Abstract
The rapid reduction in coral reefs worldwide has led to increasing attention toward protecting and restoring coral reef ecosystems. Coral reefs not only have a rich diversity of coral species, but they can also provide important products and services for human beings. One type of coral, Echinogorgia, has important scientific research value and application prospects. To understand the diversity of coral species, diving surveys were conducted in Xiamen Bay in 2017 and 2021, and a total of 928 samples were collected. Taxonomic research was conducted using methods such as morphological identification through electron microscopy. Specific phylogenetic trees of the COI gene, mtMuts gene, and ITS1 gene were analyzed. There were 47 specimens of Echinogorgia coral included among 928 samples. Fifteen species of Echinogorgia were identified, including Echinogorgia ramosa, Echinogorgia flexilis, Echinogorgia russelli, Echinogorgia ramulosa, and Echinogorgia gracilima (which represent the newly recorded species in the waters of China). This study increases the species diversity records in China and contributes to new geographical distribution information of Echinogorgia worldwide. The primary data also serve as the baseline data for long-term biomonitoring programs to estimate the status of octocorals in Xiamen Bay.
1. Introduction
Coral reefs, often referred to as an “oasis in tropical ocean deserts” or “tropical rain forest in the ocean”, are known for their ability to provide excellent habitats for numerous organisms; in addition, they form unique and highly productive ecosystems [1]. According to the statistics, coral reefs are distributed in more than 100 countries and territories. Even though they cover less than 0.2% of the seafloor, they support multiple functions, including marine species diversity, coastal protection, food, and economic value [2]. Coral reef ecosystems are estimated to provide 2.85% of the value and services of marine ecosystems [3] and they are considered to have made an inestimable contribution to the world. Since the discovery of recorded corals, the investigation and classification of coral resources have been one of the research focuses of scholars.
Corals are a global biological resource. In terms of distribution, the Indo–Pacific region, including the Pacific Ocean, Southeast Asia Sea, Indian Sea, and Red Sea, accounts for 92% of the world’s coral reef area [4]. According to a certain report [5], the second most populous place for coral reefs in Southeast Asia is the South China Sea, with an estimated area of 37,935 square kilometers, which accounts for about 5%. In China, coral reefs also appear in certain sea areas, such as the Fujian, Guangdong, Guangxi, and Hainan coasts [4]. Zou [4] believes that the hermatypic corals community in Dongshan, Fujian is the northernmost sea area where hermatypic corals appear in China.
In China, a number of coral species have been recorded, including 80 genera, or subgenera, and more than 700 species. A total of 1422 species of cnidarians are included in the “List of Marine Biota of the China Sea” [6]. Among them, there are 763 species of coral, including 41 species of Cerriantipatharia of Anthozoa, 328 species of Octocorollia, and 394 species of Seleactinia of Hexacorollia. As of 2020, China’s corals have been recorded in two categories, as well as in sixteen families, seventy-seven genera, and four hundred forty-five species [7]. In the waters of Fujian, there are fifty-four species of gorgonians in twenty-two genera and nine families [8]. The places where Scleractinian corals are distributed in Fujian include Xiamen, Dongshan, and Niushan Island, as well as in the Taishan Islands and other such islands [8,9]. However, there are currently few studies on the coral classification in Xiamen Bay. In 2006, a total of 38 species of coral were recorded in Xiamen Bay. Among them, there were twenty-eight species in seven families of Alcyonacea, and ten species in six families of Scleractinia [10]. In 2007, Huang et al. [11] reported on the species of Octocorallia corals in the waters near Wuyu Island, and these belonged to four families and seven genera. In 2017, Ni et al. [12] found six species of scleractinian corals in Baiha Reef, Xiamen Bay. In 2022, Liu et al. [13] reported three species of Astrogorgia coral inhabiting Xiamen Bay. In 2023, Yang et al. [14] found a newly recorded species in Xiamen Bay.
The functions of coral reefs support marine biodiversity and provide important products and services to humans [15,16]. Octocorallia includes blue corals, soft corals, sea pens, and gorgonians (sea fans and sea whips) within three orders: Alcyonacea, Helioporacea, and Pennatulacea [17]. There are two orders and seventy-nine families in Octocorallia [18]. Octocorallia coral plays an important role in maintaining the structural integrity and biodiversity of marine ecosystems [19]. The genus Echinogorgia belongs to the Malacalcyonacea of Octocorallia, which is similar to the Paramuriceidae. Echinogorgia was established in 1865, and the type species are E. sasappo (Esper, 1794), E. reticulata (Esper, 1791), and E. umbratica (Esper, 1791) [20]. This genus has the following characteristics: the colonies surface is flat, it is in a network-shaped fan shape, and it has short lateral branches on the main branch [20]. Moreover, the sclerites are spindle-shaped and it has leaf-shaped sclerites in the coenenchymata. These corals are red, brown, yellow, or white. As of June 2023, WORMS (https://www.marinespecies.org, accessed on 2 March 2023) records 42 valid species of this genus. The species of this genus occur in many sea areas, including the offshore waters of mainland China, Hong Kong, Germany, and the Indian Ocean [21]. Among them, eight species of this genus have been observed in the waters of China, namely E. pseudosassapo, E. ridley [22], E. sassapo, E. flora, E. furfuracea, E. complexa, E. aurantiaca, and E. lami [8,11,23]. Among them, E. ridley is recorded in the offshore waters of Taiwan, E. pseudosassapo is recorded in the offshore waters of Taiwan and Fujian, while the other six species are mainly distributed in the offshore waters of Fujian.
At present, the following two issues are of concern to the genus Echinogorgia: one is taxonomic identification and the other is natural active substances. The issue of natural active substances includes the extraction of substances [24,25], as well as the structure and biological activity of substances [26,27,28]. Most of the above studies show the status of unspecified species [29,30], and this uncertainty is not conducive to future research and production applications. Uncertainty in species classification is mainly due to the fact that previous authors did not assign the holotype and that the specimens and sclerites are mostly insufficient in correctly identifying the species. Furthermore, certain species have been identified based on only a few specimens or fragments [31]. Therefore, an in-depth taxonomic study of the species in this genus is crucial.
With the rapid development of molecular technology, these tools are also widely used in coral classification and identification. Certain studies have shown that 18S rDNA [32,33], ITS [34,35], COI, ND2, and mtMuts gene fragments [36,37,38] are suitable for the phylogenetic study of corals. On the basis of morphological identification, Xu et al. [39] established a phylogenetic tree of the mtMuts gene to study the taxonomy and phylogeny of Octocorallia in western Pacific seamounts. Li et al. [33] clearly demonstrated the phylogenetic relationship of Gorgonian coral by establishing a 18S rDNA phylogenetic tree. McFadden et al. [18] used the mtMuts gene to establish a phylogenetic tree and conducted a phylogenetic analysis on 185 taxa of Octocorallia coral. Hume et al. [40] studied coral species diversity using ITS2 markers.
In this study, the morphological classification method was used to identify the samples of Echinogorgia collected from Xiamen Bay. Also, via molecular techniques, phylogenetic analysis was conducted using the COI gene sequences, mtMuts gene sequences, and ITS gene sequences. The determination of the specific types of corals in Xiamen Bay will help with providing data support for biodiversity conservation in Xiamen Bay.
2. Materials and Methods
2.1. Station and Sample Collection
Nine hundred and twenty-eight coral samples were collected via diving sampling in 2017 and 2021. Among them, 823 samples were collected in 10 stations in 2017. In 2021, 105 samples were collected from 4 stations. The information collected from the sampling stations is shown in Table 1, and the schematic diagram of the sampling stations is shown in Figure 1.

Table 1.
The sampling station information from 2017 and 2021.

Figure 1.
Schematic diagram of sampling stations.
2.2. Material and Sample Processing
The samples were collected and brought back to the laboratory in order to take photos of the living corals, and these were numbered for preservation. Suitable lengths of coral samples were cut and placed in centrifuge tubes filled with 95% ethanol for traditional morphological and molecular biology experiments. The remaining samples were stored in an ultralow temperature refrigerator at −70 °C.
2.3. Experimental Instruments
In this study, certain experimental instruments were used, including a Nikon SMZ1270 dissecting microscope, a NiKon ECLIPSE 80i microscope, a DHG-9050A air-drying oven, a G5 PhenomProX2017 electron scanning electron microscope, an A200 PCR amplification apparatus, a Z216MK centrifuge, an MK200-2 dry thermostat, a DYY-8C electrophoresis apparatus, and a JS-20.2 gel imaging system.
2.4. Identification Method
Generally, octocorals are identified, classified, and described primarily on the basis of external and internal morphology. These morphological characteristics include the colonies’ size, shape, color, and calyx structures, as well as their sclerite content, dominance, shape, size, and arrangement [31,32,37,41].
(1) Exterior feature observation
First, certain characteristics were observed, including the coral appearances, colors, polyps, surfaces, spindles, branches, bases, top colonies, etc. In addition, actions were carried out, including measuring, photographing, describing, and recording the relevant parts of the coral.
