
Effect of Different Land Use Types on the Taxonomic and Functional Diversity of Macroinvertebrates in an Urban Area of Northern China


Abstract
:1. Introduction
2. Materials and Methods
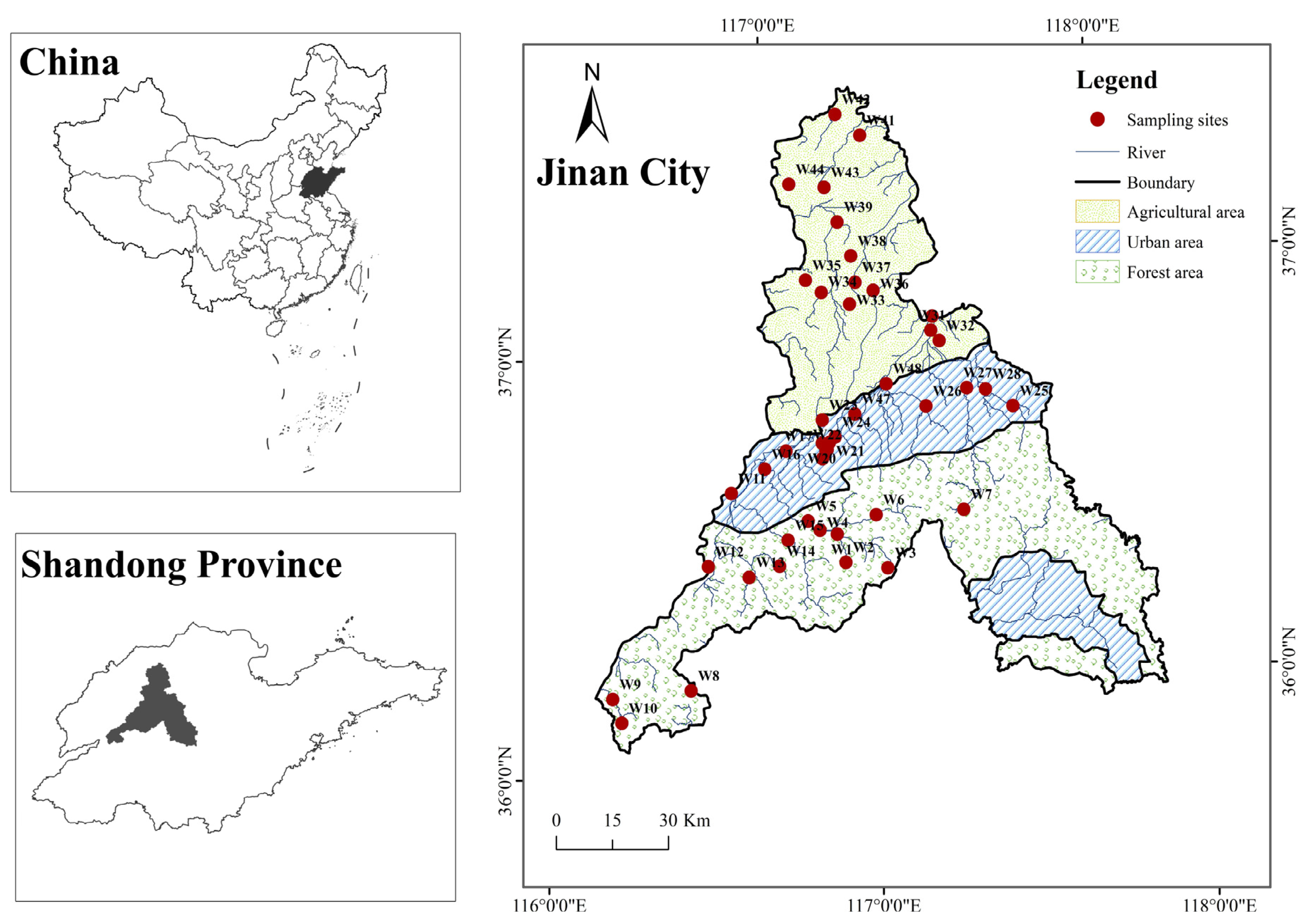
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
3.1. Environmental Variables
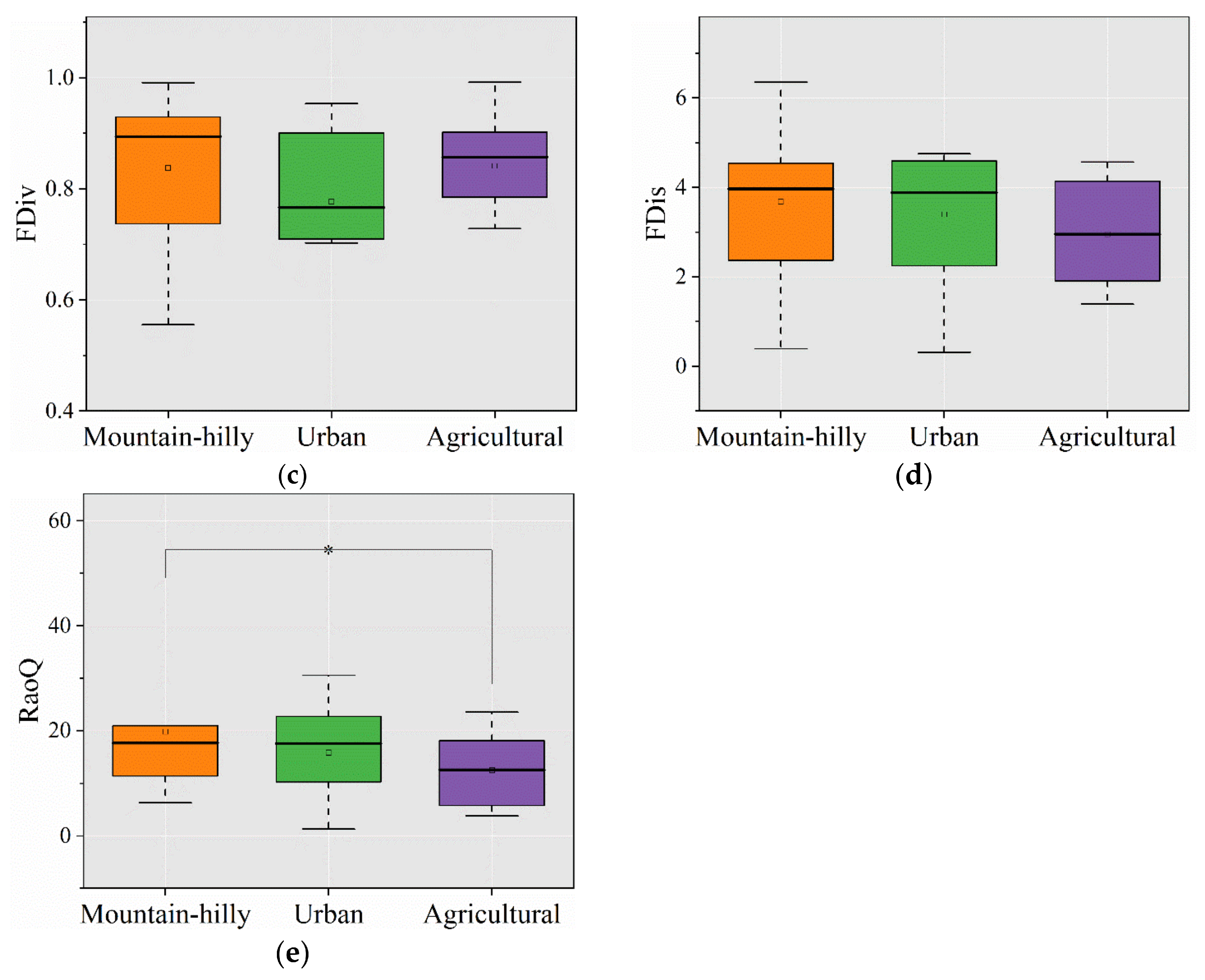
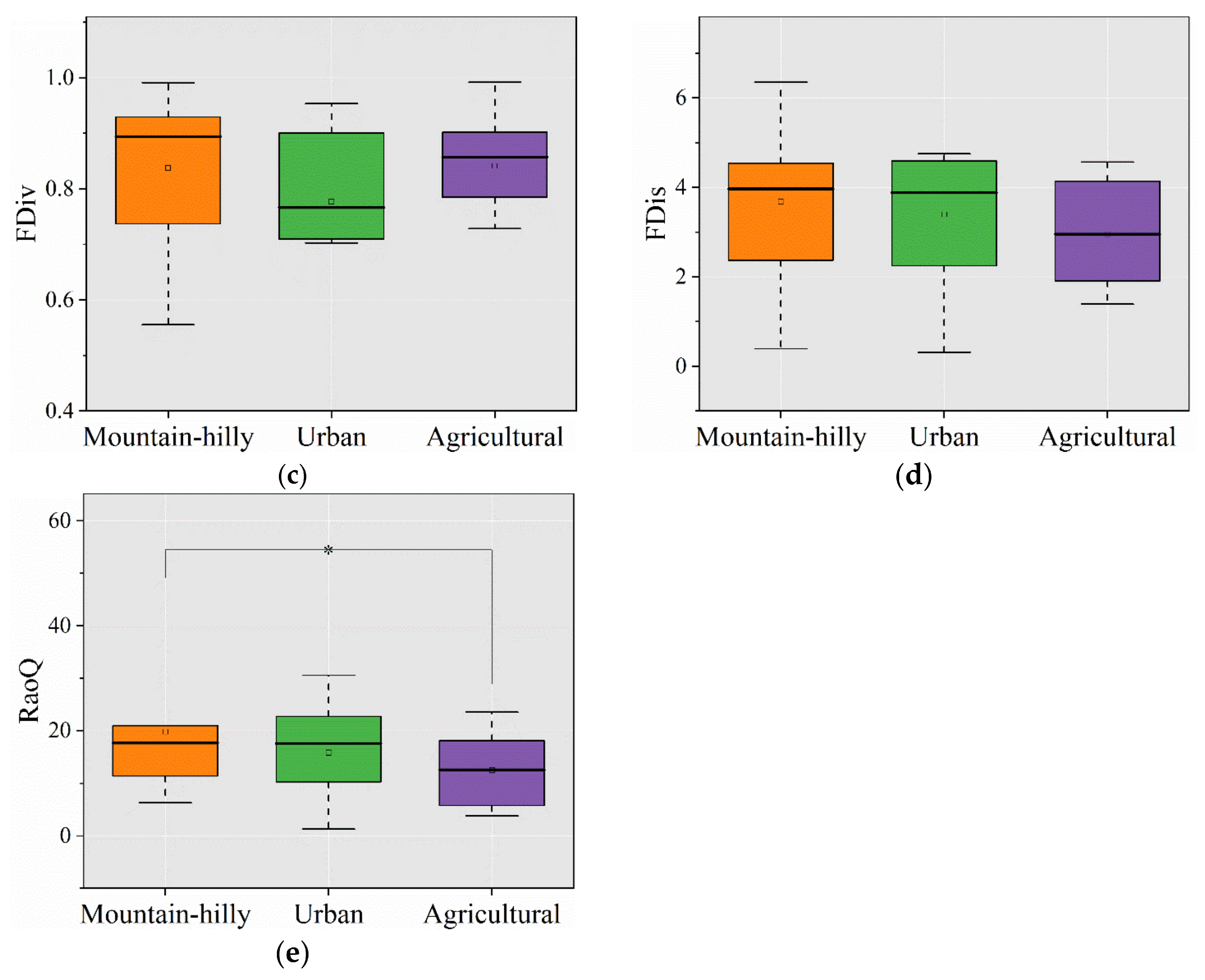
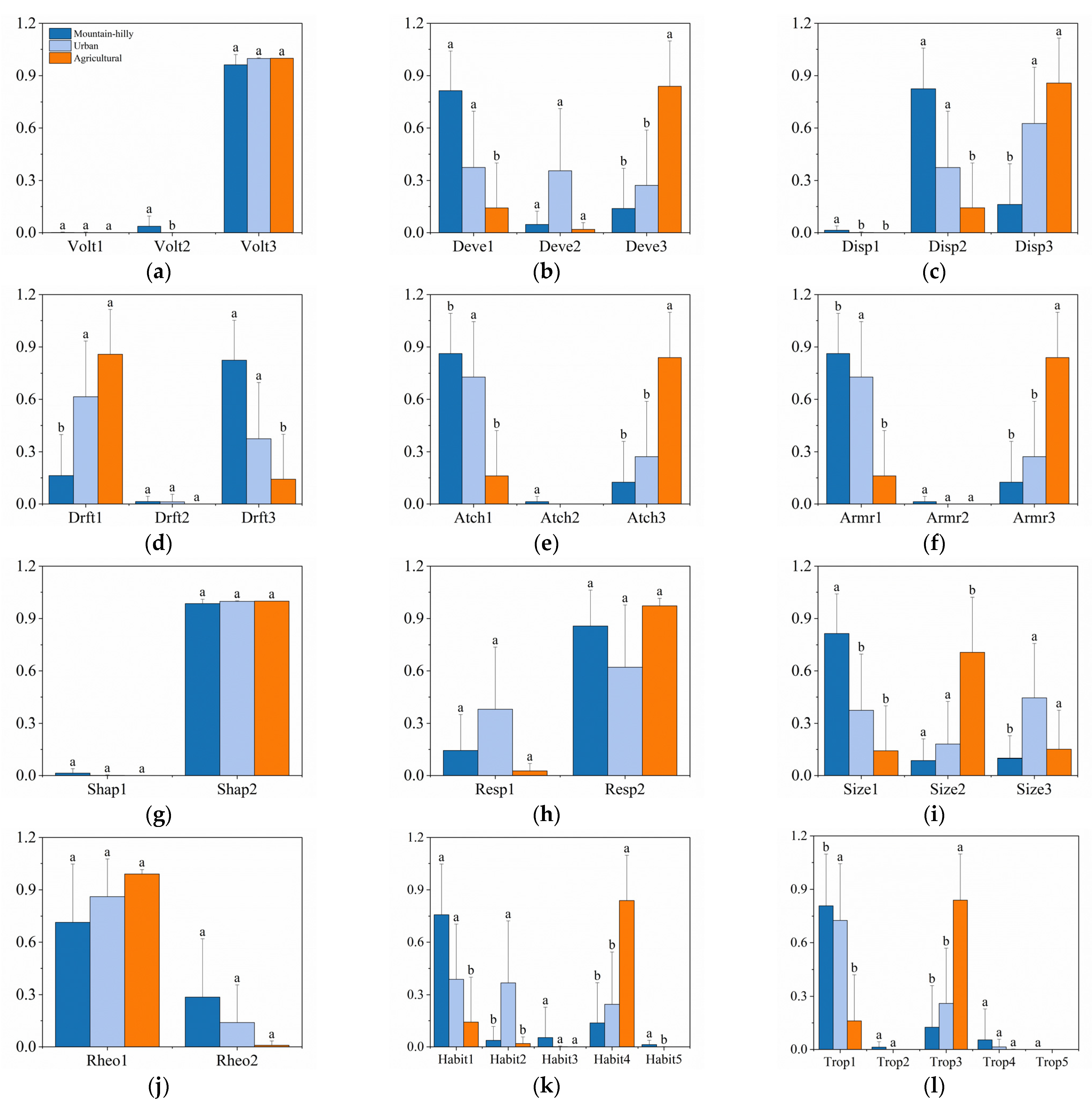
3.2. Taxonomic and Functional Diversity
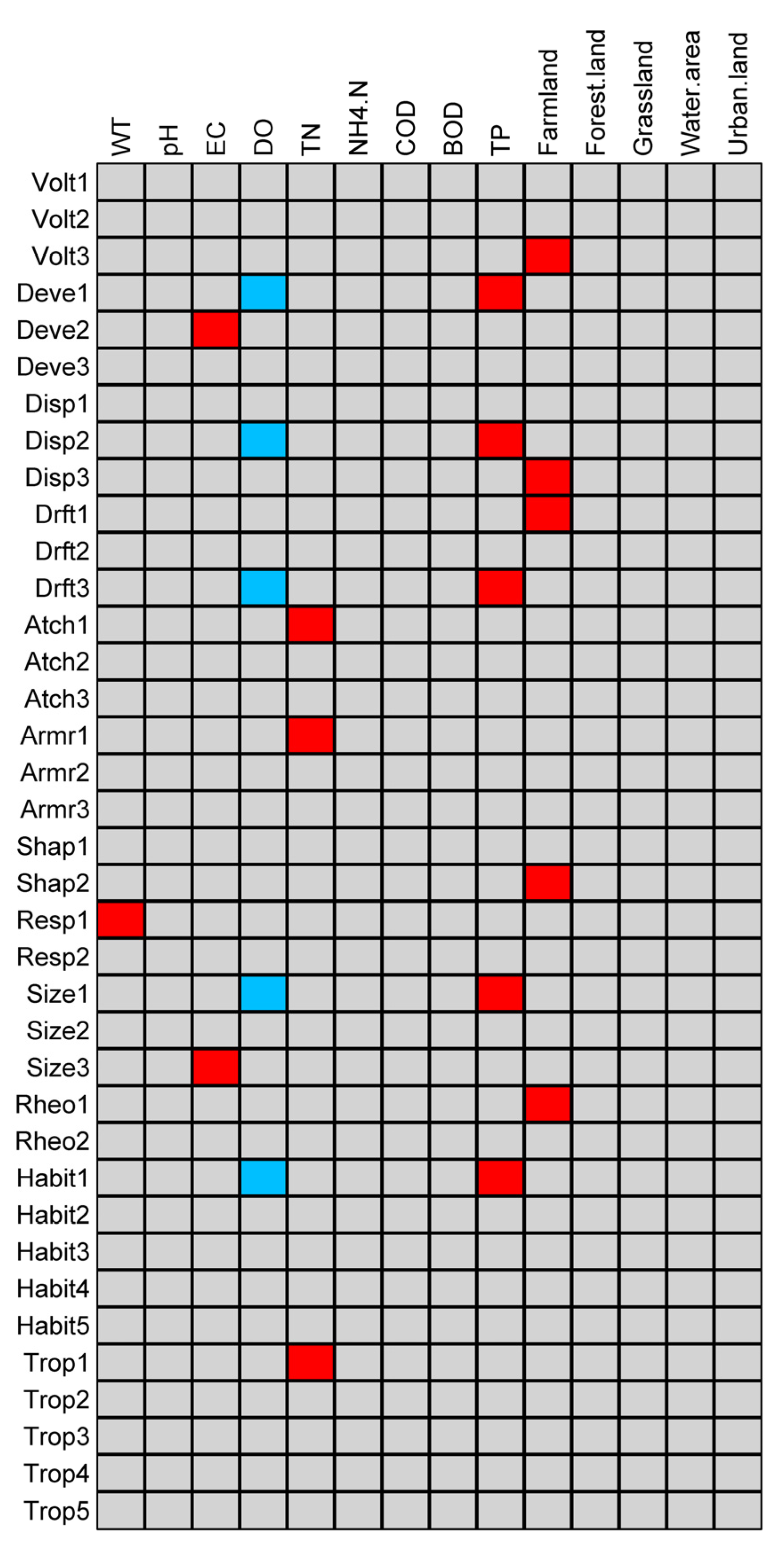
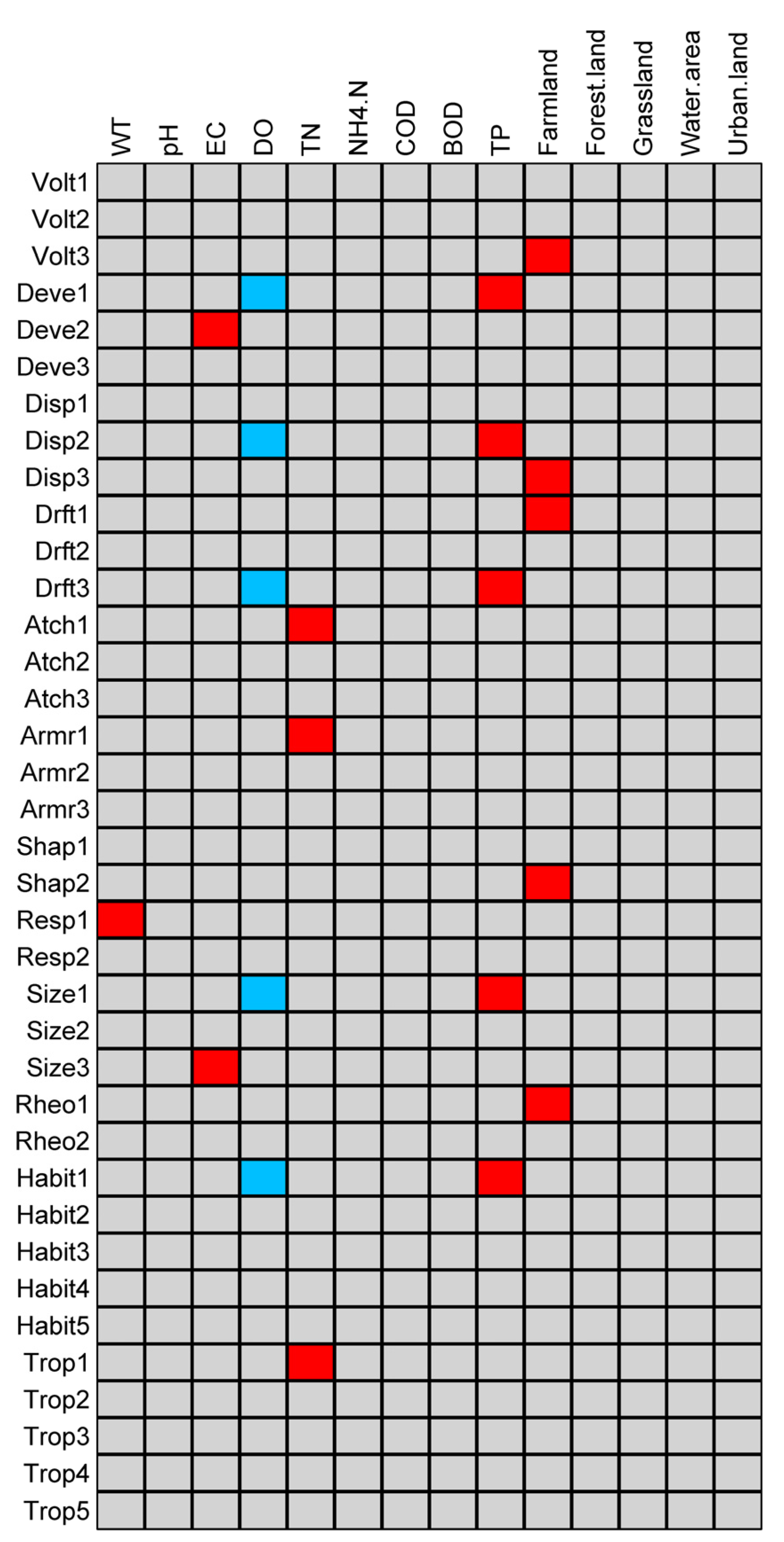
3.3. Relationship between Functional Diversity and Environmental Variables
4. Discussion
4.1. Response of Taxonomic Diversity to Different Types of Land Use
4.2. Response of Functional Diversity to Different Types of Land Use
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, S.; Techato, K.; Monprapussorn, S.; Yuangyai, C. Integrating Land use and Water quality for Environmental based land use planning for U-tapao River Basin, Thailand. In Proceedings of the PSU-USM International Conference on Humanities and Social Sciences, Hat Yai, Thailand, 3–4 October 2011; pp. 556–563. [Google Scholar]
- Barnum, T.R.; Weller, D.E.; Williams, M. Urbanization reduces and homogenizes trait diversity in stream macroinvertebrate communities. Ecol. Appl. 2017, 27, 2428–2442. [Google Scholar] [CrossRef] [PubMed]
- Rueda, J.; Camacho, A.; Mezquita, F.; Hernandez, R.; Roca, J.R. Effect of episodic and regular sewage discharges on the water chemistry and macroinvertebrate fauna of a Mediterranean stream. Water Air Soil Pollut. 2002, 140, 425–444. [Google Scholar] [CrossRef]
- Covich, A.P.; Palmer, M.A.; Crowl, T.A. The role of benthic invertebrate species in freshwater ecosystems—Zoobenthic species influence energy flows and nutrient cycling. Bioscience 1999, 49, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Piano, E.; Doretto, A.; Mammola, S.; Falasco, E.; Fenoglio, S.; Bona, F. Taxonomic and functional homogenisation of macroinvertebrate communities in recently intermittent Alpine watercourses. Freshw. Biol. 2020, 65, 2096–2107. [Google Scholar] [CrossRef]
- Fierro, P.; Bertrán, C.; Tapia, J.; Hauenstein, E.; Peña-Cortés, F.; Vergara, C.; Cerna, C.; Vargas-Chacoff, L. Effects of local land-use on riparian vegetation, water quality, and the functional organization of macroinvertebrate assemblages. Sci. Total Environ. 2017, 609, 724–734. [Google Scholar] [CrossRef]
- Townsend, C.R.; Uhlmann, S.S.; Matthaei, C.D. Individual and combined responses of stream ecosystems to multiple stressors. J. Appl. Ecol. 2008, 45, 1810–1819. [Google Scholar] [CrossRef]
