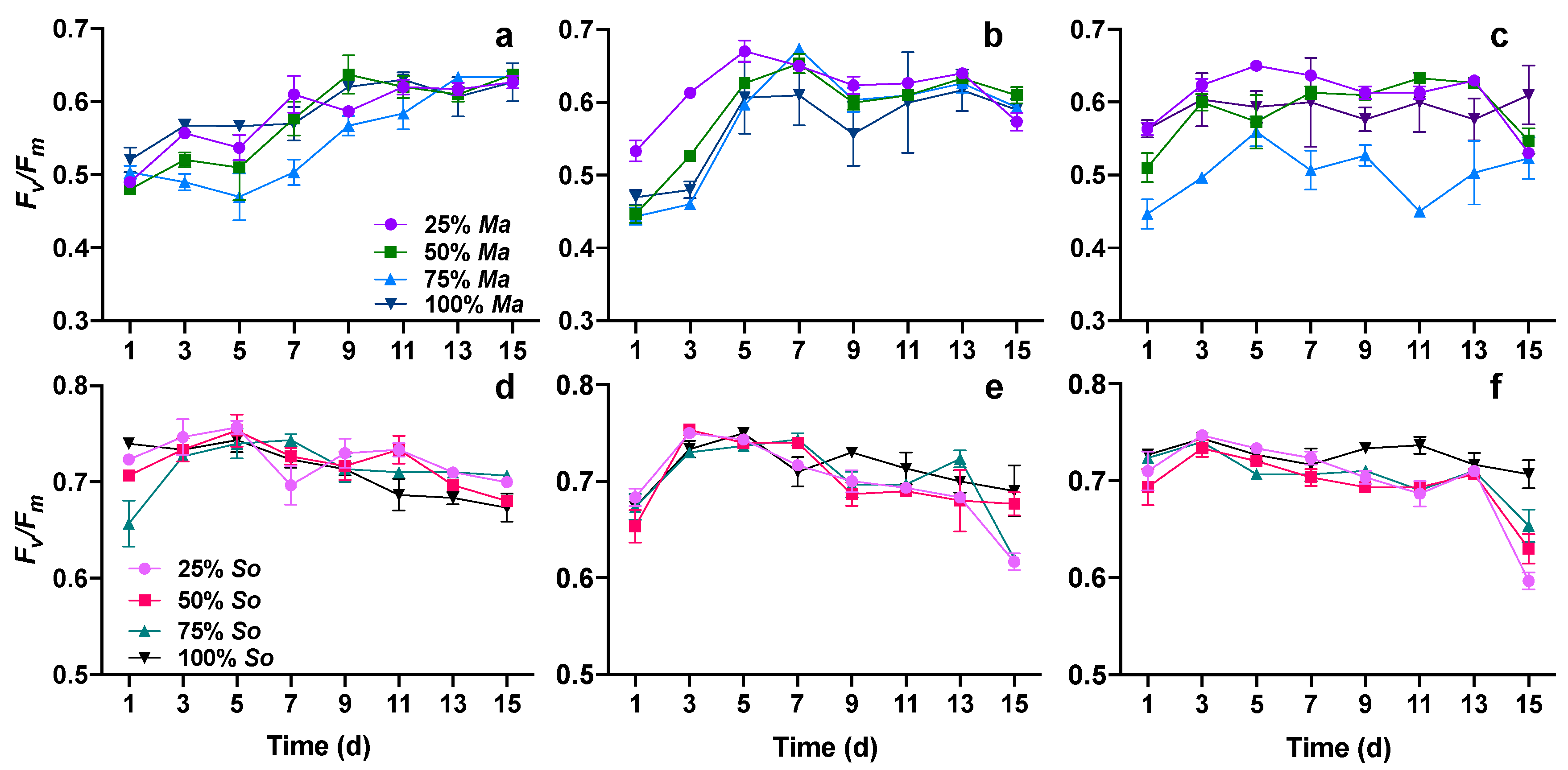
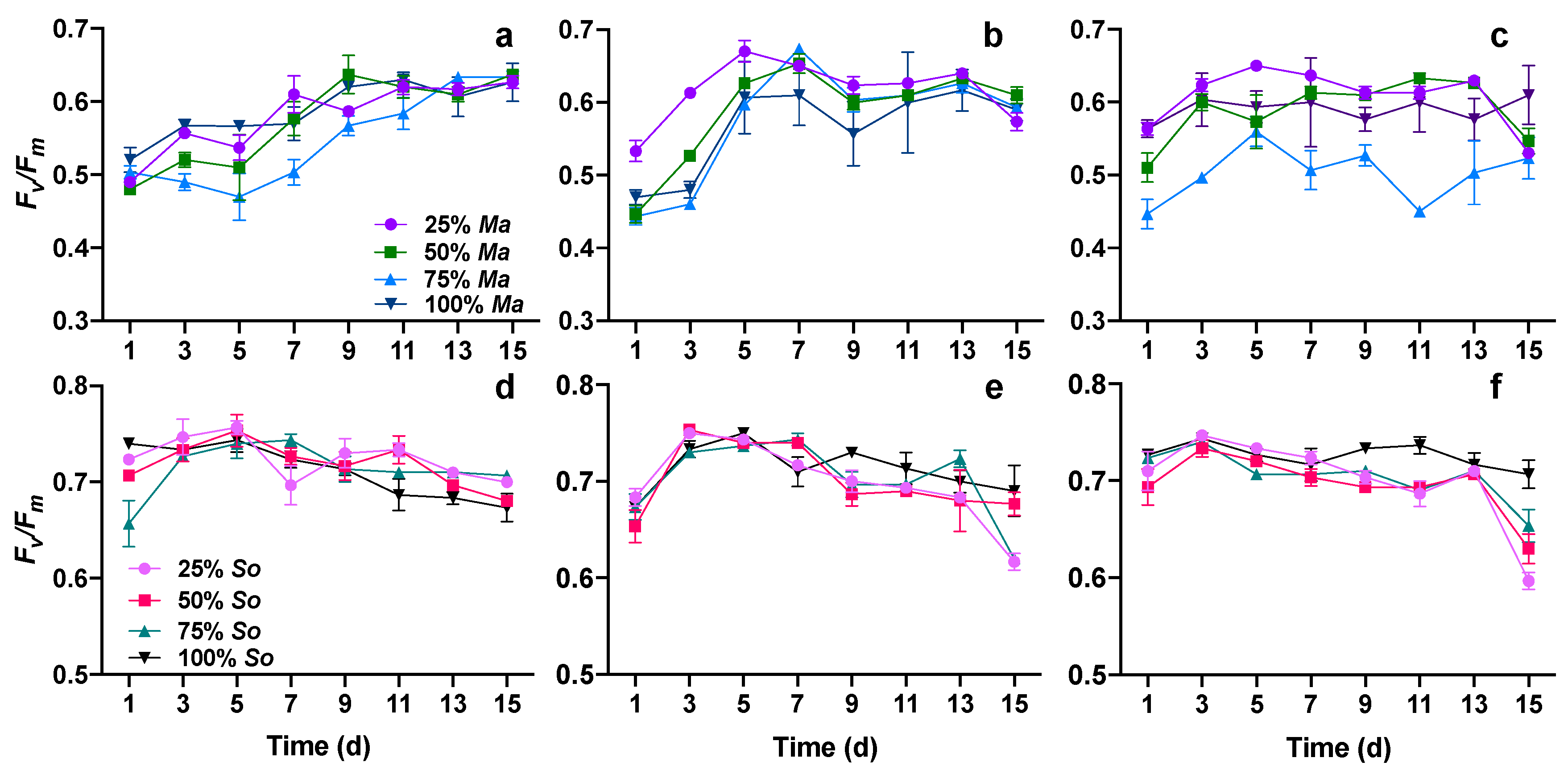
3.3. Photosynthetic Performance of Both Algae in Cocultures
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kaushal, S.S.; Likens, G.E.; Pace, M.L.; Utz, R.M.; Haq, S.; Gorman, J.; Grese, M. Freshwater salinization syndrome on a continental scale. Proc. Natl. Acad. Sci. USA 2018, 115, E574–E583. [Google Scholar] [CrossRef] [Green Version]
- Castillo, A.M.; Sharpe, D.M.; Ghalambor, C.K.; De León, L.F. Exploring the effects of salinization on trophic diversity in freshwater ecosystems: A quantitative review. Hydrobiologia 2018, 807, 1–17. [Google Scholar] [CrossRef]
- Metz, B.; Davidson, O.; Bosch, P.; Dave, R.; Meyer, L. Climate Change 2007: Mitigation of Climate Change; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2007. [Google Scholar]
- Metz, B.; Davidson, O.R.; Bosch, P.R.; Dave, R.; Meyer, L.A. (Eds.) Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; ISBN 9780521880114. [Google Scholar]
- Vidal, N.; Yu, J.; Gutierrez, M.F.; de Mello, F.T.; Tavşanoğlu, Ü.N.; Çakiroglu, A.I.; He, H.; Meerhoff, M.; Brucet, S.; Liu, Z.W.; et al. Salinity shapes food webs of lakes in semiarid climate zones: A stable isotope approach. Inland Waters 2021, 11, 476–491. [Google Scholar] [CrossRef]
- El Hamidi, L.; Larabi, A.; Faouzi, M. Numerical modeling of saltwater intrusion in the rmel-oulad ogbane coastal aquifer (Larache, Morocco) in the climate change and sea-level rise context (2040). Water 2021, 13, 2167. [Google Scholar] [CrossRef]
- El Shinawi, A.; Kuriqi, A.; Zelenakova, M.; Vranayova, Z.; Abd-Elaty, I. Land subsidence and environmental threats in coastal aquifers under sea level rise and over-pumping stress. J. Hydrol. 2022, 608, 127607. [Google Scholar] [CrossRef]
- Cunillera-Montcusí, D.; Beklioğlu, M.; Cañedo-Argüelles, M.; Jeppesen, E.; Ptacnik, R.; Amorim, C.A.; Arnott, S.E.; Berger, S.A.; Brucet, S.; Dugan, H.A.; et al. Freshwater salinisation: A research agenda for a saltier world. Trends Ecol. Evol. 2022, 37, 440–453. [Google Scholar] [CrossRef]
