
Removal of Fecal Indicator Bacteria by River Networks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
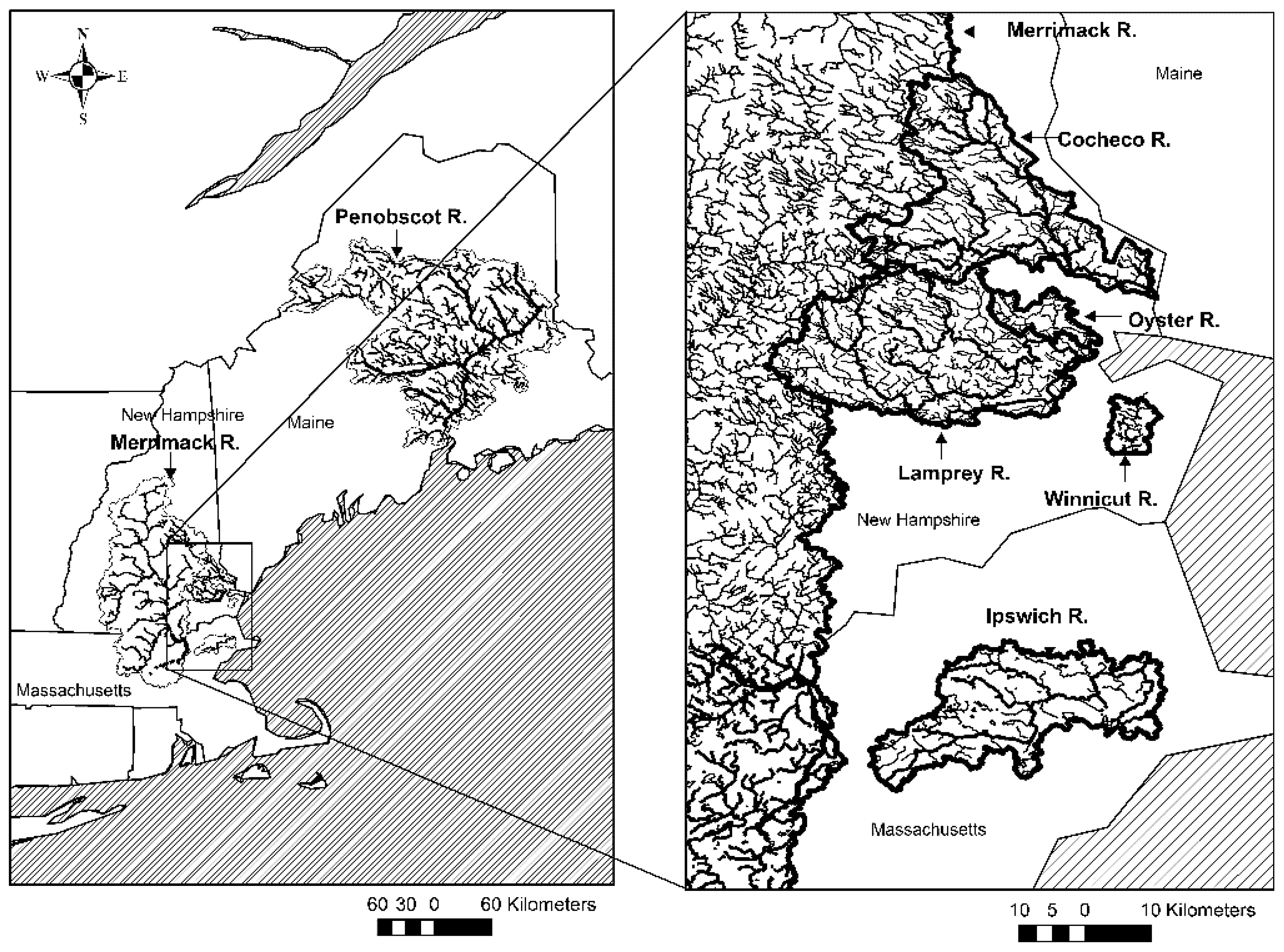
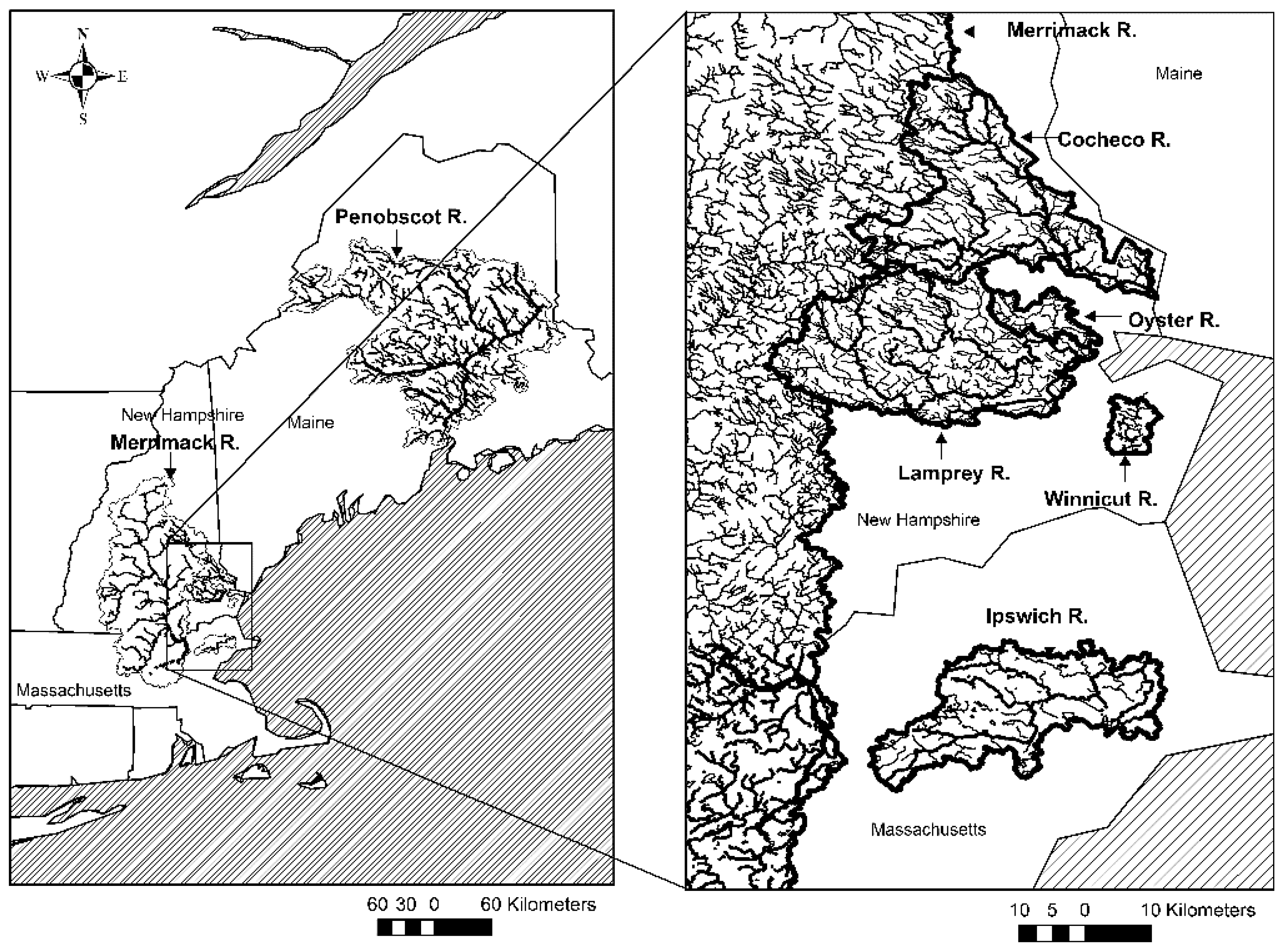
2.1. Study Sites
2.2. E. coli River Network Model Description
2.3. E. coli Model Parameterization and Testing
2.4. Hypothesis Testing
2.5. Skewness Index
2.6. E. coli Exceedance
3. Results
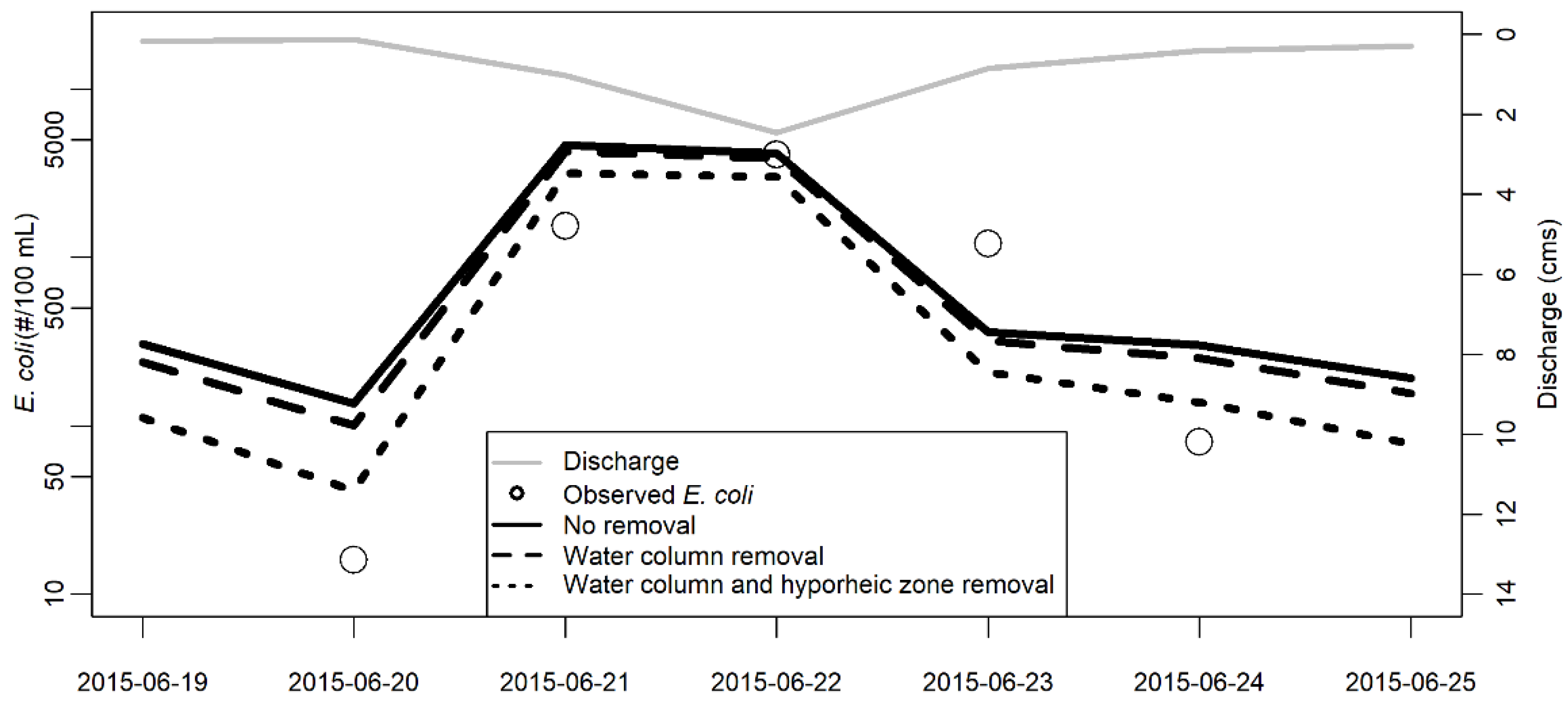
