
Diet and Trophic Structure of the Fish Community in a Small Sub-Tropical Lake in Central Mexico

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
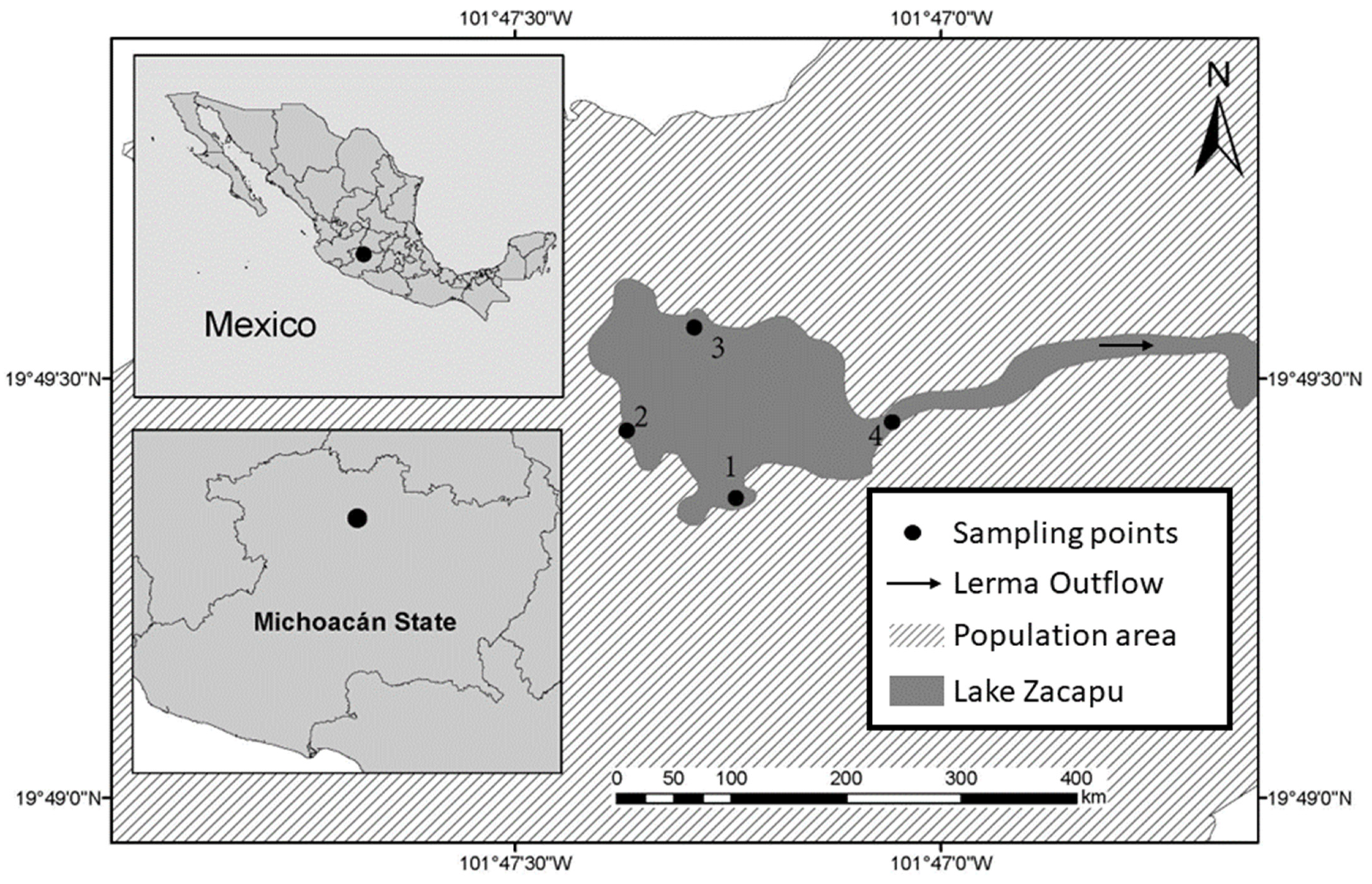
2.1. Study Area
2.2. Environmental Variables
2.3. Fish Collection
2.4. Invertebrate Collection
2.5. Laboratory Analysis
2.6. Data Analysis
2.6.1. Environmental Variables
2.6.2. Diet Analysis
2.6.3. Stable Isotopes
3. Results
3.1. Environmental Variables
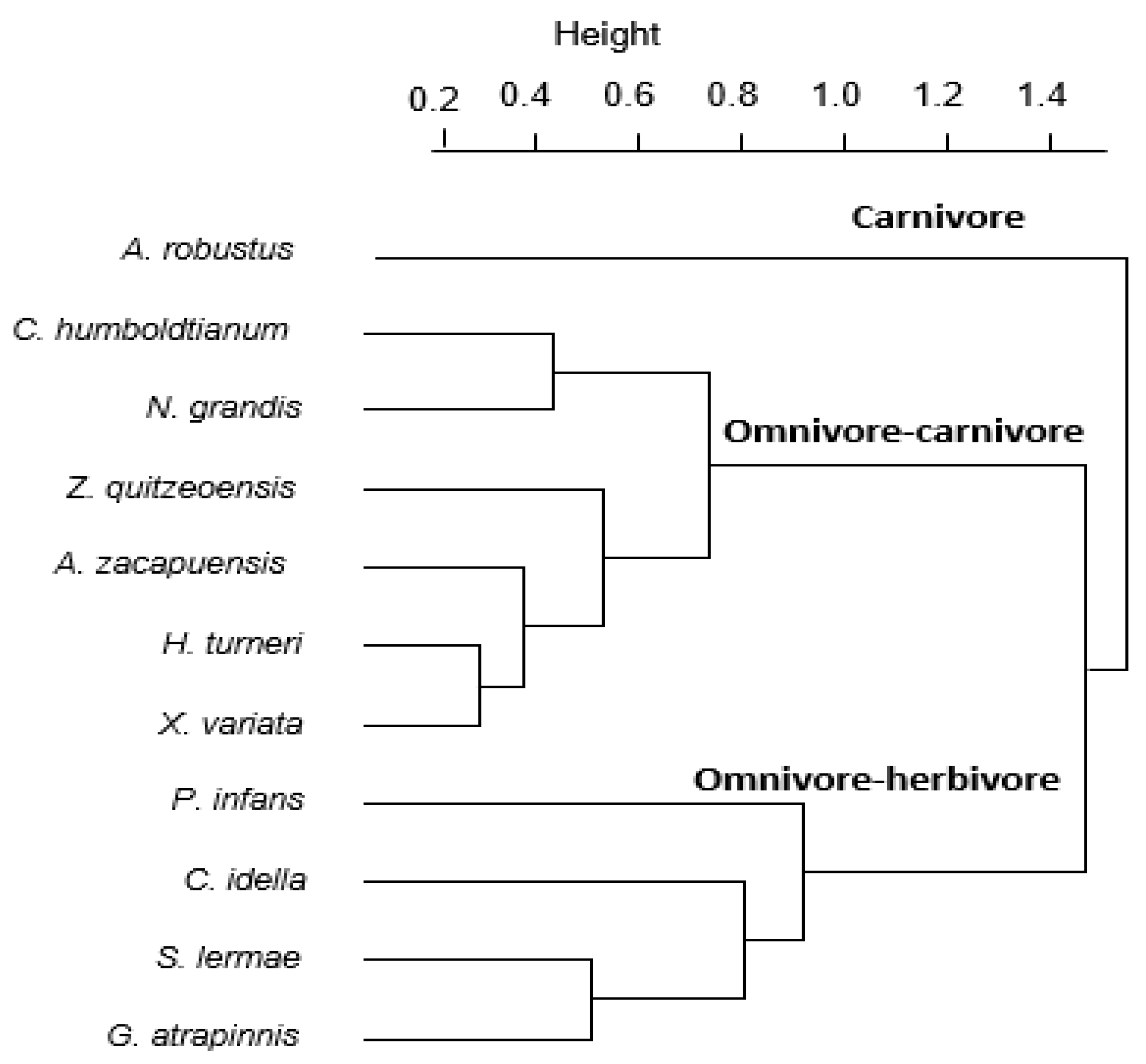
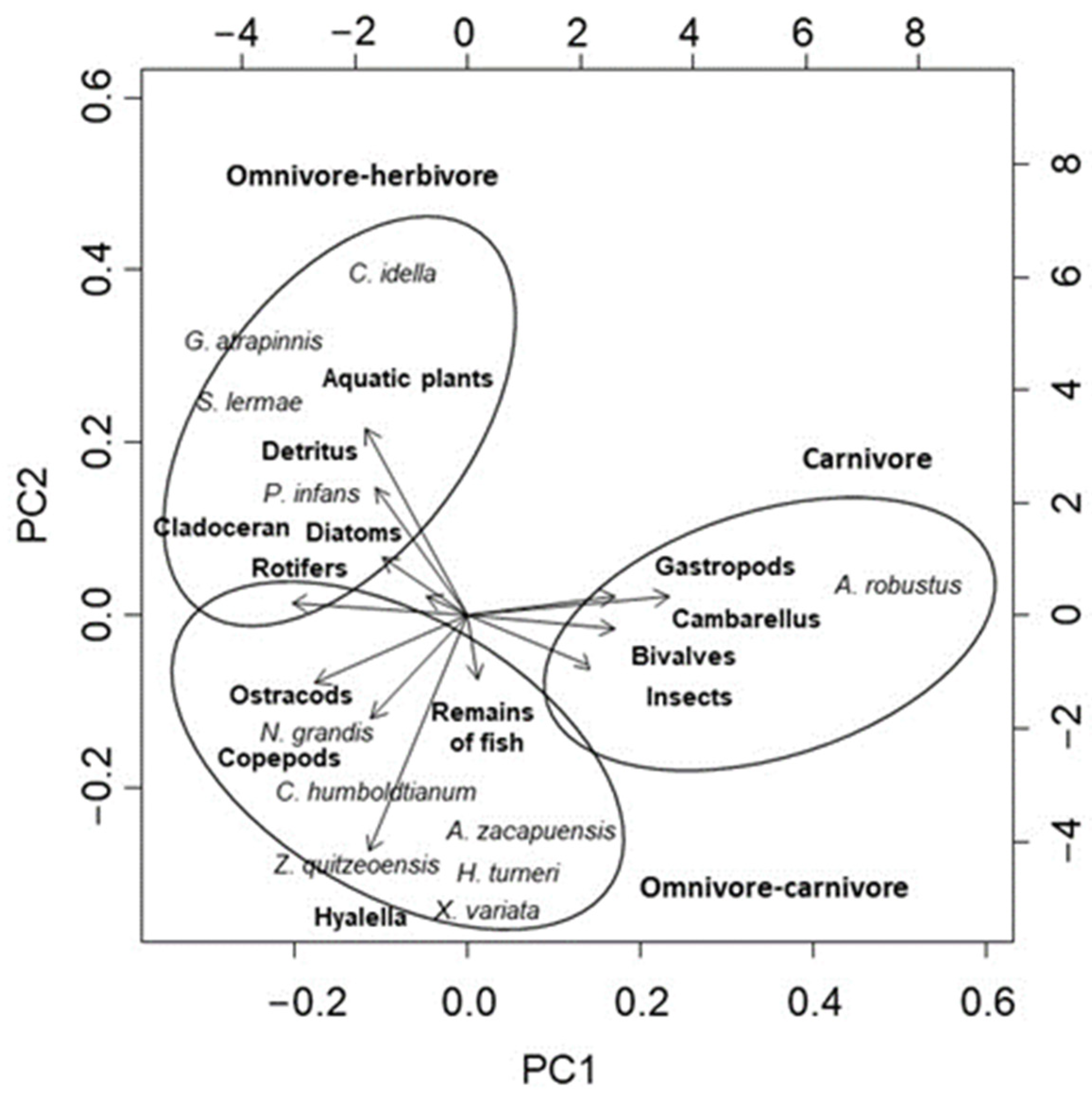
3.2. Diet Analysis
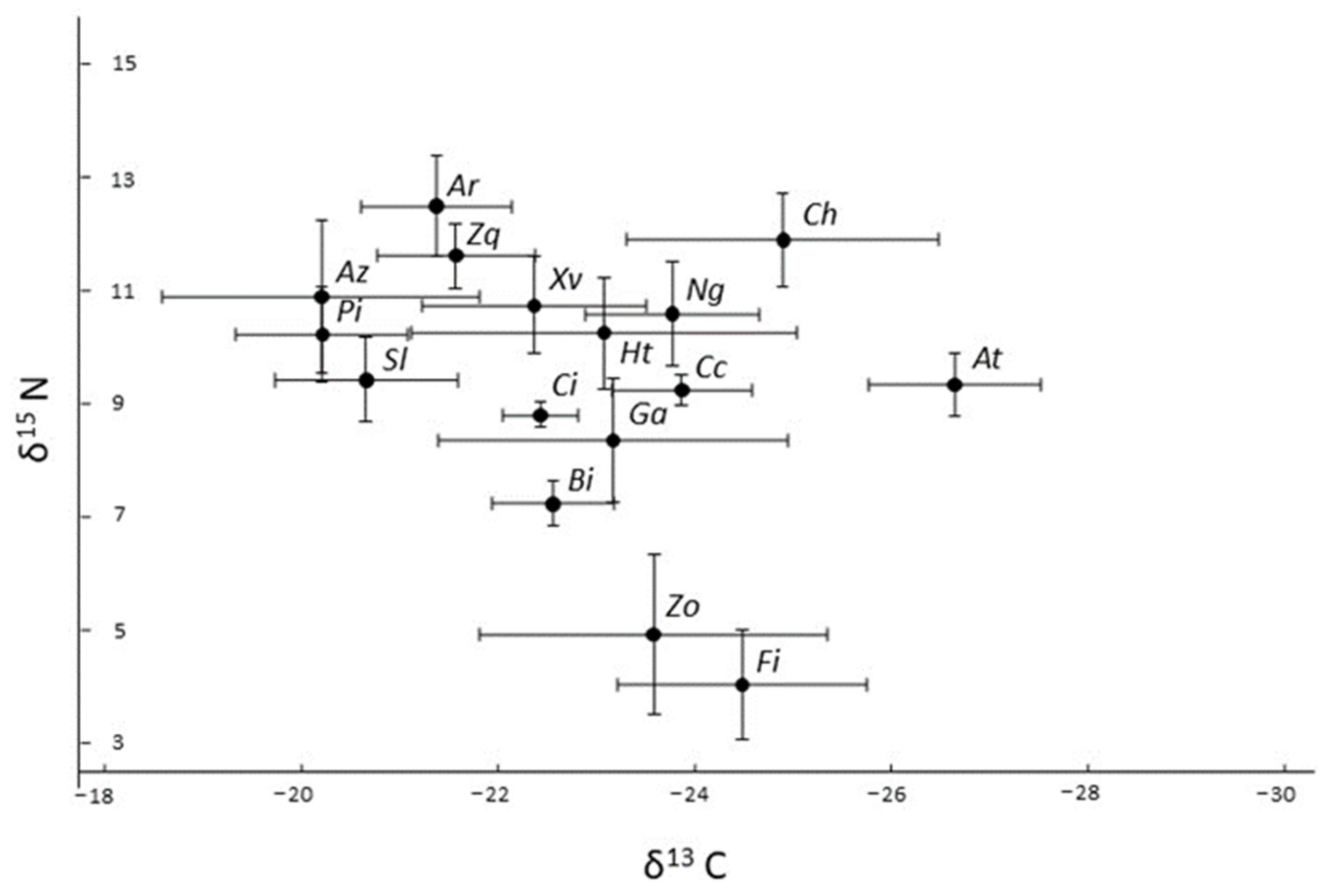
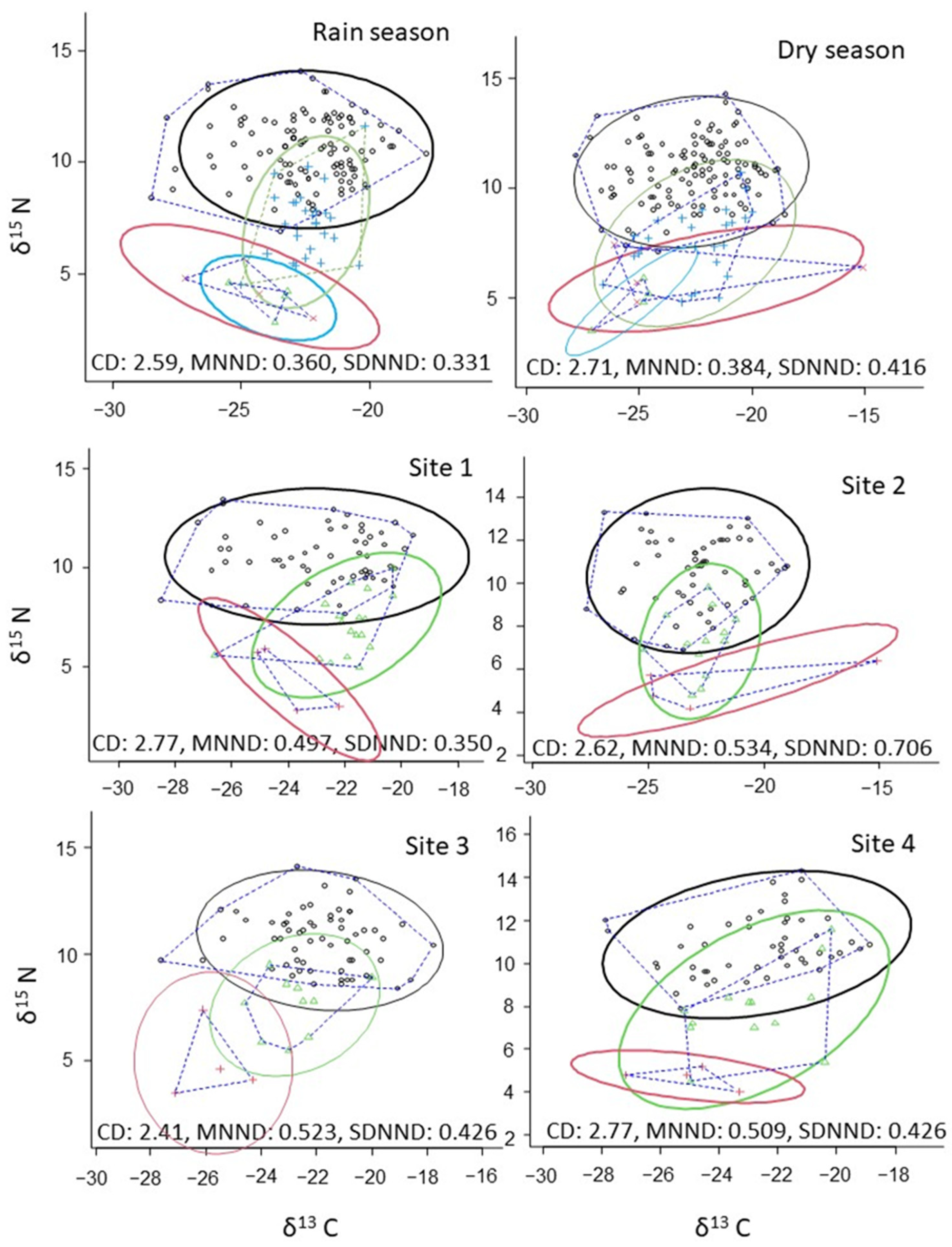
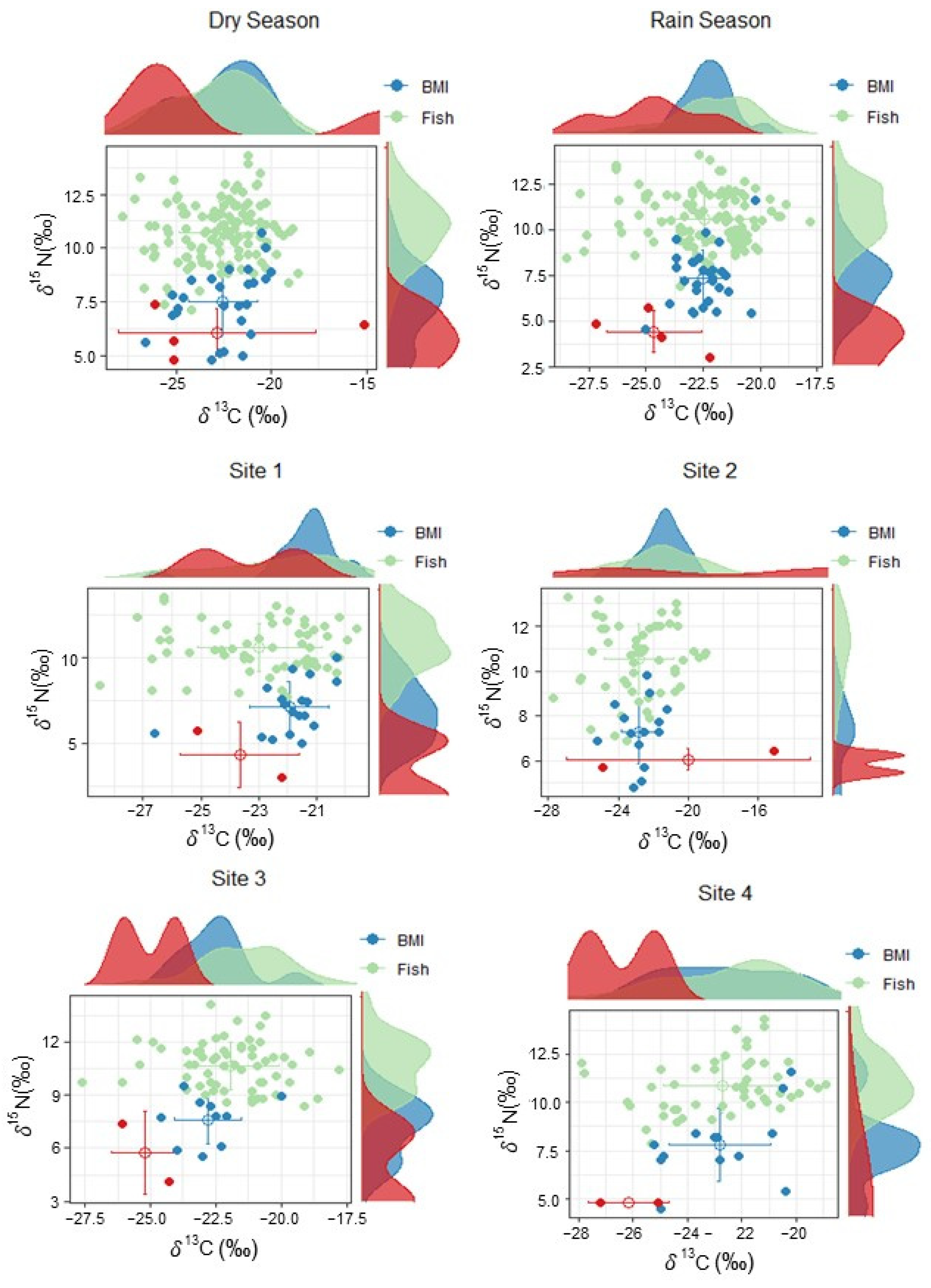
3.3. Stable Isotopes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biggs, J.; von Fumetti, S.; Kelly-Quinn, M. The importance of small waterbodies for biodiversity and ecosystem services: Implications for policy makers. Hydrobiologia 2017, 793, 3–39. [Google Scholar] [CrossRef]
- Hoverman, J.T.; Johnson, P.T.J. Ponds and Lakes: A Journey Through the Life Aquatic. Nat. Sci. Educ. 2012, 3, 17. [Google Scholar]
- Strong, D.R.; Frank, K.T. Human Involvement in Food Webs. Annu. Rev. Environ. Resour. 2010, 35, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Crowder, L.B.; Ray, R.; Drenner, W.; Chair, W.; Kerfoot, C.; McQueen, D.J.; Mills, E.; Sommer, U.; Spencer, C.N.; Vanni, M.J. Food web interactions in lakes. In Interactions in Lake Communities; Carpenter, S.R., Ed.; Springer: New York, NY, USA, 1988. [Google Scholar] [CrossRef]
- Delong, M.; Thorp, J.M.; Thons, M.S.; Mcintosh, L. Trophic niche dimensions of fish communities as a function of historical hydrological conditions in a Plains River. River Syst. 2011, 19, 177–187. [Google Scholar] [CrossRef]