- Talebi, A.F.; Tabatabaei, M.; Mohtashami, S.K.; Tohidfar, M.; Moradi, F. Comparative salt stress study on intracellular ion concentration in marine and salt-adapted freshwater strains of microalgae. Not. Sci. Biol. 2013, 5, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Demetriou, G.; Neonaki, C.; Navakoudis, E.; Kotzabasis, K. Salt stress impact on the molecular structure and function of the photosynthetic apparatus—The protective role of polyamines. BBA-Bioenerg. 2007, 1767, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Christensen, I.; Pedersen, L.K.; Søndergaard, M.; Lauridsen, T.L.; Tserenpil, S.; Richardson, K.; Amorim, C.A.; Pacheco, J.P.; Jeppesen, E. Impact of zooplankton grazing on phytoplankton in north temperate coastal lakes: Changes along gradients in salinity and nutrients. Hydrobiologia 2022. [Google Scholar] [CrossRef]
- He, H.; Jeppesen, E.; Bruhn, D.; Yde, M.; Hansen, K.J.; Spanggaard, L.; Madsen, N.; Liu, W.; Søndergaard, M.; Lauridsen, T.L. Decadal changes in zooplankton biomass, composition and body mass in four temperate shallow brackish lakes subjected to various degrees of eutrophication. Inland Waters 2020, 10, 186–196. [Google Scholar] [CrossRef]
- Jensen, E.; Brucet, S.; Meerhoff, M.; Nathansen, L.; Jeppesen, E. Community structure and diel migration of zooplankton in brackish lakes: Role of salinity and predators. Hydrobiologia 2010, 646, 215–229. [Google Scholar] [CrossRef]
- Szöcs, E.; Coring, E.; Bäthe, J.; Schäfer, R.B. Effects of anthropogenic salinization on biological traits and community composition of stream macroinvertebrates. Sci. Total Environ. 2014, 468, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, G.; Yuan, H.; Chen, J.; Lin, X.; Li, H.; Yu, L.; Wang, C.; Li, L.; Zhuang, Y.; et al. Eukaryotic plankton community assembly and influencing factors between continental shelf and slope sites in the northern South China Sea. Environ. Res. 2023, 216, 114584. [Google Scholar] [CrossRef] [PubMed]
- Omidi, A.; Pflugmacher, S.; Kaplan, A.; Kim, Y.J.; Esterhuizen, M. Reviewing interspecies interactions as a driving force affecting the community structure in lakes via cyanotoxins. Microorganisms 2021, 9, 1583. [Google Scholar] [CrossRef] [PubMed]
