
Mesocosm Design and Implementation of Two Synchronized Case Study Experiments to Determine the Impacts of Salinization and Climate Change on the Structure and Functioning of Shallow Lakes
 , , , , , , , , , , , add
Show full author list
, , , , , , , , , , , add
Show full author list
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Mesocosm Location and Infrastructure
2.2. Mesocosm Ecosystems and Inoculation of Aquatic Communities
2.3. Experiment 1
2.4. Experiment 2
2.5. Sampling and Analyses
2.6. Statistical Analyses
3. Results and Discussion
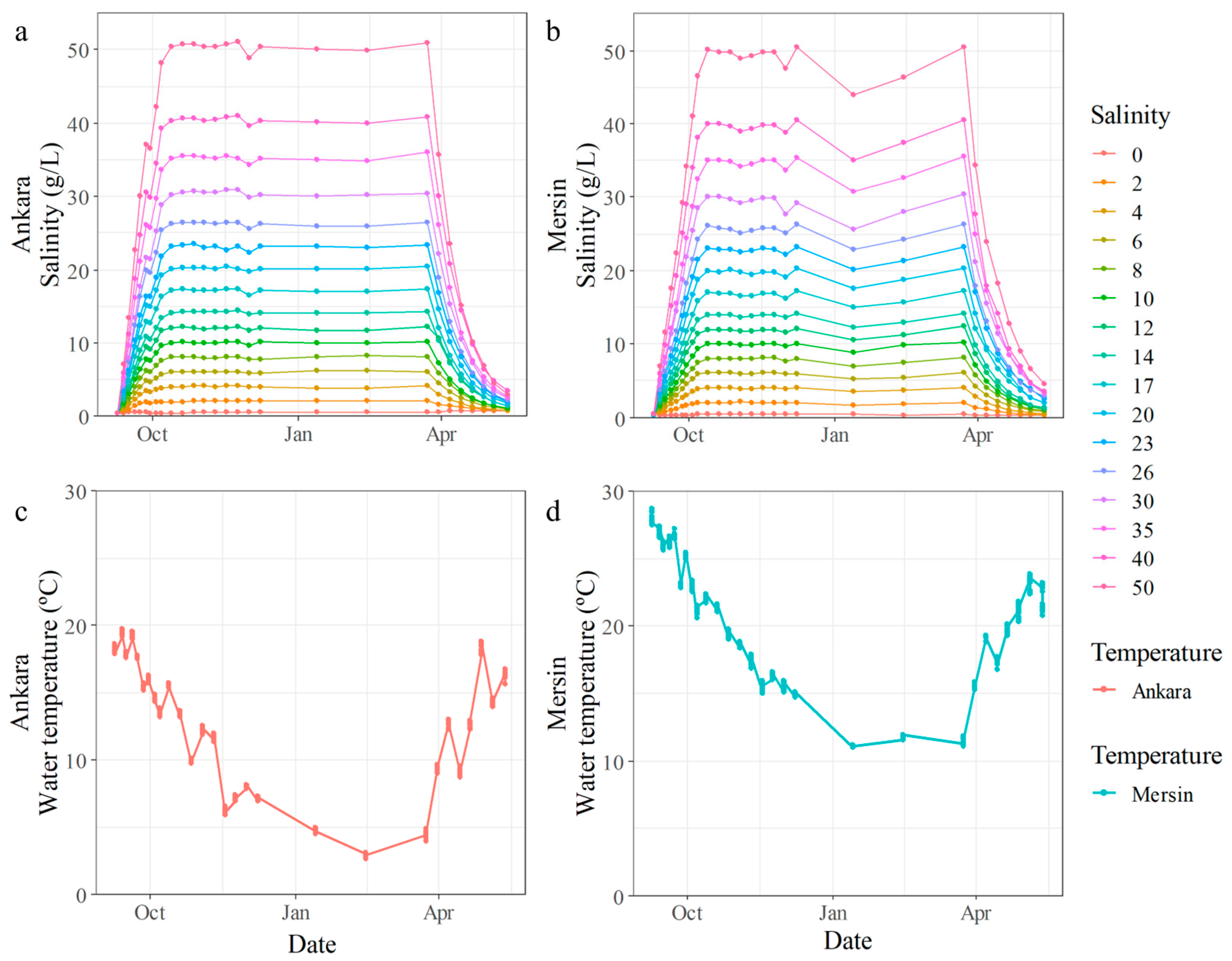
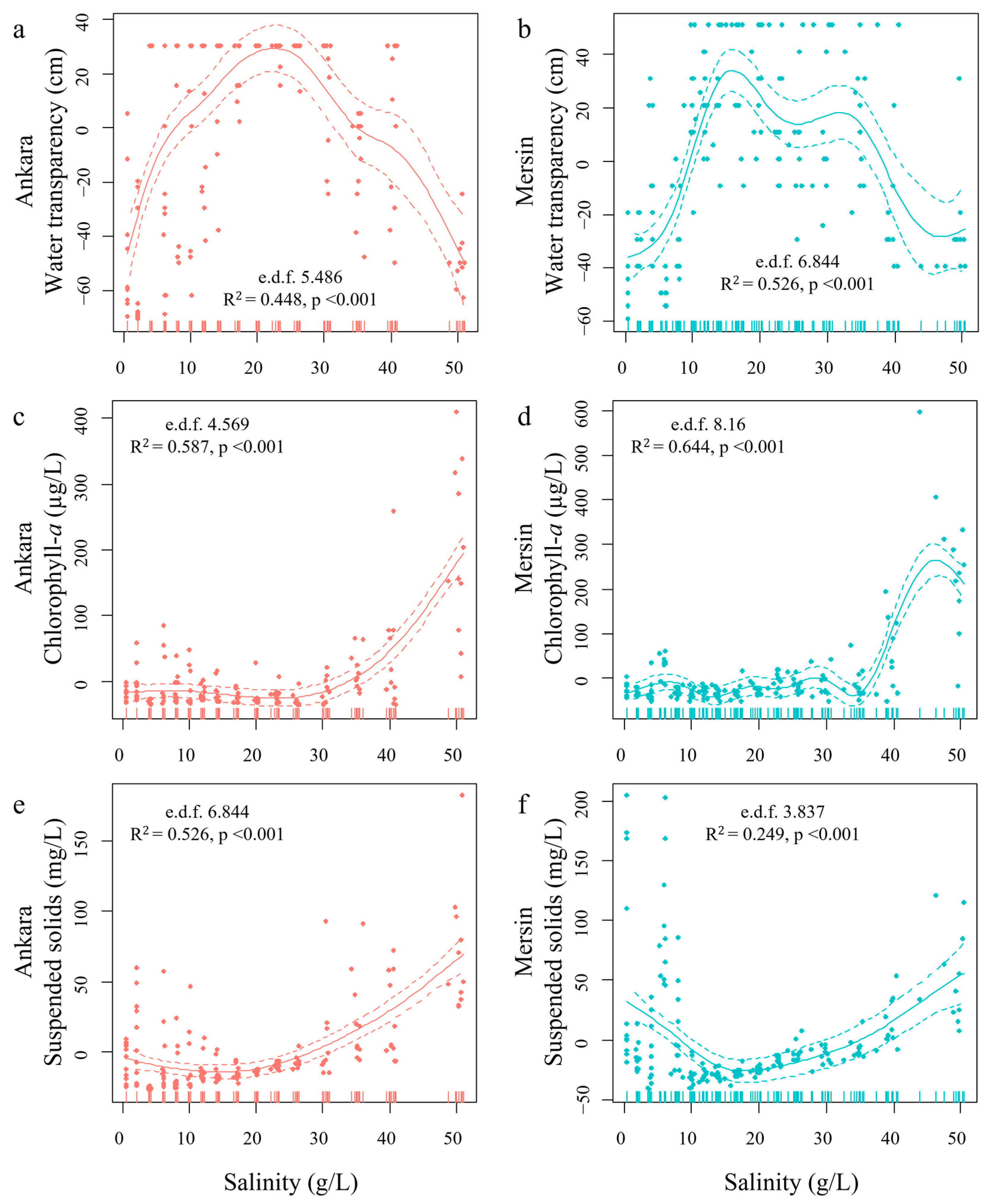
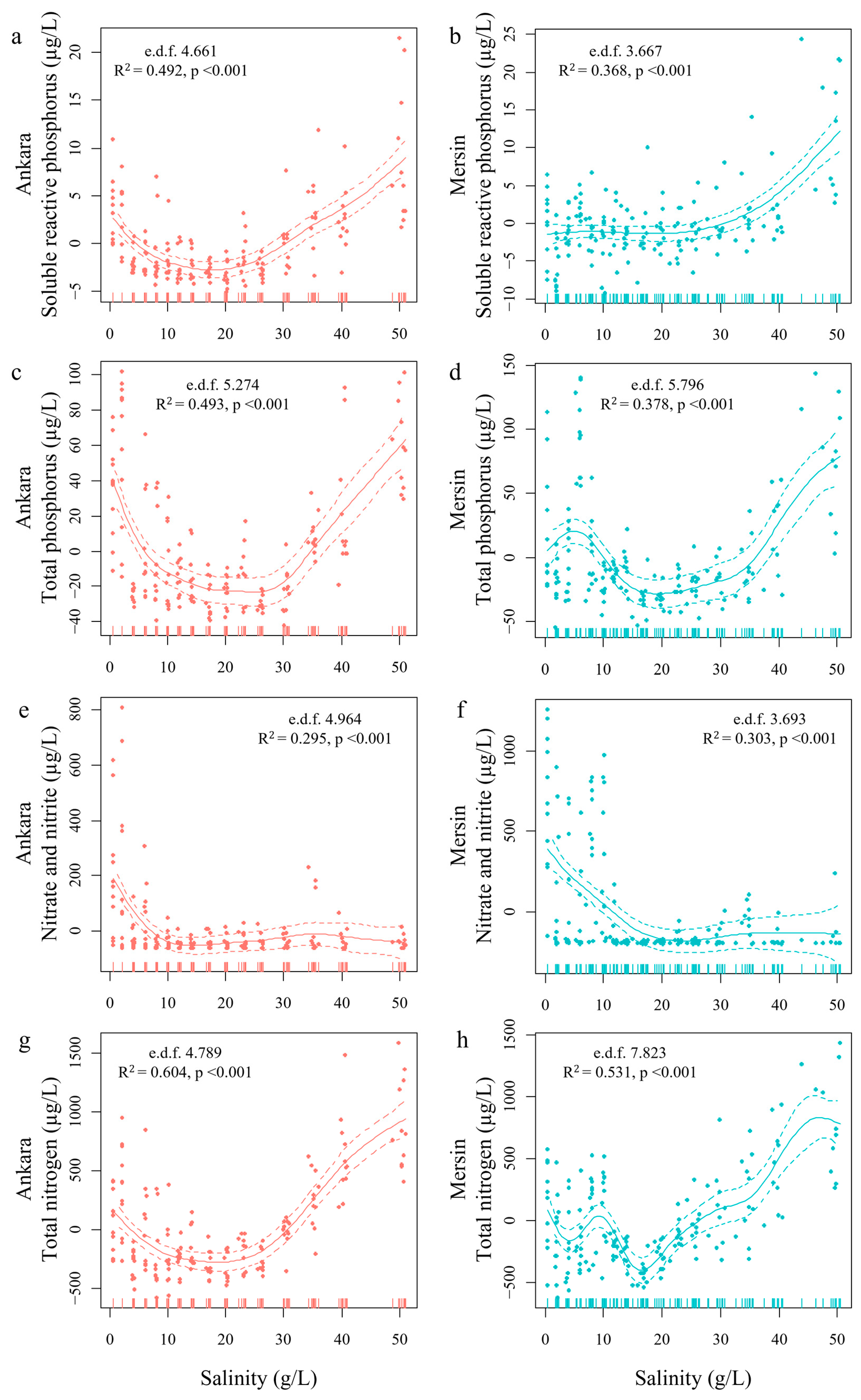
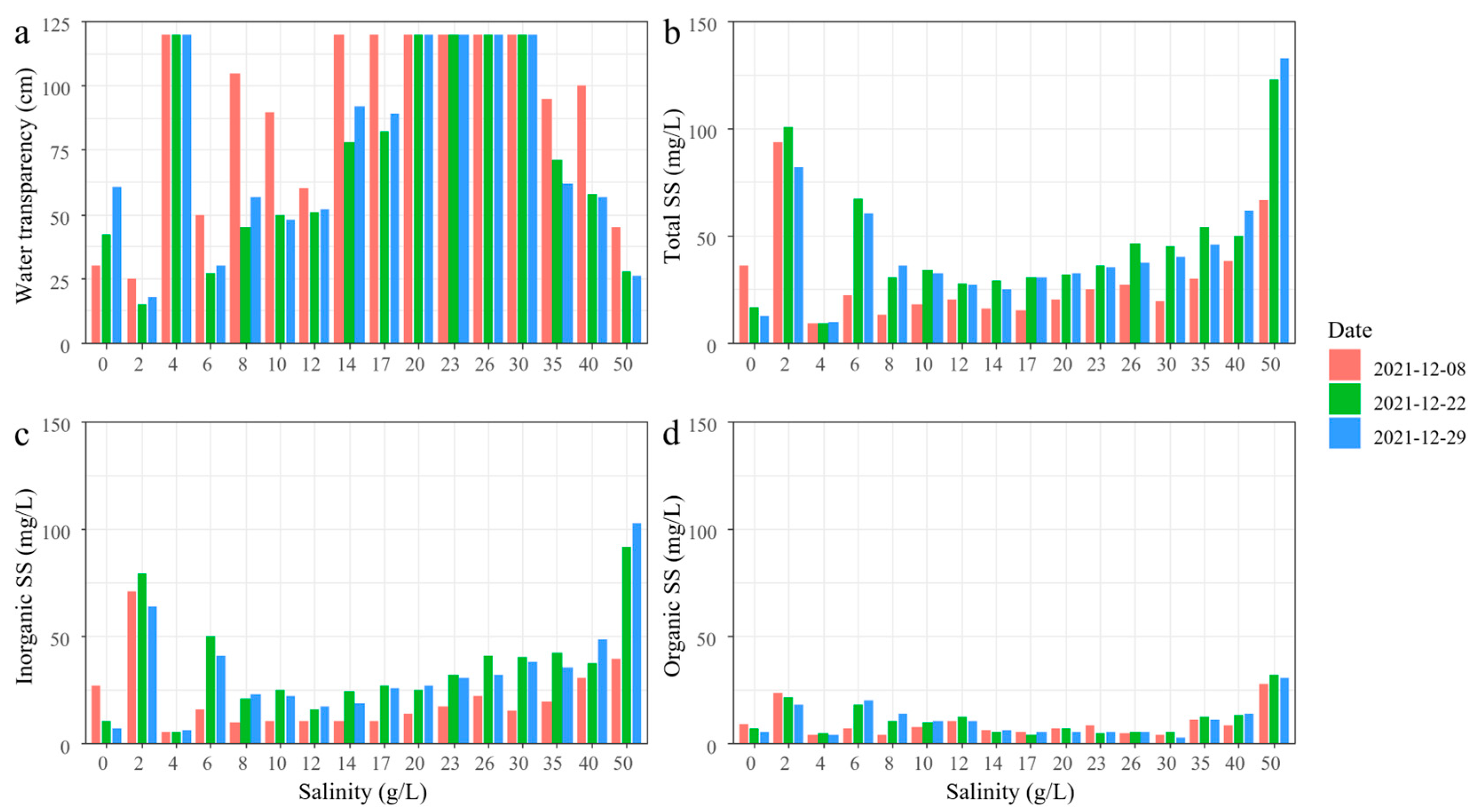
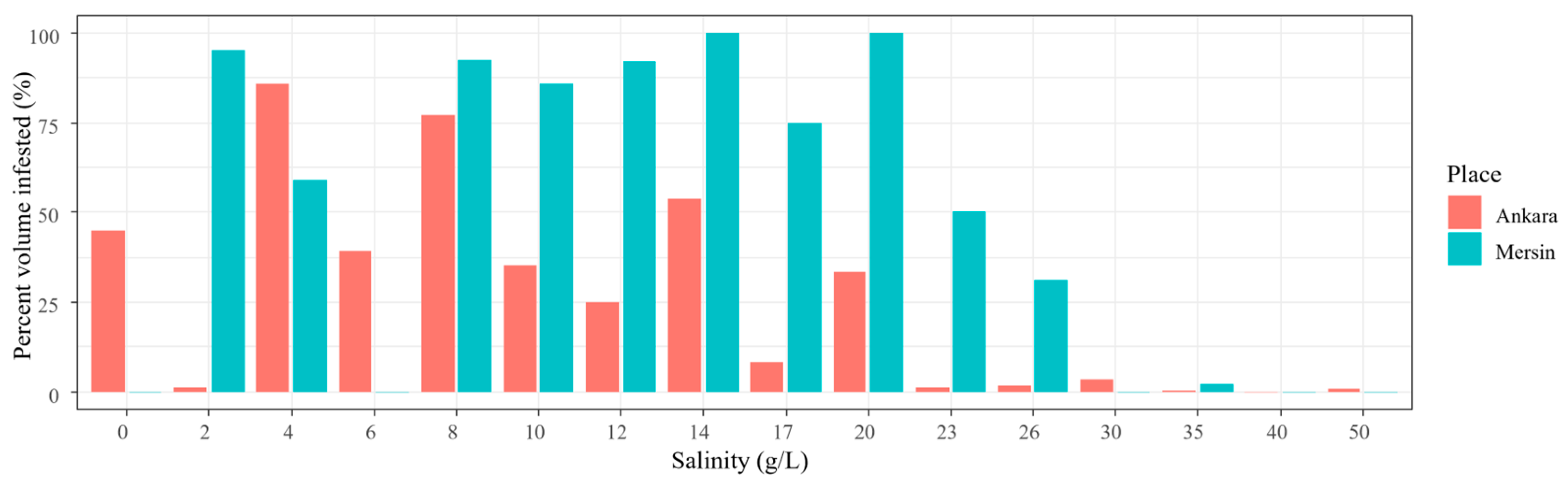
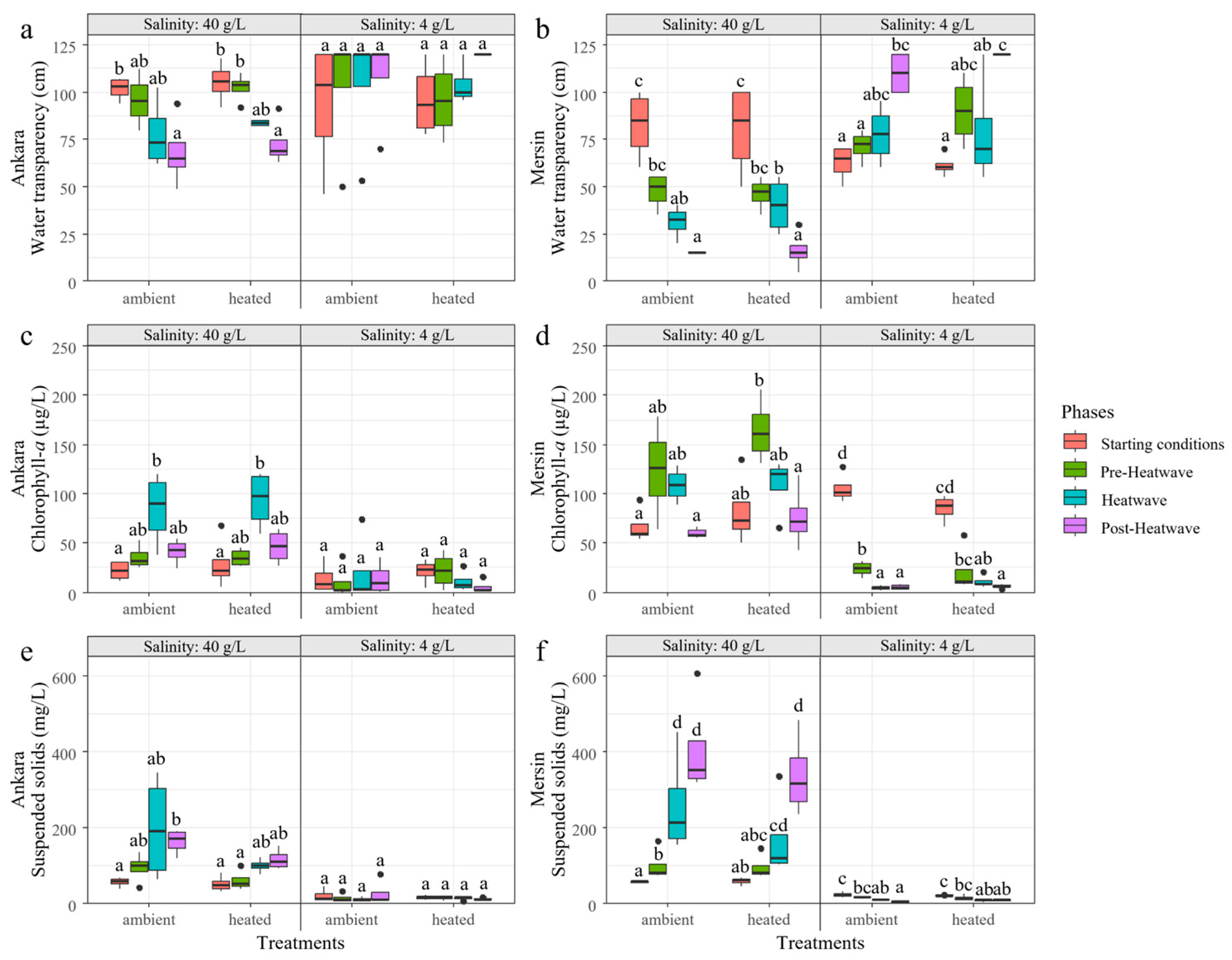
3.1. Experiment 1
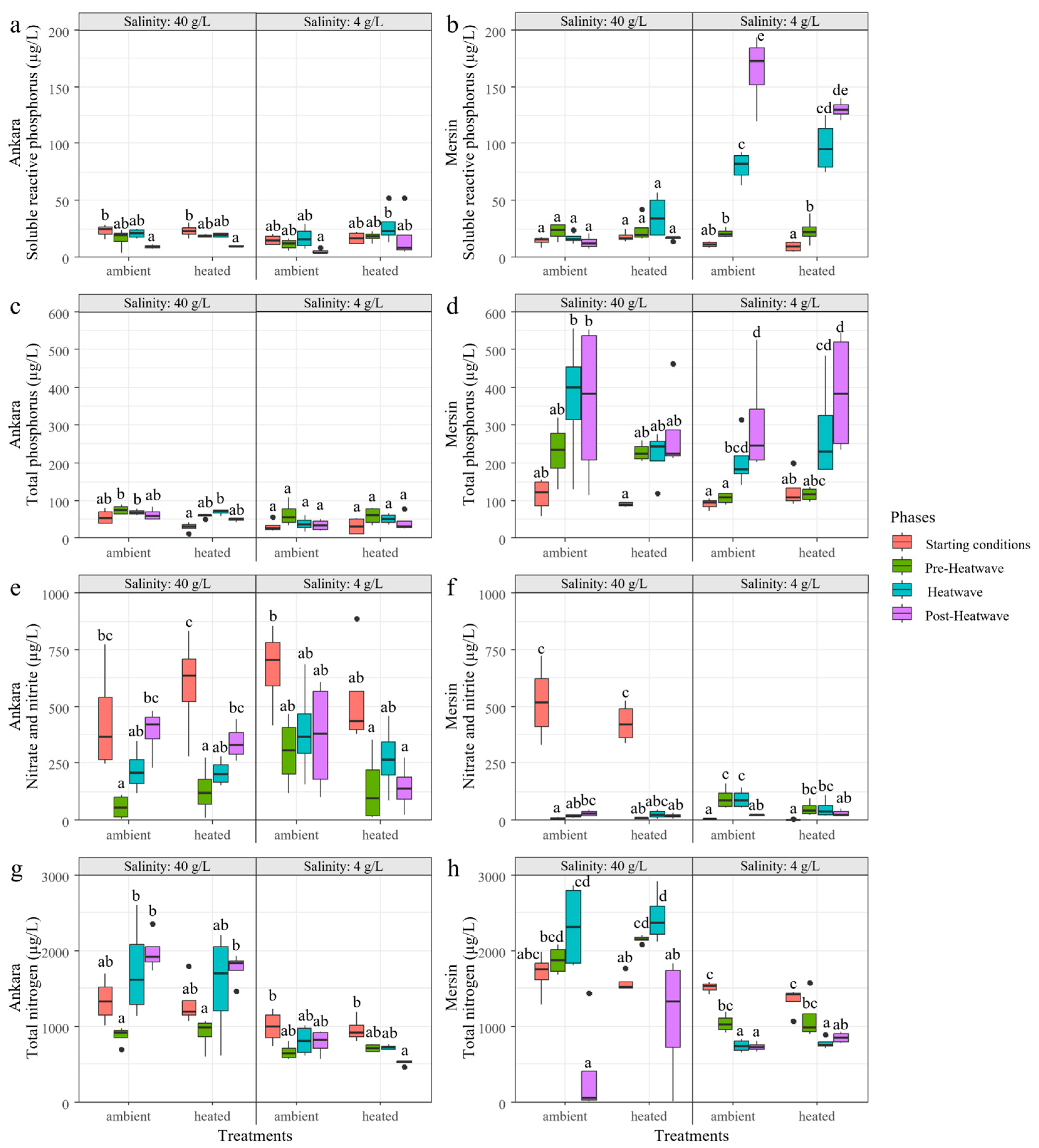
3.2. Experiment 2
4. Conclusions and Recommendations
- (1)
- High salinities cause a corrosive working environment, which is further exacerbated by UV radiation in exposed mesocosms. Therefore, the design of the mesocosms, sensors, and all other design elements should be resistant to corrosion. HDPE tanks are readily available as they are also used in the food industry and are a good choice due to their resistance to salinity and UV. However, they have less structural durability and can easily be damaged by physical forces. Therefore, enforced ground (concrete or compacted sand if the groundwater table is high), as well as thick support material for walls, should be employed. Another advantage of HDPE tanks is that they can be repaired using heat in case of minor damage. The sensors should also be resistant to high corrosion environments, requiring the use of, for instance, plastic or titanium-coated materials. If stainless steel is desired, the type of steel used must be able to resist salinities higher than marine levels. The corrosion risk is especially concerning for heating elements. Conventional stainless steel or Teflon-coated heaters could easily fail as an electric field in saline waters entails fast corrosion of metal parts. Conventional aquarium heaters are resistant to saline waters, but they have limited capacity and require a large amount of heater use, and high-capacity aquarium heaters generally work with a circulation pump, which is not desired for natural experiments. We, therefore, recommend the application of specially designed high-capacity industrial heating elements enclosed in heat-resistant glass tubes.
