1. Introduction
Despite the high cost of materials and labor (between
$25,000 and
$100,000 ha
−1), protected cultivation has gained popularity in Chilean fruit orchards during the last few years. Blueberries and other fruit crops can be grown under tunnels, greenhouses, or netting to protect plants from climatic adversities [
1,
2,
3] or to accelerate ripening [
4]. For example, fruit sunburn was reduced by 26% [
5] and 44% [
6] in apple and cherry orchards covered with shade nets. In Chile, earlier harvests and higher yields have been reported in blueberry crops covered with polyethylene film due to changes in microclimate conditions and net photosynthesis [
7]. Plastic film tunnels are preferred to protect blueberry plants from frost and accelerate ripening in northern Chile, while shade nets protect berries from high solar radiation in central growing regions. Plastic film roofs protect blueberry orchards from rain and minimize fruit cracking or rotting in southern Chile.
From a water conservation perspective, the use of protected cultivation has been associated with a reduction in transpiration [
8] or a delay in the onset of severe water stress [
9]. The decrease in the transpiration rate of covered plants has been attributed to the impact of the covering on the atmospheric water demand (VPD) of orchards [
10]. However, the effect of the covering on VPD is not always the same. In kiwifruit (
Actinidia deliciosa Chev.), plants covered with a low-density plastic film had similar VPD values during the growing season as uncovered plants [
9]. Conversely, the use of a 50% red shade netting increased xylem water flow and water demand in apple trees [
11]. These results demonstrate the need to measure VPD to understand how plant water demand is altered by the use of protected cultivation.
Depending on the amount and quality of light transmitted to the trees, the use of coverings can stimulate shoot elongation and alter leaf anatomy and function [
12,
13]. The higher the vegetative growth, the higher the plant water loss through transpiration, emphasizing the importance of measuring canopy size or density (i.e., leaf area index) to understand how protected cultivation affects plant–water relations.
Highbush blueberry (
Vaccinium corymbosum L.) is highly sensitive to water stress because the depth of its root system is limited to the upper soil layers (0–0.4 m) and root hairs are not produced [
14]. Therefore, blueberry plants require sensitive stomatal control to reduce transpiration and thus tolerate water deficits. Highbush blueberry plants begin to show an exponential decrease in stomatal conductance (gs) when the leaf water potential is less than −0.2 MPa at midday [
15]. Rapid stomatal closure in response to water stress reduces transpiration and susceptibility to xylem cavitation. A reduction in xylem hydraulic conductivity of almost 50% has been found when the leaf water potential is between −1.2 and −1.0 MPa [
16]. Although stomatal closure is an efficient mechanism to limit water loss, moderate water stress (SWP between −1.0 and −0.8 MPa) for more than 21 days can reduce plant photosynthesis by more than 30% [
17]. When the midday SWP is close to −1.8 MPa, net photosynthesis can be reduced by 50% in only 1 week [
18].
Several studies have reported that canopies can induce an increase in
gs. In hazelnut (
Corylus avellana L.), plants covered with netting showed an increased leaf stomatal density [
19]. In kiwifruit (
Actinidia deliciosa Chev.), plastic-covered vines showed a 20% increase in gs [
20]. The higher gs in covered plants under optimal water conditions may increase the transpiration rate, but not necessarily carbon assimilation and water use efficiency. These results clearly show that the role of coverings in plant–water relations goes beyond reducing the water demand of fruit orchards.
Blueberry cultivation in areas with high evaporative demand (<−3.0 kPa) has been severely affected by climate change, as the increase in evapotranspiration and the progressive decrease in precipitation have reduced water availability for agriculture. For example, in several blueberry orchards in the Maule region (Chile’s main blueberry production area) and other production areas with similar climatic conditions, a 10% increase in ETc and a 45% decrease in annual precipitation have been recorded over the last decade [
21].
In recent years, the Chilean blueberry industry has begun to plant new cultivars that have higher yields and better quality characteristics than the old cultivars, such as ‘Duke’, ‘Blue Crop’, and ‘Elliot’, among others. ‘Blue Ribbon’ and ‘Top Shelf’ are two mid-season cultivars that are adapted to mild winters and produce firm, sweet, and large berries [
22]. Currently, ‘Blue Ribbon’ and ‘Top Shelf’ are among the seven most commercialized cultivars in Chile, and both have been grown under a wide range of conditions, including the use of protected cultivation with different cover materials (nets and low- and high-density plastic films). Unfortunately, there is a lack of knowledge on how these new cultivars respond to low water availability under protected cultivation, especially considering the existing variability in covering materials. This is particularly important for new cultivars, as the response of plants to drought is highly dependent on the genetic characteristics of each cultivar. The objective of this study was to determine the effect of different types of coverings on plant–water relations in two new blueberry cultivars in recently planted orchards under two temperate climate types.
2. Materials and Methods
2.1. Study Sites
This study was conducted for two consecutive seasons (2018–2019 and 2019–2020) in two drip-irrigated highbush blueberry (Vaccinium corymbosum L.) orchards. The first orchard was established in 2018 with cv. Top Shelf and cv. Blue Ribbon in Linares, the Maule Region, Chile (35°81′04.97″ S; 71°32′27.36″ W), with 1 m spacing between plants and 3 m between rows. The soil corresponds to the Linares series and belongs to the mixed, thick clay thermal family of typical xerorthents (Entisol), with a dominant clay surface texture. The other orchard (the Traiguen orchard) was planted in 2017 with cv. Top Shelf in Traiguén, the Araucanía Region, Chile (38°33′82.37″ S; 71°54′02.45″ W), with a 1 m plant spacing and 3.2 m row spacing. The soil is mapped as the Chufquén series and belongs to the mixed, loamy fine mesic family of the Fluventic Humic Dystrudepts (Inceptisol), with a surface texture that is predominantly silty loam. The climatic conditions in Linares and Traiguén correspond to the temperate Mediterranean and humid temperate climates, respectively. The physical soil analysis showed that the field capacity was 40.9% and 28.8% for Linares and Traiguén, respectively. The permanent wilting point was 28.4% and 17.6% for Linares and Traiguén, respectively. The bulk density was 1.28 g cm−3 in Linares and 1.48 g cm−3 in Traiguén.
2.2. Experimental Design
This study was conducted in a completely randomized block design with four replications and repeated measurements over two years. A deficit irrigation treatment was implemented in a split-plot design in the Linares orchard during the 2019–2020 season. The main plot was the cover treatment, the first subplot was the cultivar, and the second subplot was the irrigation treatment. In the Traiguén orchard, all plants were irrigated in a similar way to avoid the occurrence of water stress throughout the growing season (midday stem water potential above −0.5 MPa). A total of 4 covering treatments (control: uncovered plants; netting: plants covered with a 20% black shade net; woven: plants covered with a high-density polyethylene film; and plastic: plants covered with a low-density polyethylene film) were applied in each orchard from bud break until the onset of dormancy. The covering materials were placed at a height of 3 m above the ground, with a width of 2.5 m and an angle of 28º to the ground. The nets were mounted on a structure made of impregnated pine blocks or concrete and spaced 12 m apart in both orchards (
Figure 1).
The uncovered plants remained in open field conditions throughout the season. Cover treatments were installed for 240 consecutive days (from 1 September to 1 May) each season. The cover treatments were installed over the main plot of 12 plants in both orchards, at a height of 0.70 m above each of the 3 consecutive rows for each combination of block and treatment (10 pl row−1).
