
Sediment Fungal Communities of Constructed Wetlands Dominated by Zizania latifolia and Phragmites communis and Their Effect on Organic Pollutant Removal

Abstract
1. Introduction
2. Methods and Materials
2.1. Experimental Site and Treatment System
2.2. Water Quality Analysis
2.3. Plant Community Analysis
2.4. Microbial Community Analysis
2.5. Statistical Analysis
3. Results and Discussions
3.1. Pollutant Removal Rates
3.2. Effect of Plant Community on Removal Rate
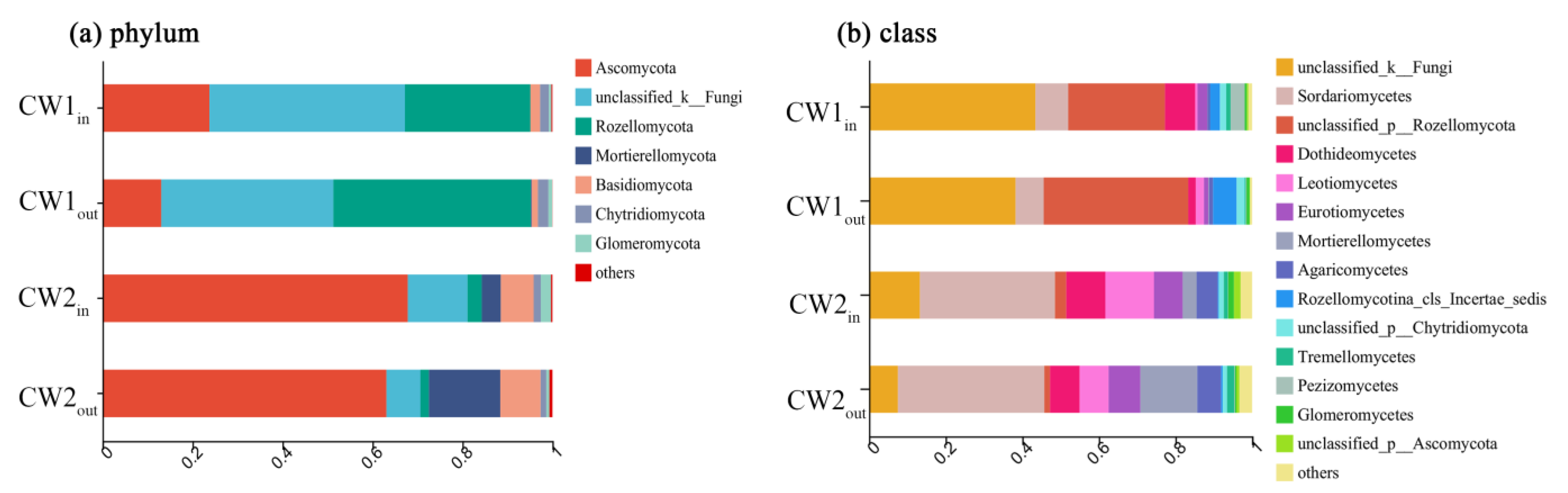
3.3. Variation in Fungal Abundance and Diversity
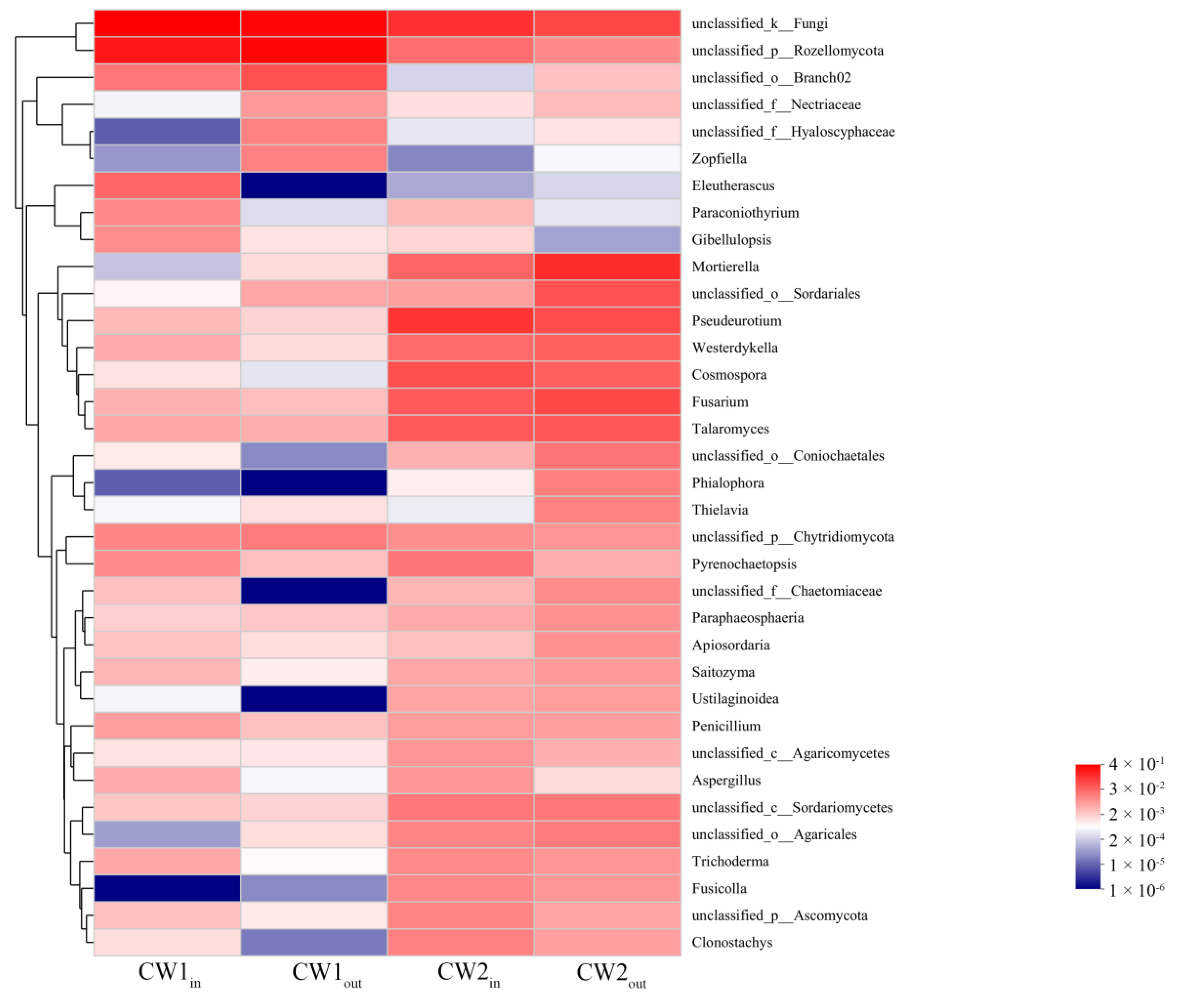
3.4. Correlation between Fugal Genera and Target Pollutants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sithamparanathan, E.; Sutton, N.B.; Rijnaarts, H.H.; Kujawa-Roeleveld, K. Controlling nitrogen removal processes in improved vertical flow constructed wetland with hydroponic materials: Effect of influent cod/n ratios. Water 2023, 15, 1074. [Google Scholar] [CrossRef]
- Spieles, D.J. Wetland construction, restoration, and integration: A comparative review. Land 2022, 11, 554. [Google Scholar] [CrossRef]
- Kurade, M.B.; Ha, Y.; Xiong, J.; Govindwar, S.P.; Jang, M.; Jeon, B. Phytoremediation as a green biotechnology tool for emerging environmental pollution: A step forward towards sustainable rehabilitation of the environment. Chem. Eng. J. 2021, 415, 129040. [Google Scholar] [CrossRef]
- Fujita, K.; Inui, H. How does the Cucurbitaceae family take up organic pollutants (POPs, PAHs, and PPCPs)? Rev. Environ. Sci. Bio/Technol. 2021, 20, 751–779. [Google Scholar] [CrossRef]
- Peña-Guzmán, C.; Ortiz-Gutierrez, B.E. Evaluation of three natural coagulant from moringa oleifera seed for the treatment of synthetic greywater. Civ. Eng. J. 2022, 8, 3842–3853. [Google Scholar] [CrossRef]
