



Environmentally Realistic Waterborne Atrazine Exposure Affects Behavior in Poecilia latipinna


Department of Biology, Merrimack College, North Andover MA 01845, USA
Water 2023, 15(2), 306; https://doi.org/10.3390/w15020306
Submission received: 23 November 2022
/
Revised: 6 January 2023
/
Accepted: 9 January 2023
/
Published: 11 January 2023
(This article belongs to the Special Issue Impacts of Human Activities and Climate Change on Freshwater Fish, Volume II)
Abstract
:The present study examined the effects of environmentally realistic exposure to atrazine (ATZ) on the behavior of sailfin mollies, Poecilia latipinna. ATZ is one of the most commonly used pesticides in the US and a known endocrine disrupting chemical (EDC). The behavior of sailfin mollies is well documented in the scientific literature. Moreover, they are ecologically important indicators of environmental health, widely distributed among the mangroves along the Gulf Coast of the Southeastern US where significant amounts of ATZ are introduced via runoff and storm water drainage from coastal households, businesses, and farms. Four sets of experiments designed to assess ATZ’s impact on various aspects of male and female reproductive behavior, aggression, anxiety, and boldness were conducted following 12 weeks of exposure to 1 or 15 ppb water-borne ATZ, along with a no-exposure control group. Results indicated that the behavior of ATZ-exposed individuals differed from those of controls: ATZ exposure affected which stimulus fish (a male vs. a female) subject females preferred to associate with while also affecting female strength of preference for males of larger body size and their sexual receptivity to conspecific males in general. ATZ-exposed males also showed reduced overall responsiveness to conspecific stimuli and directed significantly less aggression toward their mirror image compared with controls. Finally, ATZ exposure affected multiple aspects of male and female behavior that are often used as proxies for boldness and anxiety. Overall, ATZ exposure resulted in alterations across a variety of behaviors attributed to sexual receptivity, mate choice and motivation to mate, aggression, as well as boldness and anxiety. These ATZ-induced behavioral changes may adversely affect the long-term health of natural populations exposed to similar, environmentally realistic concentrations and add to a growing body of empirical data demonstrating substantial fitness consequences of exposure to sublethal concentrations of this known EDC.
1. Introduction
Endocrine disrupting compounds (EDCs) are a common form of environmental pollution that disrupts proper hormonal functioning by affecting steroid hormone synthesis and/or their target cell receptors. Some of the more commonly identified EDCs found in nature include organochlorines, organophosphates, polychlorinated biphenyls (PCBs), phthalates, phytoestrogens, among other synthetic hormone mimics or blockers released into soil and water from industrial effluents, pesticide use, plastics, and sewage [1,2]. Given their potential to affect molecular and physiological processes [3,4,5,6,7,8,9], EDC exposure may induce a variety of maladaptive behavioral changes that compromise the health and integrity of natural populations even at very low, non-lethal, environmentally realistic concentrations [10,11,12,13]. Identification and monitoring of these changes may serve as an excellent indicator of toxin-induced environmental effects on wildlife that may often be observed long before physiological changes can be identified, e.g., [14,15,16,17]. Moreover, understanding individual- as well as population-level effects of toxin-altered behavior is important to conservation biologists in their efforts to restore contaminated ecosystems.
EDCs discharged into waterways have widespread effects on the sexual development and reproductive behavior of aquatic wildlife, particularly fish [18], the most common of which involve the disruption of species-specific courtship signals/displays, spawning, parental care, and/or positioning within social hierarchies [19,20,21,22,23,24]. Additional reproductive effects may include changes in the development of secondary sexual traits and/or delayed sexual maturity [13,25,26,27] for review. Moreover, since EDC exposure may also suppress aggression, it can disrupt social interactions necessary for successful reproduction. Diminished aggression notwithstanding, changes in anxiety level and/or boldness, among other non-reproductive behaviors, may also be affected by EDC exposure, which further compromise fitness [21,26,28,29,30,31,32,33,34].
The purpose of the present study was to examine the behavioral effects of atrazine (2-chloro-4-ethylamino-6-isopropylamino-1,3,5-triazine) exposure on the sailfin molly, Poecila latipinna. Atrazine (ATZ) is one of the most commonly used pesticides in the US [35,36] and known to possess neuroendocrine disrupting properties [37,38,39,40,41,42]. It often leeches into the aquatic environment [43] and is resistant to degradation [36,44]. As a consequence, many non-target aquatic species can be exposed to significant levels for most of their lives, particularly those living near agricultural fields [2].
ATZ has been implicated in the disruption of several molecular pathways within animal nervous and endocrine systems [37,38,41,45,46,47,48,49,50,51,52] including the conversion of the testosterone to estradiol, thereby ‘feminizing’ exposed individuals [53,54]. Previous research has demonstrated that ATZ exposure in fish can suppress levels of intrasexual aggression in male courtship behavior [2,19] and interfere with other reproductive and non-reproductive behavioral traits [34] including certain forms of social interaction [55]. In addition to studies of its effects on fish, ATZ exposure has been implicated in maladaptive changes in the personality phenotypes of crayfish [56] as well as increased anxiety and reduced spatial memory and learning in rodents [41,57,58].
The vast majority of studies addressing the effects of ATZ exposure have focused on males while its potential effects on females have been largely overlooked. Could the exposure to ecologically relevant levels of ATZ affect female physiology and behavior in a manner detrimental to their survival and/or reproductive success? Using the live-bearing fish species P. latipinna, the present study addresses whether chronic (12 weeks) exposure to ecologically realistic/biologically relevant levels of ATZ affect the behavior of both sexes. The behavior of P. latipinna is well documented and easily quantified [59]. Moreover, they are ecologically important indicators of environmental health, widely distributed among the mangroves along the Gulf Coast of the Southeastern US where significant, measurable amounts of ATZ are introduced via runoff, storm water drainage, etc. from coastal households, businesses, and farms.
Four sets of behavioral experiments were conducted using separate cohorts of male and female fish following 12 weeks of static/chronic water-borne ATZ exposure of 1 ppb (LD), 15 ppb (HD), or a no-ATZ control environment. The levels chosen were based on the maximum concentrations permitted in drinking water for most regions around the world (particularly in Asia and the US), which range from 0.1 to 5 ppb. However, environmental concentrations of 30 ppb in ground and surface waters have been reported [60], with levels in some locations occasionally exceeding 300 ppb [61].
The first two expts. (I and II) involved assessing the association preferences and overall responsiveness of fish presented with pairs of opposite sex conspecific dummy fish using a standard dichotomous choice design. The third expt. (III) examined female strength of preference (SOP) when given opportunities to select among dummy males of varying body sizes. Previous studies of P. latipinna, among other closely related species, show that females prefer to associate and mate with males of larger dorsal fin and body size [59,62,63,64,65,66]. Could the exposure to ecologically-relevant levels of ATZ affect these mate choice decisions? The fourth expt. (IV) measured male interactions with a mirror as a proxy for assessing the impact of AZT exposure on male aggression. Finally, time spent in the bottom zone (BZT) and latency to enter the tank’s top zone (LTZ) were measured during each subject’s period of acclimation to the novel test arena for all four experiments in order to assess anxiety and boldness following chemical exposure. Various iterations of these ‘novel arena tests’ have been used in previous studies of the behavioral impacts of chemical exposure, including ATZ, e.g., [34,67,68].
Subject behavior was compared among the LD, HD, and control groups in all four experiments with two primary objectives: 1. To assess whether exposure to ecologically realistic levels of ATZ affect sexual receptivity, mate choice and motivation to mate, aggression, boldness, and/or anxiety in P. latipinna—a model system in the study of animal behavior and ecology and important indicator species of environmental health within the mangroves along the Gulf Coast of the Southeastern US; and 2. To strengthen the viability of a relatively simple, practical, inexpensive protocol for use in addressing the behavioral impacts of potentially harmful environmental contaminants on fishes.
2. Materials and Methods
The research methods presented herein were described in Research Protocol 1RDM1018 and approved by the Institutional Animal Care and Use Committee of Merrimack College. All animal experiments comply with the National Institutes of Health guide for the care and use of Laboratory animals (NIH Publications No. 8023, revised 1978) and these guidelines have been followed.
2.1. Subjects, Housing, and Atrazine Preparation
ATZ was purchased from TCI, Tokyo Chemical Industry (Cat# A1650; Lot# 77P7D; CAS #1912-24-9). Test subjects were sexually mature, lab-reared P. latipinna raised from samples collected in Punta Gorda, FL, USA (26.9298° N, 82.0454° W). The fish, separated by sex, were initially maintained in several 70-l stock tanks under a 12:12 h light:dark regime with broad spectrum fluorescent lighting, at an average water temperature of 25 °C, and fed ad libitum once a day with flake food (Tetramin), tubifex worms, daphnia, or Artemia nauplii. A total of 150 test subjects (77 females and 73 males) were randomly assigned to one of three treatments. The treatments included a dimethylsulfoxide (DMSO, 6 ppb) as the solvent control, ATZ low-dose (1 ppb), and ATZ high-dose (15 ppb). A solvent control was used since ATZ was dissolved in DMSO [2]; all treatments received the same concentration of DMSO. ATZ concentrations used were based on US EPA estimated environmental concentrations [69]. Fish were transferred individually to separate ‘isolation tanks’ with 1.6 L of aged, conditioned water that contained ATZ dissolved in DMSO at a ‘low-dose’ (LD) concentration of 1 ppb (n = 50; 25 males and 25 females), ATZ dissolved in DMSO at a ‘high-dose’ concentration of 15 ppb (n = 50; 25 males and 25 females), or water prepared in an identical fashion to those containing ATZ, but containing no trace of the chemical (control; n = 50; 23 males and 27 females). Subjects remained in their randomly assigned isolation tanks for 12 weeks before becoming eligible for use as subjects in expts. I–IV described below. The concentration of atrazine in the water column was ascertained from three randomly selected LD, HD, and control tanks at the end of the first week of exposure using methodology employed in a similar study of ATZ exposure in guppies [2]. The average concentration was determined to be 0.35 and 13.22 ppb for the low- and high-dose, respectively, with negligible loss over the 7 days. No atrazine was detected in the control samples. Water was changed once a week with static renewal of chemical treatments for the duration of the 12-week exposure period and mortality was recorded every day. All aspects of data collection and analysis were conducted between June 2018 and March 2020 as time permitted.
Although the study described herein did not include analytical verification of ATZ concentrations beyond the first week of exposure (i.e., immediately prior to the first water change), all exposures were conducted in a closed system using the same exposure regime employed in similar studies where ATZ concentrations were verified [2,19,37,38]. Furthermore, the LD and HD concentrations of 1 and 15 ppb were identical to those used by Shenoy [2,19] in which negligible decreases were documented over 4–7 days without renewal in a closed system. Nonetheless, as recognized in similar studies [34], the use of non-measured concentrations may introduce some minor error.
2.2. Dummy Construction
In order to control for variation in behavior among other traits, dummy fish were constructed for use as stimuli in expts. I-III described below [70]. The dummies were constructed as described in MacLaren et al. [59]. For expts. I and II, two P. latipinna images were used to construct a pair of dummy stimuli, one of a displaying male measuring 60 mm in total length and a second image of a female measuring 50 mm in total length (approximate population means for Punta Gorda males and females used in this study; MacLaren, personal observation). For expt. III, the overall size of a single image of a displaying P. latipinna male was manipulated to generate ‘small’ (S), ‘intermediate’ (I), and ‘large’ (L) dummy stimuli measuring 40, 60, and 80 mm in total length, respectively (within the natural size range of males in this population; unpubl. data) in a similar fashion to those used in previous studies of this species [71,72] (Figure 1).
2.3. Testing Apparatus and Procedures for Experiments I–III
The testing environment for expts. I-III consisted of three 17.5-l aquaria (50 × 26 × 27 cm each) lined up end to end using an apparatus and protocol similar to the one used in previous poecilid mate preference experiments [59,62,63,64,65,66,72,73,74]. Subjects were transferred from their isolation tanks into an aquarium that was divided into three zones by two black vertical lines drawn on the front wall: 30 × 26 × 13.5 cm ‘neutral zone’ flanked on each side by a 10 × 26 × 13.5 cm ‘preference zone’. A pair of aquaria flanking the female’s tank each housed an adjustable motorized belt and pulley system to which the dummy stimuli were attached (see [62] for further details; Figure 2a).
For expt. III, all three possible pairwise combinations of three dummy males (L vs. S; L vs. I; and I vs. S) were presented in random order to each female test subject from the LD, HD, and control groups over a 2-h period. Subjects were fed approx. 15 min before introduction into the test arena. The female was then placed in the test arena and allowed to acclimate to the novel environment for a period of 10 min during which bottom zone time (BZT) and latency to enter the tank’s top zone (LTZ) were measured (see below for further details). Following the acclimation period, opaque screens blocking the female’s view of the dummies were removed and the apparatus was turned on in order for the dummies to begin moving. The female was videorecorded using an iPad for a period of 10 min before replacing the opaque screens for a 3-min mid-trial break during which dummies were switched to opposite sides of the arena. The opaque screens were then removed once more, resuming the trial for another 10 min of data collection. The opaque screens were again re-positioned in order to obstruct the female’s view of the dummies. This ‘dummy switching’ was conducted in order to control for possible side biases. The subject was left undisturbed in the arena for 5 min before proceeding to the second of the three pair-wise combinations of the three dummy males, attaching the appropriate, predetermined dummy males to their respective sides of the testing apparatus. The second and third trials proceeded in an identical fashion to the first.
For expts. I and II, a dummy male measuring 60 mm in total length and dummy female measuring 50 mm in total length, were randomly assigned to opposite ends of the motorized belt and pulley system flanking the test arena (Figure 2a). The behavioral assay then proceeded as described above (including measures of BZD and LTZ during each fish’s acclimation period) with the test subject exposed to both dummies simultaneously for 10 min. Next, the dummies were switched to opposite ends of the arena and tested for another 10 min. The only procedural difference between expts. I and II was the sex of the test subjects—females in I and males in II.
2.4. Procedure for Experiment IV
The testing environment for expt. IV consisted of a single 17.5-l aquarium (50 × 26 × 27 cm) divided into two zones by a black vertical line drawn on the front wall: 40 × 26 × 13.5 cm ‘neutral zone’ on one end of the tank flanked by a 10 × 26 × 13.5 cm ‘mirror zone’ on the opposite end (Figure 2b). Similar to expts. I–III, the male test subject was placed in the arena and allowed to acclimate to the novel environment for 10 min during which BZT and LTZ were measured. A mirror was then positioned against the outside of the glass on the ‘mirror zone’ end of the tank. Next, the male subject was videorecorded for a period of 10 min during which the time spent (s) interacting with the mirror (within the 10 cm mirror zone) was recorded.