In the Linares orchard, 2 irrigation treatments were randomly applied to a subplot of 6 plants within each main plot 1 week after veraison (when 50% of the berries turned from green to red), with the aim of obtaining significant differences in plant water status near harvest time, when water availability in both locations often reaches the minimum value of the season (midday stem water potential below −0.8 MPa). Control (WET) plants were irrigated to provide at least 100% ETc throughout the season, trying to maintain an average midday stem water potential (Ψs) above −0.8 MPa. In the regulated deficit irrigation (RDI) treatment, water was withheld from both covered and uncovered plants for 14 consecutive days starting on 16 January. Irrigation was resumed in all plants when deficit-irrigated subplots reached an average midday stem water potential (Ψs) of −1.2 MPa.
2.3. Environmental Conditions and Water Requirements
The atmospheric evaporative demand was evaluated as the vapor pressure deficit (VPD) under each cover using meteorological stations (PCE-FWS 20, PCE Instruments, Southampton, UK). Air temperature and relative humidity (°C) were measured with a sampling frequency of 15 min from 10 August to 30 March in the 2018–2019 season, and from 10 August to 30 March in the 2019–2020 season. The VPD was calculated using the following formula [
23]:
where RH is the relative humidity, and Ta is the air temperature.
Photosynthetic photon flux density (PPFD) was measured weekly above and below the plants using a ceptometer (LP-80, Decagon Instruments, Washington, DC, USA) from 11 November to 3 February and from 15 October to 21 January in Linares and Traiguén, respectively. In 2019, data collection was performed every fourteen days at noon, with a sampling frequency of one measurement above the plant canopy and four measurements below the plant canopy. Above-canopy PPFD was measured 1.5 m above the ground, while below-canopy PPFD was measured at the center of the row, directly below the plant canes at 0, 0.15, and 0.3 m from the canopy, and at the edge of the canopy at 0.45 m from the canopy. The ceptometer used the PPFD data to calculate the leaf area index (LAI) after first entering the coordinates of each orchard to estimate the azimuth for that geographic area.
Reference evapotranspiration (ETo) and precipitation data were obtained from the weather station Santa Amada (Agroclima weather network) (35°76′03″ S 71°57′46″ W) for the Linares orchard, and from the weather stations Caballería (38°45′72″ S 72°74′02″ W) and La Providencia (38°28′92″ S 72°61′19″ W) (Agroclima weather network) for the Traiguen orchard. The weather stations were located approximately 10 km away from the study sites.
Irrigation requirements were estimated based on the evapotranspiration of blueberry plants:
where ETo is the reference evapotranspiration, and Kc is the crop coefficient. FC is the estimate of a dimensionless crop cover factor (0 < FC < 1) associated with Pc, which corresponds to the shading fraction of a plant at midday (0.1 < Pc < 0.7); F1 and F2 are crop-associated cover factors; Ds is the diameter of the vegetation shade at midday (m) determined from measurements of the plant’s shadow projection on the ground; and Eeh and Esh correspond to the spacing between rows (m) and between plants (m), respectively. The crop coefficients (kc) for young blueberry plants used in this study from bud break to harvest were obtained from Holzapfel et al. [
24].
Volumetric soil water content was measured in a single block (no replicates) under well-irrigated conditions using capacitance sensors (GS-1, Decagon Devices, Washington, DC, USA) connected to a data logger that recorded measurements every 15 min. Each sensor was installed at the midpoint between 2 irrigation emitters within the same irrigation line, at 2 depths (10 and 20 cm). Measurements were taken from 4 September to 13 March 2019. Water flow meters (Dishnon, Arad Ltd., Dalia, Israel) were installed at the beginning of each irrigation line in each main plot to estimate the irrigation water supply from 15 October to 3 February in the 2019–2020 season.
2.4. Plant Water Status and Physiological Responses
The water status of the blueberry plants was determined by measuring the stem water potential (SWP) at midday (from 12:30 to 15:30) using a pressure chamber (PMS 60, PMS Instruments, Washington, DC, USA). However, because blueberry plants have small petioles, whole shoots were sampled from the lower third of the canopy to improve the visualization of the meniscus during pressurization. We selected 1 shoot and covered it with an opaque and airtight bag for 45 min, according to previous studies [
25]. In the 2018–2019 season, SWP was measured only once, seven days after harvest (5 February 2019), to avoid reducing the leaf area in very young plants. In the 2019–2020 season, SWP was measured every fourteen days from fruit set to veraison and every seven days from veraison to harvest in one shoot per plant in each treatment.
Stomatal conductance (gs) was measured on three leaves per plant using a steady-state porometer (SC-1, Decagon Instruments, Washington, USA). Photosystem II (PSII) efficiency was assessed at the beginning and at the end of the season using a chlorophyll fluorimeter (Pocket PEA, Hansatech Instruments, England, UK) and determined on the basis of a variable (
Fv/Fm), where
Fv = [
Fm −
Fo], and
Fm = maximum fluorescence performance [
26], by previously darkening the leaves for 20 min [
27] and using claps with a measuring area of 4 mm in diameter. Both assessments were performed in the upper third of the canopy on mature, healthy, and sun-exposed leaves without chlorosis or obvious damage, using the same plant on which Ψ
s was determined.
2.5. Statistical Analysis
Data were subjected to analysis of variance (ANOVA) after testing for a normal distribution of errors (Shapiro–Wilks), homogeneity of variances (Levene), and additivity of effects (Tukey). In addition, we evaluated the relationship between environmental and physiological variables using linear and quadratic regression analysis. Regression coefficients were compared using analysis of covariance (ANCOVA). Differences between means were determined using the LSD test at a significance level of 95% (p ≤ 0.05). All statistical analyses were performed with SAS Studio software (SAS Institute, Cary, NC, USA).
3. Results
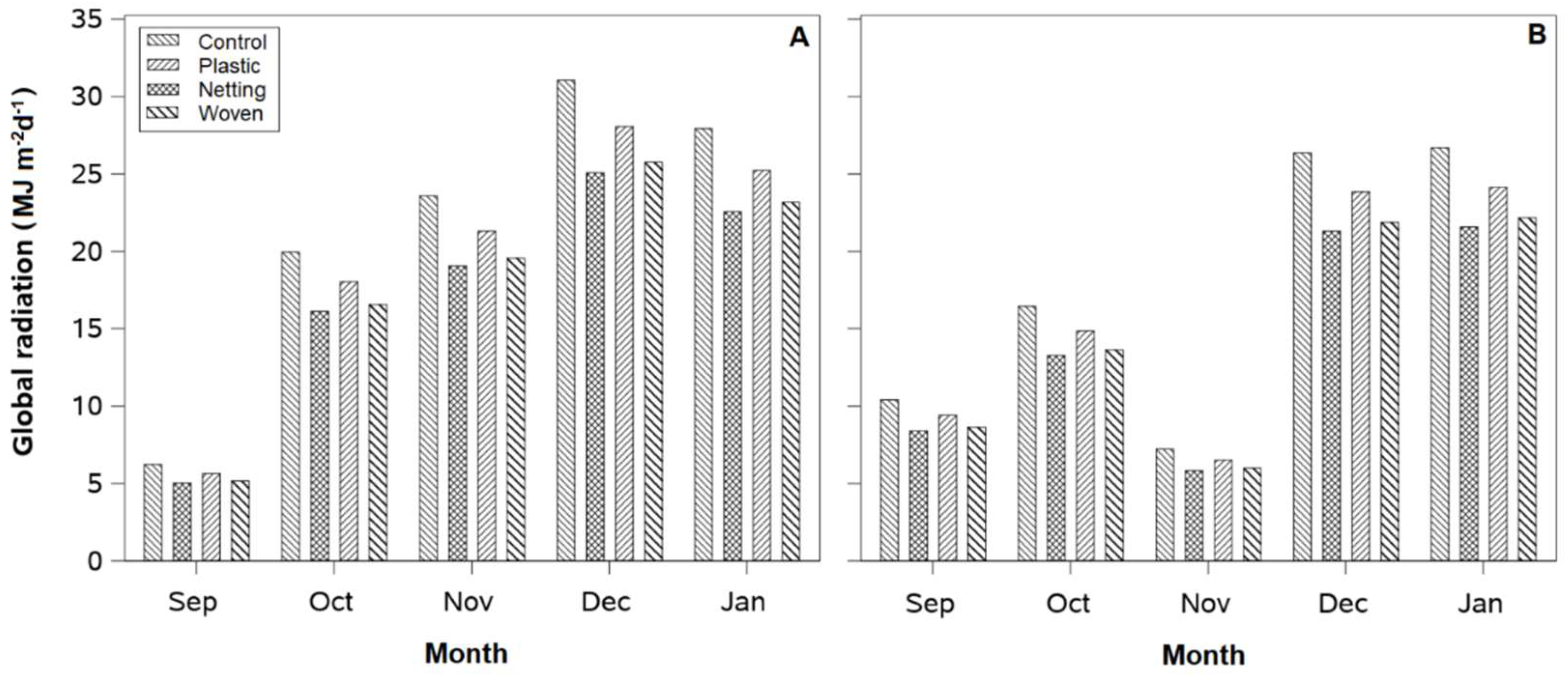
In general, global solar radiation (300–1100 nm) was reduced by 10% under the plastic cover and by 20% under the netting and woven covers (
Figure 2A,B). Meanwhile, PPFD decreased by approximately 25%, 38%, and 46% under the net, woven, and plastic covers, respectively (
Figure 3A,B). In December and January, under uncovered conditions, the Linares orchard had approximately 20% and 30% higher global solar radiation and PPFD values than the Traiguén orchard, respectively.