(2) Observation of local appearance and morphology
A dissecting microscope (Nikon SMZ1270) was used to observe (as well as photograph) the color and shape of the polyps, as well as the calyx structure.
(3) Observation of sclerites
After the tissue samples were dissolved in sodium hypochlorite, different parts of the sclerite samples (polyps, coenenchymata, and tentacles) were observed and examined. The sclerites were washed with distilled water and subsequently dried using a blast dryer. The samples were sent to the scanning electron microscope (SEM) (G5 PhenomProX2017) set up in the National Key Laboratory of Xiamen University for observation. Then, detailed images of the sclerites through SEM were obtained. The images for each sclerite type were then optimized using Adobe Photoshop 2020. When the coral branches were placed in the Petri dish, a solution of ultra-pure water and sodium hypochlorite (with a ratio of 3:1–5:1) was injected into it for dissolution. Next, the arrangement of the spicules under a dissecting microscope were observed and pictures were taken.
2.5. Classification Basis
2.5.1. Identification Documents
Certain important documents were used for species classification and identification such as “Soft Corals and Sea Fans” [42], the World Register of Marine Species (WROMS) [21], and taxonomic terminology references [43].
2.5.2. DNA Extraction and PCR Amplification
With the use of the marine animal tissue genomic DNA extraction kit of Shanghai Biotechnology Co., Ltd., three coral individuals were prepared for total genome DNA extraction. The steps of extraction were followed as per the manufacturer’s instructions. The primers used in this study are shown in Table 2.
Table 2.
The primers used for genetic analyses.
2.5.3. Phylogenetic Analysis of Coral Samples Based on COI, mtMuts, and ITS Gene Fragments
Based on the morphological results, using molecular technology to extract the DNA from certain coral samples for amplification analysis, the COI gene sequence, mtMuts gene sequence, and ITS1 gene sequence results of the samples were compared with the GenBank database. The gene sequence fragments of the species with a similarity of over 95% were compared with the corresponding screened gene fragment information in the NCBI database using MEGA7.0 software. The neighbor joining method, based on the Kimura 2 parameter (K2P) model with 1000 bootstrap replicates, was used to construct phylogenetic trees and to conduct a phylogenetic analysis.
3. Results
3.1. Systematic Approaches
The morphological classification method was used to identify 47 samples of the genus Echinogorgia, and the results showed that these samples were classified into 15 species. A total of 15 species named Echinogorgia ramosa, E. flexilis, E. russelli, E. ramulosa, E. gracillima, E. sp1, E. sp2, E. sp3, E. sp4, E. sp5, E. sp6, E. sp7, E. sp8, E. sp9, and E. sp10 were identified. We found five new recorded species in China, including E. ramose, E. flexilis, E. russelli, E. ramulosa, and E. gracillima.
The morphological descriptions of the five newly recorded species were as follows.
3.1.1. Echinogorgia ramosa (Thomson and Henderson, 1905)
Sample collection location: Qingyu (118°07′34″, 24°21′46″), water depth: 7.5 m;
Study sample number: 20210423-QY-33;
Coral group (Figure 2a): The living colonies were 9 cm high, and most of their branches were on the same plane. The lateral branches bent outward and upward. The irregular branches were mainly distributed on one side of the main trunk. The diameter of the central shaft was greater than 0.5 mm. The central axis was divided into two layers: inner and outer (with an uneven yellow color on the inner layer and a uniform dark brown color on the outer layer). The ratio of the inner and outer layers of the axis radius was approximately 2:3;
Polyps (Figure 2a(B,C)): The polyps could freely contract and were mainly distributed on the surface of the trunk and branches. The polyps contracted into the coral calyx, which was circular in shape and had a diameter of about 0.5 mm;
Sclerites (Figure 2b): The polyps contained straight or curved spindle-shaped sclerites with a length of about 0.2–0.4 mm. The surface of the sclerites contained small and conical columnar protrusions. The coenenchymata contained leaf-shaped sclerites and a few pan-shaped sclerites with a length of about 0.1–0.25 mm. The leaf-shaped sclerites contained serrated edges, and the surface of the sclerites presented protrusions of varying sizes. Sclerites are colorless;
Color (Figure 2a(A)): reddish brown, and dark brown in alcohol;
Sample collection location: the waters near Qingyu Island in Xiamen Bay.
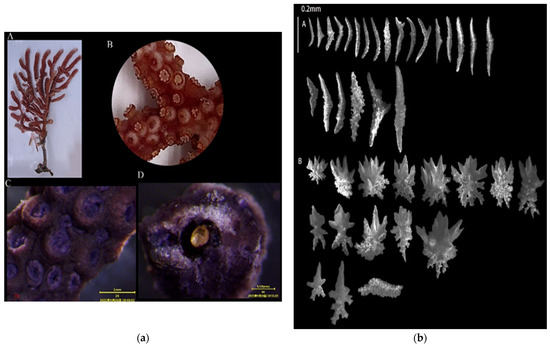
Figure 2.
E. ramosa: (a) Corals’ external morphologies ((A). Coral colonies; (B,C). Polyps; (D). Axis); (b) SEM images of the coral sclerites ((A). Sclerites of the polyps; (B). Sclerites of the coenenchymata; Scales: A = B = 0.2 mm).
3.1.2. Echinogorgia flexilis (Thomson and Simpson, 1909)
Coral group (Figure 3a): The living colonies were in a fan-shaped shape and were up to 10 cm high. The branches were irregular and on the same plane. The central axis had two layers. The inner layer was an uneven yellow, and the outer layer was a uniform dark brown. The axis radius was 3:2;
Polyps (Figure 3a(B,C)): The polyps’ monotypes were distributed on the surface of the stems and branches. The gaps between the polyps were approximately 0.5 mm in diameter. The coral calyx protruded and appeared as a dome, and the polyps contracted into the calyx. The polyps turned purple after alcohol immersion;
Sclerites (Figure 3b): Straight or slightly curved fusiform-shaped sclerites were distributed in the polyps and were about 0.18–0.3 mm long, and some of the fusiform-shaped sclerites were bifurcated at one end with columnar and prickly protrusions on the sclerites. The surface layers of the coenenchymata had leaf-shaped, flat-shaped, and irregular-shaped sclerites with a length of about 0.1–0.35 mm. The leaf-shaped sclerites had serrated edges, and the surface of the sclerites all contained warty protrusions. The inner layers of the coenenchymata contained multiple spicules (approximately 0.05–0.15 mm long), and the sclerites were colorless;
Color (Figure 3a(A)): brownish red, and dark brown in alcohol;
Sample collection location: the waters near Qingyu Island in Xiamen Bay.
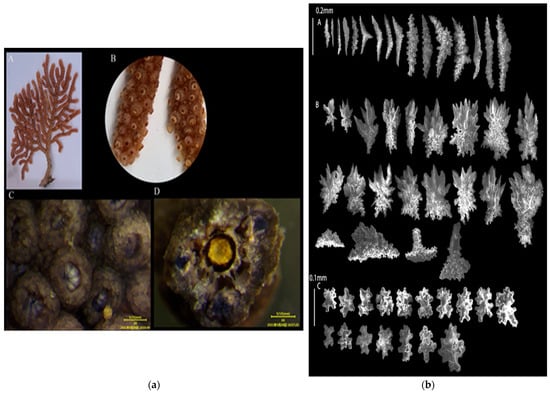
Figure 3.
E. flexilis: (a) The corals’ external morphologies ((A). Coral colonies; (B,C). Polyps; (D). Axis); (b) SEM images of the coral sclerites ((A). Sclerites of the polyps; (B). Sclerites of the surface layers of coenenchymata; (C). Sclerites of the inner layers of coenenchymata; Scales: A = B = 0.2 mm; C = 0.1 mm).
3.1.3. Echinogorgia russelli (Bayer, 1949)
Sample collection location: Wuyu (118°8′57″,24°2′51″), 5 m;
Study sample number: 20210423-WY-15;
Coral group (Figure 4a): The living colonies were 20 cm tall with dense and irregular branches that bent upward and downward. Most of the branches were at right angles with a small number of the coenenchymata shedding from the base of the main stem. The diameter of the central axis was about 1 mm and was an uneven yellow color;
Polyps (Figure 4a(A,B)): The polyps could contract completely and freely, and they were mainly distributed on the surface of the main stems and branches. The calyx slightly protruded, and it resembled a low dome-shaped wart. The polyps completely contracted into the calyx with a diameter of less than 1 mm. After alcohol immersion, the polyps turned purple;
Sclerites (Figure 4b): The polyps contained columnar- and spindle-shaped sclerites, about 0.18–0.3 mm long, with columnar and spiny processes on the sclerites. The outer layers of the coenenchymata contained leaf-shaped sclerites, approximately 0.18–0.25 mm long, with serrated edges on the sclerites. The inner layers of the coenenchymata contained quadrangular sclerites with a length of approximately 0.05–0.08 mm. The sclerites were colorless;
Color (Figure 4a(A)): light brownish red, and dark brown in alcohol;
Sample collection location: the waters near Wuyu Island in Xiamen Bay.