- Bonada, N.; Prat, N.; Resh, V.H.; Statzner, B. Developments in aquatic insect biomonitoring: A comparative analysis of recent approaches. Annu. Rev. Entomol. 2006, 51, 495–523. [Google Scholar] [CrossRef] [Green Version]
- Serrano Balderas, E.C.; Grac, C.; Berti-Equille, L.; Armienta Hernandez, M.A. Potential application of macroinvertebrates indices in bioassessment of Mexican streams. Ecol. Indic. 2016, 61, 558–567. [Google Scholar] [CrossRef]
- Cortelezzi, A.; Sierra, M.V.; Gomez, N.; Marinelli, C.; Capitulo, A.R. Macrophytes, epipelic biofilm, and invertebrates as biotic indicators of physical habitat degradation of lowland streams (Argentina). Environ. Monit. Assess. 2013, 185, 5801–5815. [Google Scholar] [CrossRef]
- Blann, K.L.; Anderson, J.L.; Sands, G.R.; Vondracek, B. Effects of Agricultural Drainage on Aquatic Ecosystems: A Review. Crit. Rev. Environ. Sci. Technol. 2009, 39, 909–1001. [Google Scholar] [CrossRef]
- Matomela, N.; Chakona, A.; Kadye, W. Comparative assessment of macroinvertebrate communities within three Afromontane headwater streams influenced by different land use patterns. Ecol. Indic. 2021, 129, 107972. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Z.; Castro, D.M.P.; Tan, X.; Jiang, X.; Meng, X.; Ge, Y.; Xie, Z. Effects of different types of land-use on taxonomic and functional diversity of benthic macroinvertebrates in a subtropical river network. Environ. Sci. Pollut. Res. 2021, 28, 44339–44353. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.L.; Resh, V.H. After site selection and before data analysis: Sampling, sorting, and laboratory procedures used in stream benthic macroinvertebrate monitoring programs by USA state agencies. J. N. Am. Benthol. Soc. 2001, 20, 658–682. [Google Scholar] [CrossRef] [Green Version]
- Menezes, S.; Baird, D.J.; Soares, A.M.V.M. Beyond taxonomy: A review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Gayraud, S.; Statzner, B.; Bady, P.; Haybachp, A.; Schöll, F.; Usseglio-Polatera, P.; Bacchi, M. Invertebrate traits for the biomonitoring of large European rivers: An initial assessment of alternative metrics. Freshw. Biol. 2003, 48, 2045–2064. [Google Scholar] [CrossRef] [Green Version]
- Bady, P.; Dolédec, S.; Fesl, C.; Gayraud, S.; Bacchi, M.; Schöll, F. Use of invertebrate traits for the biomonitoring of European large rivers: The effects of sampling effort on genus richness and functional diversity. Freshw. Biol. 2005, 50, 159–173. [Google Scholar] [CrossRef]
