
The Variation in Groundwater Microbial Communities in an Unconfined Aquifer Contaminated by Multiple Nitrogen Contamination Sources

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
- Determine the effects of the NO3− and NH4+ contamination on the microbial communities in the groundwater;
- Characterize the most abundant phyla in the groundwater and determine their importance;
- Understand how indicators and microbial community analysis can be used in conjunction with physicochemical and environmental data to add insight and resolution to the analysis of contaminant behavior in groundwater.
2. Materials and methods
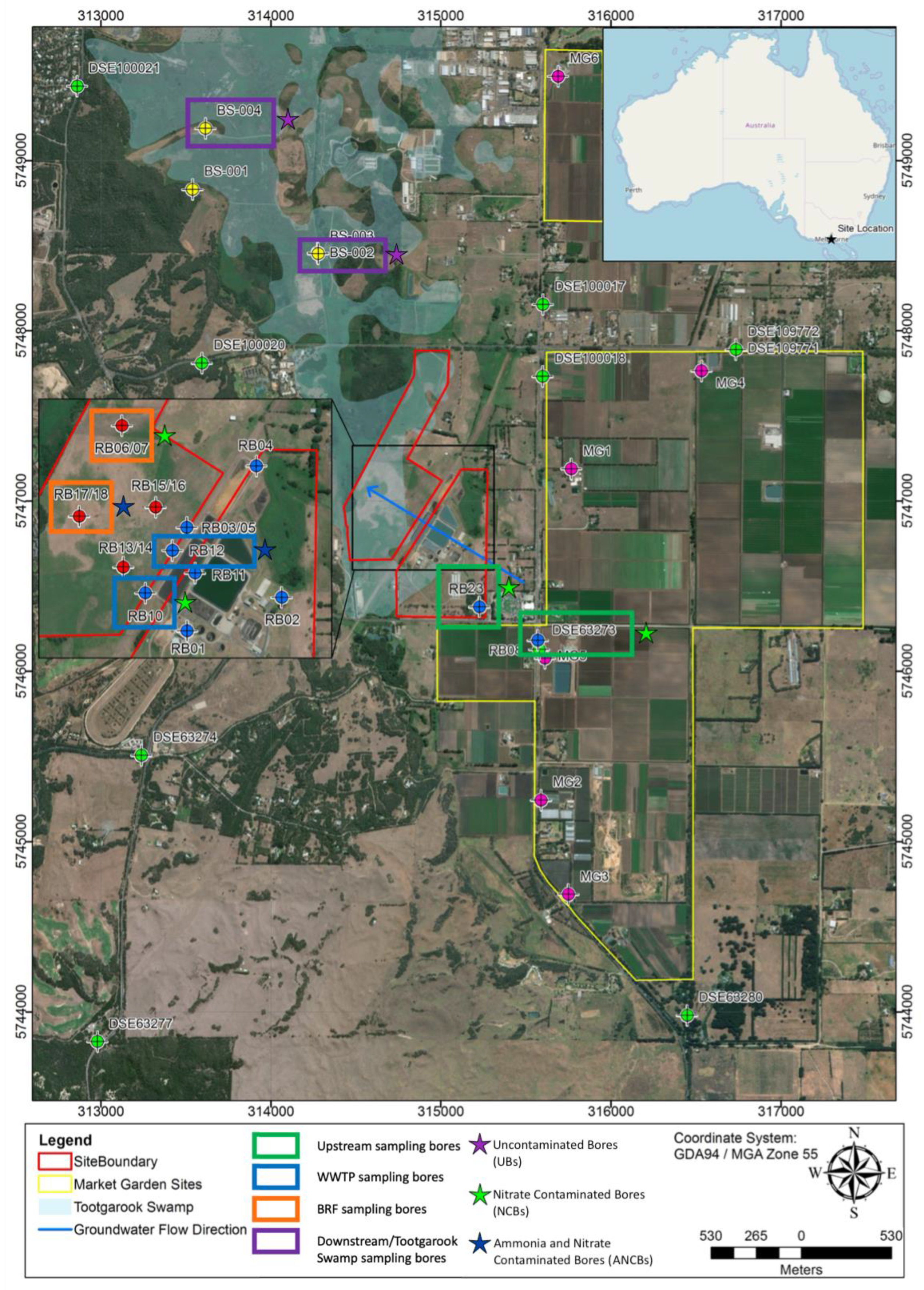