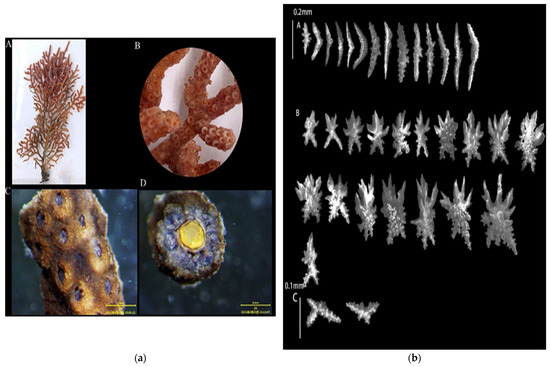
Figure 4.
E. russelli: (a) The corals’ external morphologies ((A). Coral colonies; (B,C). Polyps; (D). Axis); (b) SEM images of the coral sclerites ((A). Sclerites of the polyps; (B). Sclerites of the outer layers of coenenchymata; (C). Sclerites of the inner layers of coenenchymata; Scales: A = B = 0.2 mm; C = 0.1 mm).
3.1.4. Echinogorgia ramulosa (Gray, 1870)
Sample collection location: Wuyu (118°8′57″, 24°2′51″), 5 m;
Study sample number: 20210423-WY-15;
Coral group (Figure 5a): The living colonies were branching and were up to 14 cm high with abundant short branches. The small branches appeared at right angles to the main branch, and the short branches were slightly swollen at the end. The coenenchymata were thicker, and the diameter of the central axis was less than 0.5 mm. The central axis was uniformly yellow;
Polyps (Figure 5a(B,C)): The polyps appeared uniformly on the surface of the trunks and branches, with a diameter of about 0.5 mm. The calyx protruded, resembling a circle, and some of the polyps contracted into the calyx. After soaking in alcohol, the polyp turned purple;
Sclerites (Figure 5b): The polyps contained columnar- and spindle-shaped sclerites about 0.1–0.4 mm long. The spindle-shaped sclerites were relatively flat, and the surface of the spicules contained more verrucous protrusions. The surface layers of the coenenchymata contained leaf-shaped sclerites and irregular sclerites with a length of about 0.1–0.2 mm, and the edges of the sclerites contained serrated protrusions. The inner layers of the coenenchymata contained multiple spicules with a length of approximately 0.1–0.15 mm. The sclerites were colorless;
Color (Figure 5a(A)): brick red, and dark brown in alcohol;
Sample collection location: the waters near Wuyu Island in Xiamen Bay.
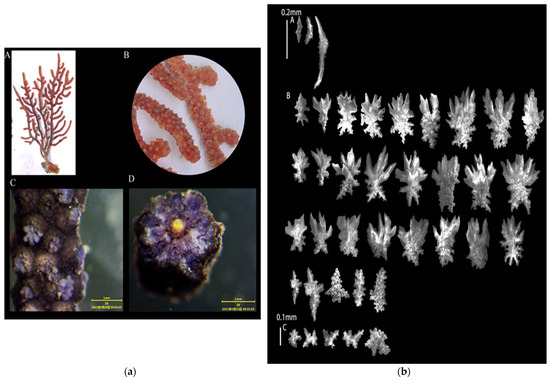
Figure 5.
E. ramulosa: (a) Corals’ external morphologies ((A). Coral colonies; (B,C). Polyps; (D). Axis); (b) SEM images of the coral sclerites ((A). Sclerites of the polyps; (B). Sclerites of the outer layers of coenenchymata; (C). Sclerites of the inner layers of coenenchymata; Scales: A = B = 0.2 mm; C = 0.1 mm).
3.1.5. Echinogorgia gracillima (Kükenthal, 1917)
Sample collection location: Wuyu (118°8′57″, 24°2′51″), 5 m;
Study sample number: 20210423-WY-11;
Coral group (Figure 6a): The living colonies were fan-shaped and were up to 10 cm with abundant and overlapping branches. There were a large number of lateral branches on one plane with slightly enlarged ends. The inner layer of the central axis was uneven and orange–yellow, and the outer layer was an uneven dark brown. The radius of the axis was about 2:1 for the inner and outer layers;
Polyps (Figure 6a(B,C)): The polyps appeared on the surface of the trunks and branches with protruding calyxes. The polyps could fully contract into the calyx with a diameter of about 0.5 mm. After alcohol immersion, the polyps turned yellow;
Sclerites (Figure 6b): The polyps contained columnar- and spindle-shaped sclerites, which were about 0.1–0.25 mm long. The sclerites contained verrucous protrusions, and some of the spindle-shaped sclerites were bifurcated at one end. The outer layers of the coenenchymata contained leaf-shaped sclerites, approximately 0.15–0.2 mm long, with serrated protrusions at the edges of the sclerites. The inner layers of the coenenchymata contained radiating sclerites, approximately 0.03–0.04 mm in length. The sclerites were colorless.
Color (Figure 6a(A)): yellow, and gray-brown in alcohol.
Sample collection location: the waters near Wuyu Island in Xiamen Bay.

Figure 6.
E. gracillima: (a) Corals’ external morphologies ((A). Coral colonies; (B,C). Polyps; (D). Axis); (b) SEM images of the coral sclerites ((A). Sclerites of the polyps; (B). Sclerites of the outer layers of coenenchymata; (C). Sclerites of the inner layers of coenenchymata; Scales: A = B = 0.2 mm; C = 0.1 mm).
3.2. DNA Analysis
The gene sequence of the successfully amplified Echinogorgia coral sample was uploaded to the NCBI database to obtain the gene sequence number, as shown in Appendix A. The gene sequences of the samples were compared with and screened using NCBI’s Nucleotide BLAST software (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome, accessed on 12 March 2023), and the corresponding gene fragment information screened in the NCBI database was used in the MEGA7.0 software. The K2P model and NJ method were used to model the phylogenetic trees of the COI gene, mtMuts gene, and ITS1 gene, respectively. The confidence level was derived from 1000 non-parametric bootstrap analyses. The specific phylogenetic trees of the COI gene, mtMuts gene, and ITS1 gene depicted the evolutionary lineages of the different species. The results of the phylogenetic trees are shown in Figure 7 (the login numbers in parentheses in the figure represent the samples of this study).
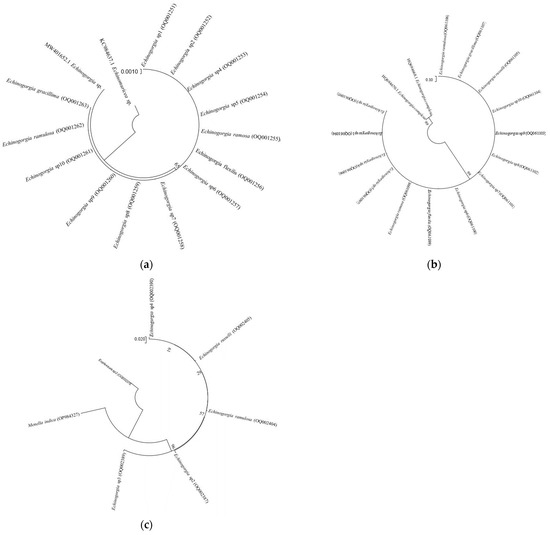
Figure 7.
NJ phylogenetic tree: (a) Based on fragments of the COI gene sequences; (b) Based on fragments of the mtMuts gene sequence; (c) Based on fragments of the ITS1 gene sequences.
Figure 7a depicts a phylogenetic tree that was established using COI gene sequences. From the figure, it can be concluded that the COI gene sequence can be used to distinguish the class of the genus Echinogorgia from the other genus. Our molecular phylogenetic analysis showed that E. sp1, E. sp2, E. sp4, E. sp5, E. ramosa, E. flexilis, E. sp6, E. sp7, E. sp8, E. sp9, E. sp10, E. ramulosa, and E. gracillima are genetically closer.
Figure 7b depicts a phylogenetic tree established using mtMuts gene sequences. From the figure, it can be concluded that the mtMuts gene sequence can be used to distinguish the class of the genus Echinogorgia from the other genus. Our molecular phylogenetic analysis showed that E. ramulosa, E. gracilima, E. russelli, E. sp10, E. sp9, E. sp8, E. sp7, E. sp6, E. flexilis, E. ramosa, E. sp5, E. sp4, E. sp1, and E. sp3 are genetically closer.
Figure 7c is a phylogenetic tree established based on the ITS1 gene sequence. From the figure, it can be concluded that the ITS gene sequence can be used to distinguish the species of the genus Echinogorgia. Our molecular phylogenetic analysis showed that E. sp4, E. russelli, E. ramulosa, E. sp2, and E. sp3 are genetically closer.