- Doledec, S.; Statzner, B. Invertebrate traits for the biomonitoring of large European rivers: An assessment of specific types of human impact. Freshw. Biol. 2008, 53, 617–634. [Google Scholar] [CrossRef]
- Dolédec, S.; Phillips, N.; Townsend, C. Invertebrate community responses to land use at a broad spatial scale: Trait and taxonomic measures compared in New Zealand rivers. Freshw. Biol. 2011, 56, 1670–1688. [Google Scholar] [CrossRef]
- Townsend, C.R.; Hildrew, A.G. Species traits in relation to a habitat templet for river systems. Freshw. Biol. 1994, 31, 265–275. [Google Scholar] [CrossRef]
- Zuellig, R.E.; Schmidt, T.S. Characterizing invertebrate traits in wadeable streams of the contiguous US: Differences among ecoregions and land uses. Freshw. Sci. 2012, 31, 1042–1056. [Google Scholar] [CrossRef] [Green Version]
- Alahuhta, J.; Erős, T.; Kärnä, O.-M.; Soininen, J.; Wang, J.; Heino, J. Understanding environmental change through the lens of trait-based, functional, and phylogenetic biodiversity in freshwater ecosystems. Environ. Rev. 2018, 27, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Song, D.; Ming, K.; Jin, X.; Wang, H.B.; Wang, L.; Liu, H.; Zhao, C.; Huo, T.B. Response of macroinvertebrate communities to land use and water quality in Wudalianchi Lake. Ecol. Evol. 2021, 11, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Castro-Lopez, D.; Rodriguez-Lozano, P.; Arias-Real, R.; Guerra-Cobian, V.; Prat, N. The Influence of Riparian Corridor Land Use on the Pesqueria River’s Macroinvertebrate Community (NE Mexico). Water 2019, 11, 1930. [Google Scholar] [CrossRef] [Green Version]
- de Castro, D.M.P.; Doledec, S.; Callisto, M. Land cover disturbance homogenizes aquatic insect functional structure in neotropical savanna streams. Ecol. Indic. 2018, 84, 573–582. [Google Scholar] [CrossRef]
- Firmiano, K.R.; Castro, D.M.P.; Linares, M.S.; Callisto, M. Functional responses of aquatic invertebrates to anthropogenic stressors in riparian zones of Neotropical savanna streams. Sci. Total Environ. 2021, 753, 141865. [Google Scholar] [CrossRef]
- Leitao, R.P.; Zuanon, J.; Mouillot, D.; Leal, C.G.; Hughes, R.M.; Kaufmann, P.R.; Villeger, S.; Pompeu, P.S.; Kasper, D.; de Paula, F.R.; et al. Disentangling the pathways of land use impacts on the functional structure of fish assemblages in Amazon streams. Ecography 2018, 41, 219–232. [Google Scholar] [CrossRef]
- Liu, S.; Dong, X.; Zhou, X.; Chen, S. Characteristic analysis of weather and climate memory in temperature: Taking Jinan and Qingdao as examples. J. Mar. Meteorol. 2022, 42, 107–114. [Google Scholar] [CrossRef]