2.1. Study Area
2.2. Sampling Timeline and Locations
2.3. Sampling Techniques, Technology and Guidelines
2.4. Major Ions and Nutrients Analysis
2.5. Microbial Analysis
2.5.1. Sampling
2.5.2. DNA Extraction and Sequencing
2.5.3. File Preparation and Data Analysis
3. Results and Discussion
3.1. Physicochemical Parameters
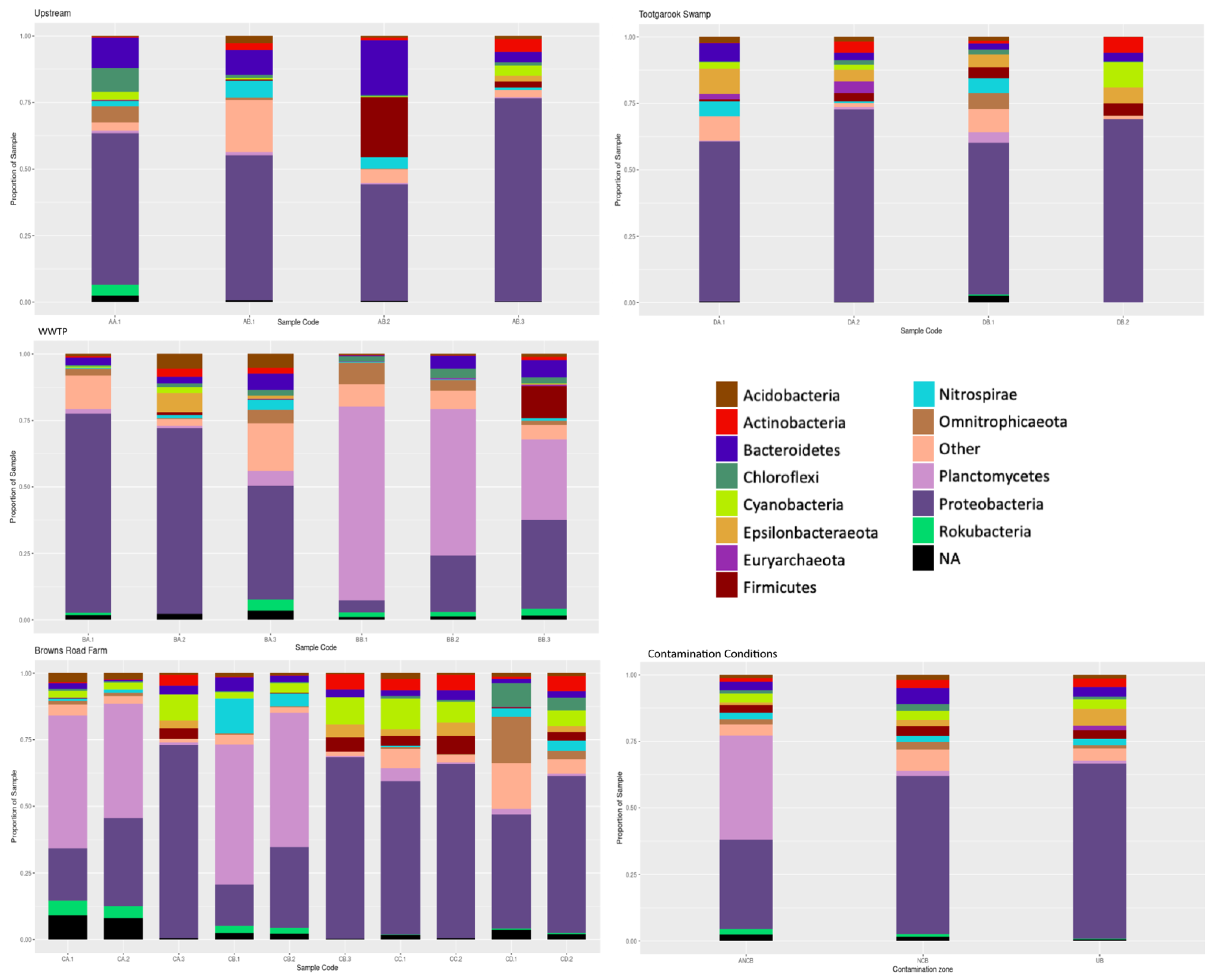
3.2. Microbial Community Structure and Variation
3.2.1. Diversity Metrics
- Bores uncontaminated by nitrogen, BS-002 and BS-004 (henceforth UBs);
- NO3− contaminated bores DSE63273, RB23, RB10 and RB6/7 (henceforth NCBs);
- NH4+ and NO3− contaminated bores RB12 and RB17/18 (henceforth ANCBs).