4. Discussion
4.1. Key Points for the Morphological Identification of the Genus Echinogorgia
The external characteristics are the main basis for the classification and identification of Octocorallia corals. The characteristics—such as coral specimens (color, branching, size, and the distribution of the coral), polyps (shape during relaxation and contraction), and sclerites (shape, size, and color)—were observed [22,31,32,41,46]. The living coral sample with the sample number of 20210423-QY-33 was found to be reddish-brown. Most of the branches were on the same plane. There were a few anastomoses between the branches. The branches were irregular and mainly distributed on one side of the main trunk. The polyps contained straight or curved spindle-shaped sclerites, and the coenenchymata contained leaf-shaped sclerites and a small number of pan-shaped sclerites. In the original literature of the species of Econogorgia ramosa (Thomson and Henderson, 1905), it was described that the coral body contains a large number of branches, some of which are anastomosed. In addition, the polyps contain spindle-shaped sclerites, the coenenchymata contain leaf-shaped sclerites and pan-shaped sclerites, and the color of the coral body is brick-red [47]. After comparing the morphological characteristics of the coral sample with the original literature description and image of E. ramosa (Thomson and Henderson, 1905), it was identified as E. ramosa (Thomson and Henderson, 1905).
The coral sample with the sample number 20210423-QY-27 had a fan-shaped coral body, branching coral bodies, and small gaps between the polyps. The calyx protruded and appeared in a dome-like shape. The polyps were distributed with straight or slightly spindle-shaped sclerites. The surface of the coenenchymata contained leaf-shaped, pan-shaped, and irregular-shaped sclerites. The leaf-shaped sclerites contained serrated edges, and the surface of the sclerites contained warty projections. The inner layer of the coenenchymata contained multiple sclerites. The original literature on the species described that the coral body is fan-shaped, the coral body is branched, the calyx is prominent, the polyps are distributed with spindle-shaped sclerites, and the symphysis contains multiple sclerites in Econogorgia flexilis (Thomson and Simpson, 1909) [48]. The morphological characteristics of the coral body with the sample number 20210423-QY-27 corresponded to the original literature, and it was identified as the species of E. flexilis (Thomson and Simpson, 1909).
The sample number was found to be 20210423-WY-15. The living colonies exhibited dense branching with anastomoses between the branches. The branches were irregular and grew upward and downward. Most of the branches were at right angles. The polyps contained columnar- and spindle-shaped sclerites, and the outer layer of the common flesh contained leaf-shaped bone needles. The coenenchymata in the inner layers contained quadrangular sclerites. The morphological description of its coral body, branches, and the sclerites of polyps corresponded to that described in the original literature of Econogorgia ruselli (Bayer, 1949). Its coral body was found to be upright, and the coral body had many branches with many anastomoses between the branches and curved branches; in addition, many of the branches were at right angles. The polyps contained spindle-shaped sclerites, and the coenenchymata contained multi-shoot-shaped sclerites [47]. Therefore, the sample with sample number 20210423-WY-15 was identified as E. russelli (Bayer, 1949).
The sample with the sample number 20210423-WY-6 was found to have a branching coral body with abundant short branches. The small branches were at right angles to the main branch, and the end of the short branch was slightly enlarged. The calyx protruded, resembling a circle, and the polyps contracted into the calyx. The polyps contained columnar- and spindle-shaped sclerites that were about 0.1–0.4 mm long. They also had flat spindle-shaped sclerites, and the surface of the sclerites contained more verrucous protrusions. The surface layers of the coenenchymata contained leaf-shaped and irregular-shaped sclerites with a length of approximately 0.1–0.2 mm. The inner layers of the coenenchymata contained multiple spicules with a length of approximately 0.1–0.15 mm. In the original literature of Echinogorgia ramulosa (Gray, 1870), the coral was described as having a branching shape, rich branches, as well as especially short and small branches with right angles between the branches and ends. The polyps are distributed on the surface of twigs. The calyx protrudes. The polyps contain spindle-shaped sclerites with a length of 0.08–0.3 mm. The surface of the sclerites contains many warty projections [49]. By comparing the morphological description of sample number 20210423-WY-6 with the description of the original literature on E. ramulosa (Gray, 1870) [50], it was identified as E. ramulosa (Gray, 1870) [50].
Sample number 20210423-WY-11 showed a fan-shaped coral body with a height of 10 cm and had abundant and overlapping branches. There were a large number of lateral branches on one plane, and the ends of the branches were slightly enlarged. The polyps had a diameter of about 0.5 mm and protruding coral calyxes. The polyps contained columnar- and spindle-shaped sclerites, which were about 0.1–0.25 mm long. The sclerites contained verrucous protrusions, and some spindle-shaped sclerites were bifurcated at one end. The leaf-shaped sclerites were shown on the outer layers of the coenenchymata, while the inner layers of the coenenchymata contained radiating sclerites, which were colorless. The coral living organisms were yellow. In the original literature description of Echinogorgia gracilima (Kükenthal, 1917), the coral bodies had numerous lateral branches on a plane. The branch parts overlapped. The polyps were small. The distance between the polyps was about 1 mm, and the diameter was about 0.5 mm. The sclerites of the coral polyps were spindle-shaped with a length of 0.18 mm. The sclerites contained warty protrusions, and the coenenchymata contained leaf-shaped sclerites [51]. The morphological description of the sample with sample number 20210423-WY-11 corresponded to the original literature description of E. gracilima (Kükenthal, 1917); as such, it is identified as E. gracilima (Kükenthal, 1917).
As of 2 May 2023, WORMS (https://www.marinespecies.org, accessed on 2 March 2023) has recorded 42 valid species of this genus. After collecting and organizing the relevant original literature on forty-two species, it was found that the original literature on seven of the species could not be collected, and the original literature on nineteen of the species did not attach a species comparison map. In addition, the original literature describing the species of the genus Echinogorgia also had certain limitations such as simplistic morphological descriptions, a lack of corresponding example maps, and incomplete literature [20,52,53]. This made it difficult for certain samples to be identified as certain species through their morphology alone. Therefore, there are still 10 undetermined species in this study.
In addition to the corals of this genus, the identification of other genera was also the same, let alone for those at the species level. Koido et al. [54] mentioned that species identification is difficult due to the high morphological variability and plasticity of the Xeniidae family. Regarding the classification of Chrysogorgia, Untiedt et al. [55] believed that morphological information is still needed for taxonomic revisions. In particular, species identification relies on the observation of subtle features that are not easily accessible to non-experts. Furthermore, phenomena such as homogeneous traits, subtle differences, and environmental changing forces may blur the boundaries of coral species [56,57,58].
The descriptions of many octocorals still rely on morphological evidence, and an integrated approach is clearly needed but not yet prevalent [59]. Untiedt et al. [55] mentioned that an integrated approach assessing morphological and molecular variation is needed to address classification issues. Information integration is often considered the key to alleviating octocoral taxonomic problems [60,61,62]. The integration of genetic and morphological data is often considered key to mitigating taxonomic issues in octocorals [60,61,62]. As early as 2001, the scholar Bayer mentioned that new technologies in molecular analysis will allow for the possibility of solving the concept of species and genus, especially in problems that change with long-distance changes in geographical and ecological conditions [63].
4.2. Phylogenetic Analysis of the Genus Echinogorgia
Various molecular markers, such as mtMuts, ND2, and 28S rDNA have been used to differentiate octocoral species [64,65,66,67]. Few molecular phylogenetic analyses of xeniids have been performed at the species level. Therefore, COI, mtMuts, ND2, and 28S rDNA were used to compare some of the genera of xeniids. However, there are still some problems and limitations [64]. Furthermore, molecular techniques require large amounts of high-quality DNA, and the need to extract it from recently collected material is critical [68,69].
Given the challenges faced in the morphological identification of certain genera of coral species in Octocorallia, many scholars have conducted phylogenetic analysis on coral species to further clarify their species relationships and to help confirm their species status. Quattrini et al. [70] established a phylogenetic tree based on mtMuts and 28S rRNA gene sequence fragments for the purposes of phylogenetic analysis, but the results were inconsistent with the morphological results. Arrigoni et al. [71] used COI gene fragments for molecular markers to study the molecular biology of coral species in the Indian Ocean; however, these also differed from the morphological results. Through the specific amplification and sequencing of COI genes, Li et al. [34] compared and analyzed the sequence of COI gene fragments of eight species of corals in Xuwen Island. They clearly pointed out that the traditional morphological classification results differed slightly from the results of their molecular phylogenetic analysis [34]. Therefore, the classification and identification of the corals required a combination of morphology descriptions and molecular biology [18,72].
(1) In this study, the phylogenetic analysis of the genus Echinogorgia, the COI gene sequence fragments, and the mtMuts gene sequence fragments could only be identified as a genus class, while the ITS gene sequence fragments could be identified as a species class. Nehoray et al. [73] mentioned that phylogenetic analysis of corals in the Eilat Sea area of the Red Sea using COI gene. That analysis identified 14 families, 39 genera, and 94 species. In the paper of Li et al. [74], the phylogenetic relationships of eight species of scleractinian corals were analyzed using mitochondrial COI genes. The results showed that the COI gene sequence fragments could identify the species class of the scleractinian corals, but there were certain differences in terms of morphological identifications. Zhou et al. [75] selected the mitochondrial msh1 gene and COI gene sequences for species identification when identifying the Alcyonacea of Sarcophyton in the sea area near Hainan Island. The results showed that, of the twenty-six samples collected, five species were identified. Xiao et al. [76] used ITS gene fragments to study the evolutionary relationship of the scleractinian coral systems in the outer Lingding Island, and the results showed that the ITS gene fragment could help with identifying the species class. Xie et al. [35] used the ITS gene to analyze the phylogenetic relationship of scleractinian corals in the waters of the Dapeng Peninsula. The results showed that ITS could help with identifying the species class, but there was a significant difference found in comparison with the morphological characteristics. According to current research, ITS gene fragments are mainly used for the phylogenetic analysis of coral. In this study, ITS gene fragments were used for a systematic analysis of the coral collected from Xiamen Bay, and the results showed that the ITS gene fragment could help with identifying the species class. Further research and analysis are required to test the application of ITS gene fragments in the coral species.