- Li, H.; Wang, S.; Ji, G.; Zhang, L. Changes in land use and ecosystem service values in Jinan, China. Energy Procedia 2011, 5, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Jiang, X.; Li, Z.; Meng, X.; Heino, J.; Xie, Z.; Wang, X.; Yu, J. Changes in multiple facets of macroinvertebrate alpha diversity are linked to afforestation in a subtropical riverine natural reserve. Environ. Sci. Pollut. Res. 2018, 25, 36124–36135. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef] [Green Version]
- Edegbene, A.O.; Arimoro, F.O.; Odume, O.N. How does urban pollution influence macroinvertebrate traits in forested riverine systems? Water 2020, 12, 3111. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Lorenz, A.W.; Zimmermann, S.; Wepener, V.; Sures, B.; Smit, N.J.; Malherbe, W. A diversity and functional approach to evaluate the macroinvertebrate responses to multiple stressors in a small subtropical austral river. Ecol. Indic. 2021, 131, 108206. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, Y.; Gao, Y.; Cui, D.; Zhang, W.; Jiao, Z.; Yao, F.; Zhang, Z.; Yang, H. The Impacts of Different Anthropogenic Disturbances on Macroinvertebrate Community Structure and Functional Traits of Glacier-Fed Streams in the Tianshan Mountains. Water 2022, 14, 1298. [Google Scholar] [CrossRef]
- Kuzmanovic, M.; Dolédec, S.; de Castro-Catala, N.; Ginebreda, A.; Sabater, S.; Muñoz, I.; Barceló, D. Environmental stressors as a driver of the trait composition of benthic macroinvertebrate assemblages in polluted Iberian rivers. Environ. Res. 2017, 156, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.M.; Alonso, M.L.S.; Gutierrez, M. Biological traits of stream macroinvertebrates from a semi-arid catchment: Patterns along complex environmental gradients. Freshw. Biol. 2008, 53, 1–21. [Google Scholar] [CrossRef]
- Edegbene, A.O.; Arimoro, F.O.; Odume, O.N. Exploring the distribution patterns of macroinvertebrate signature traits and ecological preferences and their responses to urban and agricultural pollution in selected rivers in the Niger Delta ecoregion, Nigeria. Aquat. Ecol. 2020, 54, 553–573. [Google Scholar] [CrossRef]
- Palmer, C.G.; Maart, B.; Palmer, A.R.; O’Keeffe, J.H. An assessment of macroinvertebrate functional feeding groups as water quality indicators in the Buffalo River, eastern Cape Province, South Africa. Hydrobiologia 1996, 318, 153–164. [Google Scholar] [CrossRef]
- Usseglio-Polatera, P.; Bournaud, M.; Richoux, P.; Tachet, H. Biological and ecological traits of benthic freshwater macroinvertebrates: Relationships and definition of groups with similar traits. Freshw. Biol. 2000, 43, 175–205. [Google Scholar] [CrossRef]