- (2)
- High-salinity water has high conductivity for electricity, which poses challenges for elevated electrification hazards, especially if heating is involved. Frequent safety training as well as safety switches are recommended. Due to the high salinity and consequent high conductivity, there is also an increased risk of sensor interference. The sensors should be well grounded, and sensors with circuit-verified electricity shield components should be selected. A trial before a bulk purchase is recommended.
- (3)
- The majority of the sensors available on the market are focused on freshwater and marine research (salinity < 38 g/L). Therefore, the overall design and calibration algorithms are generally not optimized for very high salinities (>40 g/L). Consequently, it should be verified with the suppliers if the sensors are calibrated for high salinities and if the potential interferences with major ions are not a problem. It can also be checked with the suppliers to see if it is possible to add extra calibration parameters to the sensor algorithms.
- (4)
- High salinities create a challenge for chemistry analyses. The majority of the chemical analyses can only be conducted within specific salinity ranges, and some of the reactions are prone to interference at a high concentration of major ions, so the analysis matrix should correspond to the salinity range of the samples. Considering that the majority of traditional laboratory facilities optimize their workflow and know-how with a focus on either freshwater or marine ecosystems, the laboratory procedures should be thoroughly verified by the scientific and technical teams before designing and implementing a mesocosm experiment. Experimental salinity treatments may result in a large salinity gradient in the sample pool, which requires different salinity matrixes in the eluent as well as potentially different analysis methods. Dilution of samples, as long as the desired parameter has sufficiently high concentrations, could be applied. Therefore, grouping samples according to salinity concentrations and conducting the analyses according to salinity levels in these groups would be the best practice. Laboratories conducting experimental research on salinity gradients should be ready to work with different sample matrixes and analysis methods.
- (5)
- Experimental set-ups across large salinity gradients pose complications for biota inoculation. Even with a high salinity tolerance range, organisms require a longer adaptation time before being released into environments with different salinities. Therefore, higher logistics demand and longer handling times should be expected for successful inoculations. Several large and well-equipped inoculation adaptation mesocosms at targeted salinity levels could be useful for the experimental mesocosms. Furthermore, high mortality rates (of fish, for example) should be expected, and thus successive inoculations with larger inoculum populations should be anticipated.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeppesen, E.; Beklioğlu, M.; Özkan, K.; Akyürek, Z. Salinization Increase Due to Climate Change Will Have Substantial Negative Effects on Inland Waters: A Call for Multifaceted Research at the Local and Global Scale. Innovation 2020, 1, 10030. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, G.; Çolak, M.A.; Özgencil, İ.K.; Metin, M.; Korkmaz, M.; Ertuğrul, S.; Soyluer, M.; Bucak, T.; Tavşanoğlu, Ü.N.; Özkan, K.; et al. Decadal Changes in Size, Salinity, Waterbirds, and Fish in Lakes of the Konya Closed Basin, Turkey, Associated with Climate Change and Increasing Water Abstraction for Agriculture. Inland Waters 2021, 11, 538–555. [Google Scholar] [CrossRef]
- Çolak, M.A.; Öztaş, B.; Özgencil, İ.K.; Soyluer, M.; Korkmaz, M.; Ramírez-García, A.; Metin, M.; Yılmaz, G.; Ertuğrul, S.; Tavşanoğlu, Ü.N.; et al. Increased Water Abstraction and Climate Change Have Substantial Effect on Morphometry, Salinity, and Biotic Communities in Lakes: Examples from the Semi-Arid Burdur Basin (Turkey). Water 2022, 14, 1241. [Google Scholar] [CrossRef]
- Şekercioğlu, Ç.H.; Anderson, S.; Akçay, E.; Bilgin, R.; Can, Ö.E.; Semiz, G.; Tavşanoğlu, Ç.; Yokeş, M.B.; Soyumert, A.; İpekdal, K.; et al. Turkey’s Globally Important Biodiversity in Crisis. Biol. Conserv. 2011, 144, 2752–2769. [Google Scholar] [CrossRef]
- Cunillera-Montcusí, D.; Beklioğlu, M.; Cañedo-Argüelles, M.; Jeppesen, E.; Ptacnik, R.; Amorim, C.A.; Arnott, S.E.; Berger, S.A.; Brucet, S.; Dugan, H.A.; et al. Freshwater Salinisation: A Research Agenda for a Saltier World. Trends Ecol. Evol. 2022, 37, 440–453. [Google Scholar] [CrossRef]
- Stewart, R.I.A.; Dossena, M.; Bohan, D.A.; Jeppesen, E.; Kordas, R.L.; Ledger, M.E.; Meerhoff, M.; Moss, B.; Mulder, C.; Shurin, J.B.; et al. Mesocosm Experiments as a Tool for Ecological Climate-Change Research. Adv. Ecol. Res. 2013, 48, 71–181. [Google Scholar]
- Lampert, W.; Sommer, U. Limnoecology: The Ecology of Lakes and Streams, 2nd ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Benton, T.G.; Solan, M.; Travis, J.M.J.; Sait, S.M. Microcosm Experiments Can Inform Global Ecological Problems. Trends Ecol. Evol. 2007, 22, 516–521. [Google Scholar] [CrossRef]