- Cheng, J.; Gu, J.; Tao, W.; Wang, P.; Liu, L.; Wang, C.; Li, Y.; Feng, X.; Qiu, G.; Cao, F. Edible fungus slag derived nitrogen-doped hierarchical porous carbon as a high-performance adsorbent for rapid removal of organic pollutants from water. Bioresour. Technol. 2019, 294, 122149. [Google Scholar] [CrossRef]
- Giovanella, P.; Vieira, G.A.; Otero, I.V.R.; Pellizzer, E.P.; de Jesus Fontes, B.; Sette, L.D. Metal and organic pollutants bioremediation by extremophile microorganisms. J. Hazard. Mater. 2020, 382, 121024. [Google Scholar] [CrossRef]
- Chan, S.S.; Khoo, K.S.; Chew, K.W.; Ling, T.C.; Show, P.L. Recent advances biodegradation and biosorption of organic compounds from wastewater: Microalgae-bacteria consortium—A review. Bioresour. Technol. 2022, 344, 126159. [Google Scholar] [CrossRef]
- Raji, V.; Packialakshmi, S. Assessing the wastewater pollutants retaining for a soil aquifer treatment using batch column experiments. Civ. Eng. J. 2022, 8, 1482–1491. [Google Scholar] [CrossRef]
- Ismail, W.N.W.; Syah, M.I.A.I.; Abd Muhet, N.H.; Bakar, N.H.A.; Yusop, H.M.; Samah, N.A. Adsorption behavior of heavy metal ions by hybrid inulin-TEOS for water treatment. Civ. Eng. J. 2022, 8, 1787–1798. [Google Scholar] [CrossRef]
- Ekeleme, A.C.; Ekwueme, B.N.; Agunwamba, J.C. Modeling contaminant transport of nitrate in soil column. Emerg. Sci. J. 2021, 5, 471–485. [Google Scholar] [CrossRef]
- Akpor, O.B.; Otohinoyi, D.; Olaolu, D.; Aderiye, B. Pollutants in wastewater effluents: Impacts and remediation processes. Int. J. Environ. Res. Earth Sci. 2014, 3, 50–59. [Google Scholar]
- Andersson, J.; Bastviken, S.K.; Tonderski, K. Free water surface wetlands for wastewater treatment in Sweden: Nitrogen and phosphorus removal. Water Sci. Technol. 2005, 51, 39–46. [Google Scholar] [CrossRef]
- Ansola, G.; Arroyo, P.; de Miera, L.E.S. Characterisation of the soil bacterial community structure and composition of natural and constructed wetlands. Sci. Total Environ. 2014, 473, 63–71. [Google Scholar] [CrossRef]