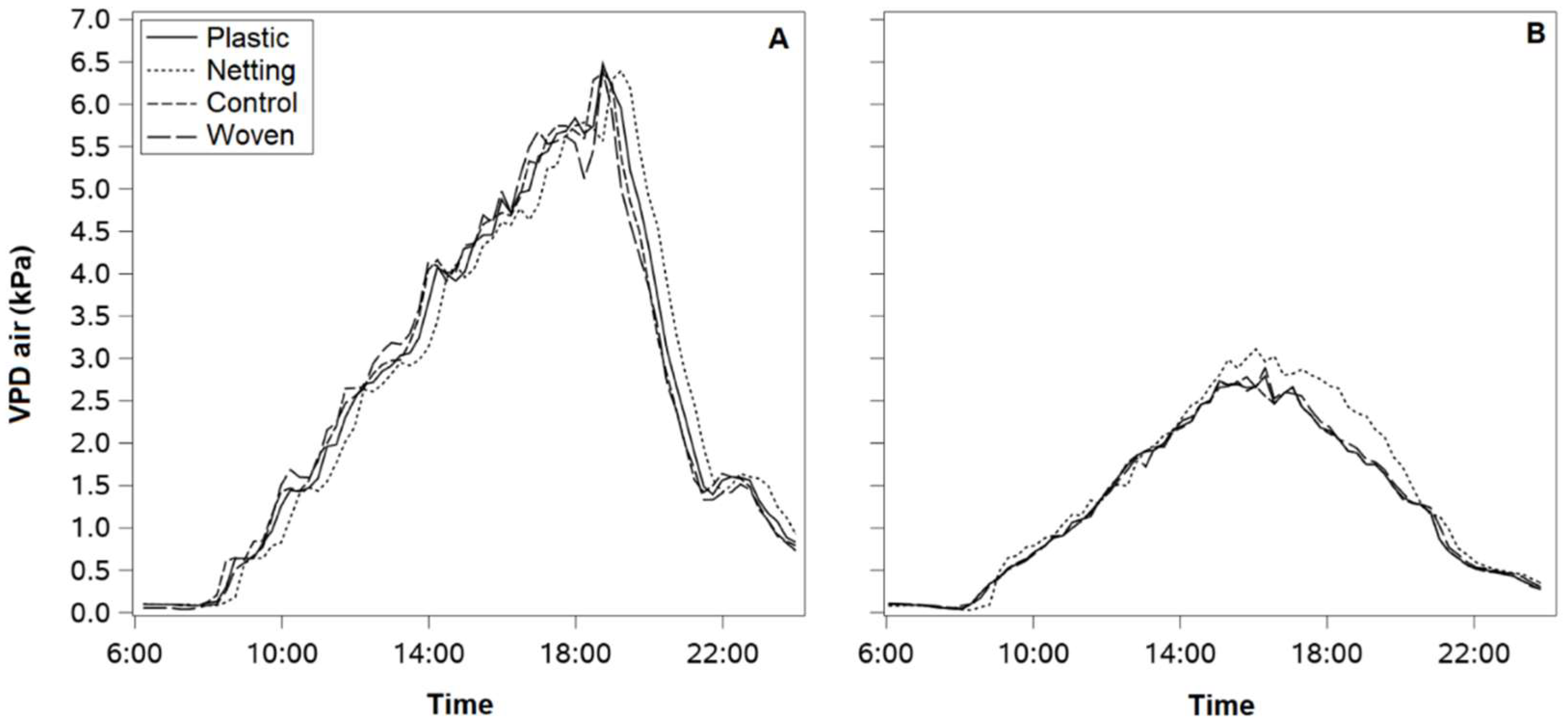
The cover treatments showed slight differences in air VPD compared with the control treatment in both study sites. In January, the month with the highest atmospheric evaporative demand, the air VPD was significantly higher in the Linares orchard, reaching maximum daily values of above 5 kPa in all canopy treatments (
Figure 4A). In the Traiguén orchard, the air VPD was often below 3.0 kPa during the same period of the year (
Figure 4B). In Linares, the cumulative ETc reached 380 mm in the plot planted with cv. Blue Ribbon and 340 mm in the plot planted with cv. Top Shelf from bud burst to leaf senescence. In Traiguén, the cumulative ETc during the 2nd season was 260 mm.
In 2019, the use of a plastic cover was often associated with the highest volumetric soil water content during the growing season (between 0.27 and 0.42 m
3 m
−3) (
Table 1). In contrast, the lowest SWC values (between 0.14 and 0.22 m
3 m
−3) were consistently measured in areas without cover. In the Linares orchard, the clearest differences in SWC between treatments were measured at a 10 cm depth, while in the Traiguén orchard, differences between treatments were found at both a 10 and 20 cm depth. The SWC values measured at a 10 and 20 cm depth tended to be similar for both cultivars at veraison. In contrast, the SWC values at a 20 cm depth were higher in plots planted with cv. Top Shelf (~5% higher than those with cv. Blue Ribbon) at harvest and post-harvest. At the end of the first season, a physical analysis of the root system of both cultivars showed that ‘Top Shelf’ had an effective plant rooting depth of 40 cm, whereas ‘Blue Ribbon’ did not exceed a depth of 20 cm. The leaf area index for both cultivars was similar for all treatments at each site (~2.0).