2.5. Novel Arena Protocol
As noted above, the novel arena test was completed during the 10 min (600 s) acclimation phase for every test fish in all four experiments. Using an iPad, videorecording began immediately upon introducing the test subject into the novel arena. Total time spent (s) in the bottom zone [the total time spent within 5 cm of the bottom of the tank (BZT)] and latency (s) to enter the test arena’s top zone [the first time a fish came within 5 cm of the water’s surface (LTZ)] were documented for each test subject. Transition to the predetermined dummy stimulus or mirror test began immediately following the completion of the novel arena test.
2.6. Behavioral Measures and Statistical Analyses
For expts. I-III, test subject association preferences were based on total time spent with one dummy relative to the total time spent with the other dummy of the pair. The video data collected on iPads in expts. I-III were analyzed as follows: The total amount of time the subject spent in the preference zones of each of the two dummy stimuli during the course of the 20-min testing period (irrespective of the number of separate visits made to a given dummy preference zone) were summed for each subject. These measures served as the raw data (Supplementary Materials) for all statistical analyses that followed.
For expts. I and II, paired sample t-tests were used to compare the total time subjects spent in association with the male versus the female dummy stimulus. Mean strength of preference (SOP), defined as time spent with the male stimulus—time with the female stimulus per 20-min test, were then calculated and compared among the LD, HD, and control groups of expts. I-III using a one-way repeated measures ANOVA, where ‘SOP’ and ‘treatment’ served as the dependent and independent variables, respectively [59,62,66]. Where appropriate, the Holm-Sidak method for multiple comparisons was used to test for differences in SOP among treatments.
For expt. III, paired sample t-tests were used to compare the total time female subjects spent in association with the larger vs. smaller male dummy stimulus for all three trials (L vs. S; L vs. I; and I vs. S) for all three treatment groups (LD, HD, and controls). Mean strength of preference (SOP), defined as the time spent with the larger male—time with the smaller male per 20-min test, were then calculated and compared among the three paired combinations of dummy males (L vs. S; L vs. I; and I vs. S) for each of the three treatment groups (LD, HD, and controls) again using a one-way repeated measures ANOVA, where ‘SOP’ and ‘treatment’ served as the dependent and independent variables, respectively [59,66,75]. As described for expts. I and II above, the Holm-Sidak method for multiple comparisons was used to test for differences in SOP among treatments where appropriate. Since each female received three trials (L vs. S; L vs. I; and I vs. S), all significance levels were adjusted to p < 0.0167 (Bonferroni procedure; Sokal and Rohlf 1981).
The ‘overall responsiveness’ of subjects to the conspecific stimuli (defined as total time spent in association with both dummy stimuli during the course of the 20-min test period) was calculated for each subject and compared among the LD, HD, and control groups for each of the first three expts. (I–III) using one-way repeated measures ANOVA, where ‘overall responsiveness’ and ‘treatment’ served as the dependent and independent variables, respectively. Once again, where appropriate, the Holm-Sidak method for multiple comparisons was used to test for differences in overall responsiveness among treatments.
For expt. IV, iPad video data were used to quantify the total time (s) male subjects spent interacting with their mirror image (within the 10 cm mirror zone). These data were then used to compare the total mirror zone time for males among the LD, HD, and control groups using one-way repeated measures ANOVA, where ‘time spent in the mirror zone’ and ‘treatment’ served as the dependent and independent variables, respectively. The Holm-Sidak method for multiple comparisons was then used to test for differences in mirror zone time (a proxy for aggression) among treatments where appropriate.
Finally, video data collected during each subject’s 10 min acclimation period were analyzed for BZT and LTZ. These data were compiled across all four experiments and compared among the LD, HD, and control groups of each experiment using one-way repeated measures ANOVA, with ‘BZT’ and ‘LTZ’ as the dependent variables and ‘treatment’ as the independent variable. The probabilities reported for all statistical analyses are two-tailed. Standard parametric tests were applied to the data as no violations of the assumptions of normality and/or equal variance were identified. All statistical tests were conducted using SigmaStat (version 11.2).
3. Results
3.1. Experiment I: Comparison of Preferences for Male vs. Female Stimuli in AZT-Exposed vs. Unexposed Females
The male-directed behaviors observed most commonly during expts. I and III included gonopore thrusts, unison swimming, circling, backing toward the male, and presenting a flexed flank, all of which attributed to the mating behavior of poecilid females [76,77,78]. Control subjects from expt. I (n = 20) spent significantly more time associating with the male than the female dummy stimulus (T = 4.556; df = 19; p < 0.001; Figure 3a), whereas LD females (n = 20) associated more with the female stimulus fish over the male (T = 3.528; df = 19; p = 0.002; Figure 3a). The HD females (n = 20) showed no significant preference for either dummy stimulus (T = 0.734; df = 19; p = 0.472; Figure 3a).
Comparisons of female SOP among the LD, HD, and control groups revealed a significant difference (F = 19.064; df = 2, 57; p < 0.001). More specifically, SOP for the male over the female dummy stimulus was significantly greater among the control females than the HD (T = 3.359; df = 37; p = 0.002) or LD group (T = 5.718; df = 38; p < 0.001). Although less than control females, the mean SOP for the male dummy stimulus among HD females was also found to be significantly greater than for LD females (T = 3.253; df = 35; p = 0.003).
The overall response of subjects to the conspecific stimuli (defined as total time spent in association with both dummy stimuli during the course of the 20-min test period) also differed among the LD, HD, and control groups (F = 32.841; df = 2, 57; p < 0.001; Figure 4a). Control females spent significantly more time associating with the dummy stimuli than females from the HD (T = 5.778; df = 37; p < 0.001) or LD (T = 8.050; df = 38; p < 0.001) groups. However, HD females spent only marginally more time in association with the dummies than LD females (T = 1.979; df = 38; p = 0.055).
3.2. Experiment II: Comparison of Preferences for Male vs. Female Stimuli in AZT-Exposed vs. Unexposed Males
No significant preference for either dummy stimulus was observed for LD (n = 20; T = 1.155; df = 19; p = 0.262), HD (n = 20; T = 0.349; df = 19; p = 0.731), or control males (n = 20; T = 1.050; df = 19; p = 0.307; Figure 3b); nor was there a significant difference in SOP among the treatment groups and controls (F = 0.290; df = 2, 57; p = 0.749). However, a significant difference was observed among treatment and control males in their overall responsiveness to (association times with) both stimulus fish combined (F = 41.609; df = 2, 57; p < 0.001; Figure 4b). Specifically, males from the LD (T = 11.257; df = 33; p < 0.001) and HD (T = 7.097; df = 25; p < 0.001) groups both spent less time in association with the stimulus fish when compared with controls. However, no difference in total time spent with the stimulus fish was found between the LD and HD groups (T = 0.596; df = 32; p = 0.555; Figure 4b).
3.3. Experiment III: Comparison of Preferences for Male Body Size in AZT-Exposed vs. Unexposed Females
Paired sample t-tests comparing the total time control females (n = 26) spent in association with the larger- vs. smaller-bodied male dummy revealed that females spent significantly more time with the larger of the paired males in each of the three trials (L vs. S; L vs. I; and I vs. S; Table 1). However, the only trial in which HD females (n = 20) preferred the larger male was ‘L vs. S’—the trial in which the difference in body size was the largest of the three pairwise combinations. Females from the LD group (n = 18) showed no preferences for either male in any of the three trials (Table 1).
Mean SOP for larger size in the ‘L vs. S’, ‘L vs. I’, and ‘I vs. S’ trials all differed among the HD, LD, and control groups (Figure 5). Specifically, the SOP of control females for the larger male body size was significantly greater than the LD females in the ‘L vs. S’, ‘L vs. I’, and ‘I vs. S’ trials (Figure 5). The SOP of control females in the ‘L vs. I’, and ‘I vs. S’ trials was significantly greater than the HD females, as well. However, no difference in SOP was observed between the control and HD females in their ‘L vs. S’ trials.
The overall responsiveness of female test subjects to the stimulus fish (i.e., total time spent with both dummies combined across all three trials) differed among the treatment groups and control (F = 7.878; df = 2,61; p < 0.001). Specifically, LD females were less responsive to the stimulus fish (mean = 1027.11 ± 53.34) in comparison with the control (mean = 1137.69 ± 23.88; F = 2.098; df = 42; p = 0.042) and HD (mean = 1265.90 ± 47.47; F = 3.348; df = 36; p = 0.002) groups. However, no difference was observed between HD females and the controls.
3.4. Experiment IV: Comparison of Time Spent in the Mirror Zone for AZT-Exposed vs. Unexposed Males
The time that males spent interacting with the mirror differed among the LD, HD, and control groups (F = 37.932; df = 2, 57; p < 0.001). Specifically, control males spent more time in the mirror zone than the LD (T = 8.539; df = 37; p < 0.001) or HD (T = 5.981; df = 38; p < 0.001) groups. Additionally, HD males spent significantly more time in the mirror zone than LD males (T = 2.246; df = 38; p < 0.031; Figure 6).
3.5. Comparison of Novel Arena Test Results for AZT-Exposed vs. Unexposed Fish from All Four Experiments
The time spent in the bottom zone ranged nearly the full length of the arena acclimation period from 43–600 s and differed among the LD, HD, and control groups for females (F = 55.481; df = 2121; p < 0.001; Figure 7a) as well as males (F = 59.913; df = 2117; p < 0.001; Figure 7a). LD females spent more time in the bottom zone during the novel arena test compared to both HD (T = 2.602; df = 73; p = 0.011) and control females (T = 9.606; df = 76; p < 0.001). Moreover, HD females spent more time in the bottom zone in comparison with controls (T = 7.781; df = 84; p < 0.001).
The results for male test subjects mirrored those of females: LD males spent more time in the bottom zone compared to HD (T = 2.632; df = 74; p = 0.010) and control (T = 10.089; df = 74; p < 0.001) males (Figure 7a). Moreover, HD males spent more time in the bottom zone in comparison with controls (T = 8.561; df = 78; p < 0.001; Figure 7a). Of note, male fish from all three groups combined (LD + HD + controls; n = 120) spent less time in the bottom zone than females from all three groups combined (n = 124), averaging 306.63 ± 13.09 s and 344.02 ± 12.66 s, respectively (T = 2.053; df = 241; p = 0.041).
Exposure to AZT significantly increased the latency to enter the top zone of the novel arena in both the LD and HD treatment groups compared to controls for females (F = 47.687; df = 2121; p < 0.001; Figure 7b) and males (F = 19.074; df = 2.117; p < 0.001; Figure 7b). Similar to the results for BZT, LD females took significantly longer to enter the top zone compared to the HD (T = 2.177; df = 76; p = 0.033) or control females (T = 9.793; df = 45; p < 0.001) and HD females took longer to enter the top zone than control as well (T = 7.243; df = 49; p < 0.001).
Once again, the LTZ results for male subjects largely mirror those observed for females: Both LD (T = 5.869; df = 57; p < 0.001) and HD males (T = 6.483; df = 68; p < 0.001) took significantly longer to enter the top zone in comparison with controls (Figure 7b). However, no significant difference in mean LTZ was observed between the LD and HD males (T = 1.830; df = 72; p = 0.071). Once again, similar to the BZT data, male fish from all three groups combined (LD + HD + controls; n = 120) took significantly less time to enter the top zone (mean for males = 153.00 ± 12.70 s; n = 120) than females (mean for females = 247.98 ± 16.11 s; n = 124; T = 4.631; df = 231; p < 0.001). Finally, ATZ treatments did not influence mortality rates of males or females in comparison with their respective control groups.
4. Discussion
Although ATZ treatments did not influence mortality rates, the results of the present study indicate that the behavior of male and female P. latipinna was affected by exposure to the chemical, particularly at the 1 ppb level (LD).
4.1. Effects on Female Behavior
ATZ exposure affected which (if either) dummy stimulus the female subjects preferred in expt. I. The LD and control females showed opposite preferences for the female and male stimulus fish, respectively; while the HD females preferred neither dummy. These data indicate that ATZ (particularly at 1 ppb) reduced female responsiveness to/interest in associating with males, favoring avoidance rather than the affiliative behavior observed among controls. Similar to most poecilids, male P. latipinna are sexually coercive in their efforts to secure mating opportunities, particularly males of smaller body size (personal observation.). Therefore, for sexually non-receptive females (whether AZT-induced or otherwise), male avoidance behavior could minimize the risk of injury and/or other forms of stress.
ATZ exposure not only affected interest in males, but their mating preferences, as well. The control females of expt. III preferred males of larger size, as observed in previous studies of P. latipinna and related species [59]. However, LD females showed no preferences, spending less time with either stimulus male when compared with controls. Therefore, ATZ exposure affected the species-typical preference for male body size and, in combination with the results from expt. I, further suggest that ATZ exposure suppressed female tendency/motivation to associate with conspecific males regardless of size.
ATZ exposure, particularly at 1 ppb (LD), diminished sexual receptivity to males in expts. I and III, facilitating the preference for the female stimulus fish in the former and lowering responsiveness to all stimulus males in the latter when compared with the control females. Furthermore, the fact that LD females from both experiments were less responsive to either dummy compared to the controls indicates that the 1 ppb concentration may have had a more generalized effect on the affiliative behavior of females, regardless of sex. For a species such as P. latipinna, for which shoaling is an important anti-predator behavior, these ATZ-induced changes in social behavior may decrease the survivorship of affected individuals, not to mention the long-term fitness consequences of reduced sexual receptivity to males.
LD females exhibited more pronounced male avoidance behavior and were less responsive to the dummies overall in comparison not only with the control group, but HD females as well. This type of non-monotonic, ‘inverted U-shaped’ response curve is often noted in the endocrinology literature, e.g., [19,34] and has been reported for EDCs [79,80,81] including ATZ [19,34,82,83,84,85]. Shenoy [19] attributed these response curves to the effects of hormone receptor saturation that stymy further enhancement of signal transduction pathways—cellular responses that would otherwise continue rising in concert with elevated hormone levels [81]. These abnormally high blood-borne hormone (or hormone mimic) concentrations may also inhibit further hormone synthesis, facilitating the removal of excess hormone receptors [19]. In a similar study of ATZ’s behavioral effects on a molly congener, P. reticulata, Shenoy [2,19] found that males prenatally exposed to 1 ppb were consistently less aggressive (within a mating context) than those prenatally exposed to 15 ppb. Shenoy [19] further noted that the effects of the 15 ppb ATZ exposure for 16 weeks were often indistinguishable from the controls. Similar to the females of expts. I and III, a similar non-monotonic response curve was observed among males involved in the mirror test of expt. IV discussed below. The data for P. latipinna and P. reticulata [19] collectively show the detrimental effects that even (and perhaps especially) very low concentrations of contaminants can have on wildlife, and illustrate how low-dose exposure may be more impactful than exposure to higher doses. Namely, it is important to note that exposure to higher EDC levels cannot be considered benign as they may affect other aspects of animal health that were not addressed in these studies.