- Fordham, D.A. Mesocosms Reveal Ecological Surprises from Climate Change. PLoS Biol. 2015, 13, e1002323. [Google Scholar] [CrossRef] [Green Version]
- Odum, E.P. The Mesocosm. Bioscience 1984, 34, 558–562. [Google Scholar] [CrossRef]
- Landkildehus, F.; Søndergaard, M.; Beklioglu, M.; Adrian, R.; Angeler, D.G.; Hejzlar, J.; Papastergiadou, E.; Zingel, P.; Çakiroğlu, A.I.; Scharfenberger, U.; et al. Climate Change Effects on Shallow Lakes: Design and Preliminary Results of a Cross-European Climate Gradient Mesocosm Experiment. Est. J. Ecol. 2014, 63, 71. [Google Scholar] [CrossRef] [Green Version]
- Liboriussen, L.; Landkildehus, F.; Meerhoff, M.; Bramm, M.E.; Søndergaard, M.; Christoffersen, K.; Richardson, K.; Søndergaard, M.; Lauridsen, T.L.; Jeppesen, E. Global Warming: Design of a Flow-through Shallow Lake Mesocosm Climate Experiment. Limnol. Oceanogr. Methods 2005, 3, 1–9. [Google Scholar] [CrossRef]
- Stephen, D.; Balayla, D.M.; Becares, E.; Collings, S.E.; Fernandez-Alaez, C.; Fernandez-Alaez, M.; Ferriol, C.; Garcia, P.; Goma, J.; Gyllstrom, M.; et al. Continental-Scale Patterns of Nutrient and Fish Effects on Shallow Lakes: Introduction to a Pan-European Mesocosm Experiment. Freshw. Biol. 2004, 49, 1517–1524. [Google Scholar] [CrossRef] [Green Version]
- Donaghay, P.; Klos, E. Physical, Chemical and Biological Responses to Simulated Wind and Tidal Mixing in Experimental Marine Ecosystems. Mar. Ecol. Prog. Ser. 1985, 26, 35–45. [Google Scholar] [CrossRef]
- Brockmann, U. Pelagic Mesocosms: II. Process Studies. Enclosed Exp. Mar. Ecosyst. A Rev. Recomm. 1990, 37, 81–108. [Google Scholar]
- Nejstgaard, J.; Hygum, B.; Naustvoll, L.; Båmstedt, U. Zooplankton Growth, Diet and Reproductive Success Compared in Simultaneous Diatom- and Flagellate-Microzooplankton-Dominated Plankton Blooms. Mar. Ecol. Prog. Ser. 2001, 221, 77–91. [Google Scholar] [CrossRef]
- Riebesell, U.; Czerny, J.; von Bröckel, K.; Boxhammer, T.; Büdenbender, J.; Deckelnick, M.; Fischer, M.; Hoffmann, D.; Krug, S.A.; Lentz, U.; et al. Technical Note: A Mobile Sea-Going Mesocosm System—New Opportunities for Ocean Change Research. Biogeosciences 2013, 10, 1835–1847. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Søndergaard, M.; Jensen, J.P.; Kanstrup, E.; Petersen, B. Macrophytes and Turbidity in Brackish Lakes with Special Emphasis on the Role of Top-Down Control. In The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; pp. 369–377. [Google Scholar]
- Christensen, I.; Pedersen, L.K.; Søndergaard, M.; Lauridsen, T.L.; Tserenpil, S.; Richardson, K.; Amorim, C.A.; Pacheco, J.P.; Jeppesen, E. Impact of Zooplankton Grazing on Phytoplankton in North Temperate Coastal Lakes: Changes along Gradients in Salinity and Nutrients. Hydrobiologia 2022, 27, 1–18. [Google Scholar] [CrossRef]
- Hintz, W.D.; Mattes, B.M.; Schuler, M.S.; Jones, D.K.; Stoler, A.B.; Lind, L.; Relyea, R.A. Salinization Triggers a Trophic Cascade in Experimental Freshwater Communities with Varying Food-Chain Length. Ecol. Appl. 2017, 27, 833–844. [Google Scholar] [CrossRef]
- Moffett, E.R.; Baker, H.K.; Bonadonna, C.C.; Shurin, J.B.; Symons, C.C. Cascading Effects of Freshwater Salinization on Plankton Communities in the Sierra Nevada. Limnol. Oceanogr. Lett. 2023, 8, 30–37. [Google Scholar] [CrossRef]
- Hintz, W.D.; Arnott, S.E.; Symons, C.C.; Greco, D.A.; McClymont, A.; Brentrup, J.A.; Cañedo-Argüelles, M.; Derry, A.M.; Downing, A.L.; Gray, D.K.; et al. Current Water Quality Guidelines across North America and Europe Do Not Protect Lakes from Salinization. Proc. Natl. Acad. Sci. USA 2022, 119, e2115033119. [Google Scholar] [CrossRef]
- Hébert, M.; Symons, C.C.; Cañedo-Argüelles, M.; Arnott, S.E.; Derry, A.M.; Fugère, V.; Hintz, W.D.; Melles, S.J.; Astorg, L.; Baker, H.K.; et al. Lake Salinization Drives Consistent Losses of Zooplankton Abundance and Diversity across Coordinated Mesocosm Experiments. Limnol. Oceanogr. Lett. 2023, 8, 19–29. [Google Scholar] [CrossRef]
- Reas, C.; Fry, B. Processing: Programming for the Media Arts. AI Soc. 2006, 20, 526–538. [Google Scholar] [CrossRef]
- Carlson, R.E. A Trophic State Index for Lakes1. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Kratzer, C.R.; Brezonik, P.L. A Carlson-Type Trophic State Index for Nitrogen in Florida Lakes. J. Am. Water Resour. Assoc. 1981, 17, 713–715. [Google Scholar] [CrossRef]
- Bartlett, K.B.; Bartlett, D.S.; Harriss, R.C.; Sebacher, D.I. Methane Emissions along a Salt Marsh Salinity Gradient. Biogeochemistry 1987, 4, 183–202. [Google Scholar] [CrossRef]
- Poffenbarger, H.J.; Needelman, B.A.; Megonigal, J.P. Salinity Influence on Methane Emissions from Tidal Marshes. Wetlands 2011, 31, 831–842. [Google Scholar] [CrossRef]
- Barnes, R.O.; Goldberg, E.D. Methane Production and Consumption in Anoxic Marine Sediments. Geology 1976, 4, 297. [Google Scholar] [CrossRef]
- Irakoze, W.; Prodjinoto, H.; Nijimbere, S.; Rufyikiri, G.; Lutts, S. NaCl and Na2SO4 Salinities Have Different Impact on Photosynthesis and Yield-Related Parameters in Rice (Oryza sativa L.). Agronomy 2020, 10, 864. [Google Scholar] [CrossRef]
- Jespersen, A.-M.; Christoffersen, K. Measurements of Chlorophyll-a from Phytoplankton Using Ethanol as Extraction Solvent. Arch. Hydrobiol. 1987, 109, 445–454. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1996.