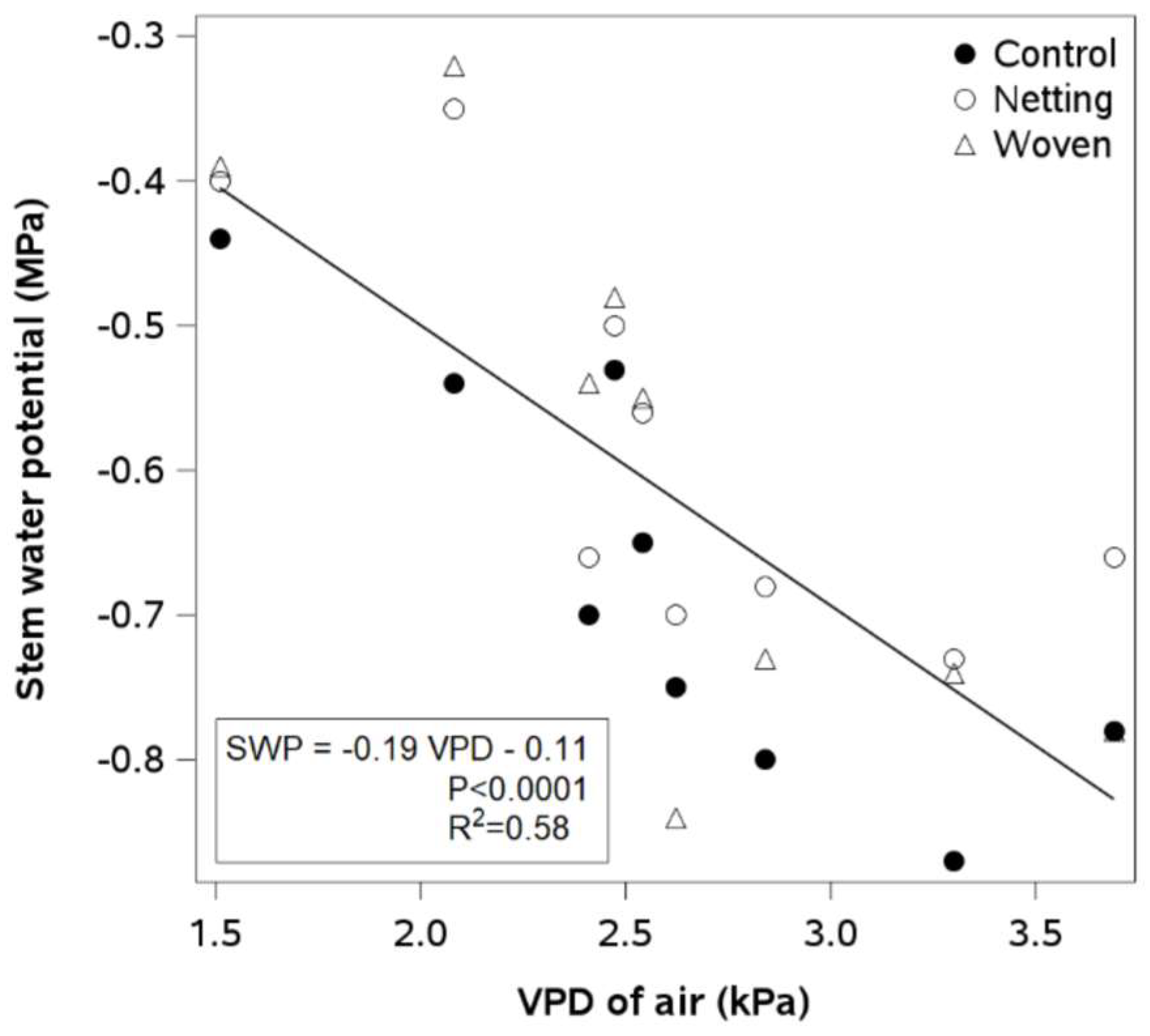
In Traiguén, there was no relationship between the midday values of VPD and SWP, regardless of the cover treatment. In Linares, the regression lines between both variables were significant for all cover treatments except plastic. The ANCOVA showed similar slopes and intercepts for the control, netting, and woven covering treatments, which explains why only one regression line was presented for all cover treatments (
Figure 5). The resulting negative linear regression indicated that lower SWP values were associated with a higher VPD. The values of the SWP ranged from −0.8 to −0.4 MPa, while the VPD values ranged from 1.5 to 3.5 kPa.
Under commercial irrigation practices, all cover treatments showed similar SWP values at the end of the first growing season in Linares and Traiguén, ranging between −0.7 and −0.6 MPa (
Table 2). In the second season, control plants from the Linares orchard consistently showed the lowest midday SWP values from veraison to postharvest, ranging between −0.9 and −0.7 MPa. In Traiguén, control plants exhibited the lowest midday SWP values at veraison and harvest, but not at postharvest. On the last measurement date, the lowest SWP was recorded in plants covered with plastic film. In Linares, the effect of each covering material on the midday SWP was not clear at veraison and harvest, but all coverings induced a higher SWP (approximately 0.1 MPa) at post-harvest. In Traiguén, plants covered with the plastic film had the highest SWP values at veraison and harvest (0.2 MPa higher than the control) but the lowest SWP values at post-harvest. In Linares, the midday SWP of the control plants tended to decrease as the season progressed (from −0.7 to −0.9 MPa), while in Traiguén, the SWP of the control plants remained relatively constant throughout the season (between −0.6 and −0.5 MPa). In the second season, there were no differences in SWP between the varieties from veraison to harvest, but ‘Top Shelf’ plants had a slightly higher SWP than ‘Blue Ribbon’ plants (0.1 MPa).
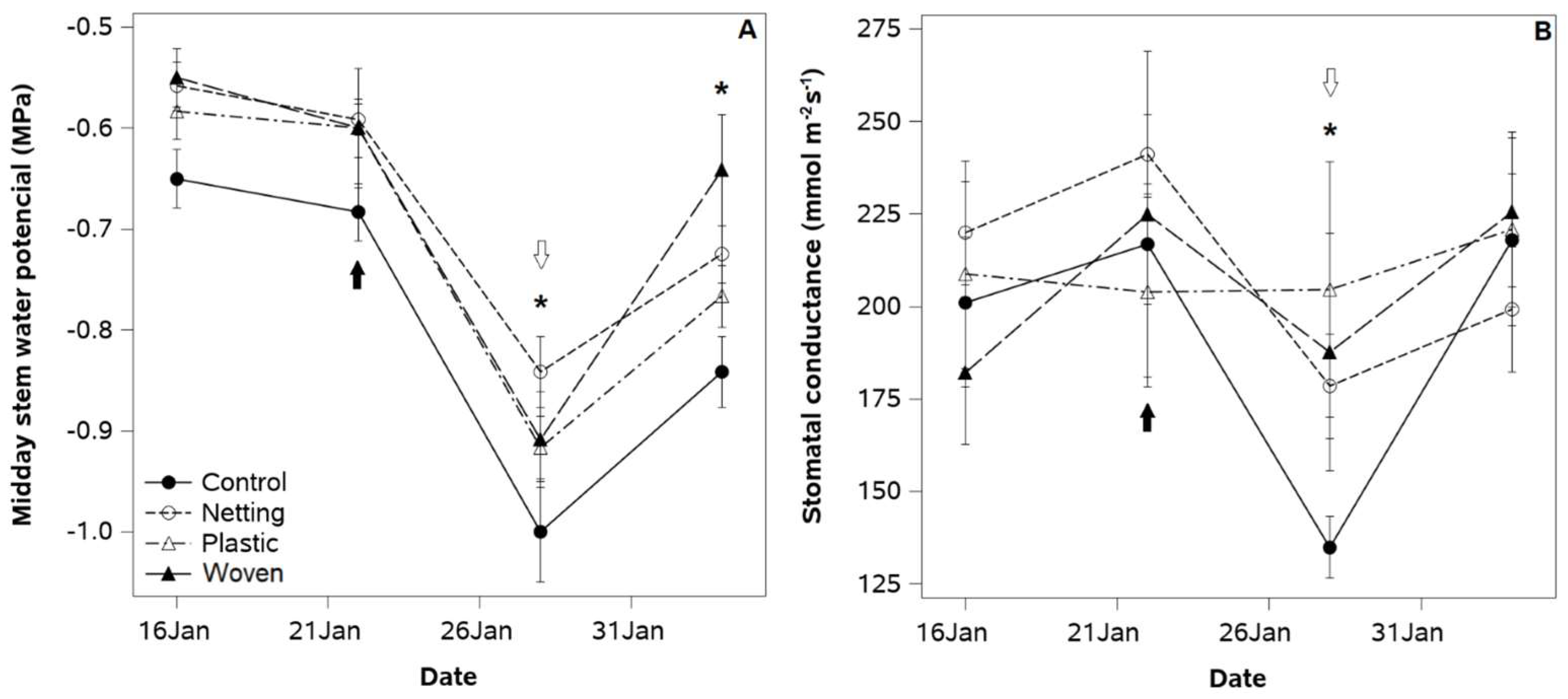
While in the first season, irrigation was provided continuously from bud break to leaf senescence, in 2019, irrigation completely ceased in both study sites after harvest time (January) for one week to test the effects of the covering treatments on deficit-irrigated plants. The results show that all coverings exhibited higher SWP values than the control (0.1 MPa) 1 week after irrigation was ceased in the whole orchard (
Figure 6A). Once irrigation was resumed, control plants still showed the lowest SWP among the treatments. However, midday SWP values in all treatments remained slightly lower than those registered before irrigation was cut off.