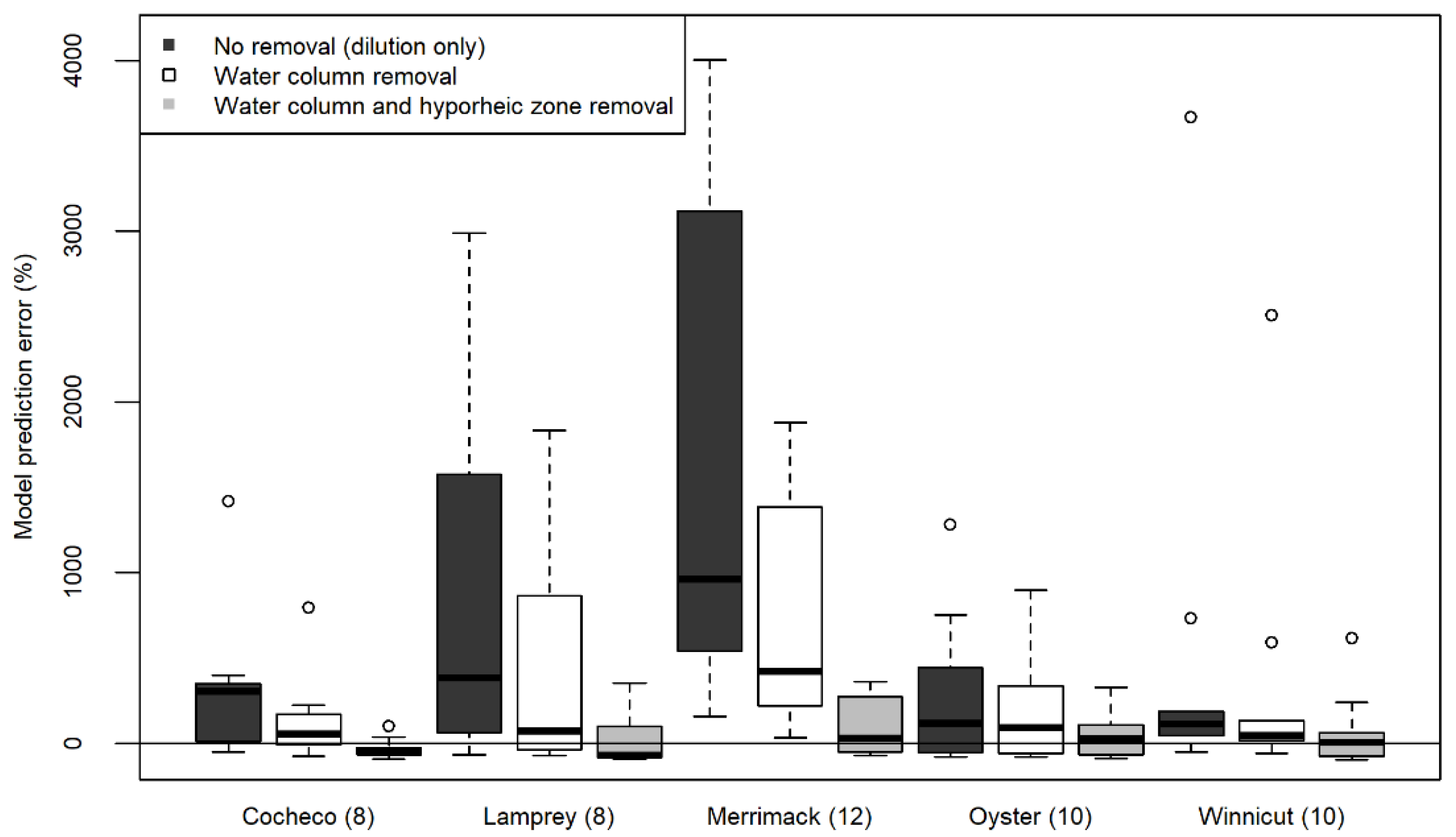
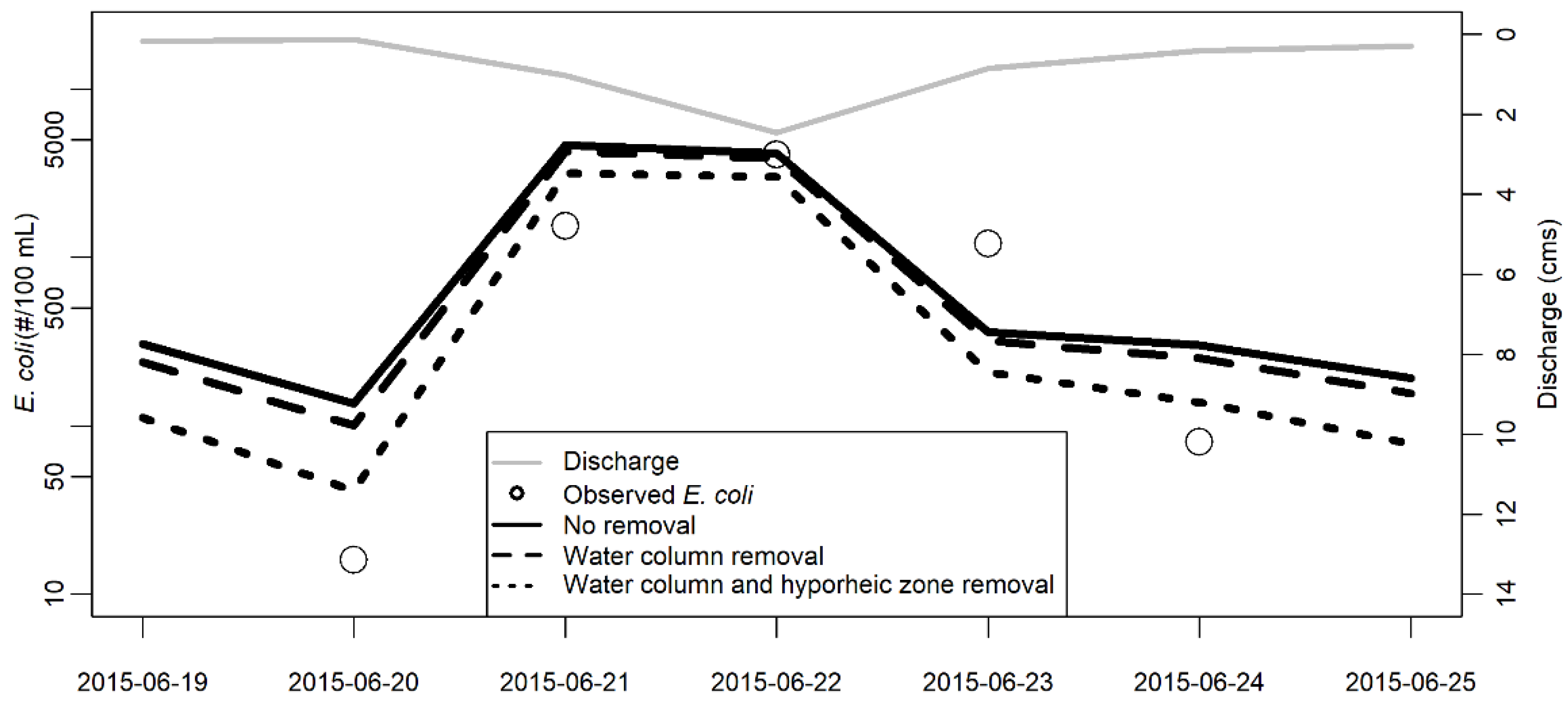
3.1. Model Validation
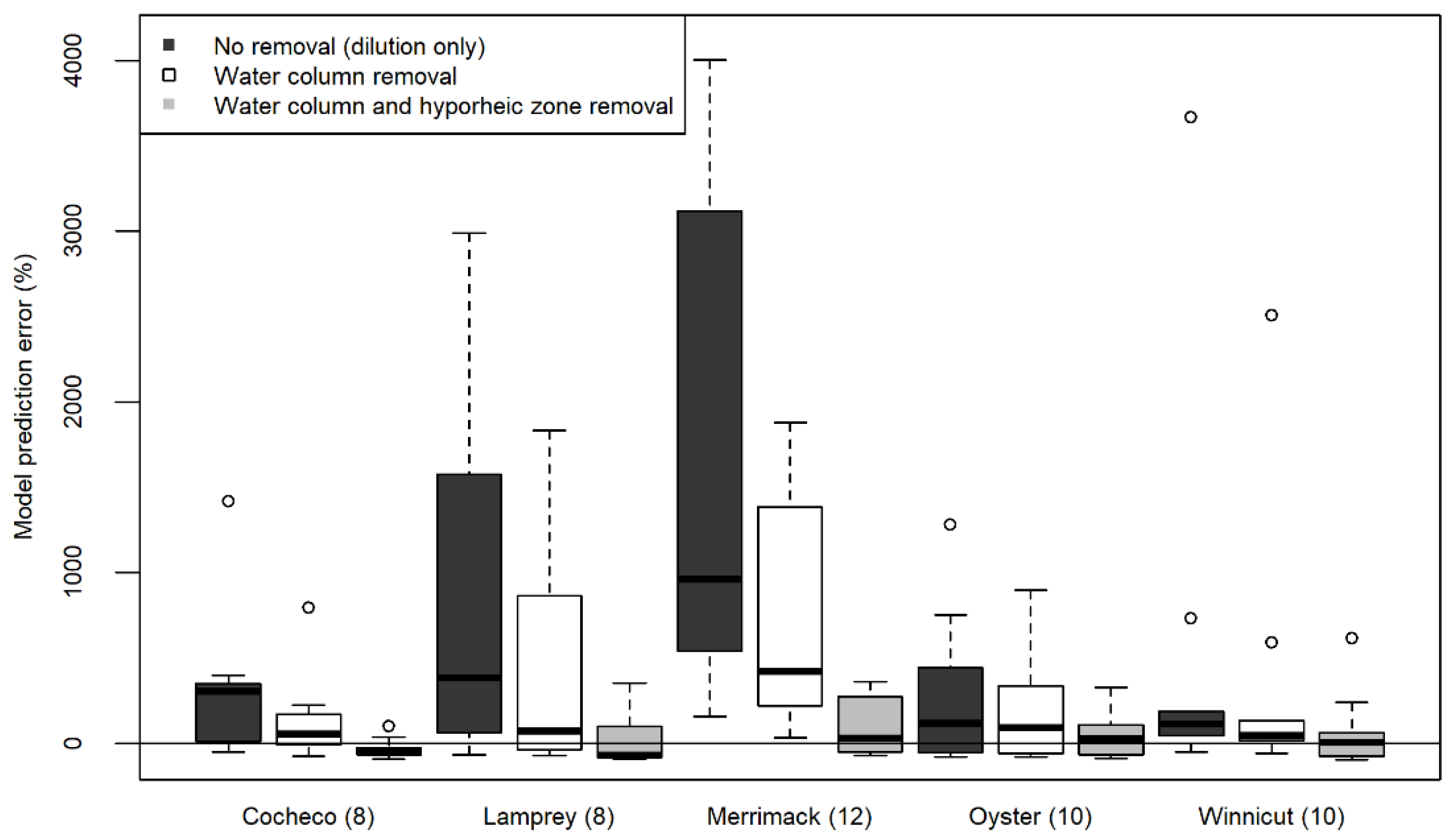
3.2. Sensitivity Analysis
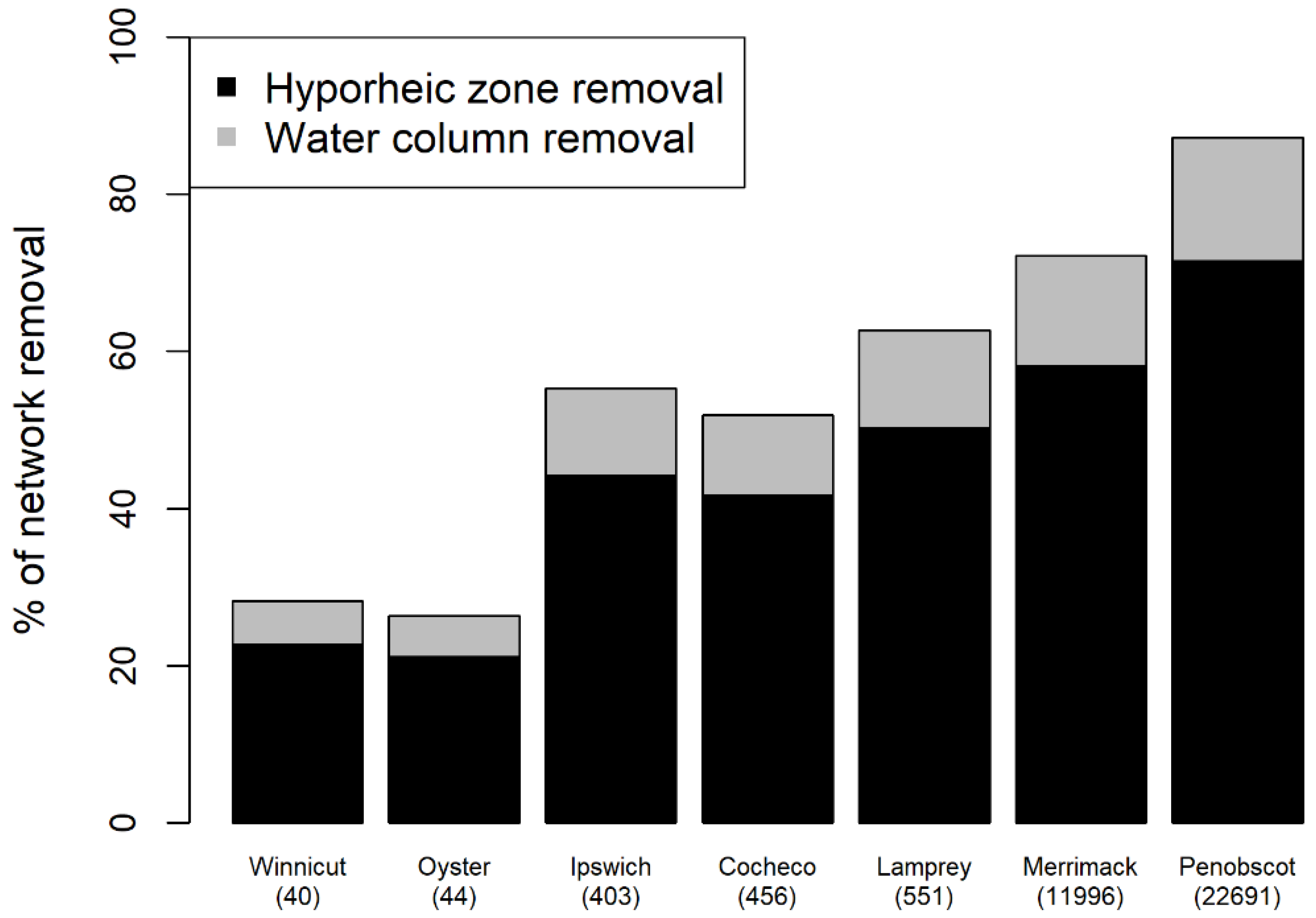
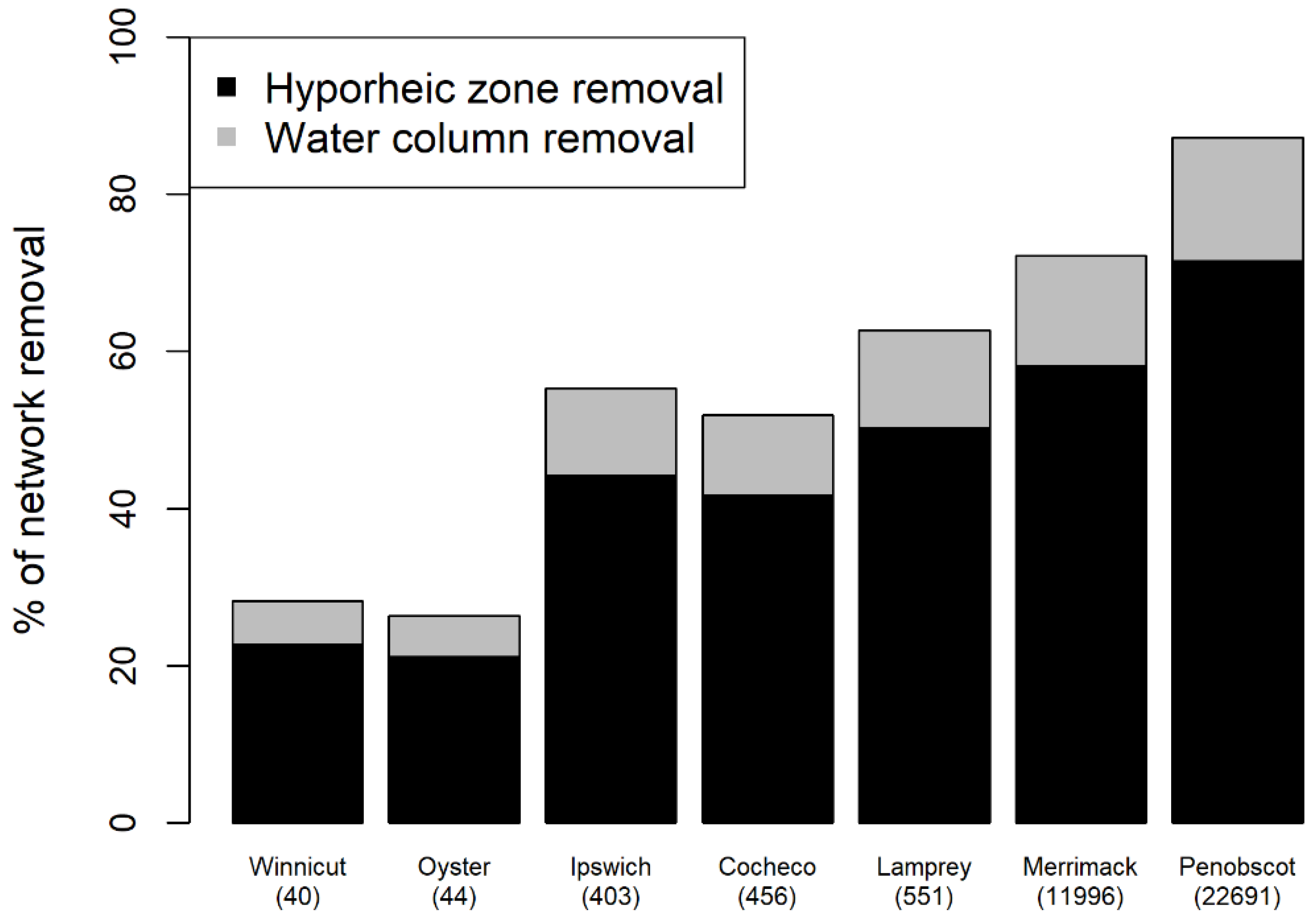
3.3. Whole River Network E. coli Removal Using Full Removal Scenario
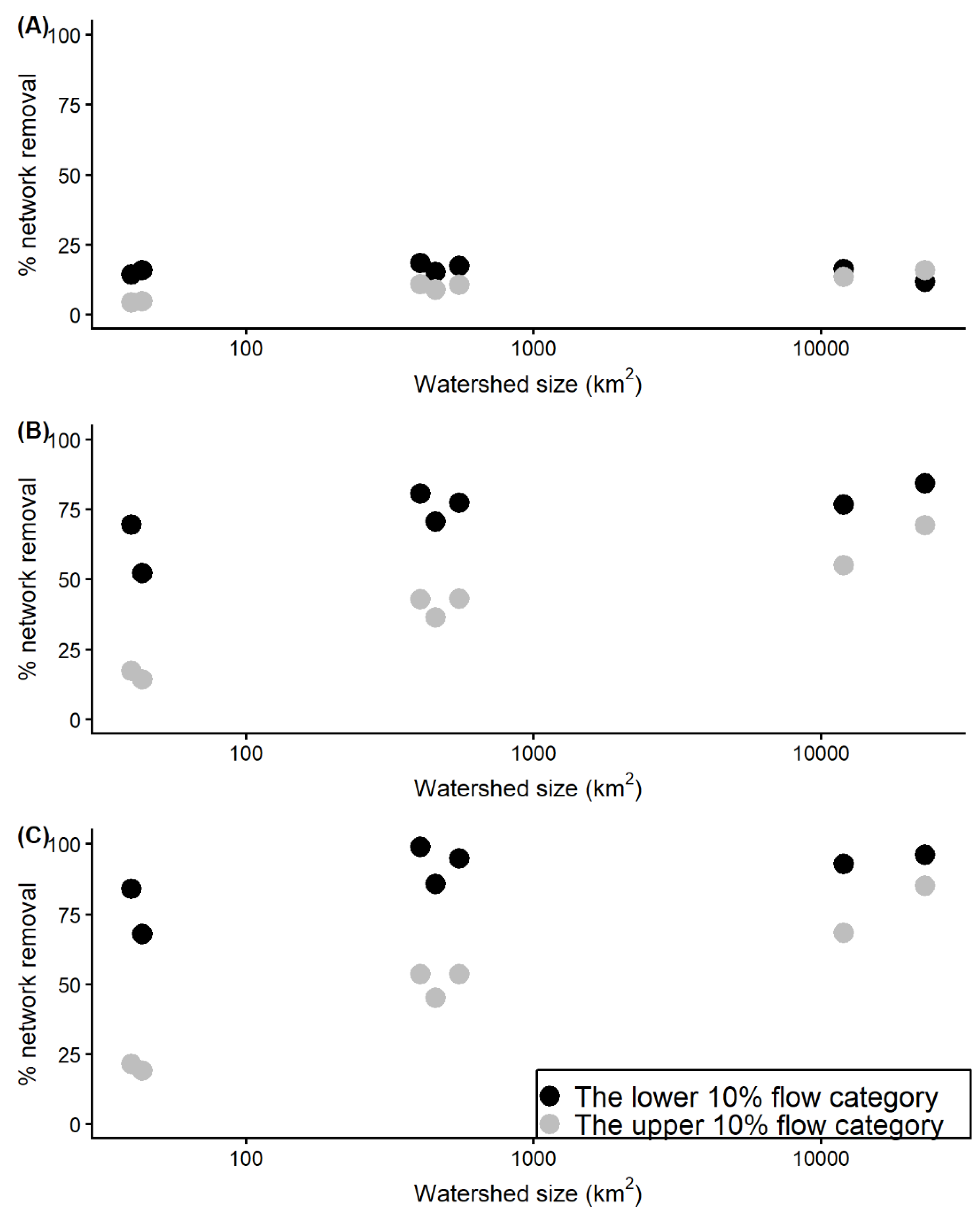
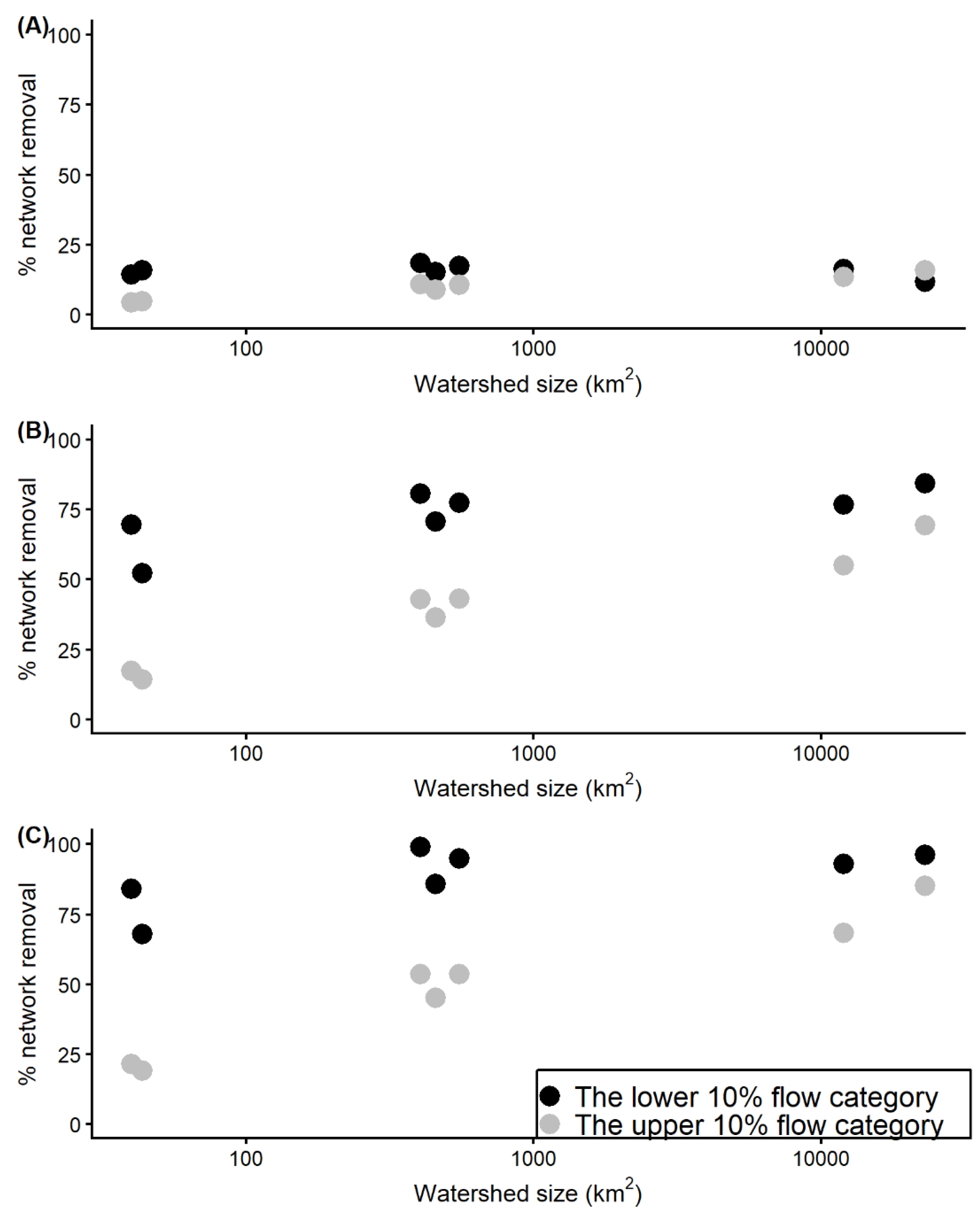