4.2. Effects on Male Behavior
In contrast to females in expt. I, neither the ATZ-exposed nor control males preferred the female or male stimulus fish in expt. II. However, the lack of preference for the female dummy is not necessarily evidence for ATZ-induced suppression of interest/motivation to court and mate since no preference was observed among control males. Similarly, the lack of significant association time with the male dummy cannot be taken as stand-alone evidence for chemically-triggered reduction in aggressive behavior for the same reason. However, similar to the results for females in expt. I, both the LD and HD males of expt. II were less responsive to the dummy stimuli overall in comparison to controls.
Data from the mirror test (expt. IV) revealed a similar trend of diminished male responsiveness. ATZ exposure reduced interactions with a mirror—results that corroborate those described in a recent study of zebrafish, Danio rerio [34] wherein paternal exposure to ATZ during juvenile development (at 0.3 and 30 ppb) was shown to significantly influence the mirror interactions of their unexposed F1 adult offspring using a similar behavioral assay. In fact, numerous studies of fish behavior have employed some form of ‘mirror test’ to assess an individual’s level of aggression (e.g., D. rerio [34,86,87] and Betta splendens [88]). Similar to many fishes, male P. latipinna are thought to perceive their mirror image as a conspecific rival (personal observation). The diminished activity levels of ATZ-exposed individuals observed in the mirror test suggest that exposed mollies may be more cautious and less motivated to engage in aggressive interactions with potential rivals than control males—a phenomenon described in previous studies of male-male interactions in guppies [2] as well as zebrafish [34] that corroborates the suppressed aggression described in similar studies of male sexual and social behavior following direct exposure to ATZ [2,19,34,39,40].
4.3. Novel Area Test
ATZ exposure altered both the latency to enter the top zone of a tank (LTZ) and bottom zone time (BZT) for both sexes during the ‘novel arena test’ phases of all four experiments—an effect found to be more pronounced in females than males, as observed in a similar study of ATZ exposure in zebrafish [34]. As typical of many aquarium fishes, e.g., [34,67,89], when exposed to an unfamiliar environment, P. latipinna tend to dive to the bottom of the tank and remain in the lowest portion (bottom zone), avoiding the higher portions of the water column for a variable period of time (usually a few minutes) before exploring other areas of the tank (MacLaren, personal observation).
Various iterations of the novel arena test have been used to assess anxiety, activity, and exploration in fishes following chemical exposure, including ATZ, e.g., [34,67,68]. The results presented herein mirror those described in similar published reports. A recent zebrafish study, for example, showed that paternal exposure to ATZ during juvenile development significantly influenced both BZT (at 0.3 ppb) and LTZ (at 0.3, 3, and 30 ppb) of unexposed F1 offspring at adulthood [34]. The authors suggested that the trend of increased time spent in the bottom zone in the novel arena may result from ATZ-induced increases in anxiety—a conclusion supported by results obtained in previous studies in fishes [90] as well as rodents [41,49].
Increases in LTZ may also result from changes in anxiety due to ATZ exposure [28,67,68,89,91]. Under more natural circumstances, swimming to the surface or otherwise spending time in the top portion of the water column undoubtedly exposes mollies, among other small fishes, to an increased risk of predation [91,92,93,94]. It is relatively commonplace for P. latipinna to engage in defensive behaviors, such as increased BZT when placed in a novel environment and/or when they perceive an eminent predation threat. With time, however, they will generally begin (or resume) exploration of their environment, including the area close to the surface [67,68,95]. Prolonged latency to enter the top zone or even move from a stationary position at the bottom of the novel tank (as observed to a larger extent among ATZ treated mollies compared to controls) might reflect a compromised ability to assess risk and thereby diminish motivation to engage in more bold, exploratory behavior. Under more natural circumstances, environmental risk assessment is a critical aspect of successful foraging, exploratory, and anti-predator behavior for many fishes [96]. Although ‘bolder’ individuals subject themselves to greater predation pressure [97,98], their overly anxious counterparts may also suffer fitness consequences with respect to growth and reproduction given its impact on foraging efficiency among other stress-induced physiological and behavioral effects [34,99,100].
4.4. Behavior As An Indicator of Environmental Health
It is important to emphasize that all behavioral changes observed in the present study were effects of exposure to environmentally relevant ATZ concentrations (1–15 ppb are doses well within the ranges likely to be encountered in the environment [43]). As the blossoming science of behavioral ecotoxicology continue to show [101], behavior may serve as an excellent indicator of the environmental health of a population and/or ecosystem ([14,15,16], but see [17]). Therefore, understanding individual- as well as population-level effects of toxin-altered behavior is very important to conservation biology. Many contaminants, such as ATZ are persistent and remain in the environment at substantial concentrations for several years, spanning multiple generations in short-lived species, such as P. latipinna [102]. The cross-generational impact of toxin-altered behavior patterns, particularly in the context of reproduction (e.g., weakened mate choice preferences, diminished sexual responsiveness to courting males, motivation to mate and/or sexual receptivity in general) may threaten the continued existence of affected species in these polluted ecosystems [27].
4.5. Possible Mechanisms Underlying Atrazine’s Effects on Behavior
Numerous studies addressing the health ramifications of exposure to sublethal, environmentally realistic concentrations of water-borne ATZ have described ‘estrogenic effects’ [54,103] with detrimental downstream impacts on fecundity and sperm counts as well as changes in gonadal morphology and hormone profiles (see [104] for review of several of these studies). Regarding its mechanism(s) of action, ATZ is believed to enhance the production of aromatase, an enzyme involved in the metabolic conversion of testosterone to estradiol [53,54]. This upregulation of aromatase reduces the blood levels of testosterone while elevating estradiol, which then suppresses further testosterone synthesis [104,105]. The reduction in testosterone levels impacts the synthesis of dihydrotestosterone (DHT) and 11- ketotestosterone (11-KT), both of which are critical for proper sexual development [106,107]. For example, 11-KT, has been shown to be important for courtship behavior in male mosquitofish, Gambusia holbrooki [108] and guppies [109,110], and for sex recognition and courtship in male goldfish, Carassius auratus [111] and sailfin mollies [112]. Moreover, aromatase is involved in the conversion of androstenedione to estrone [19]. Since testosterone is produced from androstenedione and estradiol from estrone, elevated aromatase levels can further alter testosterone:estradiol ratios critical for proper development and expression of species-typical sexual traits and behaviors [19]. Additional proposed mechanisms of ATZ-induced endocrine disruption include direct inhibition of gonadal testosterone production [113] as well as reduced levels of 5α-reductase—an enzyme involved in converting testosterone to DHT [114,115].
While the effects of ATZ exposure on male behavior and physiology have been addressed in several studies, very few have addressed ATZ’s impact on females, particularly in the context of reproductive behavior. The results of the present study provide clear evidence of potentially harmful behavioral effects of environmentally realistic ATZ exposure in females as well, especially at very small (1 ppb) concentrations. ATZ’s more pronounced effects on BZT and AZT in females compared to males coupled with the diminished sexual receptivity observed among exposed females in the present study bring to light a need for additional research that addresses the underlying mechanistic basis of these behavioral changes and the extent to which they may be observed in other species.
It is possible that ATZ’s impact on sexual behaviors in the present study may be due to poor health in general, given the parrel changes in non-sexual behaviors that were also observed. Alternatively, it could be that ATZ had a negligible impact on hormone production, secretion, metabolism, and/or the receptivity of target cells in P. latipinna. Rather, ATZ may be affecting their behavior via an entirely different as yet unidentified mechanism, although a large and growing body of evidence suggests that is highly unlikely. The dearth of studies addressing how endocrine disrupting chemicals may interfere with normal behavior in P. latipinna and poecilid fishes in general make the results of the present study all the more important a contribution to advancing our understanding of the ecological impacts of pesticides among other anthropogenic chemicals as they are introduced into the environment.
It is also important to note the use of static and constant exposures in the present study as these conditions do not adequately simulate those experienced by aquatic life in their natural environment. Chemicals from wastewater treatment plants, farmlands, among other anthropogenic sources, are dispersed in a manner where organisms will never experience constant levels. Moreover, aquatic organisms are exposed to many different chemicals all at once, which may impact the effect any one of them has in the absence of others. Once again, additional research is necessary in order for conservationists to achieve a better grasp on the myriad of possible health effects associated with exposure to ATZ in combination with other potentially harmful chemical contaminants in nature.
4.6. Justification of Methods
Time spent in association with a stimulus fish (i.e., within a ‘preference zone’) is conventionally used as a proxy for assessment of mating preferences and are positively correlated with probability of copulation in several species wherein the relationship has been studied (e.g., guppies: [116,117]; gobies: [118]; and pipefish, Syngnathus typhle: [119]). Nonetheless, the results and conclusions of the present study should be interpreted with caution. In an effort to control for as many outside variables as possible, single male and female photographs were used to create all dummy stimuli used in expts. I-III. This approach made for a more direct examination of the research questions at hand [Do subjects prefer to associate with a male or female stimulus fish? (expts. I and II) and Do females prefer males of larger or smaller body size when given the opportunity to choose? expt. III)]. However, it can also result in psuedoreplication. Subject responses to dummy stimuli may also be affected by other aspects of body and/or fin shape, size, or coloration, among other cues. Using the same fish image throughout negates this issue. Moreover, the methodology herein has been used in several previously published studies of mating preferences for visual traits in Poeciliid fishes including P. latipinna, e.g., [59,71,72,73], X. helleri [62,74], X. variatus [64], and P. reticulata [63].
Finally, the behavioral responses that subjects directed to the dummy stimuli in the present study were comparable to those observed in previous studies of P. latipinna using similar methodology [59,71,72,73] as well as experiments wherein live male P. latipinna were used as stimuli [120,121,122], thereby strengthen the viability of this relatively simple, practical, inexpensive new methodology for use in studies of the potentially harmful behavioral impacts of EDCs and other contaminants on fishes.
5. Conclusions
ATZ exposure resulted in alterations across a variety of behaviors attributed to sexual receptivity, mate choice and motivation to mate, aggression, as well as boldness and anxiety in P. latipinna—a model system in the study of animal behavior and ecology and important indicator species of environmental health within the mangroves along the Gulf Coast of the Southeastern US. The ATZ-induced behavioral changes seen here for P. latipinna are sufficiently significant to be of concern, potentially impacting the long-term health of natural populations exposed to similar, environmentally realistic concentrations. There are clear fitness implications for P. latipinna among many other species native to areas exposed to ATZ from myriad of anthropogenic sources. Understanding the effects on sexual behavior is particularly critical given its impact on reproduction and populations dynamics. The present study demonstrates a continued need for careful examination of ATZ’s impact on aquatic life. Studies of the effects of ATZ at a wider range of doses, over longer periods of exposure, and in other species, will offer a better understanding of the endocrine and behavioral impacts of exposure and its effects on survival and reproductive success of both males and females in the aquatic environment. Moreover, since ATZ’s mechanism of action is similar across most vertebrate taxa studied to date, there are clear implications for human health, as well [2,103].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15020306/s1, Table S1: P. latipinna raw data for expts. I–IV for water. Raw behavioral data collected and used in all statistical analyses conducted in the present study.
Funding
This study was supported in part by 2018, 2019, and 2020 Merrimack College Faculty Development Grants and a 2018 Merrimack College Provost Innovation Grant. All four forms of funding are internal and include no grant numbers or funds from external sources requiring references to the funder’s websites. The funders (Merrimack College) had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.
Acknowledgments
The author would like to express their gratitude to all those who contributed to this effort including Thuy Tran for her assistance in purchasing materials and lab maintenance and Merrimack undergraduates N. Nanof, H. McKeever, A. Nolan, A. Figucia, and G. Eldridge for their assistance in data collection and analysis.
Conflicts of Interest
The author declares no conflict of interest.