- Fauvelle, C.; Diepstraten, R.; Jessen, T. A meta-analysis of home range studies in the context of trophic levels: Implications for policy-based conservation. PLoS ONE 2017, 12, e0173361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrison, L.P.; Link, J.S. Fishing effects on spatial distribution and trophic guild structure of the fish community in the Georges Bank region. J. Mar. Sci. 2000, 57, 723–730. [Google Scholar] [CrossRef] [Green Version]
- Dantas, D.V.; Barletta, M.; Ramos, J.d.A.A.; Lima, A.R.A.; Costa, M.F. Seasonal diet shifts and overlap between two sympatric catfishes in an estuarine nursery. Estuar. Coast. 2013, 36, 237–256. [Google Scholar] [CrossRef]
- Pianka, E.R. Competition and niche theory. In Theoretical Ecology: Principles and Applications; May, R.M., Ed.; Blackwell Scientific Publication: Oxford, UK, 1981; pp. 167–196. [Google Scholar]
- Sánchez-Hernández, J.; Vieira-Lanero, R.; Servia, M.J.; Cobo, F. Feeding habits of four sympatric fish species in the Iberian Peninsula: Keys to understanding coexistence using prey traits. Hydrobiologia 2011, 667, 119–132. [Google Scholar] [CrossRef]
- Carrara, F.; Giometto, A.; Seymour, M.; Rinado, A.; Altermatt, F. Inferring species interactions in ecological communities: A comparison of methods at different levels of complexity. Methods Ecol. Evol. 2015, 6, 895–906. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montaña, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Córdova-Tapia, F.; Contreras, M.; Zambrano, L. Trophic niche overlap between native and non-native fishes. Hydrobiologia 2015, 746, 291–301. [Google Scholar] [CrossRef]
- Cicala, D.; Polgar, G.; Mor, J.R.; Piscia, R.; Brignone, S.; Zaupa, S.; Volta, P. Trophic Niches, Trophic Positions, and Niche Overlaps between Non-Native and Native Fish Species in a Subalpine Lake. Water 2020, 12, 3476. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef] [Green Version]