- Mackereth, F.J.H.; Heron, J.; Talling, J.F. Water Analysis: Some Revised Methods for Limnologists. Freshw. Biol. Assoc. Sci. Pub. 1978, 36, 117. [Google Scholar]
- Kroon, F.J. Total Nitrojen. In Standard Methods for the Examination of Water and Wastewater; Kemp, A.L.W., Ed.; American Public Health Association: Washington, DC, USA, 1993; pp. 5–23. [Google Scholar]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis; Verlag Chemie Weinheim: Deerfield Beach, FL, USA, 1983; p. 419. [Google Scholar]
- Schriver, P.; Bogestrand, J.; Jeppesen, E.; Sondergaard, M. Impact of Submerged Macrophytes on Fish-Zooplanl Phytoplankton Interactions: Large-Scale Enclosure Experiments in a Shallow Eutrophic Lake. Freshw. Biol. 1995, 33, 255–270. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2023. [Google Scholar]
- Wickham, H.; Chang, W.; Wickham, M.H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; CRS Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Kay, M.; Elkin, L.A.; Higgins, J.J.; Wobbrock, J.O. ARTool: Aligned Rank Transform for Nonparametric Factorial ANOVAs; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Droppo, I.G. Rethinking What Constitutes Suspended Sediment. Hydrol. Process. 2001, 15, 1551–1564. [Google Scholar] [CrossRef]
- Shen, X.; Lee, B.J.; Fettweis, M.; Toorman, E.A. A Tri-Modal Flocculation Model Coupled with TELEMAC for Estuarine Muds Both in the Laboratory and in the Field. Water Res. 2018, 145, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, L.; Scheffer, M.; Martínez-Ramos, M. Catastrophic Response of Lakes to Benthivorous Fish Introduction. Oikos 2001, 94, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Liu, C.; Hu, Y.; Shao, K.; Tang, X.; Zhang, L.; Gao, G.; Qin, B. Climate-Induced Salinization May Lead to Increased Lake Nitrogen Retention. Water Res. 2023, 228, 119354. [Google Scholar] [CrossRef]
- Zhu, J.; He, Y.; Zhu, Y.; Huang, M.; Zhang, Y. Biogeochemical Sulfur Cycling Coupling with Dissimilatory Nitrate Reduction Processes in Freshwater Sediments. Environ. Rev. 2018, 26, 121–132. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Özkan, K.; Korkmaz, M.; Amorim, C.A.; Yılmaz, G.; Koru, M.; Can, Y.; Pacheco, J.P.; Acar, V.; Çolak, M.A.; Yavuz, G.C.; et al. Mesocosm Design and Implementation of Two Synchronized Case Study Experiments to Determine the Impacts of Salinization and Climate Change on the Structure and Functioning of Shallow Lakes. Water 2023, 15, 2611. https://doi.org/10.3390/w15142611
Özkan K, Korkmaz M, Amorim CA, Yılmaz G, Koru M, Can Y, Pacheco JP, Acar V, Çolak MA, Yavuz GC, et al. Mesocosm Design and Implementation of Two Synchronized Case Study Experiments to Determine the Impacts of Salinization and Climate Change on the Structure and Functioning of Shallow Lakes. Water. 2023; 15(14):2611. https://doi.org/10.3390/w15142611
Chicago/Turabian StyleÖzkan, Korhan, Mustafa Korkmaz, Cihelio Alves Amorim, Gültekin Yılmaz, Meltem Koru, Yasemin Can, Juan Pablo Pacheco, Vildan Acar, Mehmet Arda Çolak, Gül Canan Yavuz, and et al. 2023. "Mesocosm Design and Implementation of Two Synchronized Case Study Experiments to Determine the Impacts of Salinization and Climate Change on the Structure and Functioning of Shallow Lakes" Water 15, no. 14: 2611. https://doi.org/10.3390/w15142611