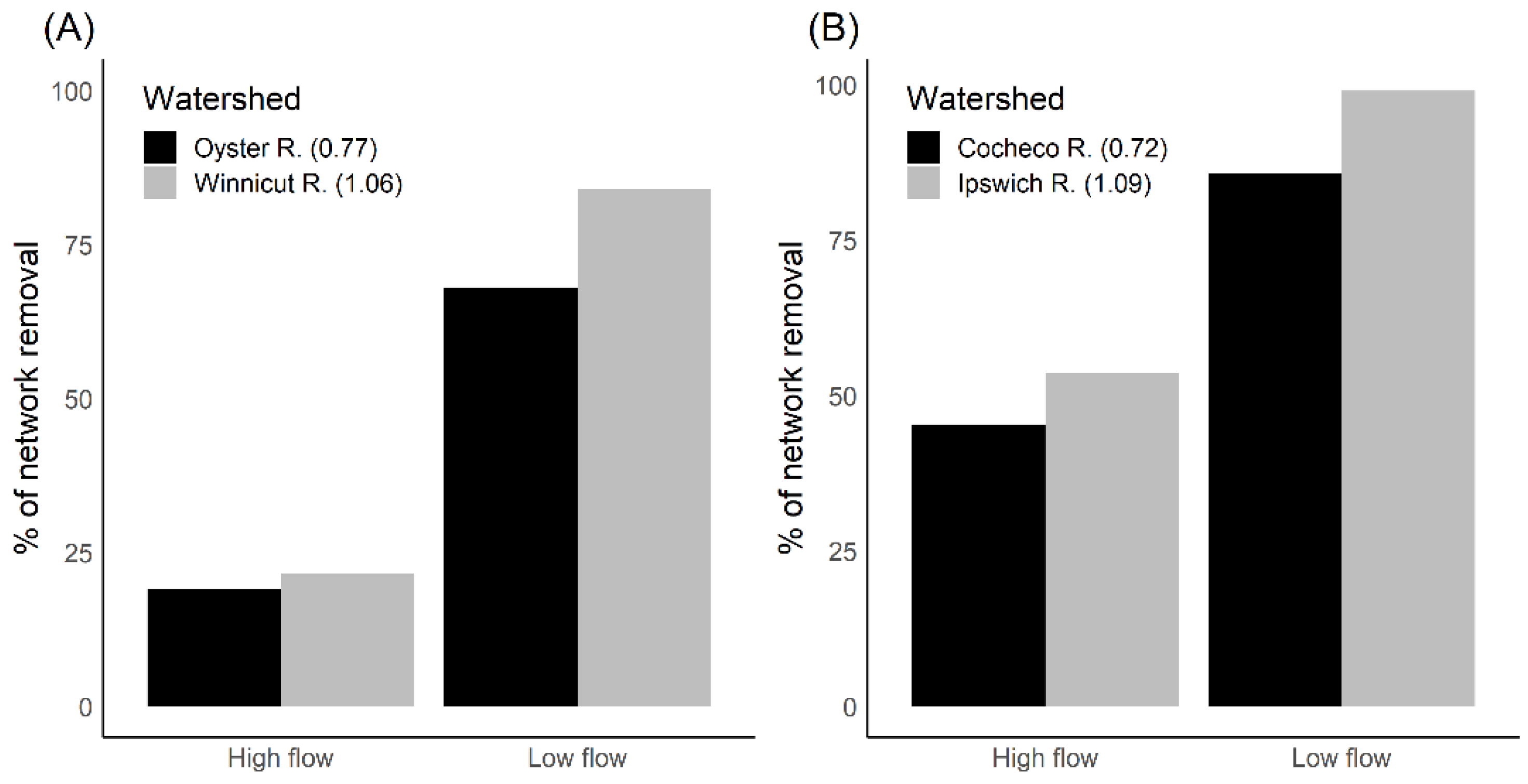
3.4. Effect of Watershed Size
3.5. Skewness Index Analysis
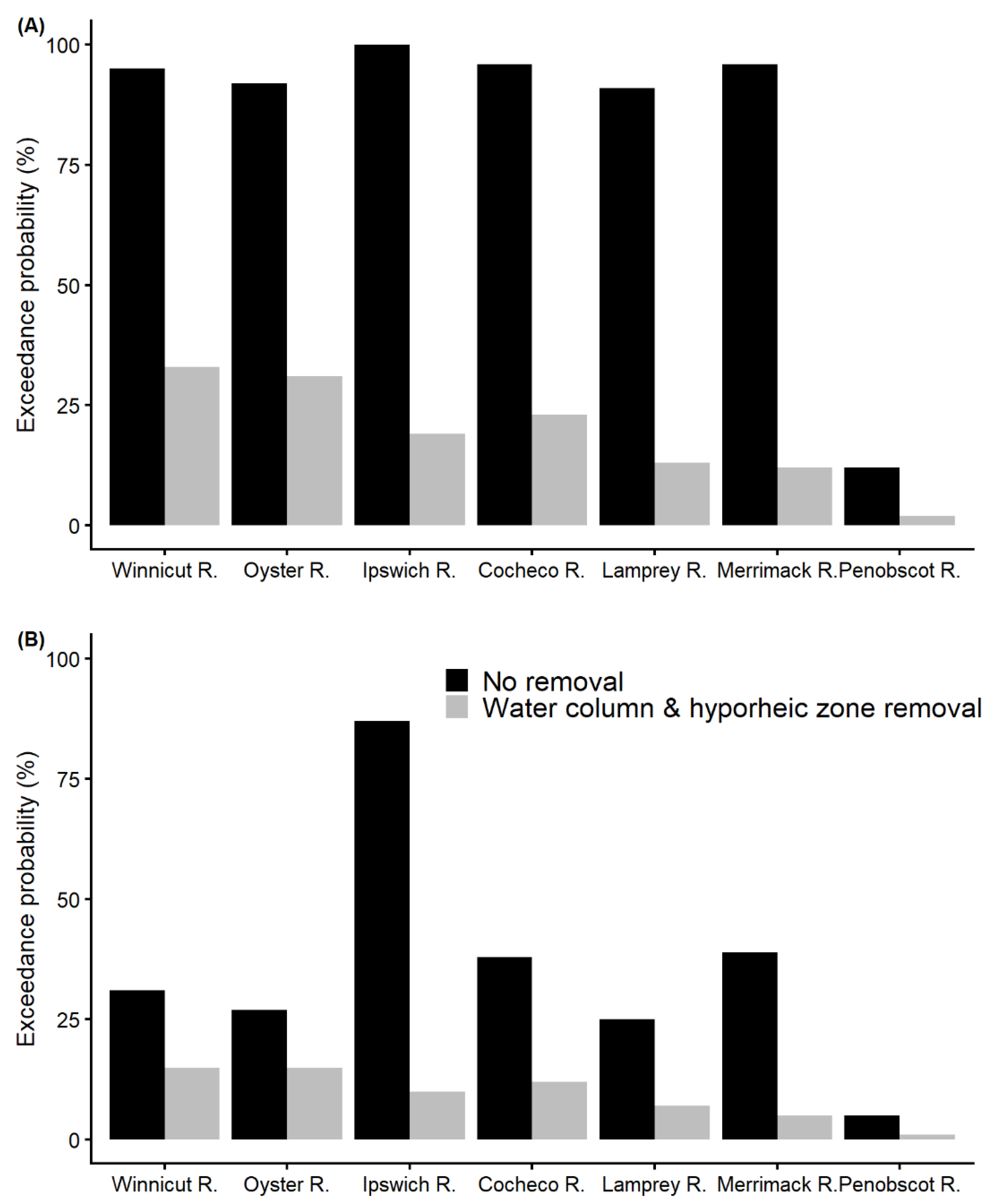
3.6. Prediction of E. coli Level Exceedance Probabilities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pal, M.; Ayele, Y.; Hadush, M.; Panigrahi, S.; Jadhav, V.J. Public health hazards due to unsafe drinking water. Air Water Borne Dis. 2018, 7, 2. [Google Scholar]
- Pandey, P.K.; Kass, P.H.; Soupir, M.L.; Biswas, S.; Singh, V.P. Contamination of water resources by pathogenic bacteria. Amb Express 2014, 4, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, A.; Oliver, D.M.; Bivins, A.; Sherchan, S.P.; Pitkänen, T. Bathing water quality monitoring practices in Europe and the United States. Int. J. Environ. Res. Public Health 2021, 18, 5513. [Google Scholar] [CrossRef] [PubMed]
- Bitton, G. Microbiology of Drinking Water Production and Distribution; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014. [Google Scholar]
- United States Environmental Protection Agency, National Cumulative TMDLs by Pollutant. National Summary of State Information. Available online: https://iaspub.epa.gov/waters10/attains_nation_cy.control?p_report_type=T#causes_303d (accessed on 9 June 2021).