- Arroyo, P.; Ansola, G.; de Miera, L.E.S. Effects of substrate, vegetation and flow on arsenic and zinc removal efficiency and microbial diversity in constructed wetlands. Ecol. Eng. 2013, 51, 95–103. [Google Scholar] [CrossRef]
- Kim, S.; Kang, H.; Megonigal, J.P.; McCormick, M. Microbial activity and diversity vary with plant diversity and biomass in wetland ecosystems. Estuaries Coasts 2021, 45, 1434–1444. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, Y.; Wang, B.; Zhang, H.; Ding, J. Effects of stepwise adjustment of C/N during the start-up of submerged membrane bioreactors (SMBRs) on the aerobic denitrification of wastewater. Water 2021, 13, 3251. [Google Scholar] [CrossRef]
- Ge, Y.; Han, W.; Huang, C.; Wang, H.; Liu, D.; Chang, S.X.; Gu, B.; Zhang, C.; Gu, B.; Fan, X.; et al. Positive effects of plant diversity on nitrogen removal in microcosms of constructed wetlands with high ammonium loading. Ecol. Eng. 2015, 82, 614–623. [Google Scholar] [CrossRef]
- Weragoda, S.; Tanaka, N.; Jinadasa, K.; Sasaki, Y. Impact of influent inorganic nitrogen on nitrate removal efficiency of submerged plant microcosms. Chem. Ecol. 2009, 25, 179–188. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.Y.; Chen, X.; Huang, Z. Meta-analysis shows positive effects of plant diversity on microbial biomass and respiration. Nat. Commun. 2019, 10, 1332. [Google Scholar] [CrossRef]
- Chen, M.; He, S.; Li, J.; Hu, W.; Ma, Y.; Wu, L.; Gang, G. Co-occurrence patterns between bacterial and fungal communities in response to a vegetation gradient in a freshwater wetland. Can. J. Microbiol. 2019, 65, 722–737. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S.-I.; Zabed, H.M.; Okubo, A. Relationships between soil microbial diversity and plant community structure in seminatural grasslands. Grassl. Sci. 2008, 54, 117–124. [Google Scholar] [CrossRef]