- Burns, J.H.; Strauss, S. More Closely related species are more ecologically similar in an experimental test. Proc. Natl. Acad. Sci. USA 2011, 108, 5302–5307. [Google Scholar] [CrossRef] [Green Version]
- Hussey, N.E.; MacNeil, M.A.; McMeans, B.C.; Olin, J.A.; Dudley, S.F.; Cliff, G.; Wintner, S.P.; Fennessy, S.T.; Fisk, A.T. Rescaling the trophic structure of marine food webs. Ecol. Lett. 2014, 17, 239–250. [Google Scholar] [CrossRef]
- Romanuk, T.N.; Hayward, A.; Hutchings, J.A. Trophic level scales positively with body size in fishes. Glob. Ecol. Biogeogr. 2011, 20, 231–240. [Google Scholar] [CrossRef]
- Eloranta, A.T.; Nieminen, P.; Kimmo, K.; Kahilainen, K. Trophic interactions between introduced lake trout (Salvelinus namaycush) and native Arctic charr (S. alpinus) in a large Fennoscandian subarctic lake. Ecol. Freshw. Fish 2015, 24, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.M.; Blanchette, M.L.; Pusey, B.J.; Jardine, T.D.; Pearson, R.G. Gut content and stable isotope analyses provide com- plementary understanding of ontogenetic dietary shifts and trophic relationships among fishes in a tropical river. Freshw. Biol. 2012, 57, 2156–2172. [Google Scholar] [CrossRef]
- Svanbäck, R.; Quevedo, M.; Olsson, J.; Eklöv, P. Individuals in food webs: The relationships between trophic position, omnivory and among-individual diet variation. Oecologia 2015, 178, 103–104. [Google Scholar] [CrossRef] [Green Version]
- Vander Zanden, M.J.; Rasmussen, J.B. Primary Consumer S13C and S15N and the trophic position of aquatic consumers. Ecology 1999, 80, 1395–1404. [Google Scholar] [CrossRef]
- Fry, B. Stable Isotope Ecology; Springer: New York, NY, USA, 2006; Volume 521. [Google Scholar] [CrossRef]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Yeager, L.A. Applying stable isotopes to examine food- web structure: An overview of analytical tools. Biol. Rev. 2012, 87, 545–562. [Google Scholar] [CrossRef]