- United States Environmental Protection Agency. Recreational Water Quality Criteria; Office of Water: Washington, DC, USA, 2012.
- WHO. WHO Recommendations on Scientific, Analytical and Epidemiological Developments Relevant to the Parameters for Bathing Water Quality in the Bathing Water Directive (2006/7/EC); World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Harwood, V.J.; Staley, C.; Badgley, B.D.; Borges, K.; Korajkic, A. Microbial source tracking markers for detection of fecal contamination in environmental waters: Relationships between pathogens and human health outcomes. FEMS Microbiol. Rev. 2014, 38, 1–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, J.K.; Snyder, B.J.; Oladeinde, A.; Spidle, D.; Berrang, M.E.; Meinersmann, R.J.; Oakley, B.; Sidle, R.C.; Sullivan, K.; Molina, M. Characterizing relationships among fecal indicator bacteria, microbial source tracking markers, and associated waterborne pathogen occurrence in stream water and sediments in a mixed land use watershed. Water Res. 2016, 101, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H.; Pachepsky, Y.A.; Kim, M.; Pyo, J.; Park, M.H.; Kim, Y.M.; Kim, J.W.; Kim, J.H. Modeling seasonal variability of fecal coliform in natural surface waters using the modified SWAT. J. Hydrol. 2016, 1, 377–385. [Google Scholar] [CrossRef]
- Acuña, V.; Von Schiller, D.; García-Galán, M.J. Occurrence and in-stream attenuation of wastewater-derived pharmaceuticals in Iberian rivers. Sci. Total Environ. 2015, 503–504, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Acuña, V.; Casellas, M.; Font, C.; Romero, F.; Sabater, S. Nutrient attenuation dynamics in effluent dominated watercourses. Water Res. 2019, 160, 330–338. [Google Scholar] [CrossRef]
- Willcock, S.; Parker, A.; Wilson, C.; Brewer, T.; Bundhoo, D.; Cooper, S.; Lynch, K.; Mekala, S.; Mishra, P.P.; Rey, D.; et al. Nature provides valuable sanitation services. One Earth 2021, 4, 192–201. [Google Scholar] [CrossRef]
- Herzog, S.P.; Higgins, C.P.; McCray, J.E. Engineered streambeds for induced hyporheic flow: Enhanced removal of nutrients, pathogens, and metals from urban streams. J. Environ. Eng. 2016, 142, 04015053. [Google Scholar] [CrossRef]
- Nguyen, H.T.M.; Le, Q.T.P.; Garnier, J.; Janeau, J.L.; Rochelle-Newall, E. Seasonal variability of faecal indicator bacteria numbers and die-off rates in the Red River basin, North Viet Nam. Sci. Rep. 2016, 6, 21644. [Google Scholar] [CrossRef] [PubMed]
- Drummond, J.D.; Davies-Colley, R.J.; Stott, R.; Sukias, J.P.; Nagels, J.W.; Sharp, A.; Packman, A.I. Microbial transport, retention, and inactivation in streams: A combined experimental and stochastic modeling approach. Environ. Sci. Technol. 2015, 49, 7825–7833. [Google Scholar] [CrossRef] [PubMed]
- Whitman, R.L.; Nevers, M.B.; Korinek, G.C.; Byappanahalli, M.N. Solar and temporal effects on Escherichia coli concentration at a lake Michigan swimming beach. Appl. Environ. Microbiol. 2004, 70, 4276–4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, J.; Paracchini, M.L.; Zulian, G. A European assessment of the provision of ecosystem services. In Towards and Atlas of Ecosystem Services; Joint Research Centre, IES: Ispra, Italy, 2011. [Google Scholar]
- Terrado, M.; Sabater, S.; Chaplin-Kramer, B.; Mandle, L.; Ziv, G.; Acuña, V. Model development for the assessment of terrestrial and aquatic habitat quality in conservation planning. Sci. Total Environ. 2016, 540, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Doyle, M.W.; Stanley, E.H.; Strayer, D.L.; Jacobson, R.B.; Schmidt, J.C. Effective discharge analysis of ecological processes in streams. Water Resour. Res. 2005, 41, W11411. [Google Scholar] [CrossRef] [Green Version]
- Lenzi, M.; Mao, L.; Comiti, F. Effective discharge for sediment transport in a mountain river: Computational approaches and geomorphic effectiveness. J. Hydrol. 2006, 326, 257–276. [Google Scholar] [CrossRef]
- Raymond, P.A.; Saiers, J.E.; Sobczak, W.V. Hydrological and biogeochemical controls on watershed dissolved organic matter transport: Pulse-shunt concept. Ecology 2016, 97, 5–16. [Google Scholar] [CrossRef]
- Noble, R.T.; Lee, I.M.; Schiff, K.C. Inactivation of indicator micro-organisms from various sources of faecal contamination in seawater and freshwater. J. Appl. Microbiol. 2004, 96, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Sinton, L.W.; Hall, C.H.; Lynch, P.A.; Davies-Colley, R.J. Sunlight inactivation of fecal indicator bacteria and bacteriophages from waste stabilization pond effluent in fresh and saline waters. Appl. Environ. Microbiol. 2002, 68, 1122–1131. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, E.; Åström, J.; Pettersson, T.J.; Bergstedt, O.; Hermansson, M. Estimation of pathogen concentrations in a drinking water source using hydrodynamic modelling and microbial source tracking. J. Water Health 2012, 10, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.J.; Wollheim, W.M.; Gooseff, M.N.; Briggs, M.A.; Jacobs, J.M.; Peterson, B.J.; Hopkinson, C.S. Separation of river network-scale nitrogen removal among the main channel and two transient storage compartments. Water Resour. Res. 2011, 47, W00J10. [Google Scholar] [CrossRef] [Green Version]
- Wollheim, W.M.; Peterson, B.J.; Thomas, S.; Hopkinson, C.; Vörösmarty, C. Dynamics of N removal over annual time periods in a suburban river network. J. Geophys. Res. Biogeosci. 2008, 113, G03038. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H.; Pachepsky, Y.A.; Oliver, D.M.; Muirhead, R.W.; Park, Y.; Quilliam, R.S.; Shelton, D.R. Modeling fate and transport of fecally-derived microorganisms at the watershed scale: State of the science and future opportunities. Water Res. 2016, 100, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Sophocleous, M. Interactions between groundwater and surface water: The state of the science. Hydrogeol. J. 2002, 10, 52–67. [Google Scholar] [CrossRef]
- Surbeck, C.Q.; Jiang, S.C.; Grant, S.B. Ecological control of fecal indicator bacteria in an urban stream. Environ. Sci. Technol. 2010, 44, 631–637. [Google Scholar] [CrossRef]
- De Brauwere, A.; Ouattara, N.K.; Servais, P. Modeling fecal indicator bacteria concentrations in natural surface waters: A review. Crit. Rev. Environ. Sci. Technol. 2014, 44, 2380–2453. [Google Scholar] [CrossRef] [Green Version]
- Coffey, R.; Cummins, E.; Bhreathnach, N.; Flaherty, V.; Cormican, M. Development of a pathogen transport model for Irish catchments using SWAT. Agric. Water Manag. 2010, 97, 101–111. [Google Scholar] [CrossRef]
- Niazi, M.; Obropta, C.; Miskewitz, R. Pathogen transport and fate modeling in the Upper Salem River watershed using SWAT model. J. Environ. Manag. 2015, 151, 167–177. [Google Scholar] [CrossRef]
- Parajuli, P.B. SWAT Bacteria Sub-Model Evaluation and Application; Kansas State University: Manhattan, NY, USA, 2007. [Google Scholar]
- Whitehead, P.G.; Leckie, H.; Rankinen, K.; Butterfield, D.; Futter, M.; Bussi, G. An INCA model for pathogens in rivers and catchments: Model structure, sensitivity analysis and application to the river thames catchment, UK. Sci. Total Environ. 2016, 572, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Boithias, L.; Cho, K.H.; Silvera, N.; Thammahacksa, C.; Latsachack, K.; Rochelle-Newall, E.; Sengtaheuanghoung, O.; Pierret, A.; Pachepsky, Y.A.; et al. Hydrological modeling of fecal indicator bacteria in a tropical mountain catchment. Water Res. 2017, 119, 102–113. [Google Scholar] [CrossRef]
- Iudicello, J.J.; Chin, D.A. In-stream bacteria modeling as a function of the hydrologic state of a watershed. J. Environ. Eng. 2015, 141, 04014073. [Google Scholar] [CrossRef]
- Cho, K.H.; Pachepsky, Y.A.; Kim, J.H.; Kim, J.-W.; Park, M.-H. The modified SWAT model for predicting fecal coliforms in the Wachusett reservoir watershed, USA. Water Res. 2012, 46, 4750–4760. [Google Scholar] [CrossRef] [PubMed]
- Naganna, S.R.; Deka, P.C.; Ch, S.; Hansen, W.F. Factors influencing streambed hydraulic conductivity and their implications on stream–aquifer interaction: A conceptual review. Environ. Sci. Pollut. Res. 2017, 24, 24765–24789. [Google Scholar] [CrossRef] [PubMed]
- Rode, M.; Hartwig, M.; Wagenschein, D.; Kebede, T.; Borchardt, D. The importance of hyporheic zone processes on ecological functioning and solute transport of streams and rivers. In Ecosystem Services and River Basin Ecohydrology; Springer: Dordrecht, The Netherlands, 2015; pp. 57–82. [Google Scholar]
- Battin, T.J.; Kaplan, L.A.; Newbold, J.D.; Hendricks, S.P. A mixing model analysis of stream solute dynamics and the contribution of a hyporheic zone to ecosystem function. Freshw. Biol. 2003, 48, 995–1014. [Google Scholar] [CrossRef] [Green Version]
- Drummond, J.; Davies-Colley, R.; Stott, R.; Sukias, J.; Nagels, J.; Sharp, A.; Packman, A. Retention and remobilization dynamics of fine particles and microorganisms in pastoral streams. Water Res. 2014, 66, 459–472. [Google Scholar] [CrossRef]
- Wörman, A.; Packman, A.I.; Johansson, H.; Jonsson, K. Effect of flow-induced exchange in hyporheic zones on longitudinal transport of solutes in streams and rivers. Water Resour. Res. 2002, 38, 1001. [Google Scholar] [CrossRef] [Green Version]
- Leight, A.; Hood, R. Precipitation thresholds for fecal bacterial indicators in the Chesapeake Bay. Water Res. 2018, 139, 252–262. [Google Scholar] [CrossRef]
- NH Department of Environmental Services-Shellfish Program. Available online: https://www.des.nh.gov/water/coastal-waters/shellfish (accessed on 3 September 2021).