References
- Markman, S.; Guschina, I.A.; Barnsley, S.; Buchanan, K.L.; Pascoe, D.; Müller, C.T. Endocrine disrupting chemicals accumulate in earthworms exposed to sewage effluent. Chemosphere 2007, 70, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, K. Environmentally realistic exposure to the herbicide atrazine alters some sexually selected traits in male guppies. PLoS ONE 2012, 7, e30611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filby, A.L.; Paull, G.C.; Hickmore, T.F.; Tyler, C.R. Unravelling the neurophysiological basis of aggression in a fish model. BMC Genom. 2010, 11, 498. [Google Scholar] [CrossRef] [Green Version]
- Thornqvist, P.-O.; Hoglund, E.; Winberg, S. Natural selection constrain personality and brain gene expression differences in Atlantic salmon (Salmo salar). J. Exp. Biol. 2015, 218, 1077–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield, C.W.; Cziko, A.-M.; Robinson, G.E. Gene expression profiles in the brain predict behavior in individual honey bees. Science 2003, 302, 296–299. [Google Scholar] [CrossRef]
- Wiese, A.-S.; Needham, E.K.; Noer, C.L.; Balsby, T.J.S.; Dabelsteen, T.; Pakkenberg, B. The number of neurons in specific amygdala regions is associated with boldness in mink: A study in animal personality. Brain Struct. Funct. 2018, 223, 1989–1998. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Hotchkiss, A.K.; Rider, C.V.; Blystone, C.R.; Wilson, V.S.; Hartig, P.C.; Ankley, G.T.; Foster, P.M.; Gray, C.L.; Gray, L.E. Fifteen years after “Wingspread”—Environmental endocrine disrupters and human and wildlife health: Where we are today and where we need to go. Toxicol. Sci. 2008, 105, 235–259. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Baccarelli, A.; Pesatori, A.C.; Bertazzi, P.A. Occupational and environmental agents as endocrine disruptors: Experimental and human evidence. J. Endocrinol. Investig. 2000, 23, 771–781. [Google Scholar] [CrossRef]
- Frye, C.A.; Bo, E.; Calamandreis, G.; Calza, L.; Dessi-Fulgheri, F.; Fernández, M.; Fusani, L.; Kah, O.; Kajta, M.; Le Page, Y.; et al. Endocrine disrupters: A review of some sources, effects, andmechanisms of actions on behavior and neuroendocrine systems. J. Neuroendocrinol. 2012, 24, 144–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLachlan, J. Environmental signaling: What embryos and evolution teach us about endocrine disrupting chemicals. Endocr. Rev. 2001, 22, 319–341. [Google Scholar] [CrossRef] [PubMed]
- Clotfelter, E.D.; Bell, A.M.; Levering, K.R. The role of animal behavior in the study of endocrine-disrupting chemicals. Anim. Behav. 2004, 68, 665–676. [Google Scholar] [CrossRef]
- Little, E.E. Behavioral measures of environmental stressors in fish. In Biological Indicators of Aquatic Ecosystem Stress; Adams, S.M., Ed.; American Fisheries Society: Bethesda, MD, USA, 2002; pp. 431–472. [Google Scholar]
- Perreault, H.A.N.; Semsar, K.; Godwin, J. Fluoxetine treatment decreases territorial aggression in a coral reef fish. Physiol. Behav. 2003, 70, 719–724. [Google Scholar] [CrossRef]
- Lynn, S.E.; Egar, J.M.; Walker, B.G.; Sperry, T.S.; Ramenofsky, M. Fish on Prozac: A simple, noninvasive physiology laboratory investigating the mechanisms of aggressive behavior in Betta splendens. Adv. Physiol. Educ. 2007, 31, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Schultz, M.M.; Painter, M.M.; Bartell, S.E.; Logue, A.; Furlong, E.T.; Werner, S.L.; Schoenfuss, H.L. Selective uptake and biological consequences of environmentally relevant antidepressant pharmaceutical exposures on male fathead minnows. Aquat. Toxicol. 2011, 104, 38–47. [Google Scholar] [CrossRef]
- Tyler, C.R.; Jobling, S.; Sumpter, J.P. Endocrine disruption in wildlife: A critical review of the evidence. Crit. Rev. Toxicol. 1998, 28, 319–361. [Google Scholar] [CrossRef]
- Shenoy, K. Prenatal exposure to low doses of atrazine affects mating behaviors in male guppies. Horm. Behav. 2014, 66, 439–448. [Google Scholar] [CrossRef]
- Soffker, M.; Tyler, C. Endocrine disrupting chemicals and sexual behaviors in fish—A critical review on effects and possible consequences. Crit. Rev. Toxicol. 2012, 42, 8. [Google Scholar] [CrossRef]
- Bertram, M.G.; Saaristo, M.; Ecker, T.E.; Baumgartner, J.B.; Wong, B. An androgenic endocrine disruptor alters male mating behavior in the guppy (Poecilia reticulata). Behav. Ecol. 2018, 29, 1255–1263. [Google Scholar] [CrossRef]
- Bell, A.M. Effects of an endocrine disrupter on courtship and aggressive behaviour of male three-spined stickleback, Gasterosteus aculeatus. Anim. Behav. 2001, 62, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Clotfelter, E.D.; Rodriguez, A.C. Behavioral changes in fish exposed to phytoestrogens. Environ. Pollut. 2006, 144, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Martinović, D.; Hogarth, W.T.; Jones, R.E.; Sorensen, P.W. Environmental estrogens suppress hormones, behavior and reproductive fitness in male fathead minnows. Environ. Toxicol. Chem. 2007, 26, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisp, T.M.; Clegg, E.D.; Cooper, R.L.; Wood, W.P.; Anderson, D.G.; Baetcke, K.P.; Hoffman, J.L.; Morrow, M.S.; Rodier, D.J.; Schaeffer, J.E.; et al. Environmental endocrine disruption: An effects assessment and analysis. Environ. Health Perspect. 1998, 106, 11–56. [Google Scholar] [PubMed]
- Dzieweczynski, T.L.; Hebert, O.L. The effects of short-term exposure to an endocrine disrupter on behavioral consistency in male juvenile and adult Siamese fighting fish. Arch. Environ. Contam. Toxicol. 2013, 64, 316–326. [Google Scholar] [CrossRef]
- Shenoy, K.; Crowley, P.H. Endocrine disruption of male mating signals: Ecological and evolutionary implications. Funct. Ecol. 2011, 25, 433–448. [Google Scholar] [CrossRef]
- Volkova, K.; Caspillo, N.R.; Porseryd, T.; Hallgren, S.; Dinnétz, P.; Porsch-Hällström, I. Developmental exposure of zebrafish (Danio rerio) to 17α-ethinylestradiol affects non-reproductive behavior and fertility as adults, and increases anxiety in unexposed progeny. Horm. Behav. 2015, 73, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Dzieweczynski, T.L. Short-term exposure to an endocrine disruptor affects behavioural consistency in male threespine stickleback. Aquat. Toxicol. 2011, 105, 681–687. [Google Scholar] [CrossRef]
- Dzieweczynski, T.L.; Campbell, B.A.; Marks, J.M.; Logan, B. Acute exposure to 17α-ethinylestradiol alters boldness behavioral syndrome in female Siamese fighting fish. Horm. Behav. 2014, 66, 577–584. [Google Scholar] [CrossRef]
- Hebert, O.L.; Lavin, L.E.; Marks, J.M.; Dzieweczynski, T.L. The effects of 17α-ethinyloestradiol on boldness and its relationship to decision making in male Siamese fighting fish. Anim. Behav. 2014, 87, 203–212. [Google Scholar] [CrossRef]
- Lagesson, A.; Saaristo, M.; Brodin, T.; Fick, J.; Klaminder, J.; Martin, J.; Wong, B.B.M. Fish on steroids: Temperature-dependent effects of 17β-trenbolone on predator escape, boldness, and exploratory behaviors. Environ. Pollut. 2019, 245, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Porseryd, T.; Kellner, M.; Caspillo, N.R.; Volkova, K.; Elabbas, L.; Ullah, S.; Olsén, H.; Dinnétz, P.; Hällström, I.P. Combinatory effects of low concentrations of 17α-etinylestradiol and citalopram on non-reproductive behavior in adult zebrafish (Danio rerio). Aquat. Toxicol. 2017, 193, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Lamb, S.; Chia, J.; Johnson, S. Paternal exposure to a common herbicide alters the behavior and serotonergic system of zebrafish offspring. PLoS ONE 2020, 15, e0228357. [Google Scholar] [CrossRef]
- Barr, D.B.; Panuwet, P.; Nguyen, J.V.; Udunka, S.; Needham, L.L. Assessing exposure to atrazine and its metabolites using biomonitoring. Environ. Health Perspect. 2007, 115, 1474–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, K.R.; Baker, D.B.; Richards, R.P.; Dixon, K.R.; Klaine, S.J.; La Point, T.W.; Kendall, R.J.; Weisskopf, C.P.; Giddings, J.M.; Giesy, J.P.; et al. Ecological risk assessment of atrazine in North American surface waters. Environ. Toxicol. Chem. 1996, 15, 31–76. [Google Scholar] [CrossRef]
- Wirbisky, S.E.; Weber, G.J.; Sepúlveda, M.S.; Xiao, C.; Cannon, J.R.; Freeman, J.L. Developmental origins of neurotransmitter and transcriptome alterations in adult female zebrafish exposed to atrazine during embryogenesis. Toxicology 2015, 333, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Wirbisky, S.E.; Weber, G.J.; Sepúlveda, M.S.; Lin, T.-L.; Jannasch, A.S.; Freeman, J.L. An embryonic atrazine exposure results in reproductive dysfunction in adult zebrafish and morphological alterations in their offspring. Sci. Rep. 2016, 6, 21337. [Google Scholar] [CrossRef] [Green Version]
- Hayes, T.B.; Collins, A.; Lee, M.; Mendoza, M.; Noriega, N.; Stuart, A.A.; Vonk, A. Hermaphroditic, demasculinized frogs after exposure to the herbicide atrazine at low ecologically relevant doses. Proc. Natl. Acad. Sci. USA 2002, 99, 5476–5480. [Google Scholar] [CrossRef] [Green Version]
- Hayes, T.B.; Khoury, V.; Narayan, A.; Nazir, M.; Park, A.; Brown, T.; Adame, L.; Chan, E.; Buchholz, D.; Stueve, T.; et al. Atrazine induces complete feminization and chemical castration in male African clawed frogs (Xenopus laevis). Proc. Natl. Acad. Sci. USA 2010, 107, 4612–4617. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Dodd, C.A.; Filipov, N.M. Short-term atrazine exposure causes behavioral deficits and disrupts monoaminergic systems in male C57BL/6 mice. Neurotoxicol. Teratol. 2013, 39, 26–35. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Reidenbach, L.S.; Thanki, D.H.; Winchester, A.E.; Qualizza, B.A.; Ryan, G.A.; Egan, K.E.; Hedrick, V.E.; Sobreira, T.J.; Peterson, S.M.; et al. Embryonic atrazine exposure elicits proteomic, behavioral, and brain abnormalities with developmental time specific gene expression signatures. J. Proteom. 2018, 186, 71–82. [Google Scholar] [CrossRef]
- Graymore, M.; Stagnitti, F.; Allinson, G. Impacts of atrazine in aquatic ecosystems. Environ. Int. 2001, 26, 483–495. [Google Scholar] [CrossRef]
- Grube, A.; Donaldson, D.; Kiely, T.; Wu, L. Pesticides Industry Sales and Usage: 2006 and 2007 Market Estimates; Environmentla Protection Agency (EPA): Washington, DC, USA, 2011; 733-R-11-001. [Google Scholar]
- Richter, C.A.; Papoulias, D.M.; Whyte, J.J.; Tillitt, D.E. Evaluation of potential mechanisms of atrazine-induced reproductive impairment in fathead minnow (Pimephales promelas) and Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem. 2016, 35, 2230–2238. [Google Scholar] [CrossRef]
- Foradori, C.D.; Hinds, L.R.; Hanneman, W.H.; Legare, M.E.; Clay, C.M.; Handa, R.J. Atrazine inhibits pulsatile luteinizing hormone release without altering pituitary sensitivity to a gonadotropin-releasing hormone receptor agonist in female Wistar rats. Biol. Reprod. 2009, 81, 40–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foradori, C.D.; Zimmerman, A.D.; Hinds, L.R.; Zuloaga, K.L.; Breckenridge, C.B.; Handa, R.J. Atrazine inhibits pulsatile gonadotropin-releasing hormone (GnRH) release without altering GnRH messenger RNA or protein levels in the female rat. Biol. Reprod. 2013, 88, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.L.; Laws, S.C.; Das, P.C.; Narotsky, M.G.; Goldman, J.M.; Lee Tyrey, E.; Stoker, T.E. Atrazine and reproductive function: Mode and mechanism of action studies. Birth Defects Res. B Dev. Reprod Toxicol. 2007, 80, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Dodd, C.A.; Xiao, S.; Krishna, S.; Ye, X.; Filipov, N.M. Gestational and lactational exposure to atrazine via the drinking water causes specific behavioral deficits and selectively alters monoaminergic systems in C57BL/6 mouse dams, juvenile and adult offspring. Toxicol. Sci. 2014, 141, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, J.T.; Boerma, J.; Lansbergen, G.W.; van den Berg, M. Induction and inhibition of aromatase (CYP19) activity by various classes of pesticides in H295R human adrenocortical carcinoma cells. Toxicol. Appl. Pharmacol. 2002, 182, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.; Farombi, E.; Kashyap, M.; Pant, A. Atrazine induces transcriptional changes in marker genes associated with steroidogenesis in primary cultures of rat Leydig cells. Toxicol. In Vitro 2011, 25, 1588–1595. [Google Scholar] [CrossRef]
- Kucka, M.; Pogrmic-Majkic, K.; Fa, S.; Stojilkovic, S.S.; Kovacevic, R. Atrazine acts as an endocrine disrupter by inhibiting cAMP-specific phosphodiesterase-4. Toxicol. Appl. Pharmacol. 2012, 265, 19–26. [Google Scholar] [CrossRef]
- Hayes, T.B. Welcome to the revolution: Integrative biology and assessing the impact of endocrine disruptors on environmental and public health. Integr. Comp. Biol. 2005, 45, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Yanase, T.; Morinaga, H.; Gondo, S.; Okabe, T.; Nomura, M.; Komatsu, T.; Morohashi, K.I.; Hayes, T.B.; Takayanagi, R.; et al. Atrazine-induced aromatase expression is SF-1 dependent: Implications for endocrine disruption in wildlife and reproductive cancers in humans. Environ. Health Perspect. 2007, 115, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Schmidel, A.J.; Assmann, K.L.; Werlang, C.C.; Bertoncello, K.T.; Francescon, F.; Rambo, C.L.; Beltrame, G.M.; Calegari, D.; Batista, C.B.; Blaser, R.E.; et al. Subchronic atrazine exposure changes defensive behaviour profile and disrupts brain acetylcholinesterase activity of zebrafish. Neurotoxicol. Teratol. 2014, 44, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Steele, A.N.; Moore, P.A. Express yourself: Individuals with bold personalities exhibit increased behavioral sensitivity to dynamic herbicide exposure. Ecotoxicol. Environ. Saf. 2019, 179, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Bi, H.; Ma, K.; Li, B. Developmental exposure to atrazine impairs spatial memory and downregulates the hippocampal D1 dopamine receptor and cAMP-dependent signaling pathway in rats. Int. J. Mol. Sci. 2018, 19, 2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardullas, U.; Giordano, M.; Rodríguez, V.M. Chronic atrazine exposure causes disruption of the spontaneous locomotor activity and alters the striatal dopaminergic system of the male Sprague–Dawley rat. Neurotoxicol. Teratol. 2011, 33, 263–272. [Google Scholar] [CrossRef]
- MacLaren, R.D.; Rowland, W.J.; Morgan, N. Female preferences for sailfin and body size in the sailfin molly, Poecilia latipinna. Ethology 2004, 110, 363–379. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Chauhan, A.; Datta, S.; Wani, A.B.; Singh, N.; Singh, J. Toxicity, degradation and analysis of the herbicide atrazine. Environ. Chem. Lett. 2018, 16, 211–237. [Google Scholar] [CrossRef]
- Rohr, J.R.; McCoy, K.A. A qualitative meta-analysis reveals consistent effects of atrazine on freshwater fish and amphibians. Environ. Health Perspect. 2010, 118, 20–32. [Google Scholar] [CrossRef] [Green Version]
- MacLaren, R.D.; Daniska, D. Female preferences for dorsal fin and body size in Xiphophorus helleri: Further investigation of the LPA bias in Poeciliid fishes. Behaviour 2008, 145, 897–913. [Google Scholar] [CrossRef]
- MacLaren, R.D.; Fontaine, A. Female bias for male lateral projection area in Poecilia reticulata. Environ. Biol. Fishes 2012, 93, 105–119. [Google Scholar] [CrossRef]
- MacLaren, R.D.; Fontaine, A. Incongruence between the sexes in preferences for body and dorsal fin size in Xiphophorus variatus. Behav. Proc. 2013, 92, 99–106. [Google Scholar] [CrossRef]
- MacLaren, R.D.; He, R.; Gagnon, J. Bias for enlarged male body and dorsal fins in female Xiphophorus variatus. Behav. Proc. 2011, 87, 197–202. [Google Scholar] [CrossRef]
- MacLaren, R.D.; Rowland, W.J. Female preference for male lateral projection area in the shortfin molly Poecilia mexicana; evidence for a preexisting bias. Ethology 2006, 112, 678–690. [Google Scholar] [CrossRef]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maximino, C.; de Oliveira, D.L.; Rosemberg, D.B.; Batista, E.D.J.O.; Herculano, A.M.; Oliveira, K.R.M.; Benzecry, R.; Blaser, R. A comparison of the light/dark and novel tank tests in zebrafish. Behaviour 2012, 149, 1099–1123. [Google Scholar] [CrossRef]
- EPA. Decision Documents for Atrazine; U.S. Environmental Protection Agency: Washington, DC, USA, 2006.