- Borics, G.; Abonyi, A.; Salmaso, N.; Ptacnik, R. Freshwater phytoplankton diversity: Models, drivers and implications for ecosystem properties. Hydrobiologia 2021, 848, 53–75. [Google Scholar] [CrossRef]
- Hutchinson, G. The paradox of the plankton. Am. Nat. 1961, 95, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Chattopadhyay, J. Towards a resolution of ‘the paradox of the plankton’: A brief overview of the proposed mechanisms. Ecol. Complex. 2007, 4, 26–33. [Google Scholar] [CrossRef]
- Suttle, C.A.; Chan, A.M.; Cottrell, M.T. Infection of phytoplankton by viruses and reduction of primary productivity. Nature 1990, 347, 467–469. [Google Scholar] [CrossRef]
- Pal, M.; Yesankar, P.J.; Dwivedi, A.; Qureshi, A. Biotic control of harmful algal blooms (HABs): A brief review. J. Environ. Manag. 2020, 268, 110687. [Google Scholar] [CrossRef]
- Dawson, R.M. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Tonk, L.; Bosch, K.; Visser, P.M.; Huisman, J. Salt tolerance of the harmful cyanobacterium Microcystis aeruginosa. Aquat. Microb. Ecol. 2007, 46, 117–123. [Google Scholar] [CrossRef]
- Preece, E.P.; Hardy, F.J.; Moore, B.C.; Bryan, M. A review of microcystin detections in Estuarine and Marine waters: Environmental implications and human health risk. Harmful Algae 2017, 61, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, Y.; Hodoki, Y.; Sano, T.; Tada, K.; Watanabe, M.M. Adaptation of the freshwater bloom-forming cyanobacterium Microcystis aeruginosa to brackish water is driven by recent horizontal transfer of sucrose genes. Front. Microbiol. 2018, 9, 1150. [Google Scholar] [CrossRef] [Green Version]
- Takeya, K.; Kuwata, A.; Yoshida, M.; Miyazaki, T. Effect of dilution rate on competitive interactions between the cyanobacterium Microcystis novacekii and the green alga Scenedesmus quadricauda in mixed chemostat cultures. J. Plankton Res. 2004, 26, 29–35. [Google Scholar] [CrossRef]
- Ji, X.; Verspagen, J.M.; Stomp, M.; Huisman, J. Competition between cyanobacteria and green algae at low versus elevated CO2: Who will win, and why? J. Exp. Bot. 2017, 68, 3815–3828. [Google Scholar] [CrossRef] [Green Version]