(2) For a phylogenetic analysis of corals of different genera, it is necessary to identify the characteristic gene fragments of that genus. With the rapid development of molecular biology, the use of DNA barcoding technology has become an important method of genetic differentiation among closely related species [77,78]. However, at present, DNA barcoding is not applicable to the identification of all coral species. In this study, only ITS gene fragments were applicable to the differentiation of the species of the genus Echinogorgia, while COI gene sequence fragments and mtMuts gene sequence fragments are applicable to the differentiation of Echinogorgia. The selection of DNA barcoding is particularly important for coral species identification.
(3) Global coral researchers are needed to jointly improve coral molecular information in the NCBI database. As of February 2023, in the NCBI database, there are thirty instances of molecular sequence data regarding the genus Echinogorgia, including five COI and eight mtMuts gene sequences. Most of them are the undetermined species of the genus Echinogorgia’s gene. In this paper, the species information of the genus Echinogorgia are compared and screened in the NCBI database, based on the COI gene sequence fragments. The mtMuts gene sequence fragment was used to establish a phylogenetic tree for the phylogenetic analysis of the species of the genus Echinogorgia, which was then validated through morphological results. Further research is needed to enrich the molecular data information of the species in the genus Echinogorgia.
Recently, new techniques such as the target capture enrichment of ultra-conserved elements are often considered and used [79,80,81,82,83,84]. This technology has advantages over traditional sequencing methods such as RAD-seq [55]. Erickson et al. [83] believed that new approaches to coral species delineation are needed to overcome certain challenges. Boundaries and group structure within species of two octocoral genera (Alcyium and Sinularia) were tested by using the method described above. Quattrini et al. [84] advised that target enrichment methods have shown effectiveness in resolving relatively old phylogenetic relationships. In the future, the use of target enrichment methods in terms of phylogenetic relationships may be considered and applied.
4.3. Distribution of Species in the Genus Echinogorgia
The species of the Echinogorgia genus are mainly distributed in Hong Kong, Germany, the Indian Ocean, and other such places [21]. According to certain reports, seven species of this genus have been found, including E. pseudosassapo, E. ridley [22], E. sassapo, E. furfuracea, E. complexa, E. aurantiaca, and E. lami in the offshore waters of China [8,11,23]. Among them, E. pseudosasapo and E. ridley inhabit the offshore waters of Taiwan, while the rest inhabit the shallow waters along the coast of Fujian. Moreover, E. complexa, E. pseudosasapo, and E. lami are also distributed in the waters of Hong Kong [85]. E. pseudosasapo is distributed in the offshore waters of Hong Kong, Taiwan and in the shallow waters of Fujian. The results show that most of the discovered species of Echinogorgia in China’s offshore waters are distributed in the shallow waters off the coast of Fujian. However, the answer as to whether other coastal waters in China are inhabited by Echinogorgia species remains to be determined.
4.4. Management Recommendations
The global decline in coral cover has resulted in serious consequences, thereby requiring a reflection and evaluation of current management measures [86]. The health and abundance of coral cover are declining for many reasons, including overfishing, climate change, runoff pollution, plastic pollution, coastal construction, and dynamite fishing [87]. Jones et al. [88] showed that fish biodiversity is also threatened whenever permanent reef degradation occurs. From single corals (e.g., genetics, reproduction, and physiology) to coral populations, reef communities, and ecosystems, a variety of coral reef restoration approaches are currently being pursued [89]. Species identification and verification are, therefore, necessary for coral ecosystem management. This fundamental task in species identification in corals has not yet been fully scientifically described. In this study, the newly recorded species of Echinogorgia were revealed to have contributed to enriching the diversity of coral species in China. However, this type of meaningful information still requires attention to successfully complete complicated tasks in the future. Clearly, the work on coral population management techniques—which include the parameters of population status, as well as population dynamics and assessment—is rather limited.
Hein et al. [89] referred to the National Academies of Sciences, Engineering, and Medicine (NASEM), as well as the Reef Restoration and Adaptation Program (RRAP), as examples of some of the interventions that could improve the physiological resilience of coral to climate change [90,91]. Eleven conservation zones, with corals as the main protection targets, have been established and implemented in nature reserves in China [92]. One of them is located in the Dongshan Coral Nature Reserve in Fujian [93]. Effective conservation measures have been demonstrated in the establishment of these protected areas. Therefore, it appears advisable to establish similar coral protection areas in coral distribution and concentration areas in Xiamen Bay. According to the World Resources Institute [94] report, an estimated 2679 coral reef areas worldwide have marine protected areas (MPAs), which account for about 27% of all reef areas. Fernandez et al. [95] identified certain key success factors in protected area measures, including communication on the issues to be solved, organizing independent experts, extensive and participatory consultations, obtaining high-level support, and solving fishermen problems. To address the need for coral conservation, MPA managers need to understand why MPAs were created, the state and status of their resources, the forces that are impacting their future the most, as well as the essential management actions, equipment, people, and facilities required. Once MPAs have been established, it is important to determine how to promote and adapt the relevant management efforts [96]. Therefore, after the establishment of coral protection areas, the protection of coral species diversity in Xiamen Bay will need be implemented, including the establishment of a network of protection areas, strengthening the fishing management of fishermen, reducing land-based pollutants, and eliminating illegal coral fishing.
5. Conclusions
Xiamen Bay is a sea area with typical ecological characteristics and extremely high biodiversity. Understanding the diversity of coral species is crucial. This study has added the identification of 15 new species of the genus Echinogorgia, including E. ramosa, E. flexilis, E. russelli, E. ramulosa, and E. gracilima, which are new recorded species in Xiamen Bay and also in China. The morphological description and molecular analysis of these five species can also provide reference for other scholars to study the species of this genus. The results have also enriched the coral record. We believe that, based on the existing research results, government authorities should direct their attention and should design specific measures for coral protection as soon as possible in order to protect precious coral ecologies.
Author Contributions
Conceptualization, Y.-P.W. and J.-Y.L.; methodology, Y.-P.W. and J.-Y.L.; software, Y.-P.W., J.-Y.L., J.Y. and T.-J.C.; validation, Y.-P.W. and J.-Y.L.; investigation, J.-Y.L., Y.-P.W. and J.Y.; resources, J.-Y.L.; data curation, Y.-P.W. and J.-Y.L.; writing—original draft preparation, J.-Y.L. and Y.-P.W.; writing—review and editing, T.-J.C. and J.-Y.L.; visualization, Y.-P.W. and J.-Y.L.; supervision, J.-Y.L.; project administration, J.-Y.L.; funding acquisition, J.-Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Fujian Province (No. 2019J01690), Xiamen Ocean and Fishery Bureau (Southern Center Project) (No. 13GQT001NF14), and National Foundation Incubation Program of Jimei University (No. ZP2020021).
Data Availability Statement
Not applicable.
Acknowledgments
We thank Shi Yi-Jia for her suggestions and contributions to the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
The GenBank accession numbers and reference species involved in this study.
Table A1.
The GenBank accession numbers and reference species involved in this study.
| Number | Species | GenBank Accession Number | Gene Segment |
|---|---|---|---|
| 1 | Echinogorgia sp1 | OQ001251 | COI |
| 2 | Echinogorgia sp2 | OQ001252 | |
| 3 | Echinogorgia sp4 | OQ001253 | |
| 4 | Echinogorgia sp5 | OQ001254 | |
| 5 | Echinogorgia ramosa | OQ001255 | |
| 6 | Echinogorgia flexilis | OQ001256 | |
| 7 | Echinogorgia sp6 | OQ001257 | |
| 8 | Echinogorgia sp7 | OQ001258 | |
| 9 | Echinogorgia sp8 | OQ001259 | |
| 10 | Echinogorgia sp9 | OQ001260 | |
| 11 | Echinogorgia sp10 | OQ001261 | |
| 12 | Echinogorgia ramulosa | OQ001262 | |
| 13 | Echinogorgia gracillima | OQ001263 | |
| 14 | Echinogorgia sp. | MW401652.1 | |
| 15 | Echinomuricea sp. | KC984637.1 | |
| 16 | Echinogorgia sp1 | OQ061094 | mtMuts |
| 17 | Echinogorgia sp3 | OQ061095 | |
| 18 | Echinogorgia sp4 | OQ061096 | |
| 19 | Echinogorgia sp5 | OQ061097 | |
| 20 | Echinogorgia ramosa | OQ061098 | |
| 21 | Echinogorgia flexilis | OQ061099 | |
| 22 | Echinogorgia sp6 | OQ061100 | |
| 23 | Echinogorgia sp7 | OQ061101 | |
| 24 | Echinogorgia sp8 | OQ061102 | |
| 25 | Echinogorgia sp9 | OQ061103 | |
| 26 | Echinogorgia sp10 | OQ061104 | |
| 27 | Echinogorgia russelli | OQ061105 | |
| 28 | Echinogorgia ramulosa | OQ061106 | |
| 29 | Echinogorgia complexa | HQ694670.1 | |
| 30 | Echinogorgia complexa | HQ694668.1 | |
| 31 | Echinogorgia sp1 | OQ002388 | ITS1 |
| 32 | Echinogorgia sp2 | OQ002387 | |
| 33 | Echinogorgia sp3 | OQ002389 | |
| 34 | Echinogorgia sp8 | OQ002390 | |
| 35 | Echinogorgia russelli | OQ002405 | |
| 36 | Echinogorgia ramulosa | OQ002404 | |
| 37 | Menella indica | OP984327 | |
| 38 | Euplexaura sp2 | OQ002379 |
References
- Zhao, H.T.; Wang, L.R. The general characteristics of the coral reef ecosystem. Chin. J. Ecol. 2001, 20, 41–45. (In Chinese) [Google Scholar]
- The Sixth Status of Corals of the World: 2020 Report. Available online: https://gcrmn.net/2020-report/ (accessed on 1 May 2023).