3.2.2. Spatial Variation within the Microbial Community
3.2.3. Constrained Correspondence Analysis
3.2.4. Microbial Communities and Their Relationship with Physicochemical Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bierkens, M.F.; Wada, Y. Non-Renewable Groundwater Use and Groundwater Depletion: A Review. Environ. Res. Lett. 2019, 14, 063002. [Google Scholar] [CrossRef]
- Morrissy, J.G.; Currell, M.J.; Reichman, S.M.; Surapaneni, A.; Megharaj, M.; Crosbie, N.D.; Hirth, D.; Aquilina, S.; Rajendram, W.; Ball, A.S. Nitrogen contamination and bioremediation in groundwater and the environment: A review. Earth-Sci. Rev. 2021, 20. Available online: https://www.sciencedirect.com/science/article/pii/s0012825221003172 (accessed on 21 December 2021). [CrossRef]
- Griebler, C.; Avramov, M. Groundwater Ecosystem Services: A Review. Freshw. Sci. 2015, 34, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Griebler, C.; Lueders, T. Microbial Biodiversity in Groundwater Ecosystems. Freshw. Biol. 2009, 54, 649–677. [Google Scholar] [CrossRef]
- Colten, C.E. A Historical Perspective on Industrial Wastes and Groundwater Contamination. Geogr. Rev. 1991, 81, 215–228. [Google Scholar] [CrossRef]
- Peña-Haro, S.; Pulido-Velazquez, M.; Sahuquillo, A. A Hydro-Economic Modelling Framework for Optimal Management of Groundwater Nitrate Pollution from Agriculture. J. Hydrol. 2009, 373, 193–203. [Google Scholar] [CrossRef]
- Boulton, A.J. Recent Progress in the Conservation of Groundwaters and Their Dependent Ecosystems. Aquat. Conserv.-Mar. Freshw. Ecosyst. 2009, 19, 731–735. [Google Scholar] [CrossRef]
- Olson, B.M.; Miller, J.J.; Rodvang, S.J.; Yanke, L.J. Soil and Groundwater Quality under a Cattle Feedlot in Southern Alberta. Water Qual. Res. J. 2005, 40, 131–144. [Google Scholar] [CrossRef]
- Kallqvist, T.; Svenson, A. Assessment of Ammonia Toxicity in Tests with the Microalga, Nephroselmis Pyriformis, Chlorophyta. Water Res. 2003, 37, 477–484. [Google Scholar] [CrossRef]
- Caicedo, J.R.; van der Steen, N.P.; Arce, O.; Gijzen, H.J. Effect of Total Ammonia Nitrogen Concentration and Ph on Growth Rates of Duckweed (Spirodela polyrrhiza). Water Res. 2000, 34, 3829–3835. [Google Scholar] [CrossRef]
- Sherris, A.R.; Baiocchi, M.; Fendorf, S.; Luby, S.P.; Yang, W.; Shaw, G.M. Nitrate in Drinking Water During Pregnancy and Spontaneous Preterm Birth: A Retrospective within-Mother Analysis in California. Environ. Health Perspect. 2021, 129, 057001. [Google Scholar] [CrossRef] [PubMed]
- Schullehner, J.; Hansen, B.; Thygesen, M.; Pedersen, C.B.; Sigsgaard, T. Nitrate in Drinking Water and Colorectal Cancer Risk: A Nationwide Population-Based Cohort Study. Int. J. Cancer 2018, 143, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Rabalais, N.N. Nitrogen in Aquatic Ecosystems. Ambio 2002, 31, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Siddig, A.A.; Ellison, A.M.; Ochs, A.; Villar-Leeman, C.; Lau, M.K. How Do Ecologists Select and Use Indicator Species to Monitor Ecological Change? Insights from 14 Years of Publication in Ecological Indicators. Ecol. Indic. 2016, 60, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Adebowale, T.; Surapaneni, A.; Faulkner, D.; McCance, W.; Wang, S.; Currell, M. Delineation of Contaminant Sources and Denitrification Using Isotopes of Nitrate near a Wastewater Treatment Plant in Peri-Urban Settings. Sci. Total Environ. 2019, 651, 2701–2711. [Google Scholar] [CrossRef]
- McCance, W.; Jones, O.; Surapaneni, A.; Currell, M. Characterising Sources of Groundwater Contamination: A Guide to the Use of Novel Groundwater Tracers at Wastewater Treatment Plants. Water E-J. 2020, 5, 1–16. [Google Scholar] [CrossRef]
- McCance, W.; Jones, O.A.; Cendón, D.I.; Edwards, M.; Surapaneni, A.; Chadalavada, S.; Wang, S.; Currell, M. Combining Environmental Isotopes with Contaminants of Emerging Concern (Cecs) to Characterise Wastewater Derived Impacts on Groundwater Quality. Water Res. 2020, 182, 116036. [Google Scholar] [CrossRef]
- Leonard, J.G. Port Phillip Region Groundwater Systems-Future Use and Management; Department of Water Resources: Melbourne, VIC, Australia, 1992.
- Environmnetal Protection Agency (E.P.A.). Goundwater Sampling Guidelines. In Publication 669; State Government of Victoria: Melbourne, VIC, Australia, 2000. [Google Scholar]
- Environmnetal Protection Agency (E.P.A.). Sampling and Analysis of Waters, Wastewaters, Soils and Wastes. In Publication IWRG701; State Government of Victoria: Melbourne, VIC, Australia, 2009. [Google Scholar]
- Koshlaf, E.; Shahsavari, E.; Aburto-Medina, A.; Taha, M.; Haleyur, N.; Makadia, T.H.; Morrison, P.D.; Ball, A.S. Bioremediation Potential of Diesel-Contaminated Libyan Soil. Ecotoxicol. Environ. Saf. 2016, 133, 297–305. [Google Scholar] [CrossRef]
- Rivett, M.O.; Buss, S.R.; Morgan, P.; Smith, J.W.N.; Bemment, C.D. Nitrate Attenuation in Groundwater: A Review of Biogeochemical Controlling Processes. Water Res. 2008, 42, 4215–4232. [Google Scholar] [CrossRef]
- Hageman, R.H. Effect of Form of Nitrogen on Plant Growth. In Nitrification Inhibitors—Potentials and Limitations; 1980; Volume 38, pp. 47–62. Available online: https://acsess.onlinelibrary.wiley.com/doi/abs/10.2134/asaspecpub38.c4 (accessed on 21 December 2021).