- Mondy, C.P.; Usseglio-Polatera, P. Using fuzzy-coded traits to elucidate the non-random role of anthropogenic stress in the functional homogenisation of invertebrate assemblages. Freshw. Biol. 2014, 59, 584–600. [Google Scholar] [CrossRef]
- Chevene, F.; DolÉAdec, S.; Chessel, D. A fuzzy coding approach for the analysis of long-term ecological data. Freshw. Biol. 1994, 31, 295–309. [Google Scholar] [CrossRef]
- Frank, C.A. Exploring the community structure of Afrotropical macroinvertebrate traits and ecological preferences along an agricultural pollution gradient in the Kat River, Eastern Cape, South Africa. Ecol. Indic. 2022, 135, 108570. [Google Scholar] [CrossRef]
- Dolédec, S.; Statzner, B.; Bournard, M. Species traits for future biomonitoring across ecoregions: Patterns along a human-impacted river. Freshw. Biol. 1999, 42, 737–758. [Google Scholar] [CrossRef]
- Huang, X.; Chen, W.; Cai, Q. Survey, observation and analysis of lake ecology. In Standard Methods for Observation and Analysis in Chinese Ecosystem Research Network, Series V; Standards Press of China: Beijing, China, 1999. [Google Scholar]
- Morse, J.C.; Yang, L.; Tian, L. Aquatic Insects of China Useful for Monitoring Water Quality; Hohai University Press: Nanjing, China, 1984. [Google Scholar]
- Kawai, S. Aquatic Insects of Japan: Manual with Keys and Illustrations; Tokai University Press: Tokyo, Japan, 1925. [Google Scholar]
- Brinkhurst, R.O. Guide to the Freshwater Aquatic Microdrile Oligochaetes of North America; Department of Fisheries and Oceans: Vancouver, BC, Canada, 1986. [Google Scholar]
- Epler, J.H. Identification Manual for the Larval Chironomidae (Diptera) of Florida; Bureau of Surface Water Management, Florida Department of Environmental Protection: Tallahassee, FL, USA, 1995. [Google Scholar]
- Merritt, R.W.; Cummins, K.W. An Introduction to the Aquatic Insects of North America; Kendall Hunt: Dubuque, IA, USA, 1996. [Google Scholar]
- Wiggins, G.B. Key to Larvae of North American Families of Trichoptera; University of Toronto Press: Toronto, ON, Canada, 1996; pp. 43–48. [Google Scholar]
- Laliberte, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Cornwell, W.; Schwilk, D.; Ackerly, D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.E.; Milner, A.M. Rapid loss of glacial ice reveals stream community assembly processes. Glob. Chang. Biol. 2012, 18, 2195–2204. [Google Scholar] [CrossRef] [Green Version]
- Pavoine, S.; Dolédec, S. The apportionment of quadratic entropy: A useful alternative for partitioning diversity in ecological data. Environ. Ecol. Stat. 2005, 12, 125–138. [Google Scholar] [CrossRef]