- Huang, H.; Zhang, C.L.; Yang, J.H.; You, F.; Lian, J.S.; Tan, Y.H. Scleractinian coral community characteristics in Zhubi reefsea area of Nansha Islands. J. Appl. Oceanogr. Taiwan Strait 2012, 31, 79–84. (In Chinese) [Google Scholar]
- Spalding, M.D.; Ravilious, C.; Green, E.P. World. Atlas of Coral Reefs; University of California Press: Berkeley, CA, USA, 2001. [Google Scholar]
- Zhao, H.T.; Wang, L.R.; Yuan, J.Y. Sustainable Development of the Coral Reefs in the South China Sea Islands. Trop. Geogr. 2016, 36, 55–65. (In Chinese) [Google Scholar]
- Liu, R.Y. Catalogue of Marine Life in China; Science Press: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Huang, L.T.; Huang, H.; Jiang, L. A revised taxonomy for Chinese hermatypic corals. Biodivers. Sci. 2020, 28, 515–523. (In Chinese) [Google Scholar] [CrossRef]
- Yang, S.L.; Yang, L.; Zhao, D.B.; Ren, Y.S. Species composition and distribution of neritic Scleractinian and Gorgonian corals in coastal waters of Fujian. J. Appl. Oceanogr. 2015, 34, 209–218. (In Chinese) [Google Scholar]
- Yang, S.L.; Zhao, D.B.; Ren, Y.C.; Yang, L.; Zhang, J.J.; Ji, H.D.; Luo, M.X.; Huang, J.J. Species composition and distribution of Scleractinian corals discovered in the East Fujian waters. J. Appl. Oceanogr. 2014, 33, 29–37. (In Chinese) [Google Scholar]
- Huang, Z.G. Species Diversity in Xiamen Bay; China Ocean Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Huang, H.; Li, X.B.; Lian, J.S.; Yang, J.H.; Li, Y.C.; Zou, R.L. Species diversity and distribution of Gorgonian (Cnidaria: Anthozoa: Octocoral) in Xiamen Bay and Dongshan Bay, Fujian. J. Oceanogr. Taiwan Strait 2007, 26, 92–98. (In Chinese) [Google Scholar]
- Ni, Z.; Liu, J.Y.; Song, Q.Q. Study on Investigation and Taxonomy of the Scleractinian Coralin Baiha Reef; Jimei University: Xiamen, China, 2018. (In Chinese) [Google Scholar]
- Lin, J.Y.; Wang, Y.P.; Yang, J.; Shi, Y.J.; Zhu, D.R. Revealing the Coral Species Diversity in Xiamen Bay: Spatial Distribution of Genus Astrogorgia (Cnidaria, Alcyonacea, Plexauridae) and Newly Recorded Species. J. Water. 2022, 14, 2417. [Google Scholar]
- Yang, J.; Liu, J.Y.; Wang, Y.P.; Shi, Y.J. Spatial distribution and new recorded species of Genus Carijoa in Xiamen Bay. J. Appl. Oceanogr. 2023, 32, 1–10. (In Chinese) [Google Scholar]
- Fisher, R.O.; Leary, R.A.; Low-Choy, S.; Brainard, R.E. Species richness on coral reefs and the pursuit of convergent global estimates. J. Curr. Biol. 2015, 25, 500–505. [Google Scholar] [CrossRef]
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. J. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Daly, M.; Brugler, M.P.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; France, S.C.; McFadden, C.S.; Opresko, D.M.; Rogriguez, E.; et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 2007, 1668, 1–766. [Google Scholar] [CrossRef]
- McFadden, C.S.; Van Ofwegen, L.P.; Quattrini, A.M. Revisionary systematics of Octocorallia (Cnidaria: Anthozoa) guided by phylogenomics. Bull. Soc. Syst. Biol. 2022, 1, 8735. [Google Scholar] [CrossRef]
- Liang, J.; Zhao, R.; Sui, Y.Z.; Zhang, Y.; Hu, Y.J.; Xu, B. Bioecological status, degradation causes and conservation suggestions of octocorals. J. Appl. Oceanogr. 2022, 1–14. Available online: https://kns.cnki.net/kcms/detail//35.1319.p.20221206.1759.001.html (accessed on 12 March 2023).
- Esper, E.J.C. Die Pflanzenthiere in Abbildungen Nach der Natur Mit Farben Erleuchtet, Nebst Beschreibungen. In Raspischen Buchhandlung; 1791; Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=267402 (accessed on 7 May 2023).
- Horton, T.; Kroh, A.; Ahyong, S.; Bailly, N.; Boury-Esnault, N.; Brandão, S.N.; Mees, J. World Register of Marine Species (WoRMS); WoRMS Editorial Board, 2018; Available online: https://www.marinespecies.org/ (accessed on 1 May 2023).
- Dai, C.F. Taxonomic morphologies and taxonomic systems of octocorallia. In A Complete Map of Corals in Taiwan: Eight Corals, 1st ed.; Li, J.H., Hu, J.Y., Eds.; Owl: Taiwan, China, 2022; pp. 1–408. (In Chinese) [Google Scholar]
- Li, X.B.; Huang, H.; Lian, J.S.; Yang, J.H.; Huang, L.M. Diversity and spatial distribution of gorgonian coral in Dongshan coastal waters in Fujian, China. J. Oceanogr. Taiwan Strait 2011, 30, 92–96. (In Chinese) [Google Scholar]
- Guo, Q.; Wei, Y.X.; Wang, C.Y.; Shao, C.L.; Li, L.; Liu, X.; Guo, Q.Q. Study on chemical constituents of Echinogorgia sassapo reticulata (Esper). Chin. J. Mar. Drugs 2010, 29, 672–681. (In Chinese) [Google Scholar]
- Wu, R.C.; Zhang, H.; Li, P.L.; Tang, X.L.; Li, G.Q. Studies on chemical constituents of Echinogorgia flora. Chin. J. Mar. Drugs 2013, 32, 23–28. (In Chinese) [Google Scholar]
- Wen, L.; Wei, Q.Q.; Cai, J.N. Advances in studies on bioactive secondary metabolites from gorgonian and microbial symbionts. Chin. J. Antibiot. 2012, 37, 641–654. [Google Scholar]
- Liang, X.Y.; Cao, F.; Wang, C.Y. Isolation, identification and bioactivities of farnesyl hydroquinone glycosides from the South China Sea gorgonian Echinogorgia rebekka. Chin. J. Mar. Drugs 2016, 35, 81–86. (In Chinese) [Google Scholar]
- Xue, L.; Guan, T.W.; Yu, Y.J.; Li, P.L.; Tang, X.L.; Li, G.Q. Studies on chemical constituents of the South China Seagorgonian Echinogorgia sassapo-reticulata. Chin. J. Mar. Drugs 2014, 33, 43–47. (In Chinese) [Google Scholar]
- Liao, L.; Wang, N.; Liang, Q. Three ceramides from gorgonian Echinogorgia sp. Chin. Tradit. Herb. Drugs 2010, 41, 851–854. (In Chinese) [Google Scholar]
- Liao, L.; Liao, X.J.; Xu, S.H. Compounds with Nitrogen in Coral Echinogorg sp. Nat. Prod. Res. Dev. 2010, 22, 392–394. (In Chinese) [Google Scholar]
- Breedy, O.; Guzman, H.M. A revision of the genus Psammogorgia Verrill, 1868 (Cnidaria, Anthozoa, Octocorallia) in the tropical eastern Pacific Ocean. ZooKeys 2020, 961, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Huang, H. Study on the Gorgonian Morphology and 18SrDNA Molecuar Phylogeny in the South China Sea; Sun Yat-sen University: Guangzhou, China, 2005. (In Chinese) [Google Scholar]
- Li, X.B.; Huang, H.; Zhang, J.B.; Dong, Z.J.; Huang, L.M. Phylogenetic Relationships with the Gorgonian (Cndaria: Anthozoa: Octocorallia) Based on 18S rDNA Sequences. Mar. Sci. Bullet. 2006, 25, 10–19. (In Chinese) [Google Scholar]
- Li, W.J.; Liu, C.W.; Liu, L.; Liao, B.L. Phylogeny analysis of Porites link in Xuwen using its gene. Mar. Sci. 2012, 36, 56–60. (In Chinese) [Google Scholar]
- Xie, Z.Q.; Liao, B.L.; Xiao, B.H.; Yang, X.D.; Li, X.J.; Chen, Y.Y. Phylogenetic Relationships in Scleractinian from Water of Dapeng Peninsula Based on ITS Genes. J. Guangdong Ocean. Univ. 2017, 37, 8–15. (In Chinese) [Google Scholar]
- Brockman, S.A.; McFadden, C.S. The mitochondrial genome of Paraminabea aldersladei (Cnidaria: Anthozoa: Octocorallia) supports intramolecular recombination as the primary mechanism of gene rearrangement in octocoral mitochondrial genomes. Genome Biol. Evol. 2012, 4, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- McFadden, C.S.; France, S.C.; Sánchez, J.A.; Alderslade, P. A molecular phylogenetic analysis of the Octocorallia (Cnidaria: Anthozoa) based on mitochondrial protein-coding sequences. Mol. Phylogenetics Evol. 2006, 41, 513–527. [Google Scholar] [CrossRef]