- Rowland, W.J. Studying visual cues in fish behavior: A review of ethological techniques. Environ. Biol. Fishes 1999, 56, 285–305. [Google Scholar] [CrossRef]
- MacLaren, R.D. The effects of male proximity, apparent size, and absolute size on female preference in the sailfin molly, Poecilia latipinna. Behaviour 2006, 143, 1457–1472. [Google Scholar] [CrossRef] [Green Version]
- MacLaren, R.D. Evidence of an emerging female preference for an artificial male trait and the potential for spread via mate choice copying in Poecilia latipinna. Ethology 2019, 125, 575–586. [Google Scholar] [CrossRef]
- MacLaren, R.D. Social environment affects female preference for male body color during development in artificially selected varieties of Poecilia latipinna. Ethol. Ecol. Evol. 2017, 29, 421–435. [Google Scholar] [CrossRef]
- MacLaren, R.D. Effects of male apparent length on female preference for absolute body size in Xiphophorus helleri. Acta Ethologica 2017, 20, 27–36. [Google Scholar] [CrossRef]
- Wagner, W.E. Measuring female mating preferences. Anim. Behav. 1998, 55, 1029–1042. [Google Scholar] [CrossRef]
- Farr, J.A. Sexual selection and secondary sexual differentiation in poeciliids: Determinants of male mating success and the evolution of female choice. In Ecology and Evolution of Livebearing Fishes; Meffe, G.K., Snelson, F.F., Jr., Eds.; Prentice Hall: Englewood Cliffs, NJ, USA, 1989; pp. 91–124. [Google Scholar]
- Houde, A. Sex, Color, and Mate Choice in Guppies; Princeton University Press: Princeton, NJ, USA, 1997; 210p. [Google Scholar]
- Basolo, A.L. Congruence between the sexes in preexisting receiver responses. Behav. Ecol. 2002, 13, 832–837. [Google Scholar] [CrossRef]
- Brodeur, J.C.; Svartz, G.; Perez-Coll, C.S.; Marino, D.J.G.; Herkovits, J. Comparative susceptibility to atrazine of three developmental stages of Rhinella arenarum and influence on metamorphosis: Non-monotonous acceleration of the time to climax and delayed tail resorption. Aquat. Toxicol. 2009, 91, 161–170. [Google Scholar] [CrossRef]
- Hamlin, H.J.; Guillette, L.J. Embryos as targets of endocrine disrupting contaminants in wildlife. Birth Defects Res. C Embryo Today 2011, 93, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Welshons, W.V.; Thayer, K.A.; Judy, B.M.; Taylor, J.A.; Curran, E.M.; Vom Saal, F.S. Large effects from small exposures. I. Mechanisms for endocrine-disrupting chemicals with estrogenic activity. Environ. Health Perspect. 2003, 111, 994–1006. [Google Scholar] [CrossRef] [Green Version]
- Brodeur, J.C.; Sassone, A.; Hermida, G.N.; Codugnello, N. Environmentally-relevant concentrations of atrazine induce non-monotonic acceleration of developmental rate and increased size at metamorphosis in Rhinella arenarum tadpoles. Ecotoxicol. Environ. Saf. 2013, 92, 10–17. [Google Scholar] [CrossRef]
- Marcus, S.R.; Fiumera, A.C. Atrazine exposure affects longevity, development time and body size in Drosophila melanogaster. J. Insect. Physiol. 2016, 91, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCallum, M.L.; Matlock, M.; Treas, J.; Safi, B.; Sanson, W.; McCallum, J.L. Endocrine disruption of sexual selection by an estrogenic herbicide in the mealworm beetle (Tenebrio molitor). Ecotoxicology 2013, 22, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Riffle, B.W.; Klinefelter, G.R.; Cooper, R.L.; Winnik, W.M.; Swank, A.; Jayaraman, S.; Suarez, J.; Best, D.; Laws, S. Novel molecular events associated with altered steroidogenesis induced by exposure to atrazine in the intact and castrate male rat. Reprod. Toxicol. 2014, 47, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Way, G.P.; Ruhl, N.; Snekser, J.L.; Kiesel, A.L.; McRobert, S.P. A comparison of methodologies to test aggression in zebrafish. Zebrafish 2015, 12, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Ariyomo, T.O.; Watt, P.J. The effect of variation in boldness and aggressiveness on the reproductive success of zebrafish. Anim. Behav. 2012, 83, 41–46. [Google Scholar] [CrossRef]
- MacLaren, R.D.; Wisniewski, K.; MacLaren, C. Environmental concentrations of metformin exposure affect aggressive behavior in the Siamese fighting fish, Betta splendens. PLoS ONE 2018, 13, e0197259. [Google Scholar] [CrossRef] [Green Version]
- Maximino, C.; de Brito, T.M.; da Silva Batista, A.W.; Herculano, A.M.; Morato, S.; Gouveia, A., Jr. Measuring anxiety in zebrafish: A critical review. Behav. Brain Res. 2010, 214, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, C.E.; Lorenz, R.; Spieser, O.H. Effects of atrazine on swimming behavior of zebrafish, Brachydanio rerio. Water Res. 1995, 29, 981–985. [Google Scholar] [CrossRef]
- Volkova, K.; Caspillo, N.R.; Porseryd, T.; Hallgren, S.; Dinnetz, P.; Olsén, H.; Hällström, I.P. Transgenerational effects of 17α-ethinyl estradiol on anxiety behavior in the guppy, (Poecilia reticulata). Gen. Comp. Endocrinol. 2015, 223, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Pink, M.; Abrahams, M.V. In shallow water ecosytems the abiotic environment is more important than prey abundance for foraging terns. Environ. Biol. Fishes 2018, 101, 355–362. [Google Scholar] [CrossRef]
- Rypel, A.L.; Layman, C.A.; Arrington, D.A. Water depth modifies relative predation risk for a motile fish taxon in Bahamian tidal creeks. Estuaries Coasts 2007, 30, 518–525. [Google Scholar] [CrossRef]
- Werner, E.E.; Gilliam, J.F.; Hall, D.J.; Mittelbach, G.G. An experimental test of the effects of predation risk on habitat use in fish. Ecology 1983, 64, 1540–1548. [Google Scholar] [CrossRef]
- Luca, R.M.; Gerlai, R. Animated bird silhouette above the tank: Acute alcohol diminishes fear responses in zebrafish. Behav. Brain Res. 2012, 229, 194–201. [Google Scholar] [CrossRef]
- Pitcher, T.; Lang, S.; Turner, J. A risk-balancing trade off between foraging rewards and predation hazard in a shoaling fish. Behav. Ecol. Sociobiol. 1988, 22, 225–228. [Google Scholar] [CrossRef]
- MacPherson, B.; Mashayekhi, M.; Gras, R.; Scott, R. Exploring the connection between emergent animal personality and fitness using a novel individual-based model and decision tree approach. Ecol. Inform. 2017, 40, 81–92. [Google Scholar] [CrossRef]
- Smith, B.R.; Blumstein, D.T. Fitness consequences of personality: A meta-analysis. Behav. Ecol. 2008, 19, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Biro, P.A.; Stamps, J.A. Are animal personality traits linked to life-history productivity? Trends Ecol. Evol. 2008, 23, 361–368. [Google Scholar] [CrossRef]
- Wolf, M.; Van Doorn, G.S.; Leimar, O.; Weissing, F.J. Life-history trade-offs favour the evolution of animal personalities. Nature 2007, 447, 581. [Google Scholar] [CrossRef]
- Peterson, E.K.; Buchwalter, D.B.; Kerby, J.L.; LeFauve, M.K.; Varian-Ramos, C.W.; Swaddle, J.P. Integrative behavioral ecotoxicology: Bringing together fields to establish new insight to behavioral ecology, toxicology, and conservation. Curr. Zool. 2017, 63, 185–194. [Google Scholar] [CrossRef] [Green Version]
- EPA. Persistent Organic Pollutants: A Global Issue, A Global Response; United States Environmental protection Agency: Washington, DC, USA, 2009.
- Hayes, T.B.; Stuart, A.A.; Mendoza, M.; Collins, A.; Noriega, N.; Vonk, A.; Johnston, G.; Liu, R.; Kpodzo, D. Characterization of atrazine-induced gonadal malformations in African clawed frogs (Xenopus laevis) and comparisons with effects of an androgen antagonist (cyproterone acetate) and exogenous estrogen (17 b-estradiol): Support for the demasculinization/feminization hypothesis. Environ. Health Perspect. 2006, 114, 134–141. [Google Scholar]
- Bartke, A.; Williams, K.I.H.; Dalterio, S. Effects of estrogens on testicular testosterone production in vitro. Biol. Reprod. 1977, 17, 645–649. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Kim, J.H.; Lee, H.J.; Yoon, Y.D. Octylphenol reduces the expressions of steroidogenic enzymes and testosterone production in mouse testis. Environ. Toxicol. 2007, 22, 449–458. [Google Scholar] [CrossRef]
- Ankley, G.T.; Kahl, M.D.; Jensen, K.M.M.; Hornung, M.W.; Korte, J.J.; Makynen, E.; Leino, R. Evaluation of the aromatase inhibitor fadrozole in a short-term reproduction assay with the fathead minnow (Pimephales promelas). Toxicol. Sci. 2002, 67, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Ankley, G.T.; Jensen, K.M.; Durhan, E.J.; Makynen, E.A.; Butterworth, B.C.; Kahl, M.; Villeneuve, D.; Linnum, A.; Gray, E.; Cardon, M.; et al. Effects of two fungicides with multiple modes of action on reproductive endocrine function in the fathead minnow (Pimephales promelas). Toxicol. Sci. 2005, 86, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Toft, G.; Guillette, L.J. Decreased sperm count and sexual behavior in mosquitofish exposed to water from a pesticide-contaminated lake. Ecotoxicol. Environ. Saf. 2005, 60, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Baatrup, E.; Junge, M. Antiandrogenic pesticides disrupt sexual characteristics in the adult male guppy (Poecilia reticulata). Environ. Health Perspect. 2001, 109, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Bayley, M.; Junge, M.; Baatrup, E. Exposure of juvenile guppies to three antiandrogens causes demasculinization and a reduced sperm count in adult males. Aquat. Toxicol. 2002, 56, 227–239. [Google Scholar] [CrossRef]
- Thompson, C.J.; Ross, S.M.; Gaido, K.W. Di(n-butyl)phthalate impairs cholesterol transport and steroidogenesis in the fetal rat testis through a rapid and reversible mechanism. Endocrinology 2004, 145, 1227–1273. [Google Scholar] [CrossRef] [Green Version]
- Gabor, C.R.; Grober, M.S. A potential role of male and female androgen in species recognition in a unisexual–bisexual mating complex. Horm. Behav. 2010, 57, 427–433. [Google Scholar] [CrossRef]
- Friedman, A.S. Atrazine inhibition of testosterone production in rat males following peripubertal exposure. Reprod. Toxicol. 2002, 16, 275–279. [Google Scholar] [CrossRef]
- Babic-Gojmerac, T.; Kniewald, Z.; Kniewald, J. Testosterone metabolism in neuroendocrine organs in male rats under atrazine and deethylatrazine influence. J. Steroid Biochem. 1989, 33, 141–146. [Google Scholar] [CrossRef]
- Kniewald, J.; Mildner, P.; Kniewald, Z. Effects of s-triazine herbicides on hormonereceptor complex formation, 5 [alpha]-reductase and 3 [alpha]-hydroxysteroid dehydrogenase activity at the anterior pituitary level. J. Steroid Biochem. 1979, 11, 833–838. [Google Scholar] [CrossRef]
- Bischoff, R.J.; Gould, J.L.; Rubenstein, D.I. Tail size and female choice in the guppy (Poecilia reticulata). Behav. Ecol. Sociobiol. 1985, 17, 253–255. [Google Scholar] [CrossRef]
- Kodric-Brown, A. Female choice of multiple male criteria in guppies: Interacting effects of dominance, coloration and courtship. Behav. Ecol. Sociobiol. 1993, 32, 415–420. [Google Scholar] [CrossRef]
- Forsgren, E. Predation risk affects mate choice in a gobiid fish. Am. Nat. 1992, 140, 1041–1049. [Google Scholar] [CrossRef]
- Berglund, A. Risky sex: Male pipefishes mate at random in the presence of a predator. Anim. Behav. 1993, 46, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Gabor, C. Association patterns of sailfin mollies (Poecilia latipinna): Alternative hypotheses. Behav. Ecol. Sociobiol. 1999, 46, 333–340. [Google Scholar] [CrossRef]
- Ptacek, M.B.; Travis, J. Mate choice in the sailfin molly, Poecilia latipinna. Evolution 1997, 51, 1217–1231. [Google Scholar] [CrossRef]
- Schlupp, I.; Marler, C.; Ryan, M.J. Benefit to male sailfin mollies of mating with heterospecific females. Science 1994, 263, 373–374. [Google Scholar] [CrossRef]
Figure 1.
Dummy fish stimuli. (Left) Images used to create the 60 and 50 mm in total length male and female dummy stimuli used in all behavioral assays for expts. I and II; (Right) Images used to create the 40, 60, and 80 mm in total length male dummies used in all behavioral assays for expt. III.
Figure 1.
Dummy fish stimuli. (Left) Images used to create the 60 and 50 mm in total length male and female dummy stimuli used in all behavioral assays for expts. I and II; (Right) Images used to create the 40, 60, and 80 mm in total length male dummies used in all behavioral assays for expt. III.

Figure 2.
Diagram of apparatus and arenas used for all experiments. (A) Apparatus and arena used for expts. I-III: The subject was placed between a pair of aquaria each housing an adjustable motorized belt and pulley system. The dummy stimuli were attached to the pulley systems situated in the aquaria flanking the central test arena for all behavioral assays. When the apparatus was turned on, the dummy fish began ‘pacing’ back and forth along the glass facing the test arena. An iPad was used to record the subject’s behavior; (B) Arena set up used for expt. IV. Note: Dimensions of the test arena were the same for all experiments.
Figure 2.