- Mason, N.; Mouillot, D.; Lee, W.G.; Setl, W.H. Functional Richness, Functional Evenness and Functional Divergence: The Primary Components of Functional Diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Dolédec, S.; Chessel, D.T.; ter Braak, C.; ter Braak, C.J.F.; Champely, S. Matching species traits to environmental variables: A new three-table ordination method. Environ. Ecol. Stat. 1996, 3, 143–166. [Google Scholar] [CrossRef]
- Pallottini, M.; Cappelletti, D.; Fabrizi, A.; Gaino, E.; Goretti, E.; Selvaggi, R.; Céréghino, R. Macroinvertebrate functional trait responses to chemical pollution in agricultural–industrial landscapes. River Res. Appl. 2017, 33, 505–513. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring functional diversity from multiple traits, and other tools for functional ecology. R Package Version 2014, 1, 1–12. [Google Scholar]
- Dray, S.; Dufour, A.-B. The ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Dray, S.; Choler, P.; Dolédec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, M.; Burrows, R.M.; Lidman, J.; Fältström, E.; Laudon, H.; Sponseller, R.A. Land use influences macroinvertebrate community composition in boreal headwaters through altered stream conditions. Ambio 2017, 46, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Luo, K.; Hu, X.; He, Q.; Wu, Z.; Cheng, H.; Hu, Z.; Mazumder, A. Impacts of rapid urbanization on the water quality and macroinvertebrate communities of streams: A case study in Liangjiang New Area, China. Sci. Total Environ. 2018, 621, 1601–1614. [Google Scholar] [CrossRef]
- Martins, R.; Couceiro, S.; Melo, A.; Moreira, M.; Hamada, N. Effects of urbanization on stream benthic invertebrate communities in Central Amazon. Ecol. Indic. 2017, 73, 480–491. [Google Scholar] [CrossRef]
- Schriever, T.A.; Bogan, M.T.; Boersma, K.S.; Cañedo-Argüelles, M.; Jaeger, K.L.; Olden, J.D.; Lytle, D.A. Hydrology shapes taxonomic and functional structure of desert stream invertebrate communities. Freshw. Sci. 2015, 34, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Gerisch, M.; Agostinelli, V.; Henle, K.; Dziock, F. More species, but all do the same: Contrasting effects of flood disturbance on ground beetle functional and species diversity. Oikos 2012, 121, 508–515. [Google Scholar] [CrossRef]
- Péru, N.; Dolédec, S. From compositional to functional biodiversity metrics in bioassessment: A case study using stream macroinvertebrate communities. Ecol. Indic. 2010, 10, 1025–1036. [Google Scholar] [CrossRef]