In the first season, irrigation was applied continuously from bud break to leaf senescence. In 2019, irrigation was completely stopped for one week after harvest time (January) in both study sites to test the effects of the covering treatments on deficit-irrigated plants. The results show that all cover treatments had higher SWP values than the control (0.1 MPa) 1 week after irrigation was stopped in the whole orchard (
Figure 6A). When irrigation was resumed, the control plants still had the lowest SWP among the treatments. However, the midday SWP remained slightly lower in all treatments than before irrigation was stopped.
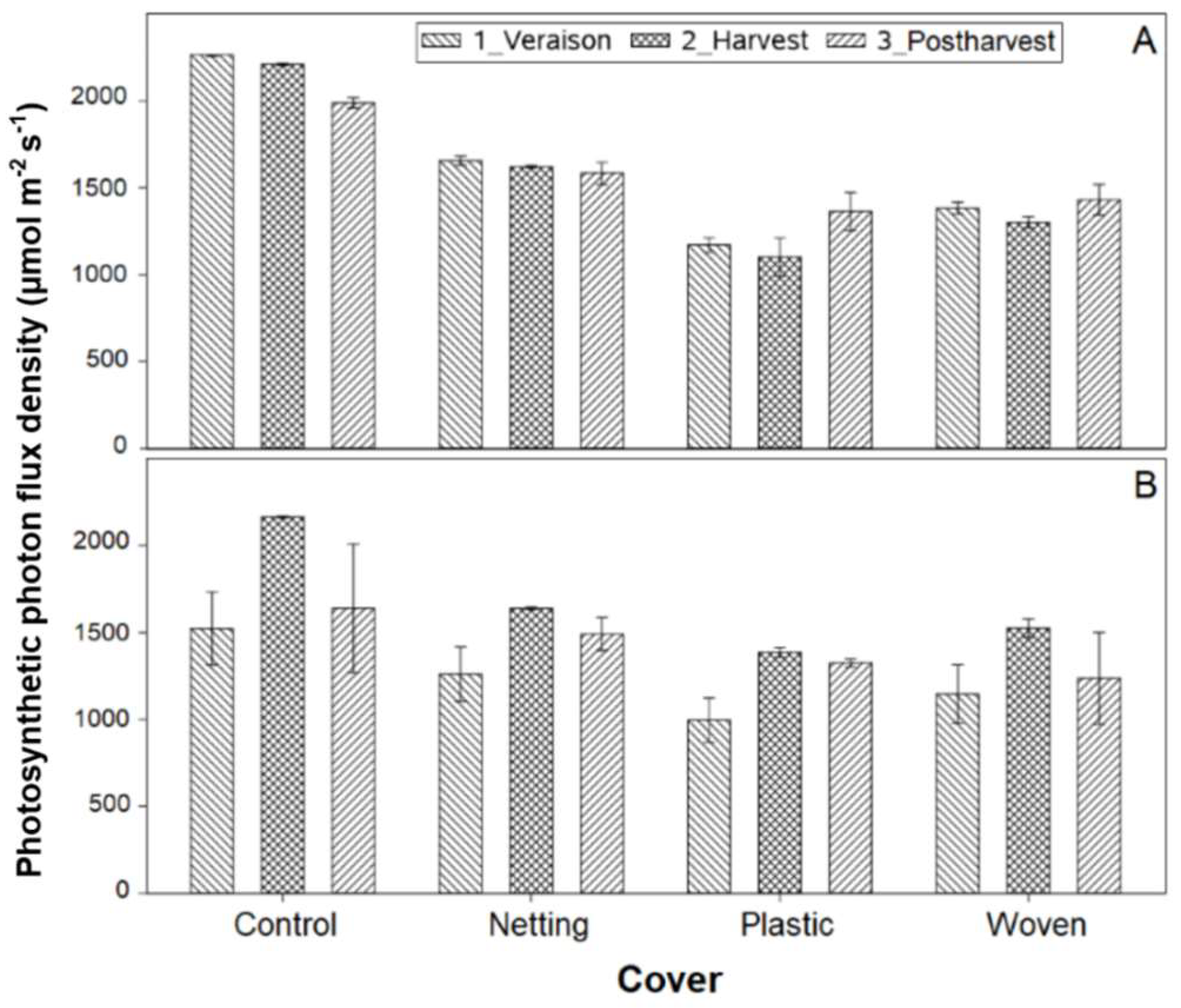
In the first season, control plants showed gs values at post-harvest that were similar to those of the cover treatments in Linares and Traiguén (
Table 3). However, in Linares, plants under the plastic cover showed a 45% reduction in gs compared with the woven and net covers. Despite the lack of differences in gs between covered and uncovered plants in both study sites, plants from the Linares orchard exhibited gs values at post-harvest that were higher than those reported in the following season from veraison to post-harvest. In the second season, there were no differences in gs between treatments at veraison and harvest. At post-harvest, the netting and plastic cover had higher gs values than the control. In Traiguén, significant differences in gs were detected only at harvest, where plants covered with netting had 20% higher gs than those covered with woven fabric. In the first season there were no differences in gs between the varieties, but in the second season, ‘Top Shelf’ tended to have higher gs than ‘Blue Ribbon’. Differences for both cultivars were significant at veraison and postharvest.
Under deficit irrigation conditions, control plants showed a 40% reduction in gs 1 week after the cessation of irrigation (
Figure 6B). On the other hand, the cover treatments showed a 15% reduction in gs in response to the same irrigation regime. Once irrigation was resumed, the gs in all treatments returned to the same values exhibited before the application of deficit irrigation.