Diagram of apparatus and arenas used for all experiments. (A) Apparatus and arena used for expts. I-III: The subject was placed between a pair of aquaria each housing an adjustable motorized belt and pulley system. The dummy stimuli were attached to the pulley systems situated in the aquaria flanking the central test arena for all behavioral assays. When the apparatus was turned on, the dummy fish began ‘pacing’ back and forth along the glass facing the test arena. An iPad was used to record the subject’s behavior; (B) Arena set up used for expt. IV. Note: Dimensions of the test arena were the same for all experiments.
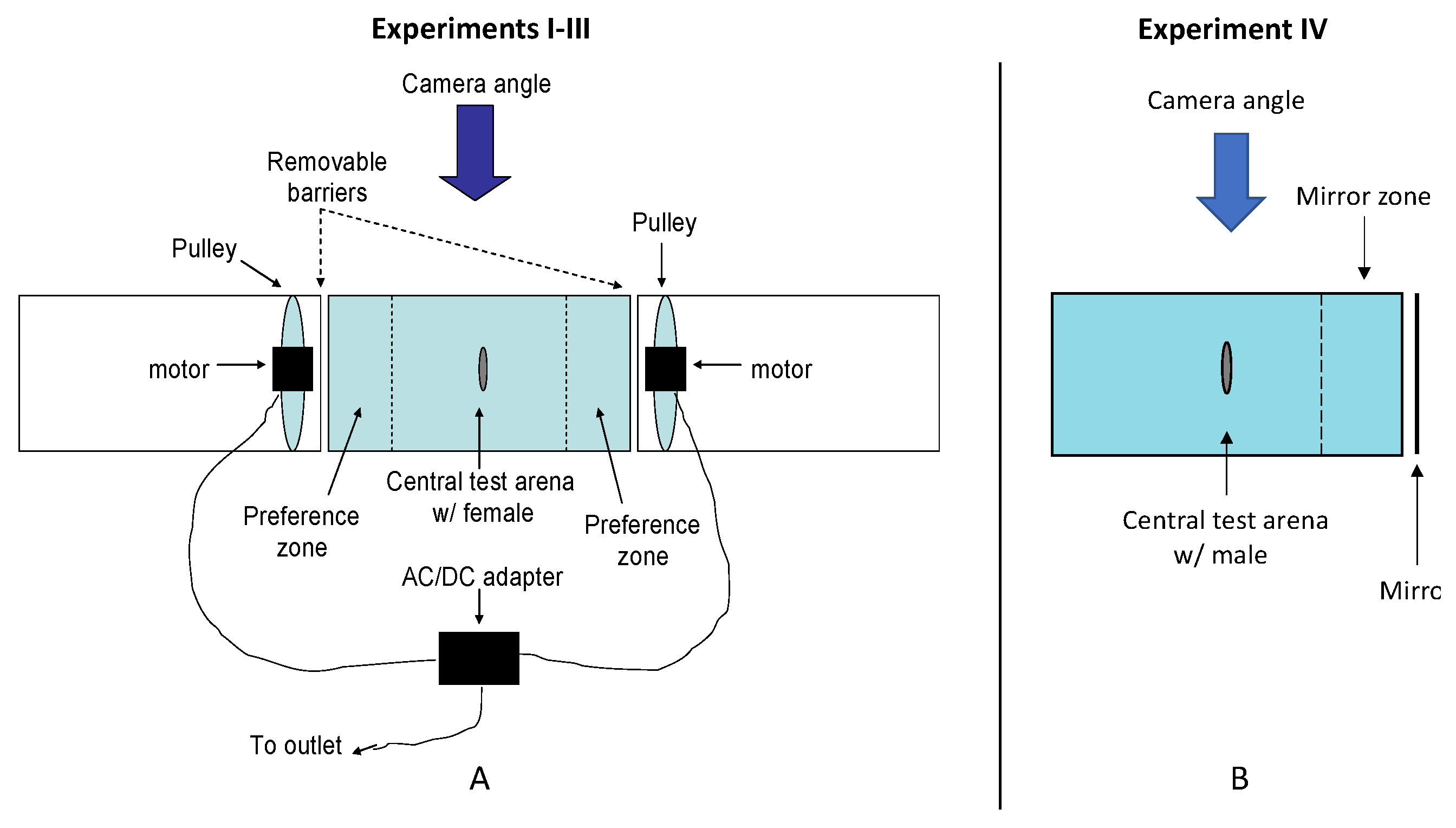
Figure 3.
Comparison of mean time spent in association with the male vs. female dummy stimuli in AZT-exposed vs. unexposed females (A) and males (B) from expts. I and II, respectively. ‘HD’ and ‘LD’ refer to subjects from the high- and low-dose exposure groups, respectively. Starred brackets (*) indicate a significant difference in mean time spent with the male vs. female dummy stimulus within a given treatment group (e.g., time LD females spent with the male vs. female dummy) using paired sample t-test.
Figure 3.
Comparison of mean time spent in association with the male vs. female dummy stimuli in AZT-exposed vs. unexposed females (A) and males (B) from expts. I and II, respectively. ‘HD’ and ‘LD’ refer to subjects from the high- and low-dose exposure groups, respectively. Starred brackets (*) indicate a significant difference in mean time spent with the male vs. female dummy stimulus within a given treatment group (e.g., time LD females spent with the male vs. female dummy) using paired sample t-test.

Figure 4.
Comparison of mean time low dose, high dose, and control females from expt. I (A) and males from expt. II (B) spent with both stimuli combined. Starred brackets (*) indicate a significant difference between the treatment groups in mean time spent with both dummies combined (paired samples t-tests).
Figure 4.
Comparison of mean time low dose, high dose, and control females from expt. I (A) and males from expt. II (B) spent with both stimuli combined. Starred brackets (*) indicate a significant difference between the treatment groups in mean time spent with both dummies combined (paired samples t-tests).
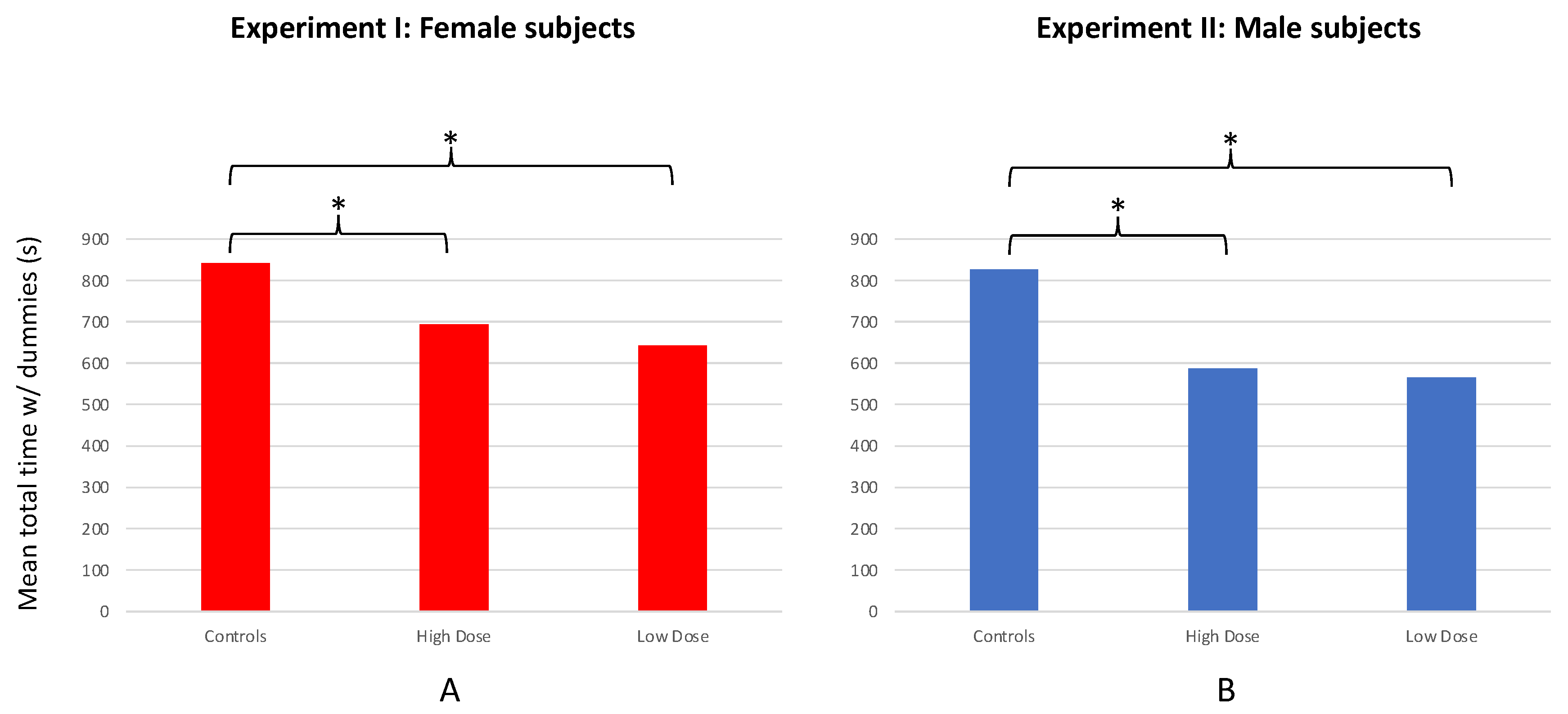
Figure 5.
Comparison of mean female strength of preference (SOP) for larger male body size among the low-dose (red), high-dose (blue), and control (green) groups of expt. III. SOP is defined as the time spent with the larger male—time spent with the smaller male per trial (A) = ‘I vs. S’ pairing; (B) = ‘L vs. I’ pairing; (C) = ‘L vs. S’ pairing). # Indicates a significant preference for the larger of the paired males in a given trial (e.g., L vs. S) using paired sample t-tests. Starred brackets (*) indicate a significant difference in mean SOP between the treatment groups (e.g., control vs. LD).
Figure 5.
Comparison of mean female strength of preference (SOP) for larger male body size among the low-dose (red), high-dose (blue), and control (green) groups of expt. III. SOP is defined as the time spent with the larger male—time spent with the smaller male per trial (A) = ‘I vs. S’ pairing; (B) = ‘L vs. I’ pairing; (C) = ‘L vs. S’ pairing). # Indicates a significant preference for the larger of the paired males in a given trial (e.g., L vs. S) using paired sample t-tests. Starred brackets (*) indicate a significant difference in mean SOP between the treatment groups (e.g., control vs. LD).
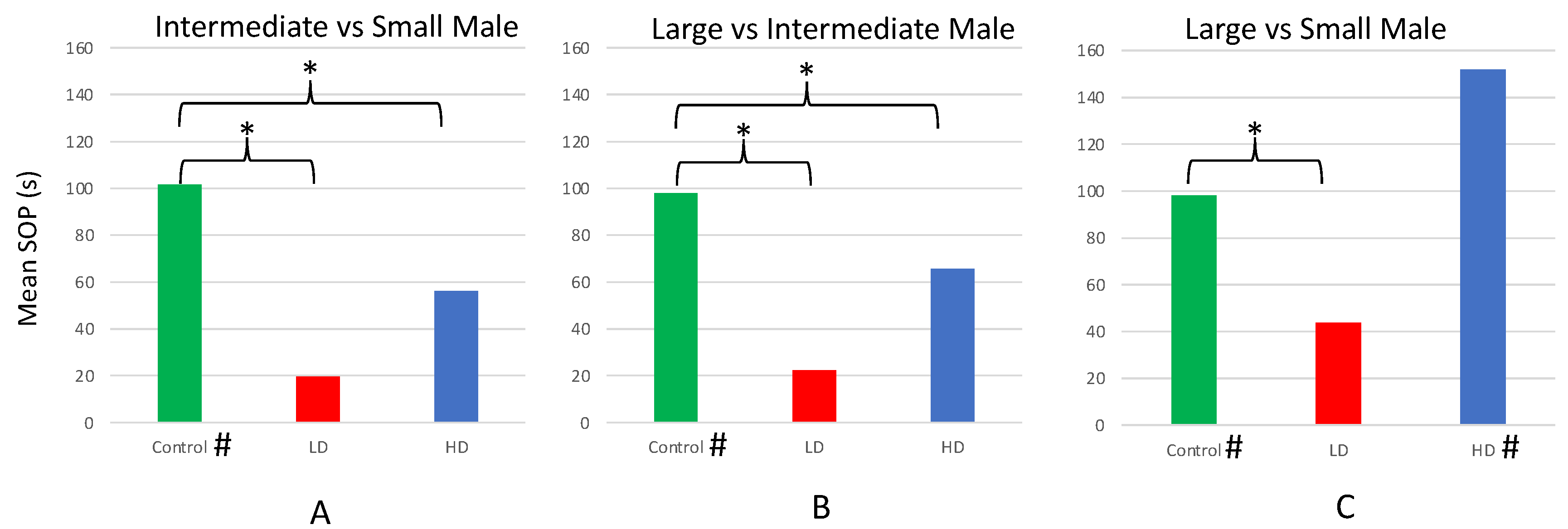
Figure 6.
Comparison of mean time spent interacting with the mirror among the LD, HD, and control males of expt. IV. Starred brackets (*) indicates a significant difference in mean mirror time between the treatment groups.
Figure 6.
Comparison of mean time spent interacting with the mirror among the LD, HD, and control males of expt. IV. Starred brackets (*) indicates a significant difference in mean mirror time between the treatment groups.