- Wang, P.Z.; Yang, C.J.; Wang, H.S.; Li, W.D. Preliminary identification of Aldersladum sp. in the South China Sea. J. Hainan Trop. Ocean. Univ. 2018, 25, 18–22. (In Chinese) [Google Scholar]
- Xu, Y. Taxonomy and Phylogeny of the Golden Coral Family Chrysogorgiidae (Anthozoa: Alcyonacea) from Seamounts in theWestern Pacific; University of Chinese Academy of Sciences: Beijing, China, 2019. (In Chinese) [Google Scholar]
- Hume, B.C.; Smith, E.G.; Ziegler, M.; Warrington, M.J.H.; Burt, A.J.; Wiedenmann, J. Sym Portal: A novel analytical frame-work and platform for coral algal symbiont next-generation sequencing ITS2 profiling. Mol. Ecol. Resour. 2019, 19, 1063–1080. [Google Scholar] [CrossRef]
- Mohammad, M.; Apandi, Z.N.; Marican, H.A.; Kamphol, N.; Mubin, N.A.; Salleh, S.; Ismail, M.N. The identification of Octocorals from the northern region of straits of Malacca. Trop. Life Sci. Res. 2016, 27 (Supp. 1), 87–93. [Google Scholar] [CrossRef] [PubMed]
- Alderslade, P.A.; Fabricius, K. Soft Corals and Sea Fans; Australian Institute of Marine Science: Cape Cleveland, Australia, 2008. [Google Scholar]
- Bayer, F.M. Key to the genera of Octocorallia exclusive of Pennatulacea (Coelenterata: Anthozoa), with diagnosis of new taxa. Proc. Biol. Soc. Wash. 1981, 94, 902–948. [Google Scholar]
- Cairns, S.D.; Wirshing, H.H. A phylogenetic analysis of the Primnoidae (Anthozoa: Octocorallia: Calcaxonia) with analyses of character evolution and a key to the genera and subgenera. BMC Evol. Biol. 2018, 18, 66. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38-Amplification and Directsequencing of Fungal Ribosomal RNA Genes for Phyogenetics; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Dai, C.F.; Qin, Q.X. Ecological map of Bafang Coral in Dongsha, 1st ed.; Cai, N.S., Wu, D.Y., Eds.; Marine National Park Administration: Kaohsiung, Taiwan, 2017; pp. 10–11. (In Chinese) [Google Scholar]
- Thomson, J.A.; Henderson, W.D. Report on the Alcyonaria Collected by Professor Herdman at Ceylon in 1902; Princeton University: Princeton, NJ, USA, 1905. [Google Scholar]
- Thomson, J.A.; Simpson, J.J. An account of the alcyonarians collected by the Royal Indian Marine Survey Ship ‘Investigator’ in the Indian Ocean. II. The alcyonarians of the littoral area. Trustees Indian Mus. Calcutta 1909, 1–202. [Google Scholar]
- Bayer, F.M. The Alcyonaria of Bikini and other atolls in the Marshall group. Part I Gorgonacea 1949, 1–20. [Google Scholar]
- Gray, J.E. XLVI.—Notes on some new genera and species of alcyonoid corals in the British Museum. J. Nat. Hist. 1870, 5, 405–408. [Google Scholar] [CrossRef]
- Kükenthal, W. Gorgonaria Wissenschaftliche Ergebnisse der deutschen Tiefsee-Expedition auf dem dampfer. Valdivia 1898, 1899, 1–946. [Google Scholar]
- Studer, T. Supplementary Report on the Alcyonaria Collected by HMS Challenger during the Years 1873–1876; Johnson Reprint Corporation: Berkeley, CA, USA, 1889. [Google Scholar]
- Kölliker, A. Die Bindesubstanz der Coelenteraten, Icones histologicae oder Atlas der vergleichenden Gewebslehre, II. Abt. I 1865. Available online: https://sc.panda321.com/scholar?hl=zh-cn&q=Die+Bindesubstanz+der+Coelenteraten%2C+Icones+histologicae+oder+Atlas+der+vergleichenden+Gewebslehre%2C+II.+Abt.+I+1865.++ (accessed on 1 May 2023).
- Koido, T.; Imahara, Y.; Fukami, H. High species diversity of the soft coral family Xeniidae (Octocorallia, Alcyonacea) in the temperate region of Japan revealed by morphological and molecular analyses. ZooKeys 2019, 862, 1–22. [Google Scholar] [CrossRef]
- Untiedt, C.B.; Quattrini, A.M.; McFadden, C.S.; Alderslade, P.A.; Pante, E.; Burridge, C.P. Phylogenetic relationships within Chrysogorgia (Alcyonacea: Octocorallia), a morphologically diverse genus of octocoral, revealed using a target enrichment approach. Front. Mar. Sci. 2021, 7, 2020. [Google Scholar] [CrossRef]
- Ament-Velásquez, S.L.; Breedy, O.; Cortés, J.; Guzman, H.M.; Wörheide, G.; Vargas, S. Homoplasious colony morphology and mito-nuclear phylogenetic discordance among Eastern Pacific octocorals. Mol. Phylogenet. Evol. 2016, 98, 373–381. [Google Scholar] [CrossRef]
- McFadden, C.S.; Haverkort-Yeh, R.; Reynolds, A.M.; Halàsz, A.; Quattrini, A.M.; Forsman, Z.H.; Toonen, R.J. Species boundaries in the absence of morphological, ecological or geographical differentiation in the Red Sea octocoral genus Ovabunda (Alcyonacea: Xeniidae). Mol. Phylogenet. Evol. 2017, 112, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, A.M.; Georgian, S.M.; Byrnes, L.; Stevens, A.; Falco, R.; Cordes, E.E. Niche divergence by deep-sea octocorals in the genus Callogorgia across the upper continental slope of the Gulf of Mexico. Mol. Ecol. 2013, 22, 4123–4140. [Google Scholar] [CrossRef] [PubMed]
- Kessel, G.M.; Alderslade, P.; Bilewitch, J.P.; Schnabel, K.E.; Gardner, J.P.A. The use of integrative taxonomy in Octocorallia (Cnidaria: Anthozoa): A literature survey. Zool. J. Linn. Soc. 2023, 198, 677–690. [Google Scholar] [CrossRef]
- Pérez, C.D.; Neves, B.; Cordeiro, R.T.S.; Williams, G.C.; Cairns, S.D. Diversity and distribution of Octocorallia. In The Cnidaria, Past, Present and Future; Stefano, G., Dubinsky, Z., Eds.; Springer International: Dordrecht, The Netherlands, 2016; pp. 109–123. [Google Scholar]
- Núñez-Flores, M.; Gomez-Uchida, D.; López-González, P.J. Molecular and morphological data reveal three new species of Thouarella Gray, 1870 (Anthozoa: Octocorallia: Primnoidae) from the Southern Ocean. Mar. Biodivers. 2020, 50, 30. [Google Scholar] [CrossRef]
- Poliseno, A.; Altuna, A.; Puetz, L.C.; Mak, S.S.T.; Rios, P.; Petroni, E.; McFadden, C.S.; Sørensen, M.V.; Gilbert, M.T.P. An integrated morphological-molecular approach reveals new insights on the systematics of the octocoral Telestula humilis (Thomson, 1927) (Octocorallia: Alcyonacea: Clavulariidae). Invertebr. Syst. 2021, 35, 261–281. [Google Scholar] [CrossRef]
- Bayer, F.M. Octocoral research: Past, present and future. Atoll Res. Bull. 2001, 494, 79–105. [Google Scholar] [CrossRef]
- McFadden, C.S.; Benayahu, Y.; Pante, E.; Thoma, J.N.; Nevarez, P.A.; France, S.C. Limitations of mitochondrial gene barcoding in Octocorallia. Mol. Ecol. Resour. 2010, 11, 19–31. [Google Scholar] [CrossRef]
- Baco, A.R.; Cairns, S.D. Comparing molecular variation to morphological species designations in the deep-sea coral Narella reveals new insights into seamount coral ranges. PLoS ONE 2012, 7, e45555. [Google Scholar] [CrossRef]
- McFadden, C.S.; Brown, A.S.; Brayton, C.; Hunt, C.B.; van Ofwegen, L.P. Application of DNA barcoding in biodiversity studies of shallow-water octocorals: Molecular proxies agree with morphological estimates of species richness in Palau. Coral Reefs 2014, 33, 275–286. [Google Scholar] [CrossRef]