- Coffey, R.; Paul, M.J.; Stamp, J.; Hamilton, A.; Johnson, T. A review of water quality responses to air temperature and precipitation changes 2: Nutrients, algal blooms, sediment, pathogens. JAWRA J. Am. Water Resour. Assoc. 2019, 55, 844–868. [Google Scholar] [CrossRef]
- Gomez-Velez, J.; Wilson, J.; Cardenas, M.; Harvey, J. Flow and residence times of dynamic river bank storage and sinuosity-driven hyporheic exchange. Water Resour. Res. 2017, 53, 8572–8595. [Google Scholar] [CrossRef]
- Tiwari, A.; Hokajärvi, A.-M.; Santo Domingo, J.; Elk, M.; Jayaprakash, B.; Ryu, H.; Siponen, S.; Vepsäläinen, A.; Kauppinen, A.; Puurunen, O.; et al. Bacterial diversity and predicted enzymatic function in a multipurpose surface water system—From wastewater effluent discharges to drinking water production. Environ. Microbiome 2021, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Hornberger, G.M.; Mills, A.L.; Herman, J.S.; Flewelling, S.A. Nitrate reduction in streambed sediments: Effects of flow and biogeochemical kinetics. Water Resour. Res. 2007, 43, W12413. [Google Scholar] [CrossRef] [Green Version]
- Mancini, J.L. Numerical estimates of coliform mortality rates under various conditions. J. Water Pollut. Control Fed. 1978, 50, 2477–2484. [Google Scholar]
- Briggs, M.A.; Gooseff, M.N.; Peterson, B.J.; Morkeski, K.; Wollheim, W.M.; Hopkinson, C.S. Surface and hyporheic transient storage dynamics throughout a coastal stream network. Water Resour. Res. 2010, 46, W06516. [Google Scholar] [CrossRef] [Green Version]
- Wollheim, W.M.; Vörösmarty, C.J.; Bouwman, A.; Green, P.; Harrison, J.; Linder, E.; Peterson, B.J.; Seitzinger, S.P.; Syvitski, J.P. Global N removal by freshwater aquatic systems using a spatially distributed, within-basin approach. Glob. Biogeochem. Cycles 2008, 22, GB2026. [Google Scholar] [CrossRef] [Green Version]
- Samal, N.R.; Wollheim, W.M.; Zuidema, S.; Stewart, R.J.; Zhou, Z.; Mineau, M.M.; Borsuk, M.E.; Gardner, K.H.; Glidden, S.; Huang, T.; et al. A coupled terrestrial and aquatic biogeophysical model of the upper Merrimack river watershed, New Hampshire, to inform ecosystem services evaluation and management under climate and land-cover change. Ecol. Soc. 2017, 22, 18. [Google Scholar] [CrossRef]
- Wollheim, W.M.; Stewart, R.J.; Aiken, G.R.; Butler, K.D.; Morse, N.B.; Salisbury, J. Removal of terrestrial DOC in aquatic ecosystems of a temperate river network. Geophys. Res. Lett. 2015, 42, 6671–6679. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, L.C.; Hofstra, N.; Kroeze, C.; Medema, G. Advancing waterborne pathogen modelling: Lessons from global nutrient export models. Curr. Opin. Environ. Sustain. 2015, 14, 109–120. [Google Scholar] [CrossRef]
- Mineau, M.M.; Wollheim, W.M.; Stewart, R.J. An index to characterize the spatial distribution of land use within watersheds and implications for river network nutrient removal and export. Geophys. Res. Lett. 2015, 42, 6688–6695. [Google Scholar] [CrossRef] [Green Version]
- Zuidema, S.; Wollheim, W.M.; Mineau, M.M.; Green, M.B.; Stewart, R.J. Controls of chloride loading and impairment at the river network scale in New England. J. Environ. Qual. 2018, 47, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, J.A.; Radoman, N.; Bergquist, J.; Wallin, M.B.; Tranvik, L.J.; Löfgren, S. Regional diversity of complex dissolved organic matter across forested hemiboreal headwater streams. Sci. Rep. 2018, 8, 16060. [Google Scholar] [CrossRef] [Green Version]
- Holcomb, D.A.; Messier, K.P.; Serre, M.L.; Rowny, J.G.; Stewart, J.R. Geostatistical prediction of microbial water quality throughout a stream network using meteorology, land cover, and spatiotemporal autocorrelation. Environ. Sci. Technol. 2018, 52, 7775–7784. [Google Scholar] [CrossRef] [PubMed]
- Carrascal, L.M.; Galván, I.; Gordo, O. Partial least squares regression as an alternative to current regression methods used in ecology. Oikos 2009, 118, 681–690. [Google Scholar] [CrossRef]
- Wang, C.; Schneider, R.L.; Parlange, J.-Y.; Dahlke, H.E.; Walter, M.T. Explaining and modeling the concentration and loading of Escherichia coli in a stream—A case study. Sci. Total Environ. 2018, 635, 1426–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fekete, B.M.; Vörösmarty, C.J.; Grabs, W. High-resolution fields of global runoff combining observed river discharge and simulated water balances. Glob. Biogeochem. Cycles 2002, 16. [Google Scholar] [CrossRef]
- Stewart, R.J.; Wollheim, W.M.; Miara, A.; Vörösmarty, C.J.; Fekete, B.; Lammers, R.B.; Rosenzweig, B. Horizontal cooling towers: Riverine ecosystem services and the fate of thermoelectric heat in the contemporary northeast US. Environ. Res. Lett. 2013, 8, 025010. [Google Scholar] [CrossRef] [Green Version]
- Mulholland, P.J.; DeAngelis, D.L. Chapter 6: Surface-Subsurface Exchange and Nutrient Spiraling. In Streams and Groundwaters; Jones, J.B., Mulholland, P.J., Eds.; Academic: San Diego, CA, USA, 2000. [Google Scholar]
- Leibowitz, S.G.; Wigington, P.J., Jr.; Schofield, K.A.; Alexander, L.C.; Vanderhoof, M.K.; Golden, H.E. Connectivity of streams and wetlands to downstream waters: An integrated systems framework. JAWRA J. Am. Water Resour. Assoc. 2018, 54, 298–322. [Google Scholar] [CrossRef]
- Reder, K.; Flörke, M.; Alcamo, J. Modeling historical fecal coliform loadings to large European rivers and resulting in-stream concentrations. Environ. Model. Softw. 2015, 63, 251–263. [Google Scholar] [CrossRef]
- Blaustein, R.; Pachepsky, Y.; Hill, R.; Shelton, D.; Whelan, G. Escherichia coli survival in waters: Temperature dependence. Water Res. 2013, 47, 569–578. [Google Scholar] [CrossRef]
- New Hampshire Department of Environmental Services, New Hampshire Surface Water Quality Standards. Available online: https://www.des.nh.gov/sites/g/files/ehbemt341/files/documents/2020-01/Env-Wq%201700.pdf (accessed on 15 February 2022).