- Hill, M.J.; Mathers, K.L.; Wood, P.J. The aquatic macroinvertebrate biodiversity of urban ponds in a medium-sized European town (Loughborough, UK). Hydrobiologia 2015, 760, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Pilger, T.J.; Gido, K.B.; Propst, D.L. Diet and trophic niche overlap of native and nonnative fishes in the Gila River, USA: Implications for native fish conservation. Ecol. Freshw. Fish 2010, 19, 300–321. [Google Scholar] [CrossRef]
- Thomas, S.; Kijunen, M.; Malinen, T.; Eloranta, A.; Amundsen, P.; Lodenius, M.; Kahilainen, K. Food-web structure and mercury dynamics in a large subarctic lake following multiple species introductions. Freshw. Biol. 2016, 61, 500–517. [Google Scholar] [CrossRef]
- Sánchez-Hernández, J.; Eloranta, A.; Finstad, A.; Amundsen, P. Community structure affects trophic ontogeny in a predatory fish. Ecol. Evol. 2017, 7, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Laske, S.; Rosenberger, A.; Wipfi, M.; Zimmeman, C. Generalist feeding strategies in Arctic freshwater fish: A mechanism for dealing with extreme environments. Ecol. Freshw. Fish 2018, 27, 767–784. [Google Scholar] [CrossRef]
- Mwijage, A.P.; Shila, D.A.; Machiwa, J.F. Differences in trophic resources and niches of two juvenile predatory species in three Pangani estuarine zones, Tanzania: Stomach contents and stable isotope approaches. J. Biol. Res. 2018, 25, 13. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.I.; Waldron, S. Combined stable isotope and gut contents analysis of food webs in plant-dominated, shallow lakes. Freshw. Biol. 2003, 48, 1396–1407. [Google Scholar] [CrossRef] [Green Version]
- Franssen, N.R.; Gido, K.B. Use of stable isotopes to test literature-based trophic classifications of small-bodied stream fishes. Am. Midl. Nat. 2006, 156, 1–10. [Google Scholar] [CrossRef]