- Fierro, P.; Valdovinos, C.; Lara, C.; Saldias, G.S. Influence of Intensive Agriculture on Benthic Macroinvertebrate Assemblages and Water Quality in the Aconcagua River Basin (Central Chile). Water 2021, 13, 492. [Google Scholar] [CrossRef]
- Odume, O.N. Searching for urban pollution signature and sensitive macroinvertebrate traits and ecological preferences in a river in the Eastern Cape of South Africa. Ecol. Indic. 2020, 108, 105759. [Google Scholar] [CrossRef]
- Odume, O.N.; Palmer, C.G.; Arimoro, F.O.; Mensah, P.K. Patterns of chironomid body-size distribution in an effluent-impacted river in the Eastern Cape, South Africa. Afr. J. Aquat. Sci. 2014, 39, 377–386. [Google Scholar] [CrossRef]
- Statzner, B.; BÊChe, L.A. Can biological invertebrate traits resolve effects of multiple stressors on running water ecosystems? Freshw. Biol. 2010, 55, 80–119. [Google Scholar] [CrossRef]
- Feio, M.J.; Dolédec, S. Integration of invertebrate traits into predictive models for indirect assessment of stream functional integrity: A case study in Portugal. Ecol. Indic. 2012, 15, 236–247. [Google Scholar] [CrossRef]
- Tomanova, S.; Usseglio-Polatera, P. Patterns of benthic community traits in neotropical streams: Relationship to mesoscale spatial variability. Fundam. Appl. Limnol. 2007, 170, 243–255. [Google Scholar] [CrossRef]
- Bryce, S.A.; Lomnicky, G.A.; Kaufmann, P.R. Protecting sediment-sensitive aquatic species in mountain streams through the application of biologically based streambed sediment criteria. J. N. Am. Benthol. Soc. 2010, 29, 657–672. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.L.; Swan, C.M. Dendritic network structure constrains metacommunity properties in riverine ecosystems. J. Anim. Ecol. 2010, 79, 571–580. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Miserendino, M.L.; Masi, C.I. The effects of land use on environmental features and functional organization of macroinvertebrate communities in Patagonian low order streams. Ecol. Indic. 2010, 10, 311–319. [Google Scholar] [CrossRef]
- Statzner, B. Mineral grains in caddisfly pupal cases and streambed sediments: Assessing resource use and its limitation across various river types. Ann. Limnol.-Int. J. Limnol. 2011, 47, 103–118. [Google Scholar] [CrossRef]
- Burdon, F.J.; Harding, M. Habitat loss drives threshold response of benthic invertebrate communities to deposited sediment in agricultural streams. Ecol. Appl. 2013, 23, 1036–1047. [Google Scholar] [CrossRef] [PubMed]