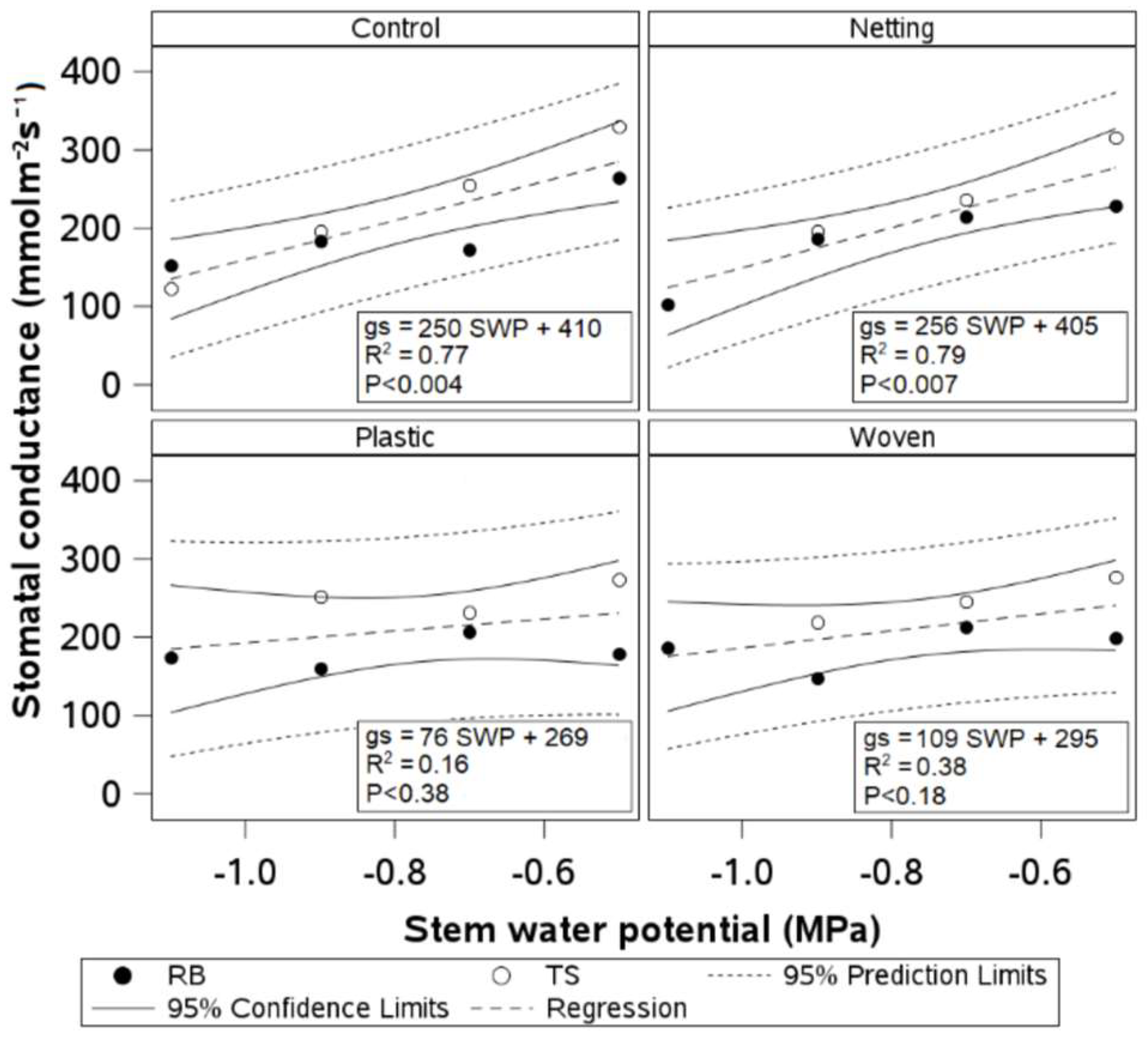
Regression analyses showed linear relationships between SWP and gs for the control and netting treatments, while no significant relationship was found for the woven and plastic treatments (
Figure 7). The covariance analysis showed the same slopes and intercepts for uncovered and netting-covered plants. Moreover, the coefficients of determination and
p-values were similar for uncovered and netting-covered plants (R
2~0.78;
p < 0.01).
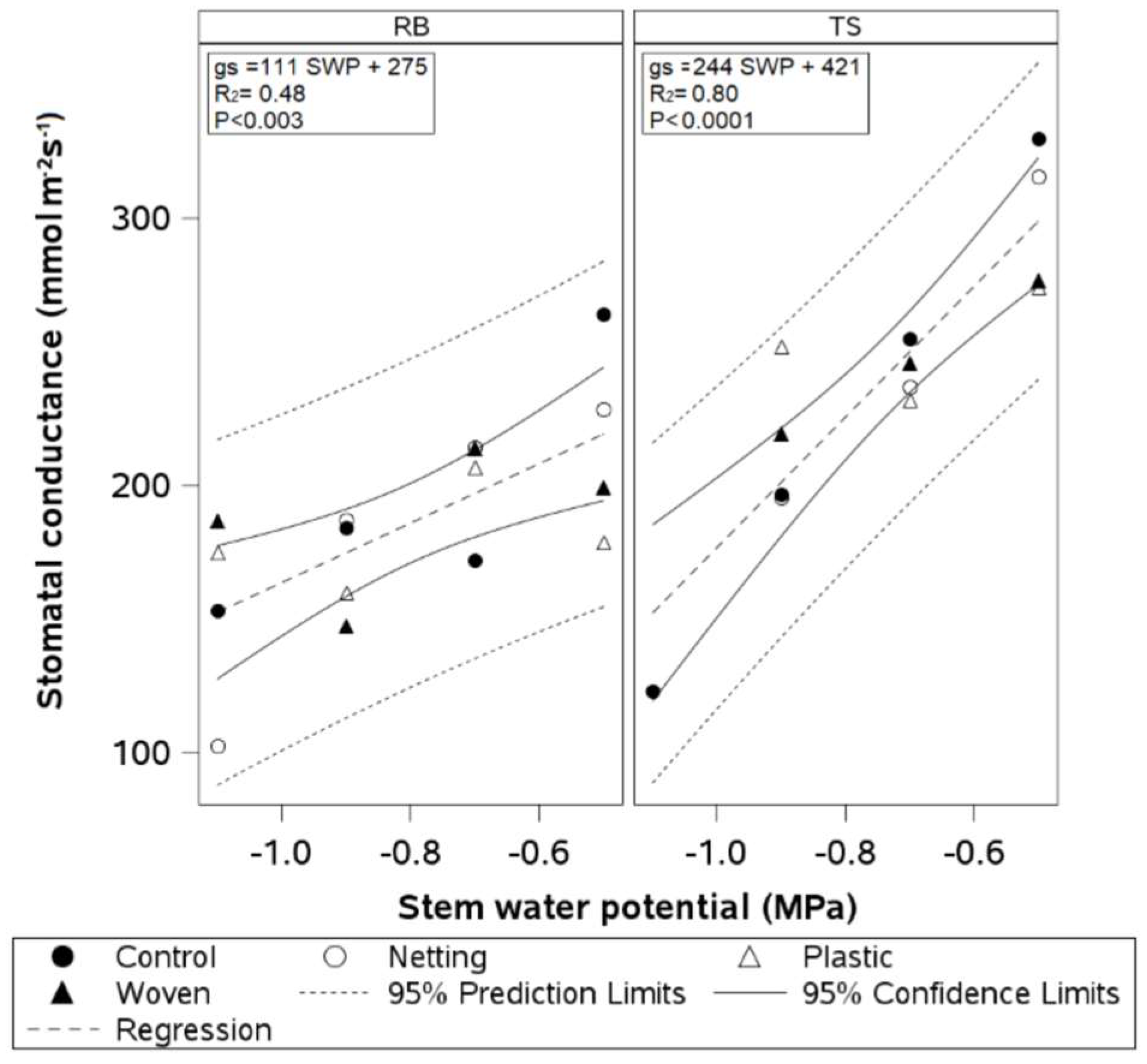
The regression analysis showed differences in the relationship between SWP and gs between cultivars (
Figure 8). The covariance analysis confirmed that the slopes and intercepts were different between cultivars. ‘Top Shelf’ had a significantly steeper slope for the resulting linear regression than ‘Blue Ribbon’ (TS slope = 244 vs. BR slope = 111). In addition, the intercepts differed between cultivars (TS intercept = 421 vs. BR intercept = 275). The coefficients of determination were higher, but the
p-values were lower, for ‘Top Shelf’ (TS R
2 = 0.80,
p < 0.0001 vs. BR R
2 = 0.48,
p < 0.003).