- Ayala-Ramírez, G.L.; Ruiz-Sevilla, G.; Chacon-Torres, A. La Laguna de Zacapu, Michoacán. In Las Aguas Interiores de México: Conceptos y Casos; De la Lanza, G., Ed.; AGT EDITOR, S.A.: Mexico City, Mexico, 2007; pp. 268–284. [Google Scholar]
- Domínguez-Domínguez, O.; Zambrano, L.; Escalera-Vázquez, L.H.; Pérez-Rodríguez, R.; Pérez-Ponce de León, G. Changes in the distribution of goodeids (Osteichthyes: Cyprinodontiformes: Goodeidae) in river basin of Central Mexico. Rev. Mex. Biodivers. 2008, 79, 501–512. [Google Scholar] [CrossRef]
- Ramírez-García, A.; Moncayo-Estrada, R.; González-Cárdenas, J.J.; Domínguez-Domínguez, O. Reproductive cycle of native viviparous fish species (Actinopterygii: Cyprinodontiformes: Goodeidae) in a subtropical Mexican lake. Neotrop. Ichthyol. 2021, 19, e210105. [Google Scholar] [CrossRef]
- Ramírez-García, A.; Domínguez-Domínguez, O.; De La Cruz-Agüero, J.; López-López, E.; Moncayo-Estrada, R. Temporary changes in the structure of the fish community separated by decades in a small subtropical lake. Ecol. Freshw. Fish 2022, 32, 53–63. [Google Scholar] [CrossRef]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). La biodiversidad en Michoacán. In Estudio de Estado 2; CONABIO: Mexico City, Mexico, 2019. [Google Scholar]
- Zubieta-Rojas, T.; Alvarado-Villanueva, R.; Ortega-Murillo, M.R.; Medina-Nava, M.; Sánchez-Heredia, J.D. Plan de Manejo del área Natural Protegida “Laguna de Zacapu y su Ribera”; Comisión Nacional de Áreas Naturales Protegidas (CONANP): Mexico City, Mexico, 2005. [Google Scholar]
- Moncayo-Estrada, R. Estructura y Función de la Comunidad de Peces de la Laguna de Zacapu, Michoacán, México. Master’s Thesis, Instituto Politécnico Nacional, Mexico City, Mexico, 1996. Available online: https://www.repositoriodigital.ipn.mx/handle/123456789/15234 (accessed on 13 April 2021).