- Beardall, J.; Raven, J.A. Cyanobacteria vs green algae: Which group has the edge? Environ. Dev. Sustain. 2017, 68, 3697–3699. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.G.; Santhakumaran, P.; Kookal, S. Phytoplankton communities of eutrophic freshwater bodies (Kerala, India) in relation to the physicochemical water quality parameters. Environ. Dev. Sustain. 2021, 23, 259–290. [Google Scholar] [CrossRef]
- Thorslund, J.; Bierkens, M.F.P.; Oude Essink, G.H.P.; Sutanudjaja, E.H.; van Vliet, M.T.H. Common irrigation drivers of freshwater salinisation in river basins worldwide. Nat. Commun. 2021, 12, 4232. [Google Scholar] [CrossRef]
- Brown, A.F.M.; Dortch, Q.; Van Dolah, F.M.; Leighfield, T.A.; Morrison, W.; Thessen, A.E.; Steidinger, K.; Richardson, B.; Moncreiff, C.A.; Pennock, J.R. Effect of salinity on the distribution, growth, and toxicity of Karenia spp. Harmful Algae 2006, 5, 199–212. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, Y.; Hou, X.; Kong, Q.; Sun, Y.; Wang, J.; Huang, Y.; Yang, Z. High temperature promotes the inhibition effect of Zn2+ on inducible defense of Scenedesmus obliquus. Chemosphere 2019, 216, 203–212. [Google Scholar] [CrossRef]
- Jia, X.; Pan, Y.; Zhu, X. Salinization and heavy metal cadmium impair growth but have contrasting effects on defensive colony formation of Scenedesmus obliquus. Sci. Total Environ. 2023, 862, 160693. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, J.; Lu, Y.; Chen, Q.; Yang, Z. Grazer-induced morphological defense in Scenedesmus obliquus is affected by competition against Microcystis aeruginosa. Sci. Rep. 2015, 5, 12743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Cui, G.; Li, B.; Zhu, X.; Yang, Z. Elevated atmospheric CO2 enhances grazer-induced morphological defense in the freshwater green alga Scenedesmus obliquus. Limnol. Oceanogr. 2018, 63, 1004–1014. [Google Scholar] [CrossRef]
- Martin-Luna, B.; Sevilla, E.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Variation in the synthesis of microcystin in response to osmotic stress in Microcystin aeruginosa PCC 7806. Limnetica 2015, 34, 205–2014. [Google Scholar]
- Black, K.; Yilmaz, M.; Phlips, E.J. Growth and toxin production by Microcystis aeruginosa pcc 7806 (kutzing) lemmerman at elevated salt concentrations. J. Environ. Prot. 2011, 2, 669–674. [Google Scholar] [CrossRef] [Green Version]