- de Caritat, P.; Cooper, M.; Wilford, J. The Ph of Australian Soils: Field Results from a National Survey. Soil Res. 2011, 49, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Hagin, J.; Olsen, S.R.; Shaviv, A. Review of Interaction of Ammonium-Nitrate and Potassium Nutrition of Crops. J. Plant Nutr. 1990, 13, 1211–1226. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A Tribute to Claude Shannon (1916–2001) and a Plea for More Rigorous Use of Species Richness, Species Diversity and the ‘Shannon–Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Strong, W.L. Biased Richness and Evenness Relationships within Shannon–Wiener Index Values. Ecol. Indic. 2016, 67, 703–713. [Google Scholar] [CrossRef]
- Ricotta, C. On Parametric Evenness Measures. J. Theor. Biol. 2003, 222, 189–197. [Google Scholar] [CrossRef]
- Spieck, E.; Bock, E. Nitrifying Bacteria. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–8. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.bm00016 (accessed on 21 December 2021).
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Nitrosomonadaceae Fam. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; p. 1. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.fbm00188 (accessed on 21 December 2021).
- Kitzinger, K.; Koch, H.; Lücker, S.; Sedlacek, C.J.; Herbold, C.; Schwarz, J.; Daebeler, A.; Mueller, A.J.; Lukumbuzya, M.; Romano, S.; et al. Characterization of the First “Candidatus Nitrotoga” Isolate Reveals Metabolic Versatility and Separate Evolution of Widespread Nitrite-Oxidizing Bacteria. MBio 2018, 9, e01186-18. [Google Scholar] [CrossRef] [Green Version]
- Fahrbach, M.; Kuever, J.; Meinke, R.; Kämpfer, P.; Hollender, J. Denitratisoma Oestradiolicum Gen. Nov., Sp. Nov., a 17β-Oestradiol-Degrading, Denitrifying Betaproteobacterium. Int. J. Syst. Evol. Microbiol. 2006, 56, 1547–1552. [Google Scholar] [CrossRef] [Green Version]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Pseudomonadaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; p. 1. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.fbm00232 (accessed on 21 December 2021).
- Brenner, D.J.; Farmer, J.J., III. Enterobacteriaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–24. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.fbm00222 (accessed on 21 December 2021).
- Starke, R.; Müller, M.; Gaspar, M.; Marz, M.; Küsel, K.; Totsche, K.U.; von Bergen, M.; Jehmlich, N. Candidate Brocadiales Dominates C, N and S Cycling in Anoxic Groundwater of a Pristine Limestone-Fracture Aquifer. J. Proteom. 2017, 152, 153–160. [Google Scholar] [CrossRef]
- Smith, R.L.; Bohlke, J.K.; Song, B.; Tobias, C.R. Role of Anaerobic Ammonium Oxidation (Anammox) in Nitrogen Removal from a Freshwater Aquifer. Environ. Sci. Technol. 2015, 49, 12169–12177. [Google Scholar] [CrossRef]
- Waite, D.W.; Vanwonterghem, I.; Rinke, C.; Parks, D.H.; Zhang, Y.; Takai, K.; Sievert, S.M.; Simon, J.; Campbell, B.J.; Hanson, T.E.; et al. Comparative Genomic Analysis of the Class Epsilonproteobacteria and Proposed Reclassification to Epsilonbacteraeota (Phyl. Nov.). Front. Microbiol. 2017, 8, 682. [Google Scholar] [CrossRef]
- Stolz, J.F.; Oremland, R.S.; Paster, B.J.; Dewhirst, F.E.; Vandamme, P. Sulfurospirillum Schumacher, Kroneck, and Pfennig 1993, 188 (Vp) (Effective Publication: Schumacher, Kroneck and Pfennig 1992, 291) Emend. Finster, Liesack and Tindall 1997d, 1216. In Bergey’s Manual of Systematic Bacteriology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 1165–1168. [Google Scholar]
- Vandamme, P.; Dewhirst, F.E.; Paster, B.J.; On, S.L.W. Campylobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–27. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.gbm01071 (accessed on 21 December 2021).
- Goodfellow, M. Actinobacteria Phyl. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–2. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.cbm00004 (accessed on 21 December 2021).
- Miao, V.; Davies, J. Actinobacteria: The Good, the Bad, and the Ugly. Antonie Van Leeuwenhoek 2010, 98, 143–150. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Schumann, P. Cellulomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–14. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.gbm00063 (accessed on 21 December 2021).
- Stackebrandt, E.; Schumann, P. Oerskovia. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–9. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.gbm00064 (accessed on 21 December 2021).
- Gupta, R. Fundamentals of Bacterial Physiology and Metabolism; Springer Nature: University of Delhi New Delhi, Delhi, India, 2021. [Google Scholar]
- Kizina, J. Insights into the Biology of Candidate Division Op3 Lim Populations; Universität Bremen: Bremen, Germany, 2017. [Google Scholar]
- Glöckner, J.; Kube, M.; Shrestha, P.M.; Weber, M.; Glöckner, F.O.; Reinhardt, R.; Liesack, W. Phylogenetic Diversity and Metagenomics of Candidate Division Op3. Environ. Microbiol. 2010, 12, 1218–1229. [Google Scholar] [CrossRef]
- Zhou, L.; Dong, X. Methanospirillum. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–5. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.gbm00511.pub2 (accessed on 21 December 2021).