- Ramírez-Herrejón, J.P. Análisis Temporal de la Calidad Ambiental de Los Ecosistemas Acuáticos en la Sub-Cuenca del río Angulo, Cuenca Lerma-Chapala. Master’s Thesis, Universidad Michoacana de San Nicolás de Hidalgo, Mexico City, Mexico, 2008. [Google Scholar]
- Valencia-Vargas, R.; Escalera-Vázquez, L.H. Abundancia de la salamandra Ambystoma andersoni con relación a la dinámica estacional y heterogeneidad espacial en el lago de Zacapu, Michoacán, México. Rev. Mex. Biodivers. 2021, 92, e923283. [Google Scholar] [CrossRef]
- Miller, R.R.; Minckley, W.L.; Norris, S.M. Peces dulceacuícolas de México; CONABIO: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2009. [Google Scholar]
- Merritt, R.; Cummins, K.; Berg, M.B. An Introduction to the Aquatic Insects of North America, 5th ed.; Kendall Hunt Publishing Company: Dubuque, IA, USA, 2019; p. 1498. [Google Scholar]
- Thorp, J.H.; Covich, A.P. Ecology and Classification of North American Freshwater Invertebrates; Academic Press: London, UK, 2010. [Google Scholar]
- Ferry, L.A.; Cailliet, G. Sample size and data analysis are we characterizing and comparing diet properly. In Feeding Ecology and Nutrition in Fish Symposium Proceedings, International Congress on the Biology of Fishes; San Francisco State University: San Francisco, CA, USA, 1996; pp. 14–18. [Google Scholar]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with tradition: Redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1896). Environ. Biol. Fishes 2012, 95, 3–20. [Google Scholar] [CrossRef]
- Pauly, D.; Froese, R.; Sala, P.; Palomares, M.L.; Christensen, V.; Rius, J. Trophlab Manual; ICLARM: Manila, Philippines, 2000. [Google Scholar]
- Christensen, V.; Pauly, D. ECOPATH II-a software for balancing steady-state ecosystem models and calculating network characteristics. Ecol. Model 1992, 61, 169–185. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology; Harper and Row: New York, NY, USA, 1989. [Google Scholar]
- Jackson, A.; Parnell, A. Package SIBER: Stable Isotope Bayesian Ellipses in R. Cran-R-Project. 2021. Available online: https://cran.r-project.org/web/packages/SIBER/SIBER.pdf (accessed on 15 June 2021).
- Abrantes, K.; Johnston, R.; Connolly, R.; Sheaves, M. Importance of Mangrove Carbon for Aquatic Food Webs in Wet–Dry Tropical Estuaries. Estuaries Coasts 2014, 39, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER-Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Quezada-Romegialli, C.; Jackson, A.L.; Hayden, B.; Kahilainen, K.K.; Lopes, C.; Harrod, C. tRophicPosition, an R package for the Bayesian estimation of trophic position from consumer stable isotope ratios. Methods Ecol. Evol. 2018, 9, 1592–1599. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-García, A.; Pérez-Munguía, R.M.; Domínguez-Domínguez, O. Spatial and temporal variation patterns of aquatic macroinvertebrates in a subtropical lake in the central Mexico. Rev. Biol. Trop. 2023, in press. [Google Scholar]
- Bean, P.T.; Williams, C.S.; Diaz, P.H.; Bonner, T.H. Habitat associations, life history, and diet of the blackspot shiner, Notropis atrocaudalis. Southeast. Nat. 2010, 9, 673–686. [Google Scholar] [CrossRef]
- Moncayo-Estrada, R. Análisis histórico de la biología de la Cherehuita (Hubbsina turneri) (Pisces: Goodeidae), especie endémica y en peligro de extinción de México. Rev. Chapingo Ser. Cienc. For. Ambiente 2012, 18, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Domínguez, O.; Pérez-Rodríguez, R.; Escalera-Vázquez, L.H.; Doadrio, I. Two new species of the genus Notropis Rafinesque, 1817 (Actinopterygii, Cyprinidae) from the Lerma River Basin in Central Mexico. Hidrobiológica 2009, 19, 159–172. [Google Scholar]
- Moncayo-Estrada, R.; De La Cruz-Agüero, J.; López-López, E.; Del Monte-Luna, P.; Díaz-Argüero, M.M.; Chacón-Torres, A.; Ramírez-García, A.; Domínguez-Domínguez, O.; Ramírez-Herrejón, J.P. Historical analysis of an imperiled fish species: Environmental variables modeling, biotic interactions, extirpation, and current restricted-range. Front. Biosci. 2022, 27, 165. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.K.; Radda, A.C.; Domínguez-Domínguez, O. Notes on the genera Neoophorus HUBBS & TURNER, 1937 and Allotoca HUBBS & TURNER, 1937, with a description of a new species of Allotoca from Laguna de Zacapu, Michoacán, Mexico (Teleostei, Cyprinodontiformes: Goodeidae). Ann. Nat. Hist. Mus. Wien. 2001, 103B, 453–460. [Google Scholar]