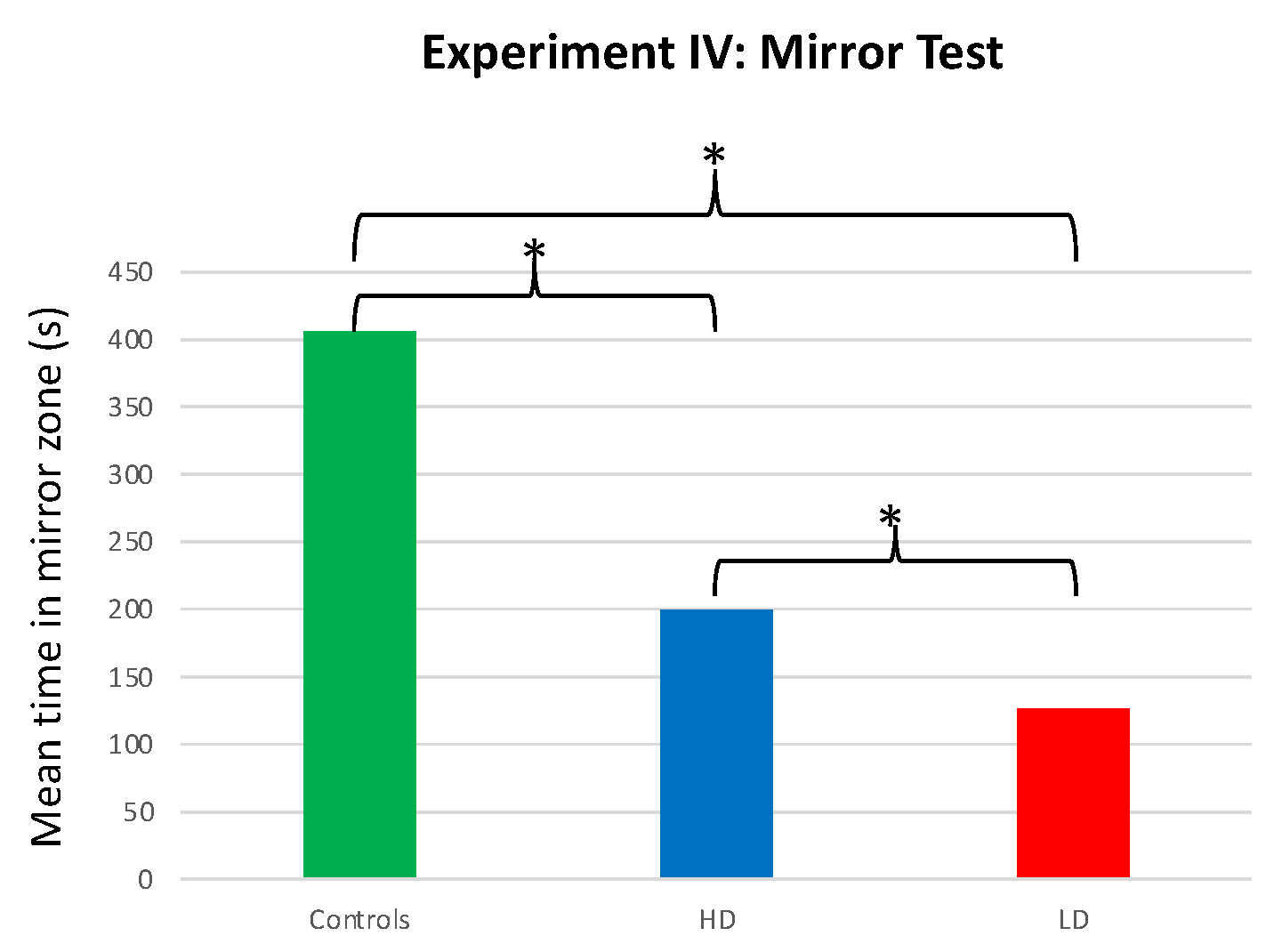
Figure 7.
Comparison of (A) time spent in the bottom zone (BZT) and (B) latency to enter the top zone (LTZ) among the LD, HD, and control females (red) and males (blue) from expts. I-IV combined. Starred brackets (*) indicate a significant difference between the treatment groups (e.g., control females vs. LD females).
Figure 7.
Comparison of (A) time spent in the bottom zone (BZT) and (B) latency to enter the top zone (LTZ) among the LD, HD, and control females (red) and males (blue) from expts. I-IV combined. Starred brackets (*) indicate a significant difference between the treatment groups (e.g., control females vs. LD females).
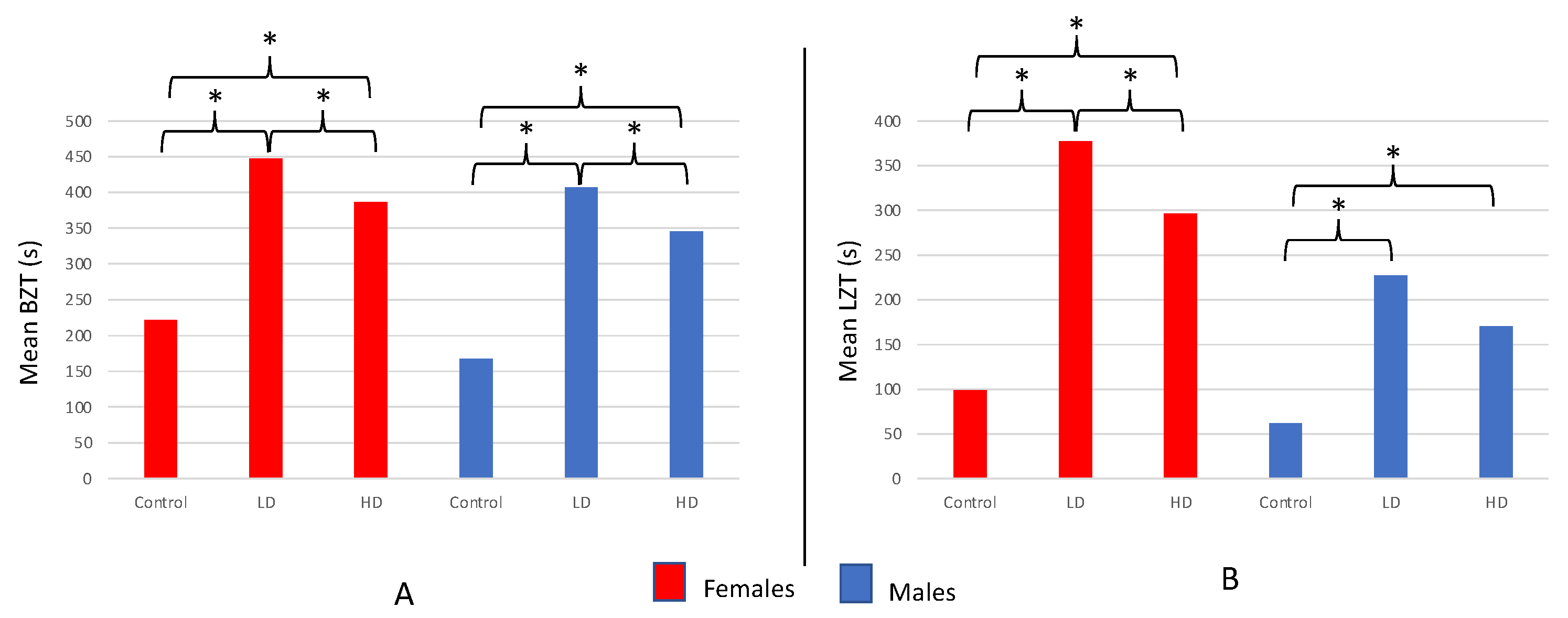

{kind=link}
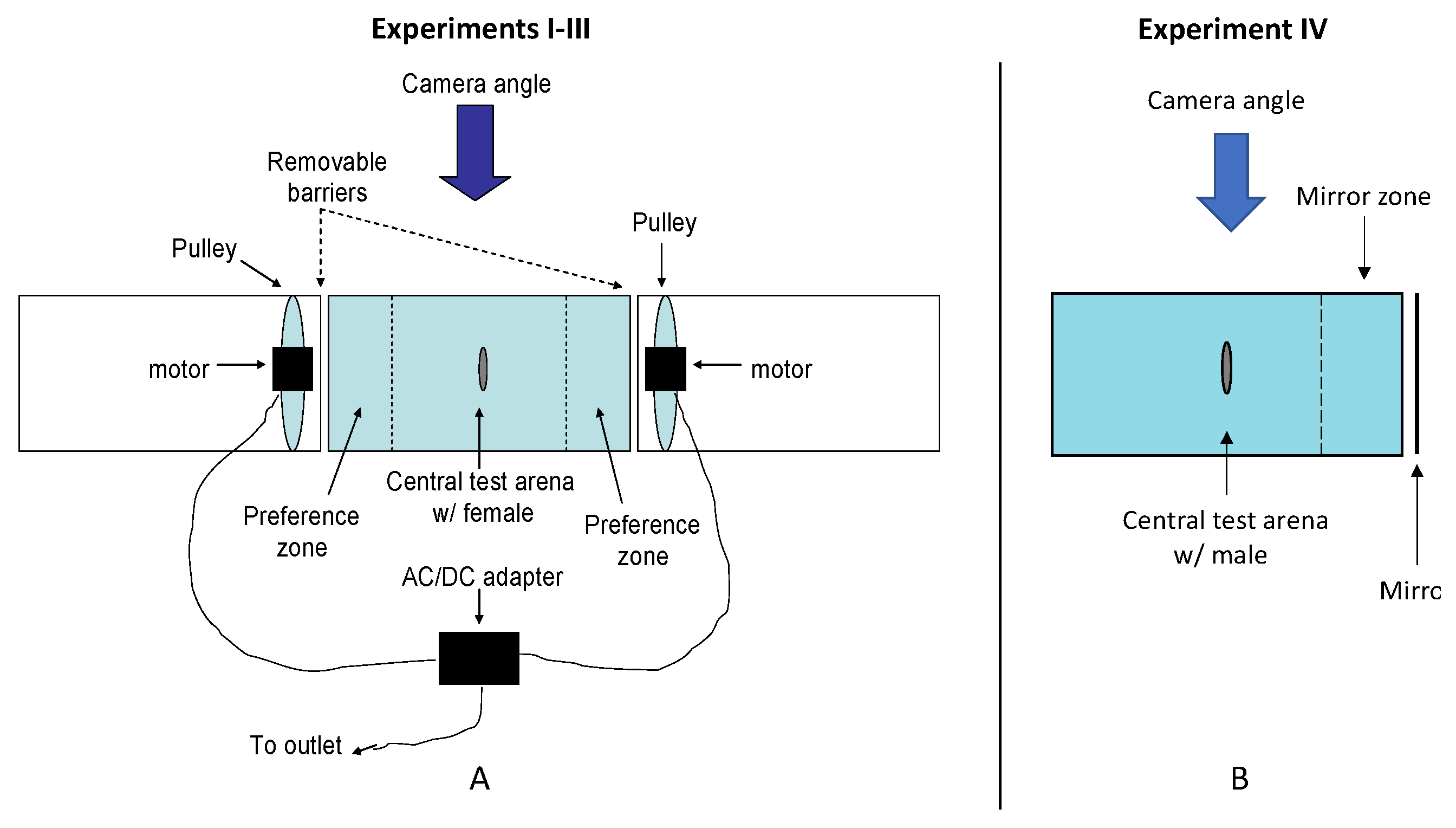{kind=link}
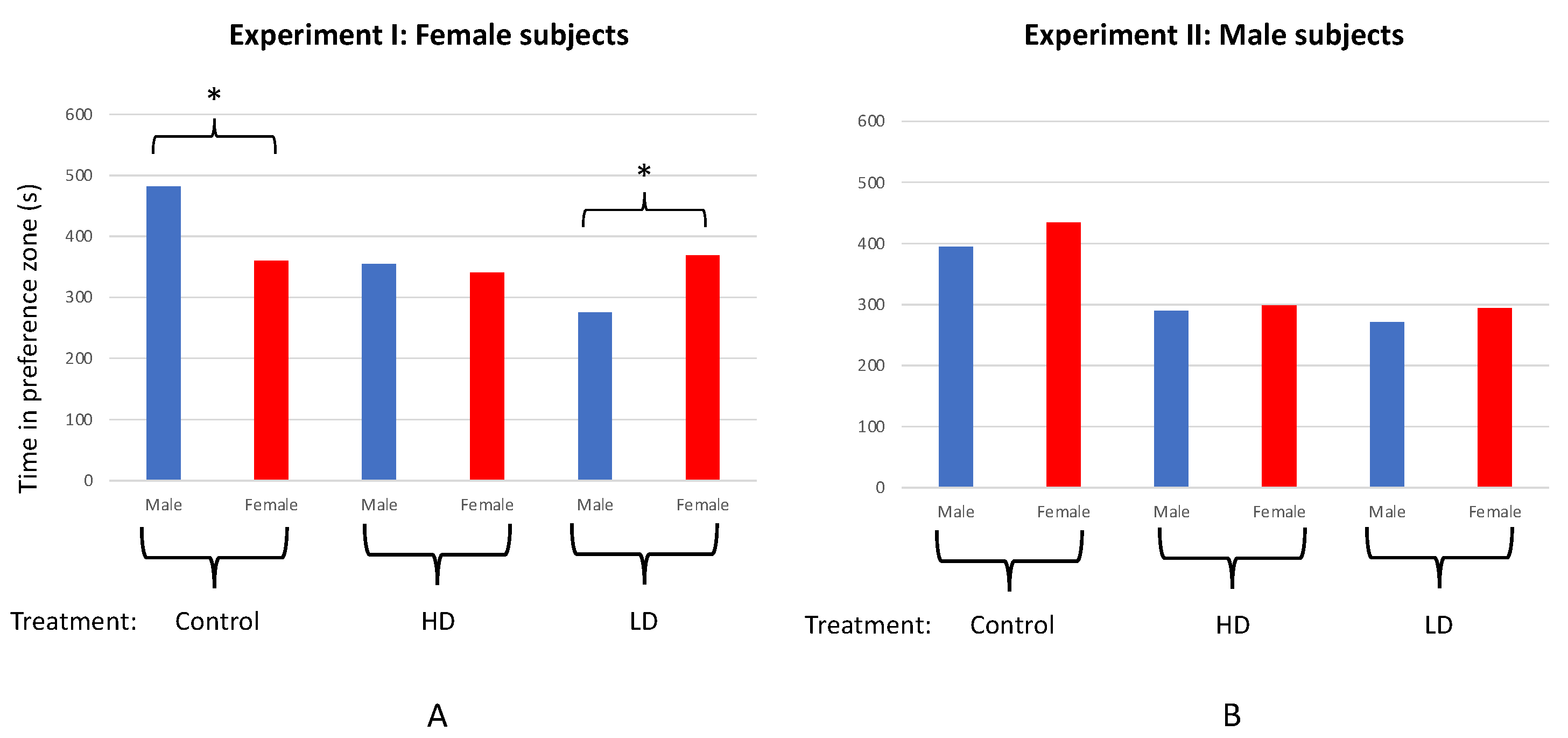{kind=link}
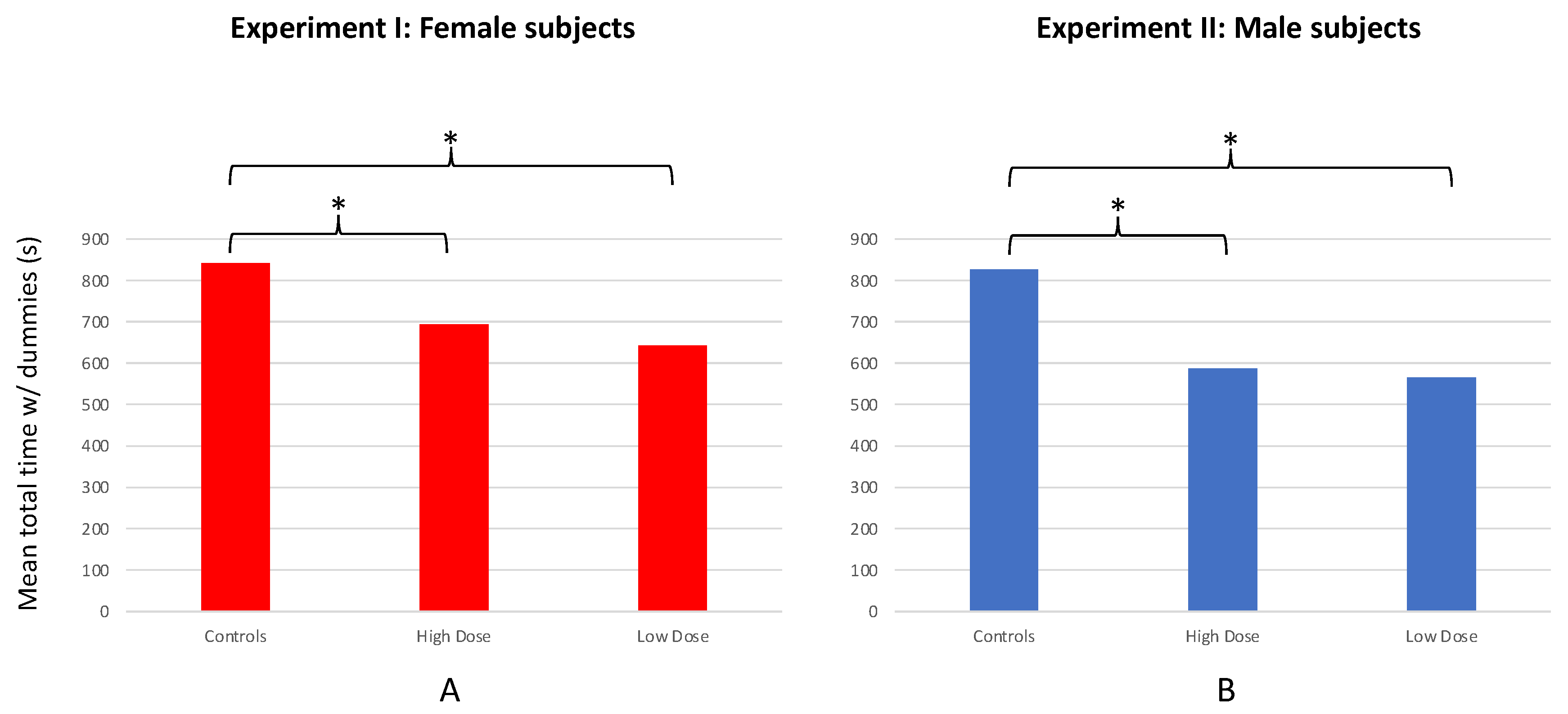{kind=link}
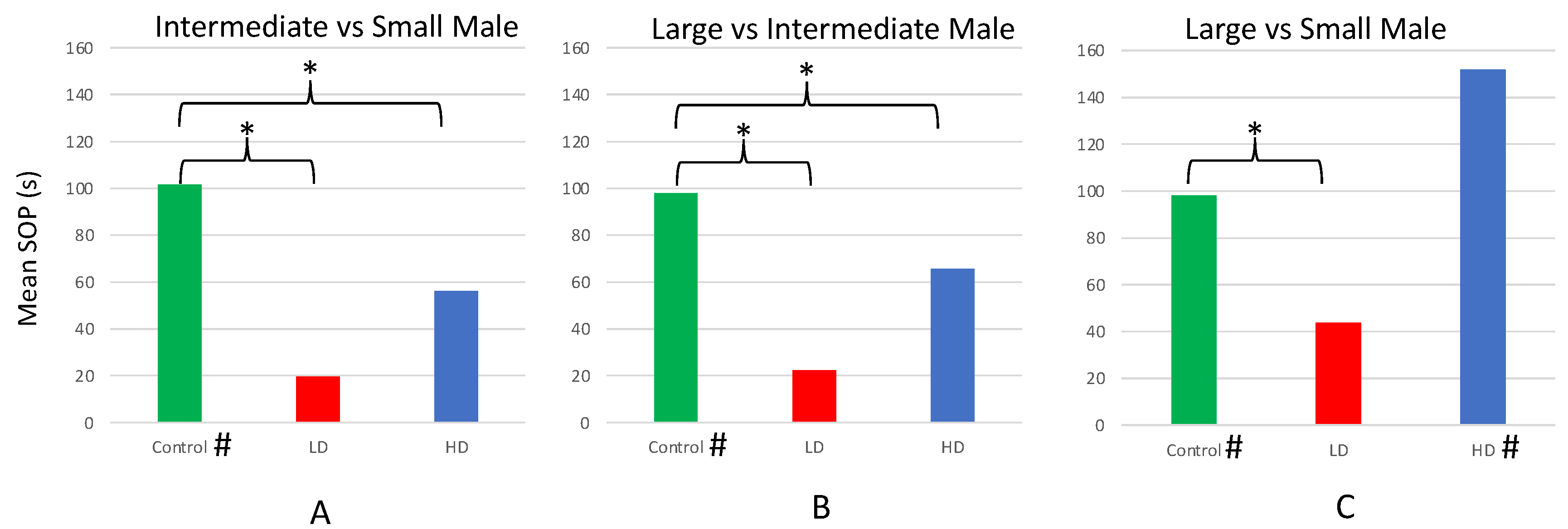{kind=link}
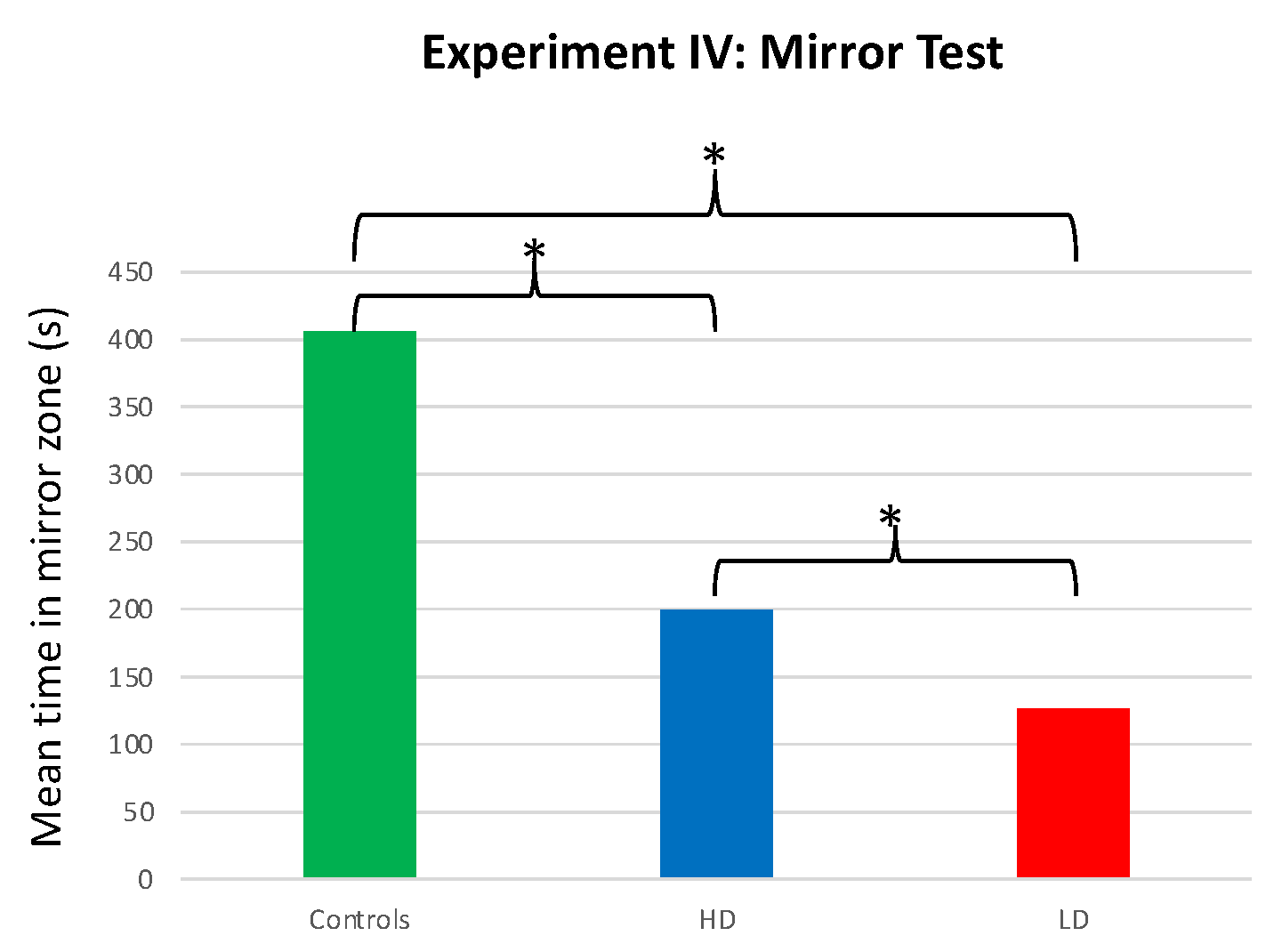{kind=link}
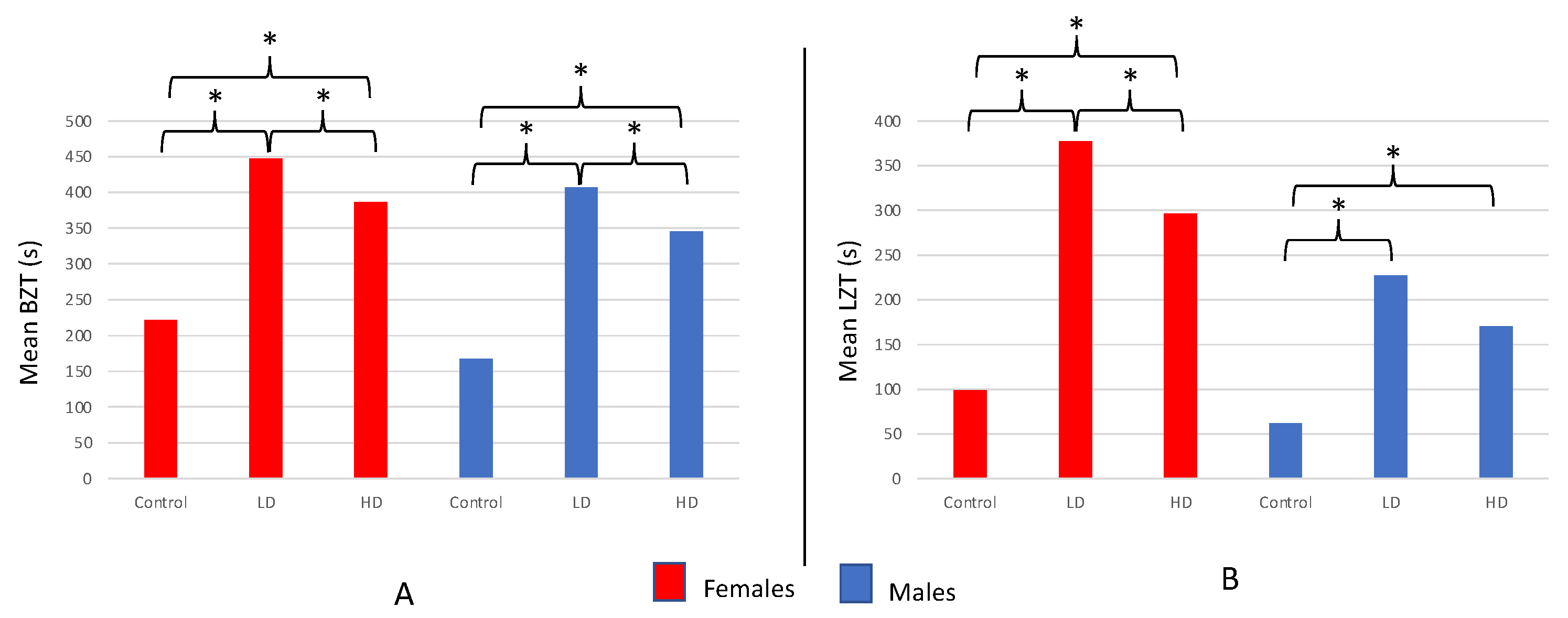{kind=link}
Table 1.
Results of paired sample t-tests comparing the mean time spent in association with the larger- versus smaller-male dummy for the LD, HD, and control groups of expt. III. ‘S’, ‘I’, and ‘L’ refer to the small, intermediate, and large male dummy stimuli, respectively.
Table 1.
Results of paired sample t-tests comparing the mean time spent in association with the larger- versus smaller-male dummy for the LD, HD, and control groups of expt. III. ‘S’, ‘I’, and ‘L’ refer to the small, intermediate, and large male dummy stimuli, respectively.