- McFadden, C.S.; Reynolds, A.M.; Janes, M.P. DNA barcoding of xeniid soft corals (Octocorallia: Alcyonacea: Xeniidae) from Indonesia: Species richness and phylogenetic relationships. Syst. Biodivers. 2014, 12, 247–257. [Google Scholar] [CrossRef]
- Faircloth, B.C.; Branstetter, M.G.; White, N.D.; Brady, S.G. Target enrichment of ultraconserved elements from arthropods provides a genomic perspective on relationships among hymenoptera. Mol. Ecol. Resour. 2015, 15, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Harvey, M.G.; Smith, B.T.; Glenn, T.C.; Faircloth, B.C.; Brumfield, R.T. Sequence capture versus restriction site associated DNA sequencing for shallow systematics. Syst. Biol. 2016, 65, 910–924. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, A.M.; Wu, T.; Soong, K.; Jeng, M.S.; Benayahu, Y.; McFadden, C.S. A next generation approach to species delimitation reveals the role of hybridization in a cryptic species complex of corals. BMC Evol. Biol. 2019, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- Arrigoni, R.; Stefani, F.; Pichon, M.; Galli, P.; Benzoni, F. Molecular phylogeny of the robust clade (Faviidae, Mussidae, Merulinidae, and Pectiniidae): An Indian Ocean perspective. Mol. Phylogenetics Evol. 2012, 65, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhan, Z.; Xu, K. Morphology and phylogeny of Chrysogorgia pinniformis sp. nov. and C. varians sp. nov., two golden corals from the Caroline seamounts in the tropical Western Pacific Ocean. J. Oceanol. Limnol. 2021, 39, 1767–1789. [Google Scholar] [CrossRef]
- Nehoray, R.E.; Omri, S.; Maayan, Y.; Francis, C.M.; Godfray, C. Initiating a DNA Barcoding Reference Library of Stony Corals from the Gulf of Eilat (Red Sea). J. Mar. Sci. Eng. 2022, 10, 174–184. [Google Scholar]
- Li, X.N.; Liu, L.; Liu, C.W. Phylogenetic relationship in 8 species of Scleractinian coral based on COI sequences. Prog. Fish. Sci. 2012, 33, 83–88. (In Chinese) [Google Scholar]
- Zhou, Y.; Yang, C.J.; Wang, P.Z.; Wang, H.S.; Xu, Y.S.; Li, W.D. Identification of the soft coral genera Sarcophyton in the coastal waters of Hainan Island. J. Appl. Oceanogr. 2019, 38, 30–41. (In Chinese) [Google Scholar]
- Xiao, B.H.; Liao, B.L.; Yang, X.D. Study of phylogenetic relationships of scleractinian from Wailingding Island based on three standard molecular markers. Reports 2017, 41, 1–8. (In Chinese) [Google Scholar]
- Haverkort-Yeh, R.D.; McFadden, C.S.; Benayahu, Y.; Berumen, M.; Halász, A.; Toonen, R.J. A taxonomic survey of Saudi Arabian Red Sea octocorals (Cnidaria: Alcyonacea). Mar. Biodivers. 2013, 43, 279–291. [Google Scholar] [CrossRef]
- Babcock, R.C.; Baird, A.H.; Piromvaragorn, S.; Thomson, D.P.; Willis, B.L. Identification of scleractinian coral recruits from Indo-Pacific reefs. Zool. Stud. 2003, 42, 211–226. [Google Scholar]
- McCormack, J.E.; Tsai, W.L.E.; Faircloth, B.C. Sequence capture of ultraconserved elements from bird museum specimens. Mol. Ecol. Resour. 2016, 16, 1189–1203. [Google Scholar] [CrossRef] [PubMed]
- Ruane, S.; Austin, C.C. Phylogenomics using formalin-fixed and 100+ year-old intractable natural history specimens. Mol. Ecol. Resour. 2017, 17, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Wood, H.M.; González, V.L.; Lloyd, M.; Coddington, J.; Scharff, N. Next-generation museum genomics: Phylogenetic relationships among palpimanoid spiders using sequence capture techniques (Araneae: Palpimanoidea). Mol. Phylogenet. Evol. 2018, 127, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Derkarabetian, S.; Benavides, L.R.; Giribet, G. Sequence capture phylogenomics of historical ethanol-preserved museum specimens: Unlocking the rest of the vault. Mol. Ecol. Resour. 2019, 19, 1531–1544. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.L.; Pentico, A.; Quattrini, A.M.; McFadden, C.S. New approaches to species delimitation and population structure of anthozoans: Two case studies of octocorals using ultraconserved elements and exons. Mol Ecol Resour. 2021, 21, 78–92. [Google Scholar] [CrossRef]
- Quattrini, A.M.; Faircloth, B.C.; Dueñas, L.F.; Bridge, T.C.L.; Brugler, M.R.; Calixto-Botía, I.F.; DeLeo, D.M.; Forêt, S.; Herrera, S.; Lee, S.M.Y.; et al. Universal target-enrichment baits for anthozoan (Cnidaria) phylogenomics: New approaches to long-standing problems. Mol. Ecol. Resour. 2018, 18, 281–295. [Google Scholar] [CrossRef]
- Lam, K.; Morton, B. Soft corals, sea fans, gorgonians (Octocorallia: Alcyonacea) and black and wire corals (Ceriantipatharia: Antipatharia) from submarine caves in Hong Kong with a checklist of local species and a description of a new species of Paraminabea. J. Nat. Hist. 2008, 42, 9–12. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef]
- The Disappearance of Coral Reefs: Causes and Consequences. Available online: https://www.gvi.ie/blog/smb-the-disappearance-of-coral-reefs-causes-and-consequences/ (accessed on 6 September 2023).
- Jones, G.P.; McCormick, M.I.; Srinivasan, M.; Eagle, J.V. Coral decline threatens fish biodiversity in marine reserves. Proc. Natl. Acad. Sci. USA 2004, 101, 8251–8253. [Google Scholar] [CrossRef]
- Hein, M.Y.; Vardi, T.; Shaver, E.C.; Pioch, S.; Boström-Einarsson, L.; Ahmed, M.; Grimsditch, G.; McLeod, I.M. Perspectives on the use of coral reef restoration as a strat-egy to support and improve reef ecosystem services. Front. Mar. Sci. 2021, 8, 618303. [Google Scholar] [CrossRef]
- Bay, L.K.; Rocker, M.; Boström-Einarsson, L.; Babcock, R.; Buerger, P.; Cleves, P.; Webster, N. Reef Restoration and Adaptation Program: Intervention Technical Summary. In A Report Provided to the Australian Government by the Reef Restoration and Adaptation Program; Australian Institute of Marine Science: Townsville, Australia, 2019; p. 89. [Google Scholar]
- National Academies of Sciences, Engineering, and Medicine (NASEM). In A Research Review of Interventions to Increase the Persistence and Resilience of Coral Reefs; The National Academies Press: Washington, DC, USA, 2019. [CrossRef]
- List of National Nature Reserves in China. Available online: http://www.zrbhq.com.cn/index.php?m=content&c=index&a=lists&catid=81 (accessed on 1 July 2023). (In Chinese).
- Huang, H.; Lian, J.S.; Li, Z.X.; Chen, Z.Q. Fujian Dongshan Coral Nature Reserve and Its Biodiversity; China Ocean Press: Beijing, China, 2009. (In Chinese) [Google Scholar]
- Marine Protected Areas in Coral Reef Regions Classified According to Management Effectiveness Rating. Available online: https://www.wri.org/data/marine-protected-areas-coral-reef-regions-classified-according-management-effectiveness-rating (accessed on 6 September 2023).
- Fernandes, L.; Day, J.; Lewis, A.; Slegers, S.; Kerrigan, B.; Breen, D.; Cameron, D.; Jago, B.; Hall, J.; Lowe, D.; et al. Establishing representative no-take areas in the Great Barrier Reef: Large-scale implementation of theory on marine protected areas. Conserv. Biol. 2005, 19, 1733–1744. [Google Scholar] [CrossRef]
- Coral Reef Protected Areas: A Guide for Management. Available online: https://www.coralreef.gov/ecosystem/strate.html (accessed on 6 September 2023).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).