- Comas, L.H.; Callahan, H.S.; Midford, P.E. Patterns in root traits of woody species hosting arbuscular and ectomycorrhizas: Implications for the evolution of belowground strategies. Ecol. Evol. 2014, 4, 2979–2990. [Google Scholar] [CrossRef] [PubMed]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root exudates regulate soil fungal community composition and diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Liang, W.; Wu, Z.; Cheng, S.; Zhou, Q.; Hu, H. Roles of substrate microorganisms and urease activities in wastewater purification in a constructed wetland system. Ecol. Eng. 2003, 21, 191–195. [Google Scholar] [CrossRef]
- Ligi, T.; Oopkaup, K.; Truu, M.; Preem, J.; Nõlvak, H.; Mitsch, W.J.; Mander, Ü.; Truu, J. Characterization of bacterial communities in soil and sediment of a created riverine wetland complex using high-throughput 16s rRNA amplicon sequencing. Ecol. Eng. 2014, 72, 56–66. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, D.; Bai, G.; Cao, T.; Liu, W.; Hu, Z.; Chen, D.; Qiu, D.; Wu, Z. Changes of microbial community structure during the initial stage of biological clogging in horizontal subsurface flow constructed wetlands. Bioresour. Technol. 2021, 337, 125405. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, D.; Dzakpasu, M.; Yang, Q.; Liu, Y.; Zhang, H.; Zhang, L.; Wang, X.C.; Zhao, Y. Effects of plants competition on critical bacteria selection and pollutants dynamics in a long-term polyculture constructed wetland. Bioresour. Technol. 2020, 316, 123927. [Google Scholar] [CrossRef]
- Li, W.; Liu, Y.; Tian, K.; Liang, Q.; Liu, H. Research on purification effect of Jian lake Zizania latifolia wetland lakefront zone in northwest of Yunnan plateau on nitrogen and phosphorus of agricultural non-point source. Agric. Sci. Technol. Hunan 2010, 11, 109–150. [Google Scholar]
- GB3838-2002; Environmental Quality Standards for Surface Water. Ministry of Ecology and Environment: Beijing, China, 2002.