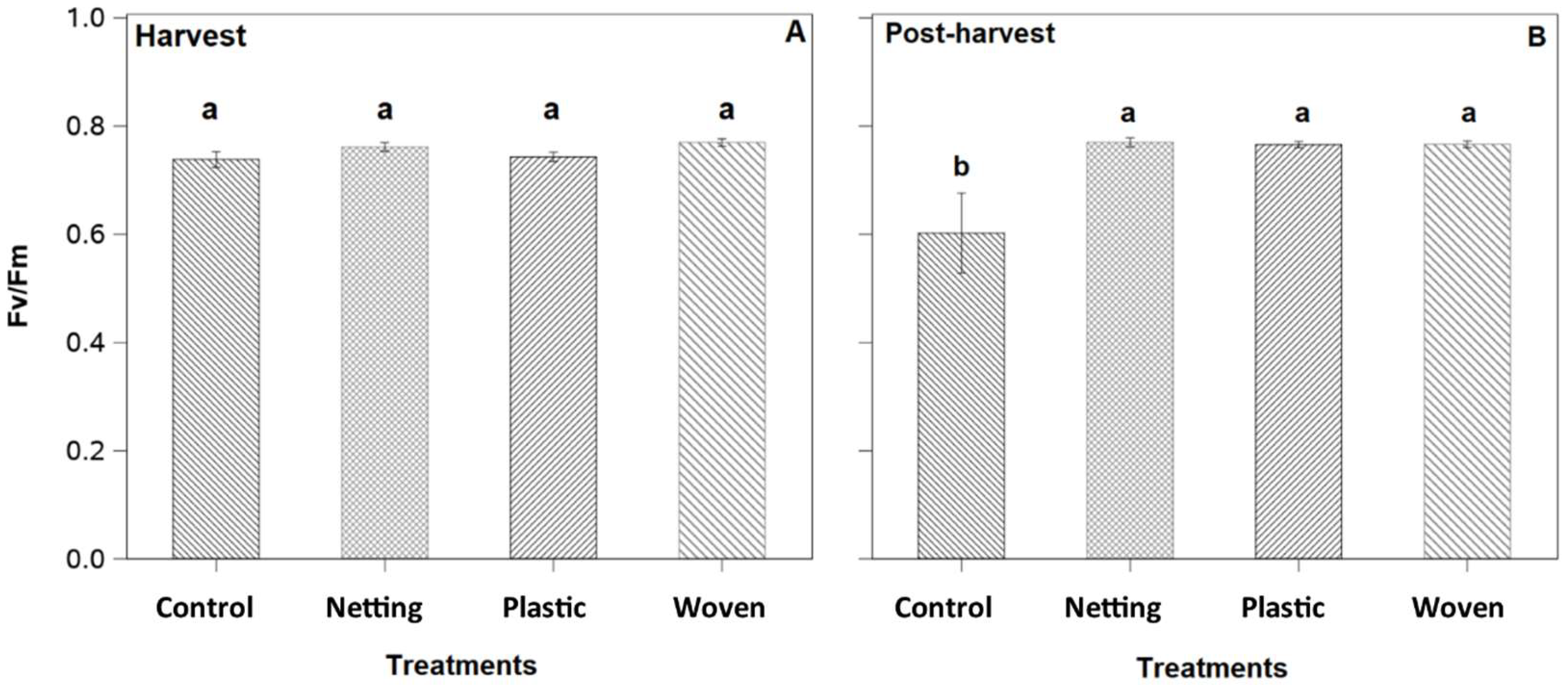
The maximum photochemical efficiency of photosystem II at harvest, estimated as
Fv/Fm, was similar for all treatments under conventional irrigation, maintaining values close to 0.8 (
Figure 9A). However, under deficit irrigation, control plants reduced the
Fv/Fm to 0.6, while the remaining treatments maintained the
Fv/Fm at 0.8 (
Figure 9B).
4. Discussion
Our results confirm that protected cultivation mitigates the adverse effects of transient drought events on the plant and soil water status and photochemistry of young blueberry plants. Under well-irrigated conditions, protected cultivation reduced desiccation in the upper soil layers (10 to 20 cm depth) by between 30% and 40% in Linares and Traiguén, respectively (
Table 1). While the shallow soil under cover was maintained within the range of available water capacity in both locations, the shallow soil without cover was close to the permanent wilting point. These results are consistent with previous studies in which protected cultivation using several cover materials conserved significant amounts of soil water [
28]. The higher water content in the upper soil layers, especially at a 10 cm depth, under coverings was attributed to a reduction in plant evapotranspiration, as has also been reported for citrus orchards [
10,
29], grapevines [
30], and apple trees [
31,
32]. In Linares and Traiguén, global solar radiation (
Figure 2A,B) and PPFD (
Figure 3A,B) decreased by between 10 and 20%, and 25 and 46% under the different coverings, respectively. Therefore, the lower soil desiccation under covered conditions was associated with a substantial reduction in the amount of incident global solar radiation and not in the atmospheric evaporative demand, since the differences in VPD between treatments were minimal in both orchards (±0.1 kPa) (
Figure 4A,B). Although fruit crops under protected cultivation have often shown an increase in vegetative growth rates [
12,
19,
33], LAI at harvest tended to be similarly lower in covered and uncovered plants. These results suggest that differences in soil desiccation are unlikely to be due to differences in canopy size.
The relationship between midday values of SWP and VPD air differed between sites and materials. In Traiguén, no relationship was observed between the two variables, regardless of the covering treatment (data not shown). In contrast, in Linares, SWP and VPD were linearly related for all coverings except plastic (
Figure 5). During the period of maximum water demand (December–January), global radiation (
Figure 2A,B) and minimum VPD (
Figure 4A,B) were 30% and 60% lower in Traiguén than in Linares, respectively. These results suggest that atmospheric water demand is more determinant of water stress for blueberry plants in locations where the driving forces of evapotranspiration are high. In Linares, the highest coefficient of determination (R
2) for the linear regression between midday values of SWP and VPD was found under uncovered conditions (R
2 = 0.72), but the analysis of covariance showed no differences in slopes and intercepts between covering treatments. Similar results were obtained in lime trees, comparing plants covered with shade netting and plants grown under uncovered conditions [
34]. The linear regression analysis (R
2 = 0.58;
p ≤ 0.0001) showed an SWP difference of 0.4 MPa for contrasting values of atmospheric water demand (VPD between 1.5 and 3.5 kPa) (
Figure 5). Well-watered plants exhibited midday SWP values, between −0.8 and −0.5 MPa, that have been associated with moderate to severe levels of water stress in previous blueberry studies [
14], highlighting the need to evaluate VPD when using SWP to measure plant water stress. Although there is no clear explanation for the lack of a relationship observed under the plastic cover, waterproof and non-porous materials such as plastic may cause a potential reduction in the vertical wind flow. Wind speed is a major determinant of boundary layer thickness in canopies and varies dramatically with height (up to 90% across the plant canopy) [
35]. Unfortunately, the installation height of the meteorological stations and the low frequency of measurements prevented us from capturing the potential changes in wind properties at various altitudes.