- German, D.P.; Horn, M.H. Gut length and mass in herbivorous and carnivorous prickleback fishes (Teleostei: Stichaeidae): Ontogenetic, dietary, and phylogenetic effects. Mar. Biol. 2006, 148, 1123–1134. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.A.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12, 692–715. [Google Scholar] [CrossRef]
- Mérona, B.D.; Rankin-de-Mérona, J. Food resource partitioning in a fish community of the central Amazon floodplain Bernard. Neotrop. Ichthyol. 2004, 2, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Herrejón, J.P.; Zambrano, L.; Mercado-Silva, N.; Torres-Téllez, A.; Pineda-García, F.; Caraveo-Patiño, J.; Balart, E.F. Long term changes in the fish fauna of Lago de Pátzcuaro in Central México. Lat. Am. J. Aquat. Res. 2014, 42, 137–149. [Google Scholar] [CrossRef]
- Zambrano, L.; Valiente, E.; Vander Zanden, M.J. Food web overlap among native axolotl (Ambystoma mexicanum) and two non-native fishes: Carp (Cyprinus carpio) and tilapia (Oreochromis niloticus) in Xochimilco, Mexico City. Biol. Invasions 2010, 12, 3061–3069. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilá, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Orbe-Mendoza, A.A.; Acevedo-García, J.; Lyons, J. Lake Patzcuaro fishery management plan. Rev. Fish Biol. Fish. 2002, 12, 207–217. [Google Scholar] [CrossRef]
- Post, P.M.; Pace, M.L.; Hairston, N.G. Ecosystem size determines food-chains in lakes. Nature 2000, 405, 1047–1049. [Google Scholar] [CrossRef]
- Jia, Y.; Jiang, Y.; Liu, Y.; Sui, X.; Feng, X.; Zhu, R.; Chen, Y. Understanding trophic structure variation in fish assemblages of subtropical shallow lakes: Combined effects of ecosystem size, productivity, and disturbance. Ecol. Indic. 2021, 129. [Google Scholar] [CrossRef]
- Mercado-Silva, N.; Lyons, J.; Moncayo-Estrada, R.; Gesundheit, P.; Krabbenhoft, T.J.; Powell, D.L.; Piller, K.R. Stable isotope evidence for trophic overlap of sympatric Mexican Lake Chapala silversides (Teleostei: Atherinopsidae: Chirostoma spp.). Neotrop. Ichthyol. 2015, 13, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Herrejón, J.P.; Moncayo-Estrada, R.; Balart, E.F.; García-Camacho, L.A.; Vital-Rodríguez, B.; Alvarado-Villanueva, R.; Ortega-Murillo, R.; Caraveo-Patiño, J. Trophic interrelations between introduced common carp, Cyprinus carpio (Actinopterygii: Cypriniformes: Cyprinidae), and fish community in a eutrophic shallow lake. Acta Ichthyol. Piscat. 2014, 44, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Hernández, J.; Amundsen, P. Ecosystem type shapes trophic position and omnivory in fishes. Fish Fish. 2018, 19, 1003–1015. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.H.; Schelske, C.L.; Hoyer, M.V. Intrapopulation feeding diversity in blue tilapia: Evidence from stable-isotope analyses. Ecology 1997, 78, 2263–2266. [Google Scholar]
- Kondoh, M. Foraging adaptation and the relationship between food-web complexity and stability. Science 2003, 299, 1388–1391. [Google Scholar] [CrossRef]
- Kao, Y.C.; Rogers, M.W.; Bunnell, D.B.; Cowx, I.G.; Qian, S.S.; Anneville, O.; Beard, T.D., Jr.; Brinker, A.; Britton, J.R.; Chura-Cruz, R.; et al. Effects of climate and land-use changes on fish catches across lakes at a global scale. Nat. Commun. 2020, 11, 2526. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Dry Season | Wet Season |
|---|---|---|
| Temp | 18.5 ± 1.5 | 19.9±0.7 |
| Season | x2 = 3.6486, p = 0.056 | |
| Sites | x2 = 1.723, p = 0.631 | |
| DO | 13.7±2.7 | 14.3±0.2 |
| Season | x2 = 0.2757, p = 0.599 | |
| Sites | x2 = 9.3971, p = 0.064 | |
| pH | 7.7±0.4 | 7.3±0.2 |
| Season | x2 = 6.419, p = 0.011 * | |
| Sites | x2 = 0.74108, p = 0.86 | |
| TDS | 107.9±2.1 | 109.2±2.4 |
| Season | x2 = 0.80397, p = 0.37 | |
| Sites | x2 = 1.0738, p = 0.78 | |
| Cond | 136.7±2.6 | 138.8±1.8 |
| Season | x2 = 1.875, p = 0.17 | |
| Sites | x2 = 1.0429, p = 0.79 | |
| NH3 | 0.07±0.02 | 0.11±0.03 |
| Season | x2 = 6.1799, p = 0.012 * | |
| Sites | x2 = 0.18107, p = 0.98 | |
| NO3 | 7.4±1.0 | 6.7±0.6 |
| Season | x2 = 2.6694, p = 0.10 | |
| Sites | x2 = 0.13889, p = 0.98 | |
| NH4 | 0.06±0.02 | 0.09±0.05 |
| Season | x2 = 2.2443, p = 0.13 | |
| Sites | x2 = 0.0171, p = 0.99 | |
| Fish Species | Fish/R | AcIn/R | Chi | Hya | Cam | Ost | Bos | Cop | Detritus |
|---|---|---|---|---|---|---|---|---|---|
| Goodeidae | |||||||||
| * Alloophorus robustus | 7.1 | 1.2 | 1.9 | 13.0 | 0.3 | 0 | |||
| ** Allotoca zacapuensis | 13.8 | 4.8 | 22.9 | 0.6 | 0.8 | 0 | |||
| * Goodea atripinnis | 4.5 | 1.5 | 6.9 | 7.5 | 2.5 | 9.7 | |||