- Vergalli, J.; Fayolle, S.; Combes, A.; Franquet, E.; Comte, K. Persistence of microcystin production by Planktothrix agardhii (Cyanobacteria) exposed to different salinities. Phycologia 2020, 59, 24–34. [Google Scholar] [CrossRef]
- Wang, W.; Sheng, Y.; Jiang, M. Physiological and metabolic responses of Microcystis aeruginosa to a salinity gradient. Environ. Sci. Pollut. Res. 2022, 29, 13226–13237. [Google Scholar] [CrossRef]
- Helmus, M.R.; Mercado-Silva, N.; Vander Zanden, M.J. Subsidies to predators, apparent competition and the phylogenetic structure of prey communities. Oecologia 2013, 173, 997–1007. [Google Scholar] [CrossRef]
- Song, H.; Lavoie, M.; Fan, X.; Tan, H.; Liu, G.; Xu, P.; Fu, Z.; Paerl, H.; Qian, H. Allelopathic interactions of linoleic acid and nitric oxide increase the competitive ability of Microcystis aeruginosa. ISME J. 2017, 11, 1865–1876. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Xu, J.; Lu, T.; Zhang, Q.; Qu, Q.; Yang, Z.; Pan, X. Responses of unicellular alga Chlorella pyrenoidosa to allelochemical linoleic acid. Sci. Total Environ. 2018, 625, 1415–1422. [Google Scholar] [CrossRef]
- Yang, J.; Tang, H.; Zhang, X.; Zhu, X.; Huang, Y.; Yang, Z. High temperature and pH favor Microcystis aeruginosa to outcompete Scenedesmus obliquus. Environ. Sci. Pollut. Res. 2018, 25, 4794–4802. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, J.; Dai, M.; Chen, X. Competition between Microcystis aeruginosa and Scenedesmus obliquus under different temperature and light regimes. J. Environ. Manag. 2021, 286, 111809. [Google Scholar]
- von Alvensleben, N.; Magnusson, M.; Heimann, K. Salinity tolerance of four freshwater microalgal species and the effects of salinity and nutrient limitation on biochemical profiles. J. Appl. Phycol. 2016, 28, 861–876. [Google Scholar] [CrossRef]
- Yamamoto, M.; Chiba, T.; Tuji, A. Salinity responses of benthic diatoms inhabiting tidal flats. Diatom Res. 2017, 32, 243–250. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, Y.; Wei, J.; Zhang, X.; Zhang, L.; Yang, Z.; Huang, Y. Ultraviolet-B radiation stress alters the competitive outcome of algae: Based on analyzing population dynamics and photosynthesis. Chemosphere 2021, 272, 129645. [Google Scholar] [CrossRef]