- Logan, N.A.; De Vos, P. Bacillaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; p. 1. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.fbm00112 (accessed on 21 December 2021).
- Schleifer, K.-H.; Bell, J.A. Staphylococcus. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–43. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.gbm00569 (accessed on 21 December 2021).
- Whiley, R.A.; Hardie, J.M. Streptococcus. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; pp. 1–86. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.gbm00612 (accessed on 21 December 2021).
- Thrash, J.C.; Coates, J.D. Phylum Xvii. Acidobacteria Phyl. Nov. In Bergey’s Manual® of Systematic Bacteriology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 725–735. [Google Scholar]
- Thrash, J.C.; Coates, J.D. Acidobacteriaceae Fam. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; p. 1. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.fbm00001 (accessed on 21 December 2021).
- Thrash, J.C.; Coates, J.D. Holophagae Fam. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2015; p. 1. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.cbm00002 (accessed on 21 December 2021).
- Moe, W.M.; Rainey, F.A. Dehalogenimonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2020; pp. 1–8. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.gbm01556 (accessed on 21 December 2021).
- Yamada, T.; Sekiguchi, Y. Anaerolineaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; 2018; pp. 1–5. Available online: https://onlinelibrary.wiley.com/doi/10.1002/9781118960608.fbm00301 (accessed on 21 December 2021).
- Becraft, E.D.; Woyke, T.; Jarett, J.; Ivanova, N.; Godoy-Vitorino, F.; Poulton, N.; Brown, J.M.; Brown, J.; Lau, M.C.Y.; Onstott, T.; et al. Rokubacteria: Genomic Giants among the Uncultured Bacterial Phyla. Front. Microbiol. 2017, 8, 2264. [Google Scholar] [CrossRef] [PubMed]
- McCance, W.; Jones, O.A.H.; Cendón, D.I.; Edwards, M.; Surapaneni, A.; Chadalavada, S.; Currell, M. Decoupling Wastewater Impacts from Hydrogeochemical Trends in Impacted Groundwater Resources. Sci. Total Environ. 2021, 774, 145781. [Google Scholar] [CrossRef]
- Australian Bureau Of Meteorology. Climate Data Online. Available online: http://www.bom.gov.au/climate/data/ (accessed on 1 November 2021).
- De Silva, S.; Ball, A.S.; Shahsavari, E.; Indrapala, D.V.; Reichman, S.M. The Effects of Vehicular Emissions on the Activity and Diversity of the Roadside Soil Microbial Community. Environ. Pollut. 2021, 277, 116744. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Location | Sample | Bore Code | Date Sampled | Bore Depth | DO | EC (uS/cm) | pH | Redox Potential (mV) | Water Temp (°C) | E. coli (cfu/100 mL) | Alkalinity as CaCO3 | HCO3− | SO4 | Anionic Strength (meq/L) | Cationic Strength (meq/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Upstream | AA.1 | DSE63273 | Aug-2018 | 25.08 | 4.46 | 2944 | 6.7 | 106.9 | 16.3 | 0 | 315 | 385 | 410 | 32 | 28 |
| AB.1 | RB23 | Aug-2018 | 11.14 | 5.92 | 1487 | 7.15 | 28.9 | 15.1 | 0 | 270 | 329.4 | 24 | 17 | 14 | |
| AB.2 | RB23 | Nov-2018 | 11.18 | 5.89 | 1463 | 6.93 | 111.3 | 15.3 | 0 | 170 | 210 | 25 | 14 | 14 | |