- Kondo, T.; Sakai, N.; Yazawa, T.; Shimizu, Y. Verifying the applicability of SWAT to simulate fecal contamination for watershed management of Selangor River, Malaysia. Sci. Total Environ. 2021, 774, 145075. [Google Scholar] [CrossRef]
- Sowah, R.A.; Bradshaw, K.; Snyder, B.; Spidle, D.; Molina, M. Evaluation of the soil and water assessment tool (SWAT) for simulating E. coli concentrations at the watershed-scale. Sci. Total Environ. 2020, 746, 140669. [Google Scholar] [CrossRef]
- Cole, J.J.; Prairie, Y.T.; Caraco, N.F.; McDowell, W.H.; Tranvik, L.J.; Striegl, R.G.; Duarte, C.M.; Kortelainen, P.; Downing, J.A.; Middelburg, J.J.; et al. Plumbing the global carbon cycle: Integrating inland waters into the terrestrial carbon budget. Ecosystems 2007, 10, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Wollheim, W.M.; Bernal, S.; Burns, D.A.; Czuba, J.; Driscoll, C.; Hansen, A.; Hensley, R.; Hosen, J.; Inamdar, S.; Kaushal, S.; et al. River network saturation concept: Factors influencing the balance of biogeochemical supply and demand of river networks. Biogeochemistry 2018, 141, 503–521. [Google Scholar] [CrossRef]
- Comer-Warner, S.A.; Gooddy, D.C.; Ullah, S.; Glover, L.; Percival, A.; Kettridge, N.; Krause, S. Seasonal variability of sediment controls of carbon cycling in an agricultural stream. Sci. Total Environ. 2019, 688, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Leight, A.; Hood, R.; Wood, R.; Brohawn, K. Climate relationships to fecal bacterial densities in Maryland shellfish harvest waters. Water Res. 2016, 89, 270–281. [Google Scholar] [CrossRef]
- Camp, E.V.; Pine III, W.E.; Havens, K.; Kane, A.S.; Walters, C.J.; Irani, T.; Lindsey, A.B.; Morris, J.G., Jr. Collapse of a historic oyster fishery: Diagnosing causes and identifying paths toward increased resilience. Ecol. Soc. 2015, 20, 45. [Google Scholar] [CrossRef]
- Fischer, H.; Kloep, F.; Wilzcek, S.; Pusch, M.T. A river’s liver-microbial processes within the hyporheic zone of a large lowland river. Biogeochemistry 2005, 76, 349–371. [Google Scholar] [CrossRef]
- Schaper, J.L.; Posselt, M.; McCallum, J.L.; Banks, E.W.; Hoehne, A.; Meinikmann, K.; Shanafield, M.A.; Batelaan, O.; Lewandowski, J. Hyporheic exchange controls fate of trace organic compounds in an urban stream. Environ. Sci. Technol. 2018, 52, 12285–12294. [Google Scholar] [CrossRef] [Green Version]
- Rehmann, C.R.; Soupir, M.L. Importance of interactions between the water column and the sediment for microbial concentrations in streams. Water Res. 2009, 43, 4579–4589. [Google Scholar] [CrossRef]
- Hrachowitz, M.; Benettin, P.; Van Breukelen, B.M.; Fovet, O.; Howden, N.J.; Ruiz, L.; Van Der Velde, Y.; Wade, A.J. Transit times—The link between hydrology and water quality at the catchment scale. Wiley Interdiscip. Rev. Water 2016, 3, 629–657. [Google Scholar] [CrossRef] [Green Version]
- Wollheim, W.M. From headwaters to rivers to river networks: Scaling in stream ecology. In Stream Ecosystems in a Changing Environment; Jones, J.B., Stanley, E., Eds.; Elsevier: Oxford, UK, 2016; pp. 349–388. [Google Scholar]
- Wollheim, W.M.; Vörösmarty, C.; Peterson, B.J.; Seitzinger, S.P.; Hopkinson, C.S. Relationship between river size and nutrient removal. Geophys. Res. Lett. 2006, 33, L06410. [Google Scholar] [CrossRef] [Green Version]
- Wollheim, W.M.; Mulukutla, G.K.; Cook, C.; Carey, R. Aquatic nitrate retention at river network scales across flow conditions determined using nested in situ sensors. Water Resour. Res. 2017, 53, 9740–9756. [Google Scholar] [CrossRef]
- Hall, R., Jr.; Baker, M.; Rosi-Marshall, E.; Tank, J.; Newbold, J. Solute-specific scaling of inorganic nitrogen and phosphorus uptake in streams. Biogeosciences 2013, 10, 7323–7331. [Google Scholar] [CrossRef] [Green Version]
- Korajkic, A.; Wanjugi, P.; Brooks, L.; Cao, Y.; Harwood, V.J. Persistence and decay of fecal microbiota in aquatic habitats. Microbiol. Mol. Biol. Rev. 2019, 83, e00005-19. [Google Scholar] [CrossRef] [PubMed]
- Petersen, F.; Hubbart, J.A. Physical factors impacting the survival and occurrence of Escherichia coli in secondary habitats. Water 2020, 12, 1796. [Google Scholar] [CrossRef]
- Badgley, B.D.; Steele, M.K.; Cappellin, C.; Burger, J.; Jian, J.; Neher, T.P.; Orentas, M.; Wagner, R. Fecal indicator dynamics at the watershed scale: Variable relationships with land use, season, and water chemistry. Sci. Total Environ. 2019, 697, 134113. [Google Scholar] [CrossRef]
- Hipsey, M.R.; Antenucci, J.P.; Brookes, J.D. A generic, process-based model of microbial pollution in aquatic systems. Water Resour. Res. 2008, 44, W07408. [Google Scholar] [CrossRef]
- Menon, P.; Billen, G.; Servais, P. Mortality rates of autochthonous and fecal bacteria in natural aquatic ecosystems. Water Res. 2003, 37, 4151–4158. [Google Scholar] [CrossRef]
- Johnson, M.F.; Wilby, R.L. Seeing the landscape for the trees: Metrics to guide riparian shade management in river catchments. Water Resour. Res. 2015, 51, 3754–3769. [Google Scholar] [CrossRef] [Green Version]
- Bolick, M.M.; Post, C.J.; Mikhailova, E.A.; Zurqani, H.A.; Grunwald, A.P.; Saldo, E.A. Evaluation of riparian tree cover and shading in the Chauga river watershed using LiDAR and deep learning land cover classification. Remote Sens. 2021, 13, 4172. [Google Scholar] [CrossRef]
- Devane, M.; Moriarty, E.; Weaver, L.; Cookson, A.; Gilpin, B. Fecal indicator bacteria from environmental sources; strategies for identification to improve water quality monitoring. Water Res. 2020, 185, 116204. [Google Scholar] [CrossRef]
- Rothenheber, D.; Jones, S. Enterococcal concentrations in a coastal ecosystem are a function of fecal source input, environmental conditions, and environmental sources. Appl. Environ. Microbiol. 2018, 84, e01038-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paerl, H.W.; Scott, J.T.; McCarthy, M.J.; Newell, S.E.; Gardner, W.S.; Havens, K.E.; Hoffman, D.K.; Wilhelm, S.W.; Wurtsbaugh, W.A. It takes two to tango: When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems. Environ. Sci. Technol. 2016, 50, 10805–10813. [Google Scholar] [PubMed] [Green Version]
- Zhou, T.; Endreny, T. The straightening of a river meander leads to extensive losses in flow complexity and ecosystem services. Water 2020, 12, 1680. [Google Scholar] [CrossRef]
- Durand, P.; Breuer, L.; Johnes, P.J.; Billen, G.; Butturini, A.; Pinay, G.; Van Grinsven, H.; Garnier, J.; Rivett, M.; Reay, D.S.; et al. Nitrogen Processes in Aquatic Ecosystems. In European Nitrogen Assessment; Sutton, M.A., Howard, C.M., Erisman, J.W., Billen, G., Bleeker, A., Grennfelt, P., van Grinsven, H., Grizzetti, B., Eds.; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.; Wollheim, W.M.; Jones, S.H. Removal of Fecal Indicator Bacteria by River Networks. Water 2022, 14, 617. https://doi.org/10.3390/w14040617
Huang T, Wollheim WM, Jones SH. Removal of Fecal Indicator Bacteria by River Networks. Water. 2022; 14(4):617. https://doi.org/10.3390/w14040617
Chicago/Turabian StyleHuang, Tao, Wilfred M. Wollheim, and Stephen H. Jones. 2022. "Removal of Fecal Indicator Bacteria by River Networks" Water 14, no. 4: 617. https://doi.org/10.3390/w14040617
APA StyleHuang, T., Wollheim, W. M., & Jones, S. H. (2022). Removal of Fecal Indicator Bacteria by River Networks. Water, 14(4), 617. https://doi.org/10.3390/w14040617