All coverings tended to maintain a higher SWP under well-irrigated conditions, but covered plants under plastic often exhibited the most consistent difference in water stress compared with uncovered plants at both sites (
Table 2). However, this effect was of physiological importance only under a moderate or high water stress severity (SWP < −0.8 MPa) [
14], which occurred at the end of the growing season (when soil water was mostly depleted) or when irrigation was withheld for 1 week in Linares. Uncovered plants reached moderate–severe levels of water stress (SWP of −1.0 MPa) when water application ceased for 1 week, while covered plants exhibited a plant water status representative of well-irrigated conditions (~0.85 MPa) (
Figure 6A) under high evaporative demand (VPD ~3.3 kPa) (
Figure 5). Maintaining adequate plant water status when water is scarce is particularly important in young blueberry orchards because drastic reductions in SWP can decrease vegetative growth due to a drop in expansive cell growth and net photosynthesis. For example, after 7 days of water deprivation, the leaf water potential and net photosynthesis in highbush blueberry plants were reduced by 35% and 48%, respectively [
18]. Changes in gas exchange capacity in response to water stress are not consistent in C3 plants because the relationship between stomatal conductance and plant water status is influenced by several factors, such as environmental conditions, genotype, or rootstock [
36]. In the present study, the comparison of g
s between cover treatments was less consistent under well-irrigated conditions (
Table 3) than under deficit irrigation (
Figure 6B). For well-irrigated conditions, higher g
s values in covered plants were only detected at post-harvest in Linares, while g
s was not affected by covering treatments at any measurement date in Traiguén (
Table 3). A recent study in highbush blueberry plants, cv. Legacy, reported that g
s was not affected by covering the orchard with polyethylene films of contrasting densities, despite the covering materials exhibiting high percentages of diffused transmittance [
37]. The weak and erratic response of gs to covering treatments may explain why
Fv/Fm was similarly high (~0.8) in all plants (
Figure 9A), indicating an optimum PSII efficiency. These results are in disagreement with previous studies in blueberry [
7], kiwifruit [
9], and European hazelnut [
19], which have measured higher PSII efficiency in covered plants under well-irrigated conditions.
Plants covered with plastic and netting had almost 50% higher g
s than uncovered plants after 1 week of water restriction (
Figure 6B). Previous studies have reported a very close relationship between water stress and gs in blueberry plants under open-field conditions, where a slight reduction in plant water status (±0.2 MPa) induced a 40% decrease in gs [
15,
18]. Although all cover materials tended to maintain high gs at moderate levels of water stress, the regression analyses showed that the stomatal response varied between cover treatments (
Figure 7). Uncovered and netting-covered plants showed a nearly 30% decrease in gs when the SWP reached −0.9 MPa. In contrast, woven and plastic-covered plants showed virtually no variation in gs at similar levels of water stress. In kiwifruit, plants covered with a low-density transparent plastic film maintained relatively stable gs under severe water stress (−1.3 MPa), while uncovered plants showed a 20% reduction in gs for a similar plant water status. These results suggest that waterproof covers, such as woven and plastic covers, may induce anisohydric behavior in blueberry plants and other fruit crops. Some plants (e.g.,
Acacia aptaneura) can maintain high values of gs at low water potentials (−1.8 MPa), which is probably due to the maintenance of high turgor in cell guards via osmotic adjustment [
38]. In a greenhouse experiment, which represents another type of protected cultivation, several blueberry cultivars showed significantly higher proline levels in the leaves of water-stressed plants [
27].
Blueberry plants are particularly susceptible to moderate water stress, since an SWP of −1.2 MPa may be enough to induce xylem embolism [
18]. Thus, effective stomatal closure in response to water stress is critical to limit irreversible plant dehydration. The anisohydric behavior of woven- and plastic-covered plants may be associated with a higher susceptibility to xylem vessel cavitation.
In this study, covered plants subjected to 1-week water restriction had higher PSII efficiencies, with an average
Fv/Fm of 0.8, while uncovered plants had an
Fv/Fm of 0.6 (
Figure 9B).
Fv/Fm values lower than 0.8 may indicate photo-inhibitory damage to PSII [
26] caused by excessive sunlight exposure of leaves with limited photosynthetic capacity due to stomatal closure. Nevertheless, the magnitude of the reduction in
Fv/Fm measured in uncovered plants under water stress was considerably less than that found in a previous study conducted under greenhouse conditions [
27], where
Fv/Fm values in deficit-irrigated plants ranged from 0 to 0.6 when significant differences were detected.
From an ecophysiological perspective, these results indicate that the combination of protected cultivation and deficit irrigation may increase water productivity in blueberry orchards. In covered plants subjected to deficit irrigation, there was a reduction in the amount of water applied that was not associated with possible changes in the rate of CO
2 diffusion to the chloroplasts and the electron transfer from water to plastoquinone, since moderate water stress had no impact on gs or
Fv/Fm. Similar results were reported for apple trees covered with 50% red shade netting [
11] and kiwifruit covered with low-density transparent plastic film [
9]. The fact that plants under plastic and woven covers had higher
Fv/Fm and gs in a moderate water stress situation may facilitate the generation of osmotically active solutes that maintain the osmotic adjustment of guard cells.
Under conventional irrigation, both cultivars showed a similar plant water status during most of the season, maintaining an SWP representative of well-irrigated conditions (between −0.7 and −0.6 MPa) (
Table 3). The lack of significant differences in SWP between cultivars of well-watered plants contrasted with a 30% higher gs in cv. Top shelf (
Table 3). The denser and deeper root system of cv. Top Shelf compared with cv. Blue Ribbon may have delayed the onset of moderate water stress. The regression and covariance analyses of the relationship between SWP and gs showed that cv. Top Shelf was more sensitive to water stress (steeper slope) than cv. Blue Ribbon (
Figure 8). These results suggest that cv. Blue Ribbon tended to be more anisohydric than cv. Top Shelf, which makes cv. Blue Ribbon more vulnerable to cavitation than cv. Top Shelf. Plant species have evolved plasticity in leaf characteristics to respond to changing environmental conditions. Studies in northern highbush blueberries have reported differences in stomatal density among cultivars. For example, cv. Brigitta had a lower stomatal density than cv. Star under identical environmental conditions [
39]. The latter could explain the persistent differences in gs observed between cultivars under conventional irrigation conditions.
Despite the differences in gs, both cultivars maintained a similar PSII efficiency, regardless of the severity of water stress. Despite cultivar differences in stomatal response, the CO2 diffusion rate would not have undergone sufficient changes to alter the photosynthetic efficiency of blueberry leaves. The maximum severity of water stress achieved in the present study (SWP of −1.0 MPa) may have induced a higher reduction in transpiration than photosynthesis in covered plants. Therefore, the severity of water stress reached in this study may serve as a reference SWP threshold in a regulated deficit irrigation program for covered blueberry orchards.
5. Conclusions
Our results show that all coverings altered plant–water relations in a young blueberry orchard by reducing solar radiation without causing substantial changes in atmospheric water demand. All covering materials delayed the occurrence of severe water stress for both cultivars when the evaporative demand was high. Nevertheless, the plastic covering showed the most consistent effect on plant and soil–water relations. In this context, the most important benefits of the plastic covering from bud break to senescence were (i) a delay in the onset of moderate–severe water stress, (ii) a reduction in soil desiccation, and (iii) lower leaf photoinhibition under severe water stress. Plastic-covered plants will tend to show higher water use efficiency than uncovered plants under water stress, which is particularly relevant in areas with Mediterranean or semi-arid climates affected by water scarcity.
The mechanisms of plant tolerance to water stress differed between treatments. Plants covered with woven and plastic materials tended to maintain stomata open from well-watered conditions to moderate water stress, reflecting an anisohydric behavior. In contrast, plants covered with netting and without any covering showed a rapid stomatal response to increasing levels of water stress, reflecting an isohydric behavior with a lower risk of xylem vessel cavitation. However, the impact of coverings on plant water stress was not high enough to economically justify the use of protected cultivation as the only method to cope with water scarcity. However, if protected cultivation is necessary for other purposes, plastic, woven, and netting materials can improve water stress tolerance.
cv. Top Shelf was more tolerant to higher levels of water stress than cv. Blue Ribbon. The higher stomatal sensitivity to water stress in cv. Top Shelf, in combination with a denser and deeper root system, had an impact on the occurrence of severe water stress and reduced the risk of xylem cavitation.
Finally, our results indicate that protected cultivation, using various covering materials, can improve water stress tolerance in irrigated blueberry orchards. The greater tolerance to water stress detected in covered orchards may save high amounts of irrigation water and improve water use efficiency.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}