| AB.3 | RB23 | May-2019 | 11.12 | 4.77 | 1434 | 7.27 | 33.8 | 17.0 | 0 | 170 | 210 | 45 | 13 | 12 | |
| Ca2+ | Na+ | K+ | Cl− | Mg2+ | NH4+ as N | NO3− as N | Total N | TOC | PO43− | TDS | Cu | Fe2+ | Zn | ||
| AA.1 | 390 | 135 | 1.2 | 230 | 41 | 0.15 | 150 | 155 | 3.7 | 0.009 | 2250 | 0.004 | 0.02 | 0.0295 | |
| AB.1 | 170 | 84 | 1 | 170 | 19 | <0.1 | 83 | 83 | 1.3 | <0.05 | 1200 | 0.003 | <0.01 | 0.006 | |
| AB.2 | 170 | 85 | <1 | 160 | 17 | <0.1 | 72 | 88 | 1.3 | <0.05 | 960 | 0.008 | <0.01 | 0.014 | |
| AB.3 | 150 | 75 | <1 | 150 | 15 | <0.1 | 64 | 65 | 2.5 | <0.005 | 1000 | 0.0092 | 0.003 | 0.0087 | |
| Wastewater Treatment Plant (WWTP) | BA.1 | RB10 | Aug-2018 | 5.10 | 0.22 | 2357 | 6.68 | 32.8 | 14.8 | 2 | 610 | 744.2 | 640 | 32 | 27 |
| BA.2 | RB10 | Nov-2018 | 5.46 | 0.51 | 2444 | 6.73 | 61.2 | 18 | 0 | 430 | 530 | 580 | 29 | 31 | |
| BA.3 | RB10 | May-2019 | 5.39 | 0.48 | 2659 | 6.72 | 21.1 | 20 | 0 | 470 | 580 | 760 | 34 | 34 | |
| BB.1 | RB12 | Aug-2018 | 5.10 | 0.15 | 2192 | 6.78 | 32.2 | 15.3 | 2 | 560 | 683.2 | 330 | 26 | 24 | |
| BB.2 | RB12 | Nov-2018 | 5.20 | 0.21 | 2641 | 6.69 | 68.7 | 18.4 | 0 | 500 | 610 | 380 | 28 | 31 | |
| BB.3 | RB12 | May-2019 | 5.14 | 0.47 | 2466 | 6.81 | 23.3 | 19.8 | 0 | 450 | 550 | 530 | 29 | 27 | |
| Sludge Lagoon | - | - | 3.59 | 1171 | 7.51 | 44.6 | 18.6 | 4426 | 460 | 396 | 24 | - | - | ||
| Ca2+ | Na+ | K+ | Cl− | Mg2+ | NH4+ as N | NO3− as N | Total N | TOC | PO43− | TDS | Cu | Fe2+ | Zn | ||
| BA.1 | 350 | 120 | 11 | 210 | 50 | <0.1 | 14 | 15 | 10 | <0.05 | 1800 | 0.002 | <0.01 | 0.01 | |
| BA.2 | 430 | 110 | 11 | 170 | 59 | <0.5 | 54 | 55 | 12 | <0.05 | 1700 | 0.001 | <0.01 | 0.013 | |
| BA.3 | 430 | 140 | 14 | 200 | 69 | <0.5 | 39 | 40 | 13 | <0.005 | 1900 | 0.0025 | 0.015 | 0.012 | |
| BB.1 | 230 | 120 | 40 | 190 | 48 | 26 | 40 | 66 | 10 | <0.5 | 1400 | 0.004 | <0.01 | 0.009 | |
| BB.2 | 320 | 160 | 40 | 210 | 61 | 13 | 63 | 76 | 12 | 0.05 | 1600 | 0.002 | <0.01 | 0.015 | |
| BB.3 | 310 | 140 | 31 | 170 | 54 | 5.5 | 54 | 60 | 11 | <0.005 | 1600 | 0.0034 | 0.009 | 0.0086 | |
| Sludge Lagoon | 37 | 82 | 44 | 107 | 20 | 54 | 0.17 | 57 | 27 | 70 | 493 | 0.11 | 3.5 | 0.25 | |
| Browns Road Farm | CA.1 | RB17 | Aug-2018 | 4.74 | 0.24 | 1486 | 6.92 | 30.5 | 14.4 | 0 | 350 | 430 | 220 | 17 | 18 |
| CA.2 | RB17 | Nov-2018 | 4.74 | 0.2 | 1723 | 6.84 | 77.2 | 15.1 | 0 | 370 | 450 | 330 | 20 | 18 | |
| CA.3 | RB17 | May20-19 | 4.68 | 0.64 | 1682 | 7.01 | 29.1 | 17.1 | 1 | 360 | 440 | 250 | 18 | 18 | |
| CB.1 | RB18 | Aug-2018 | 10.80 | 0.17 | 1381 | 6.90 | 20.7 | 15.4 | 0 | 350 | 430 | 180 | 16 | 15 | |
| CB.2 | RB18 | Nov-2018 | 10.80 | 0.19 | 1309 | 6.87 | 79.9 | 15.5 | 0 | 360 | 440 | 130 | 14 | 13 | |
| CB.3 | RB18 | May-2019 | 10.77 | 0.56 | 1480 | 7.24 | 29.0 | 16.0 | 0 | 370 | 450 | 190 | 16 | 13 | |
| CC.1 | RB06 | Aug-2018 | 9.15 | 0.12 | 2633 | 6.79 | 33.4 | 14.7 | 0 | 330 | 400 | 1100 | 37 | 37 | |
| CC.2 | RB06 | May-2019 | 9.01 | 0.61 | 1915 | 7.19 | 35.9 | 15.3 | 0 | 270 | 330 | 490 | 22 | 18 | |
| CD.1 | RB07 | Aug-2018 | 4.65 | 0.22 | 3342 | 6.78 | 24.0 | 13.6 | 0 | 410 | 490 | 1500 | 48 | 36 | |