- Hu, Q.; He, X.; Lu, X.; Zhang, X. Trend analysis of seasonal precipitation (1960–2013) in subregions of Hunan province, central south China using discrete wavelet transforms. J. Appl. Meteorol. Climatol. 2019, 58, 2159–2175. [Google Scholar] [CrossRef]
- Xiao, J.; Huang, J.; Huang, M.; Chen, M.; Wang, M. Application of basalt fiber in vertical flow constructed wetland for different pollution loads wastewater: Performance, substrate enzyme activity and microorganism community. Bioresour. Technol. 2020, 318, 124229. [Google Scholar] [CrossRef]
- Vymazal, J. Emergent plants used in free water surface constructed wetlands: A review. Ecol. Eng. 2013, 61, 582–592. [Google Scholar] [CrossRef]
- Zhang, B.; Song, X.; Zhang, Y.; Han, D.; Tang, C.; Yu, Y.; Ma, Y. Hydrochemical characteristics and water quality assessment of surface water and groundwater in Songnen plain, Northeast China. Water Res. 2012, 46, 2737–2748. [Google Scholar] [CrossRef]
- Zhai, X.; Piwpuan, N.; Arias, C.A.; Headley, T.; Brix, H. Can root exudates from emergent wetland plants fuel denitrification in subsurface flow constructed wetland systems? Ecol. Eng. 2013, 61, 555–563. [Google Scholar] [CrossRef]
- Wu, H.; Fan, J.; Zhang, J.; Ngo, H.H.; Guo, W.; Hu, Z.; Liang, S. Decentralized domestic wastewater treatment using intermittently aerated vertical flow constructed wetlands: Impact of influent strengths. Bioresour. Technol. 2015, 176, 163–168. [Google Scholar] [CrossRef]
- Allen, C.R.; Stein, O.R.; Hook, P.B.; Burr, M.D.; Parker, A.E.; Hafla, E.C. Temperature, plant species and residence time effects on nitrogen removal in model treatment wetlands. Water Sci. Technol. 2013, 68, 2337–2343. [Google Scholar] [CrossRef]
- Nivala, J.; Wallace, S.; Headley, T.; Kassa, K.; Brix, H.; van Afferden, M.; Müller, R. Oxygen transfer and consumption in subsurface flow treatment wetlands. Ecol. Eng. 2013, 61, 544–554. [Google Scholar] [CrossRef]
- Zhu, H.; Yan, B.; Xu, Y.; Guan, J.; Liu, S. Removal of nitrogen and cod in horizontal subsurface flow constructed wetlands under different influent C/N ratios. Ecol. Eng. 2014, 63, 58–63. [Google Scholar] [CrossRef]
- Liang, Y.; Zhu, H.; Bañuelos, G.; Yan, B.; Shutes, B.; Cheng, X.; Chen, X. Removal of nutrients in saline wastewater using constructed wetlands: Plant species, influent loads and salinity levels as influencing factors. Chemosphere 2017, 187, 52–61. [Google Scholar] [CrossRef]
- Jabłońska, E.; Winkowska, M.; Wiśniewska, M.; Geurts, J.; Zak, D.; Kotowski, W. Impact of vegetation harvesting on nutrient removal and plant biomass quality in wetland buffer zones. Hydrobiologia 2021, 848, 3273–3289. [Google Scholar] [CrossRef]
- Wei, F.; Shahid, M.J.; Alnusairi, G.S.; Afzal, M.; Khan, A.; El-Esawi, M.A.; Abbas, Z.; Wei, K.; Zaheer, I.E.; Rizwan, M. Implementation of floating treatment wetlands for textile wastewater management: A review. Sustainability 2020, 12, 5801. [Google Scholar] [CrossRef]
- Brisson, J.; Rodriguez, M.; Martin, C.A.; Proulx, R. Plant diversity effect on water quality in wetlands: A meta-analysis based on experimental systems. Ecol. Appl. 2020, 30, e02074. [Google Scholar] [CrossRef] [PubMed]