| * Hubbsina turneri | 3.9 | 2.7 | 31.2 | 2.3 | 4.5 | 3.9 | 0 | ||
| * Skiffia lermae | 0.7 | 4.6 | 0.9 | 0.4 | 6.0 | 1.6 | 5.1 | ||
| * Xenotoca variata | 8.1 | 8.2 | 27.4 | 9.3 | 4.9 | 0.3 | |||
| * Zoogoneticus quitzeoensis | 7.6 | 2.2 | 3.9 | 13.7 | 0.4 | 10.9 | 3.6 | ||
| Atherinopsidae | |||||||||
| * Chirostoma humboldtianum | 1.5 | 0.5 | 0.5 | 27.1 | 0.4 | 2.2 | 31.0 | 5.2 | 0 |
| Poeciliidae | |||||||||
| * Poeciliopsis infans | 3.1 | 0.2 | 0.2 | 0.6 | 0.9 | 24.6 | 6.1 | ||
| Cyprinidae | |||||||||
| + Ctenopharyngodon idella | 21.3 | ||||||||
| ** Notropis grandis | 20.3 | 13.3 | 9.6 | 1.9 | 0 | ||||
| Fish Species | Gast | Biv | Ker | Frag | Dia | Ve/R | Pot | Plu | |
| Goodeidae | |||||||||
| * Alloophorus robustus | 5.7 | 1.2 | |||||||
| ** Allotoca zacapuensis | 0.4 | ||||||||
| * Goodea atripinnis | 1.0 | 0.4 | 3.0 | 4.4 | 13.9 | ||||
| * Hubbsina turneri | 3.6 | 0.3 | 0.9 | ||||||
| * Skiffia lermae | 0.2 | 14.8 | 1.1 | 0.6 | |||||
| * Xenotoca variata | 0.6 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | |||
| * Zoogoneticus quitzeoensis | 0.4 | ||||||||
| Atherinopsidae | |||||||||
| * Chirostoma humboldtianum | 0.9 | 0.9 | 2.6 | 0.2 | |||||
| Poeciliidae | |||||||||
| * Poeciliopsis infans | 1.8 | ||||||||
| Cyprinidae | |||||||||
| + Ctenopharyngodon idella | 48.2 | 13.2 | |||||||
| ** Notropis grandis | 6.8 | 0.6 | 0.1 | ||||||
| Fish Species | Trophic Level | Omnivory Index |
|---|---|---|
| A. robustus | 3.5 ± 0.52 | 0.20 |
| A. zacapuensis | 3.3 ± 0.45 | 0.10 |
| G. atripinnis | 2.8 ± 0.37 | 0.29 |
| H. turneri | 3.1 ± 0.40 | 0.12 |
| S. lermae | 2.8 ± 0.35 | 0.22 |
| X. variata | 2.8 ± 0.37 | 0.10 |
| Z. quitzeoensis | 3.0 ± 0.39 | 0.12 |
| C. humboldtianum | 3.3 ± 0.44 | 0.25 |
| P. infans | 3.1 ± 0.46 | 0.05 |
| C. idella | 2.6 ± 0.14 | 0.01 |
| N. grandis | 2.9 ± 0.38 | 0.15 |
| Ar | Az | Ht | Sl | Xv | Zq | Ch | Pi | Ci | Ng | Ga | |
| Ar | 1 | 0.33 | 0.35 | 0.16 | 0.40 | 0.38 | 0.30 | 0.19 | 0.00 | 0.29 | 0.15 |
| Az | 1 | 0.87 | 0.37 | 0.83 | 0.72 | 0.54 | 0.13 | 0.00 | 0.52 | 0.20 | |
| Ht | 1 | 0.47 | 0.88 | 0.79 | 0.79 | 0.33 | 0.00 | 0.72 | 0.38 | ||
| Sl | 1 | 0.45 | 0.52 | 0.49 | 0.51 | 0.66 | 0.50 | 0.76 | |||
| Xv | 1 | 0.93 | 0.67 | 0.24 | 0.03 | 0.72 | 0.36 | ||||
| Zq | 1 | 0.63 | 0.33 | 0.07 | 0.66 | 0.45 | |||||
| Ch | 1 | 0.68 | 0.01 | 0.80 | 0.50 | ||||||
| Pi | 1 | 0.10 | 0.45 | 0.57 | |||||||
| Ci | 1 | 0.04 | 0.38 | ||||||||
| Ng | 1 | 0.52 | |||||||||
| Ga | 1 |
| NNR | CR | TA | SEA | SEAc | |
|---|---|---|---|---|---|
| Wet Season | 11.3 | 10.7 | 82.4 | ||
| Fish | 49.5 | 9.1 | 9.2 | ||
| Phyto | 1.7 | 2.3 | 3.5 | ||
| BMI | 21.5 | 4.8 | 4.9 | ||
| Zoo | 4.3 | 5.3 | 7.9 | ||
| Dry Season | 10.8 | 12.7 | 88.3 | ||
| Fish | 49.5 | 9.1 | 9.2 | ||
| Phyto | 1.7 | 2.3 | 3.5 | ||
| BMI | 21.5 | 4.8 | 4.9 | ||
| Zoo | 4.3 | 5.3 | 7.9 | ||
| Site 1 | 10.7 | 8.9 | 64.0 | ||
| Fish | 38.2 | 9.7 | 9.8 | ||
| BMI | 15.1 | 5.7 | 6.1 | ||
| Phyto-Zoo | 3.0 | 3.6 | 5.4 | ||
| Site 2 | 9.1 | 12.6 | 74.8 | ||
| Fish | 39.2 | 9.6 | 9.7 | ||
| BMI | 10.7 | 4.8 | 5.2 | ||
| Phyto-Zoo | 8.6 | 10.5 | 15.7 | ||
| Site 3 | 10.6 | 9.8 | 63.6 | ||
| Fish | 34.2 | 7.5 | 7.5 | ||
| BMI | 10.6 | 5.2 | 5.8 | ||
| Phyto-Zoo | 5.1 | 6.3 | 9.5 | ||
| Site 4 | 10.3 | 9 | 68.19 | ||
| Fish | 32.7 | 9.0 | 9.2 | ||
| BMI | 22.8 | 9.6 | 10.5 | ||
| Phyto-Zoo | 1.8 | 2.2 | 3.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-García, A.; Jeppesen, E.; Moncayo-Estrada, R.; Mercado-Silva, N.; Domínguez-Domínguez, O. Diet and Trophic Structure of the Fish Community in a Small Sub-Tropical Lake in Central Mexico. Water 2023, 15, 1301. https://doi.org/10.3390/w15071301
Ramírez-García A, Jeppesen E, Moncayo-Estrada R, Mercado-Silva N, Domínguez-Domínguez O. Diet and Trophic Structure of the Fish Community in a Small Sub-Tropical Lake in Central Mexico. Water. 2023; 15(7):1301. https://doi.org/10.3390/w15071301
Chicago/Turabian StyleRamírez-García, Arely, Erik Jeppesen, Rodrigo Moncayo-Estrada, Norman Mercado-Silva, and Omar Domínguez-Domínguez. 2023. "Diet and Trophic Structure of the Fish Community in a Small Sub-Tropical Lake in Central Mexico" Water 15, no. 7: 1301. https://doi.org/10.3390/w15071301
APA StyleRamírez-García, A., Jeppesen, E., Moncayo-Estrada, R., Mercado-Silva, N., & Domínguez-Domínguez, O. (2023). Diet and Trophic Structure of the Fish Community in a Small Sub-Tropical Lake in Central Mexico. Water, 15(7), 1301. https://doi.org/10.3390/w15071301