| CD.2 | RB07 | May-2019 | 4.64 | 0.76 | 3503 | 6.73 | −8.4 | 15.8 | 0 | 420 | 510 | 1500 | 48 | 36 | |
| Ca2+ | Na+ | K+ | Cl− | Mg2+ | NH4+ as N | NO3− as N | Total N | TOC | PO43− | TDS | Cu | Fe2+ | Zn | ||
| CA.1 | 200 | 120 | <1 | 150 | 28 | <0.1 | 12 | 13 | 2.4 | <0.05 | 1000 | 0.003 | <0.01 | 0.011 | |
| CA.2 | 200 | 120 | <1 | 160 | 32 | <0.1 | 15 | 16 | 7.1 | <0.05 | 1200 | <0.001 | <0.01 | 0.006 | |
| CA.3 | 200 | 130 | 0.8 | 160 | 27 | <0.1 | 14 | 16 | 1.6 | <0.005 | 1000 | 0.001 | 0.003 | 0.0043 | |
| CB.1 | 170 | 100 | 3 | 140 | 25 | 4.1 | 18 | 20 | 2.3 | <0.05 | 840 | 0.003 | <0.01 | 0.007 | |
| CB.2 | 140 | 100 | 3 | 110 | 22 | 3.3 | 15 | 18 | 6.1 | <0.05 | 830 | <0.001 | <0.01 | 0.004 | |
| CB.3 | 130 | 110 | 3.5 | 140 | 22 | 4.6 | 11 | 20 | 1.3 | 0.006 | 790 | 0.0012 | 0.003 | 0.0052 | |
| CC.1 | 480 | 170 | <1 | 250 | 63 | <0.1 | 7.5 | 8.2 | 8.1 | <0.05 | 2100 | 0.002 | <0.01 | 0.032 | |
| CC.2 | 210 | 110 | 0.8 | 180 | 34 | <0.1 | 18 | 20 | 4.4 | <0.005 | 1200 | <0.001 | <0.01 | 0.005 | |
| CD.1 | 570 | 210 | 7 | 320 | 81 | 0.1 | <0.01 | 0.9 | 16.0 | 0.08 | 2800 | 0.002 | 0.9 | 0.01 | |
| CD.2 | 470 | 160 | 1 | 300 | 70 | <0.1 | 0.22 | 1.2 | 13.0 | <0.005 | 2700 | 0.001 | 1.2 | 0.01 | |
| Downstream/Tootgarook Swamp | DA.1 | BS-002 | Nov-2018 | 26.35 | 0.24 | 940 | 7.43 | 5.5 | 15.5 | 0 | 240 | 300 | 8 | 9 | 9 |
| DA.2 | BS-002 | May-2019 | 26.35 | 0.74 | 1004 | 7.37 | −59.6 | 15.3 | 0 | 250 | 300 | 7 | 10 | 8 | |
| DB.1 | BS-004 | Nov-2018 | 5.49 | 0.17 | 718 | 7.44 | −106.9 | 15.1 | 0 | 210 | 250 | 26 | 7 | 7 | |
| DB.2 | BS-004 | May-2019 | 5.49 | 0.69 | 776 | 7.77 | −83.5 | 14.3 | <10 | 210 | 260 | 34 | 8 | 7 | |
| Ca2+ | Na+ | K+ | Cl− | Mg2+ | NH4+ as N | NO3− as N | Total N | TOC | PO43− | TDS | Cu | Fe2+ | Zn | ||
| DA.1 | 84 | 82 | 1 | 160 | 11 | 0.1 | <0.01 | 0.2 | 2.6 | <0.05 | 440 | <0.001 | 0.13 | 0.008 | |
| DA.2 | 83 | 78 | 1.3 | 170 | 11 | <0.1 | 0.01 | 0.07 | <0.5 | <0.005 | 480 | 0.0003 | 0.096 | 0.0026 | |
| DB.1 | 47 | 52 | 4 | 93 | 25 | 0.2 | <0.01 | 0.3 | 3.1 | <0.05 | 300 | <0.001 | 1.4 | 0.003 | |
| DB.2 | 51 | 49 | 4 | 100 | 28 | <2 | <0.2 | 0.66 | 2.7 | 0.05 | 360 | <0.0001 | 0.57 | 0.0007 |
| Sample Location | Sample | Bore Code | Ammonia as N (mg L−1) | Number of Replicates | Non-Chimeric Sequences | OTUs | ACE | Chao1 | Gini-Simpson | Shannon-Wiener | Pielou’s Evenness |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Upstream | AA.1 | DSE63277 | 0.15 | 3 | 6403 | 1289 | 1429 | 1524 | 0.01 | 5.41 | 0.76 |
| AB.1 | RB23 | <0.1 | 3 | 12,150 | 1153 | 1227 | 1261 | 0.01 | 5.72 | 0.81 | |
| AB.2 | RB23 | <0.1 | 3 | 26,674 | 1189 | 1251 | 1260 | 0.03 | 5.11 | 0.72 | |
| AB.3 | RB23 | <0.1 | 3 | 22,156 | 1263 | 1309 | 1404 | 0.04 | 4.60 | 0.64 | |
| Average | 0.1 | 3 | 16,846 | 1224 | 1304 | 1362 | 0.02 | 5.21 | 0.73 | ||
| WWTP | BA.1 | RB10 | <0.1 | 3 | 12,535 | 1680 | 1880 | 3333 | 0.02 | 5.62 | 0.76 |
| BA.2 | RB10 | <0.5 | 3 | 1584 | 263 | 291 | 779 | 0.11 | 3.56 | 0.64 | |
| BA.3 | RB10 | <0.5 | 3 | 21,782 | 2590 | 2717 | 2742 | 0.01 | 6.58 | 0.84 | |
| Average | <0.5 | 3 | 11,967 | 1511 | 1629 | 2285 | 0.04 | 5.25 | 0.74 | ||