- Assress, H.A.; Selvarajan, R.; Nyoni, H.; Ntushelo, K.; Mamba, B.B.; Msagati, T.A. Diversity, co-occurrence and implications of fungal communities in wastewater treatment plants. Sci. Rep. 2019, 9, 14056. [Google Scholar] [CrossRef]
- Ping, Y.; Pan, X.; Li, W.; Wang, J.; Cui, L. The soil bacterial and fungal diversity were determined by the stoichiometric ratios of litter inputs: Evidence from a constructed wetland. Sci. Rep. 2019, 9, 13813. [Google Scholar] [CrossRef]
- Zhai, J.; Yan, G.; Cong, L.; Wu, Y.; Dai, L.; Zhang, Z.; Zhang, M. Assessing the effects of salinity and inundation on halophytes litter breakdown in yellow river delta wetland. Ecol. Indic. 2020, 115, 106405. [Google Scholar] [CrossRef]
- Kharitonov, S.; Shchegolkova, N.; Alexandrova, A.; Saynchuk, A.; Michel, P.; Maciejewski, K.; Gautier, M.; Gourdon, R.; Semenov, M.; Krasnov, G. Taxonomic diversity of fungi and bacteria in Azoé-NP® vertical flow constructed wetlands. Water 2022, 14, 698. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Chen, S.; Zhao, Z.; Feng, J.; Zhang, Z.; Lu, K.; Jia, J. Water bacterial and fungal community compositions associated with urban lakes, Xi’an, China. Int. J. Environ. Res. Public Health 2018, 15, 469. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, T.; Chen, S.; Yang, X.; Lv, K.; Sekar, R. Abundance and diversity of bacteria in oxygen minimum drinking water reservoir sediments studied by quantitative PCR and pyrosequencing. Microb. Ecol. 2015, 69, 618–629. [Google Scholar] [CrossRef]
- Held, B.W.; Salomon, C.E.; Blanchette, R.A. Diverse subterranean fungi of an underground iron ore mine. PLoS ONE 2020, 15, e0234208. [Google Scholar] [CrossRef]
- Matei, G.; Matei, S.; Mocanu, V. Assessing the role of soil microbial communities of natural forest ecosystem. EuroBiotech J. 2020, 4, 1–7. [Google Scholar] [CrossRef]
- Martino, E.; Prandi, L.; Fenoglio, I.; Bonfante, P.; Perotto, S.; Fubini, B. Soil fungal hyphae bind and attack asbestos fibers. Angew. Chem. 2003, 115, 229–232. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Park, S.W.; Pangging, M.; Lee, H.B. Molecular and morphological confirmation of three undescribed species of Mortierella from Korea. Mycobiology 2019, 47, 31–39. [Google Scholar] [CrossRef]
- Ozimek, E.; Jaroszuk-Ściseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyśkiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of indoleacetic acid, gibberellic acid and acc-deaminase by Mortierella strains promote winter wheat seedlings growth under different conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Zhao, X.; Zhang, X.; Zhao, Q.; Wang, X.; Li, Y. Restructured fungal community diversity and biological interactions promote metolachlor biodegradation in soil microbial fuel cells. Chemosphere 2019, 221, 735–749. [Google Scholar] [CrossRef]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.; Zhang, C.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land Degrad. Dev. 2018, 29, 1642–1651. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unit | Area (hm2) | Dominant Species | Density (Plants/m2) | Other Species | HRT | HLR | PLR |