- Zhang, M.; Kong, F.; Xing, P.; Tan, X. Effects of Interspecific Interactions between Microcystis aeruginosa and Chlorella pyrenoidosa on Their Growth and Physiology. Int. Rev. Hydrobiol. 2007, 92, 281–290. [Google Scholar] [CrossRef]
- Hernández-Zamora, M.; Santiago-Martínez, E.; Martínez-Jerónimo, F. Toxigenic Microcystis aeruginosa (Cyanobacteria) affects the population growth of two common green microalgae: Evidence of other allelopathic metabolites different to cyanotoxins. J. Phycol. 2021, 57, 1530–1541. [Google Scholar] [CrossRef]
- Zak, A.; Musiewicz, K.; Kosakowska, A. Allelopathic activity of the Baltic cyanobacteria against microalgae. Estuar. Coast. Shelf Sci. 2012, 112, 4–10. [Google Scholar] [CrossRef]
- Sychrová, E.; Štěpánková, T.; Nováková, K.; Bláha, L.; Giesy, J.P.; Hilscherová, K. Estrogenic activity in extracts and exudates of cyanobacteria and green algae. Environ. Int. 2012, 39, 134–140. [Google Scholar] [CrossRef]
- Li, B.; Yin, Y.; Kang, L.; Feng, L.; Liu, Y.; Du, Z.; Tian, Y.; Zhang, L. A review: Application of allelochemicals in water ecological restoration—Algal inhibition. Chemosphere 2021, 267, 128869. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity | Algae | 25% So + 75% Ma | 50% So + 50% Ma | 75% So + 25% Ma |
|---|---|---|---|---|
| 0 | Ma | R2 = 0.6799 | R2 = 0.6004 | R2 = 0.5069 |
| So | R2 = 0.9180 | R2 = 0.9543 | R2 = 0.9140 | |
| Y(t) | R2 = 0.9398 | R2 = 0.8693 | R2 = 0.8013 | |
| 3 | Ma | R2 = 0.6753 | R2 = 0.5851 | R2 = 0.1538 |
| So | R2 = 0.9041 | R2 = 0.9003 | R2 = 0.9016 | |
| Y(t) | R2 = 0.8627 | R2 = 0.8746 | R2 = 0.8574 | |
| 6 | Ma | R2 = 0.6174 | R2 = 0.5690 | R2 = 0.4568 |
| So | R2 = 0.8361 | R2 = 0.9269 | R2 = 0.7749 | |
| Y(t) | R2 = 0.9349 | R2 = 0.8829 | R2 = 0.7747 |
| Algae | Factors | Nmax | tmax | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SS | DF | MS | F | p Value | SS | DF | MS | F | p Value | ||
| Microcystis | Salinity | 2.014 | 2 | 1.007 | 0.1493 | 0.8624 | 173.7 | 2 | 86.85 | 19.14 | <0.0001 |