| BB.1 | RB12 | 26 | 3 | 7353 | 1705 | 1907 | 2069 | 0.13 | 3.58 | 0.48 | |
| BB.2 | RB12 | 13 | 3 | 23,760 | 2391 | 2551 | 2669 | 0.11 | 3.84 | 0.49 | |
| BB.3 | RB12 | 5.5 | 3 | 18,086 | 2170 | 2356 | 5870 | 0.04 | 5.35 | 0.70 | |
| Average | 14.8 | 3 | 16,400 | 2089 | 2271 | 3536 | 0.09 | 4.26 | 0.56 | ||
| Browns Road Farm | CA.1 | RB17 | <0.1 | 2 | 21,920 | 1643 | 1790 | 1772 | 0.10 | 4.20 | 0.57 |
| CA.2 | RB17 | <0.1 | 2 | 17,700 | 1599 | 1758 | 1748 | 0.09 | 3.98 | 0.54 | |
| CA.3 | RB17 | <0.1 | 3 | 1542 | 415 | 432 | 574 | 0.02 | 4.75 | 0.79 | |
| Average | <0.1 | 2 | 13,721 | 1219 | 1327 | 1365 | 0.07 | 4.31 | 0.63 | ||
| CB.1 | RB18 | 4.1 | 3 | 30,838 | 1591 | 1703 | 1916 | 0.10 | 3.97 | 0.54 | |
| CB.2 | RB18 | 3.3 | 3 | 17,382 | 1374 | 1459 | 1652 | 0.10 | 3.92 | 0.54 | |
| CB.3 | RB18 | 4.6 | 3 | 1691 | 398 | 409 | 593 | 0.02 | 4.91 | 0.82 | |
| Average | 4.0 | 3 | 16,637 | 1121 | 1191 | 1387 | 0.07 | 4.26 | 0.63 | ||
| CC.1 | RB06 | <0.1 | 3 | 1964 | 758 | 837 | 1664 | 0.01 | 5.55 | 0.84 | |
| CC.2 | RB06 | <0.1 | 3 | 2284 | 556 | 578 | 858 | 0.01 | 5.19 | 0.82 | |
| CD.1 | RB07 | 0.1 | 3 | 15,628 | 2906 | 3099 | 3345 | 0.01 | 6.45 | 0.81 | |
| CD.2 | RB07 | <0.1 | 3 | 36,019 | 2894 | 3075 | 6421 | 0.01 | 6.34 | 0.80 | |
| Average | 0.1 | 3 | 13,974 | 1779 | 1897 | 3072 | 0.01 | 5.88 | 0.82 | ||
| Tootgarook Swamp | DA.1 | BS-002 | 0.1 | 3 | 9588 | 940 | 994 | 1039 | 0.03 | 4.92 | 0.72 |
| DA.2 | BS-002 | <0.1 | 3 | 33,592 | 1534 | 1589 | 1614 | 0.05 | 4.65 | 0.63 | |
| DB.1 | BS-004 | 0.2 | 3 | 13,032 | 1882 | 2066 | 2090 | 0.01 | 6.03 | 0.80 | |
| DB.2 | BS-004 | <2 | 3 | 1809 | 379 | 389 | 610 | 0.02 | 4.87 | 0.82 | |
| Average | 0.2 | 3 | 14,505 | 1184 | 1260 | 1338 | 0.03 | 5.12 | 0.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morrissy, J.G.; Currell, M.J.; Reichman, S.M.; Surapaneni, A.; Megharaj, M.; Crosbie, N.D.; Hirth, D.; Aquilina, S.; Rajendram, W.; Ball, A.S. The Variation in Groundwater Microbial Communities in an Unconfined Aquifer Contaminated by Multiple Nitrogen Contamination Sources. Water 2022, 14, 613. https://doi.org/10.3390/w14040613
Morrissy JG, Currell MJ, Reichman SM, Surapaneni A, Megharaj M, Crosbie ND, Hirth D, Aquilina S, Rajendram W, Ball AS. The Variation in Groundwater Microbial Communities in an Unconfined Aquifer Contaminated by Multiple Nitrogen Contamination Sources. Water. 2022; 14(4):613. https://doi.org/10.3390/w14040613
Chicago/Turabian StyleMorrissy, Justin G., Matthew J. Currell, Suzie M. Reichman, Aravind Surapaneni, Mallavarapu Megharaj, Nicholas D. Crosbie, Daniel Hirth, Simon Aquilina, William Rajendram, and Andrew S. Ball. 2022. "The Variation in Groundwater Microbial Communities in an Unconfined Aquifer Contaminated by Multiple Nitrogen Contamination Sources" Water 14, no. 4: 613. https://doi.org/10.3390/w14040613
APA StyleMorrissy, J. G., Currell, M. J., Reichman, S. M., Surapaneni, A., Megharaj, M., Crosbie, N. D., Hirth, D., Aquilina, S., Rajendram, W., & Ball, A. S. (2022). The Variation in Groundwater Microbial Communities in an Unconfined Aquifer Contaminated by Multiple Nitrogen Contamination Sources. Water, 14(4), 613. https://doi.org/10.3390/w14040613