|---|---|---|---|---|---|---|---|
| CW1 | 2.10 | Zizania latifolia | 5 | Trapa natansa, Canna indicaa, Acorus calamusa Oenanthe javanicab, Ipomoea trilobab, etc. | 42 | 0.0074 | 0.21 |
| CW2 | 2.14 | Phragmites communis | 5 | Myriophyllum verticillatuma, Hydrilla verticillateaErigeron canadensisb, Setaria viridisb, etc. | 38 | 0.0093 | 0.25 |
| Unit | Site | pH | COD (mg/L) | TN (mg/L) | TP (mg/L) | NH4+-N (mg/L) |
|---|---|---|---|---|---|---|
| CW1 | Inlet | 8.00 | 28.44 ± 7.86 | 1.42 ± 0.35 | 0.24 ± 0.04 | 0.51 ± 0.09 |
| Outlet | 7.67 | 18.38 ± 7.43 | 0.93 ± 0.32 | 0.16 ± 0.03 | 0.34 ± 0.12 | |
| Removal rate | R% | - | 35.9 ±13.1 | 35.2 ± 11.8 | 31.5 ± 13.6 | 33.9 ± 17.7 |
| CW2 | Inlet | 7.98 | 27.15 ± 2.75 | 1.30 ± 0.11 | 0.24 ± 0.02 | 0.45 ± 0.08 |
| Outlet | 7.70 | 19.56 ± 3.39 | 0.93 ± 0.20 | 0.19 ± 0.02 | 0.36 ± 0.11 | |
| Removal rate | R% | - | 27.6 ± 11.8 | 29.0 ± 11.9 | 23.0 ± 10.2 | 21.3 ± 13.3 |
| Class III | 6~9 | 20.0 | 1.0 | 0.2 | 1.0 |
| R (%) | COD | TN | TP | NH4+-N | ||||
|---|---|---|---|---|---|---|---|---|
| Index | CW1 | CW2 | CW1 | CW2 | CW1 | CW2 | CW1 | CW2 |
| Margalef (richness) | 0.938 | 0.946 | 0.989 | 0.957 | 0.961 | 0.827 | 0.964 | 0.923 |
| Simpson (diversity) | 0.994 | 0.970 | 0.965 | 0.984 | 0.957 | 0.887 | 0.907 | 0.911 |
| Shannon–Weiner (diversity) | 0.978 | 0.881 | 0.852 | 0.897 | 0.821 | 0.727 | 0.749 | 0.940 |
| Pielou (evenness) | 0.976 | 0.928 | 0.957 | 0.934 | 0.921 | 0.784 | 0.899 | 0.913 |
| Unit | Site | OTU | Chao | Shannon–Weiner | Good’s Coverage |
|---|---|---|---|---|---|
| Inlet | 2337 | 920.05 ± 197.35 a | 4.97 ± 0.11 a | 0.9988 | |
| CW1 | Outlet | 1017 | 350.13 ± 70.21 c | 3.95 ± 0.52 c | 0.9996 |
| Total | 3354 | ||||
| Inlet | 1881 | 627.41 ± 137.75 b | 4.49 ± 0.33 b | 0.9994 | |
| CW2 | Outlet | 1551 | 464.66 ± 95.55 bc | 4.08 ± 0.26 bc | 0.9996 |
| Total | 2432 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Huang, Z.; Li, H.; Huang, J.; Qin, X.; Wu, Z. Sediment Fungal Communities of Constructed Wetlands Dominated by Zizania latifolia and Phragmites communis and Their Effect on Organic Pollutant Removal. Water 2023, 15, 2291. https://doi.org/10.3390/w15122291
He Y, Huang Z, Li H, Huang J, Qin X, Wu Z. Sediment Fungal Communities of Constructed Wetlands Dominated by Zizania latifolia and Phragmites communis and Their Effect on Organic Pollutant Removal. Water. 2023; 15(12):2291. https://doi.org/10.3390/w15122291
Chicago/Turabian StyleHe, Yiting, Zhongliang Huang, Hui Li, Jing Huang, Xiaoli Qin, and Zijian Wu. 2023. "Sediment Fungal Communities of Constructed Wetlands Dominated by Zizania latifolia and Phragmites communis and Their Effect on Organic Pollutant Removal" Water 15, no. 12: 2291. https://doi.org/10.3390/w15122291
APA StyleHe, Y., Huang, Z., Li, H., Huang, J., Qin, X., & Wu, Z. (2023). Sediment Fungal Communities of Constructed Wetlands Dominated by Zizania latifolia and Phragmites communis and Their Effect on Organic Pollutant Removal. Water, 15(12), 2291. https://doi.org/10.3390/w15122291