| Treatment | Trial | Time w/ Smaller Male (s) | Time w/ Larger Male (s) | T | df | p |
|---|---|---|---|---|---|---|
| Control * | S vs. I | 144.69 ± 15.08 | 246.38 ± 17.90 | 3.258 | 25 | 0.003 * |
| Control * | S vs. L | 132.23 ± 9.71 | 230.46 ± 11.70 | 5.171 | 25 | <0.001 * |
| Control * | I vs. L | 143.00 ± 12.03 | 240.92 ± 14.69 | 3.893 | 25 | <0.001 * |
| LD | S vs. I | 187.06 ± 34.86 | 206.72 ± 35.54 | 0.293 | 17 | 0.773 |
| LD | S vs. L | 140.28 ± 35.52 | 183.94 ± 34.71 | 0.678 | 17 | 0.507 |
| LD | I vs. L | 143.33 ± 28.15 | 165.78 ± 32.59 | 0.405 | 17 | 0.691 |
| HD | S vs. I | 193.80 ± 31.66 | 249.10 ± 33.16 | 0.896 | 19 | 0.382 |
| HD * | S vs. L | 132.25 ± 18.06 | 284.45 ± 29.23 | 3.451 | 19 | 0.003 * |
| HD | I vs. L | 178.00 ± 28.30 | 227.85 ± 33.18 | 0.846 | 19 | 0.408 |
Note: * Indicates a significant preference for the larger of the paired males.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
MacLaren, R.D. Environmentally Realistic Waterborne Atrazine Exposure Affects Behavior in Poecilia latipinna. Water 2023, 15, 306. https://doi.org/10.3390/w15020306
AMA Style
MacLaren RD. Environmentally Realistic Waterborne Atrazine Exposure Affects Behavior in Poecilia latipinna. Water. 2023; 15(2):306. https://doi.org/10.3390/w15020306
Chicago/Turabian StyleMacLaren, R. David. 2023. "Environmentally Realistic Waterborne Atrazine Exposure Affects Behavior in Poecilia latipinna" Water 15, no. 2: 306. https://doi.org/10.3390/w15020306
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.