| Composition | 5.766 | 2 | 2.883 | 0.4273 | 0.6587 | 2.169 | 2 | 1.085 | 0.2391 | 0.7898 | |
| Interaction | 2.319 | 4 | 0.5799 | 0.08594 | 0.9857 | 44.16 | 4 | 11.04 | 2.434 | 0.085 | |
| Scenedesmus | Salinity | 25.62 | 2 | 12.81 | 5.596 | 0.0129 | 47.52 | 2 | 23.76 | 10.43 | 0.001 |
| Composition | 9.455 | 2 | 4.727 | 2.065 | 0.1558 | 7.698 | 2 | 3.849 | 1.689 | 0.2126 | |
| Interaction | 2.877 | 4 | 0.7193 | 0.3142 | 0.8647 | 6.962 | 4 | 1.74 | 0.7640 | 0.5623 | |
| Algae | Source of Variation | SS | DF | MS | F | p Value |
|---|---|---|---|---|---|---|
| Microcystis | Time | 0.437 | 7 | 0.0625 | 47.108 | <0.001 |
| Salinity | 0.0153 | 2 | 0.00765 | 5.773 | 0.004 | |
| Proportion of Ma | 0.0531 | 3 | 0.0177 | 13.341 | <0.001 | |
| Time × Salinity | 0.185 | 14 | 0.0132 | 9.973 | <0.001 | |
| Time × Proportion of Ma | 0.0773 | 21 | 0.00368 | 2.778 | <0.001 | |
| Salinity × Proportion of Ma | 0.0242 | 6 | 0.00403 | 3.043 | 0.007 | |
| Time × Salinity × Proportion of Ma | 0.0847 | 42 | 0.00202 | 1.522 | 0.031 | |
| Scenedesmus | Time | 0.152 | 7 | 0.0217 | 62.987 | <0.001 |
| Salinity | 0.00898 | 2 | 0.00449 | 13.009 | <0.001 | |
| Proportion of So | 0.00543 | 3 | 0.00181 | 5.243 | 0.002 | |
| Time × Salinity | 0.0285 | 14 | 0.00203 | 5.889 | <0.001 | |
| Time × Proportion of So | 0.0233 | 21 | 0.00111 | 3.216 | <0.001 | |
| Salinity × Proportion of So | 0.0115 | 6 | 0.00191 | 5.539 | <0.001 | |
| Time × Salinity × Proportion of So | 0.0421 | 42 | 0.001 | 2.906 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, T.; Li, Y.; Xue, W.; Pan, Y.; Zhu, X. Freshwater Salinization Impacts the Interspecific Competition between Microcystis and Scenedesmus. Water 2023, 15, 1331. https://doi.org/10.3390/w15071331
Gao T, Li Y, Xue W, Pan Y, Zhu X. Freshwater Salinization Impacts the Interspecific Competition between Microcystis and Scenedesmus. Water. 2023; 15(7):1331. https://doi.org/10.3390/w15071331
Chicago/Turabian StyleGao, Tianheng, Yinkang Li, Wenlei Xue, Yueqiang Pan, and Xuexia Zhu. 2023. "Freshwater Salinization Impacts the Interspecific Competition between Microcystis and Scenedesmus" Water 15, no. 7: 1331. https://doi.org/10.3390/w15071331