- Edegbene, A.O.; Adam, M.B.; Gambo, J.; Osimen, E.C.; Ikomi, R.B.; Ogidiaka, E.; Omovoh, G.O.; Akamagwuna, F.C. Searching for indicator macroinvertebrate traits in an Afrotropical riverine system: Implication for ecosystem biomonitoring and sustainability. Environ. Monit. Assess. 2021, 193, 711. [Google Scholar] [CrossRef]
- Chapman, L.J.; Schneider, K.R.; Apodaca, C.; Chapman, C.A. Respiratory ecology of macroinvertebrates in a swamp-river system of east Africa. Biotropica 2004, 36, 572–585. [Google Scholar] [CrossRef]
- Chadwick, M.A. Stream Ecology: Structure and Function of Running Waters. Freshw. Biol. 2008, 53, 1914. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Category | Code | Reference |
|---|---|---|---|
| Voltinism | Semivoltine | Volt1 | [3,32,44] |
| Univoltine | Volt2 | ||
| Bi- or multivoltine | Volt3 | ||
| Development | Fast seasonal | Deve1 | [32] |
| Slow seasonal | Deve2 | ||
| Nonseasonal | Deve3 | ||
| Dispersal | No | Disp1 | [3,32] |
| Low | Disp2 | ||
| High | Disp3 | ||
| Drift | Rare | Drft1 | [3,32] |
| Common | Drft2 | ||
| Very abundant | Drft3 | ||
| Attachment | Free-ranging | Atch1 | [32] |
| Sessile, sedentary | Atch2 | ||
| Both | Atch3 | ||
| Armoring | None (soft-bodied forms) | Armr1 | [32] |
| Poor (heavily sclerotized) | Armr2 | ||
| Good (e.g., some cased caddisflies) | Armr3 | ||
| Shape | Streamlined (flat, fusiform) | Shap1 | [32,44] |
| Not streamlined (cylindrical, round, or bluff) | Shap2 | ||
| Respiration | Tegument | Resp1 | [3,32,44] |
| Gills | Resp2 | ||
| Air (Plastron or spiracle) | Resp3 | ||
| Size | Small (<9 mm) | Size1 | [3,32,44] |
| Medium (9–16 mm) | Size2 | ||
| Large (>16 mm) | Size3 | ||
| Rheophily | Depositional only | Rheo1 | [3,32] |
| Depositional and erosional | Rheo2 | ||
| Erosional | Rheo3 | ||
| Habit | Burrow | Habit1 | [3,32,44] |
| Climb | Habit2 | ||
| Sprawl | Habit3 | ||
| Cling | Habit4 | ||
| Swim | Habit5 | ||
| Trophic habit | Collector–gatherer | Trop1 | [3,32,44] |
| Collector–filterer | Trop2 | ||
| Herbivore (scraper, piercer, and shedder) | Trop3 | ||
| Predator (piercer and engulfer) | Trop4 | ||
| Shredder (detritivore) | Trop5 |
| Variable | Mountain-Hilly Area | Urban Area | Agricultural Area | p | χ2 |
|---|---|---|---|---|---|
| Water temperature (°C) | 21.57 ± 4.89 | 21.48 ± 4.42 | 20.94 ± 4.88 | 0.930 | 0.145 |
| pH | 8.24 ± 0.37 | 7.91 ± 0.35 | 8.17 ± 0.33 | <0.001 | 33.132 |
| Electrical conductivity (EC) (s/cm) | 634.65 ± 393.51 | 1145.73 ± 700.55 | 1723.13 ± 863.66 | <0.001 | 118.534 |
| Dissolved oxygen concentration (DO) (mg/L) | 8.15 ± 1.52 | 7.44 ± 2.35 | 7.49 ± 2.45 | 0.199 | 3.222 |
| Total nitrogen (TN) (mg/L) | 3.94 ± 4.16 | 5.86 ± 4.49 | 4.69 ± 9.32 | <0.001 | 15.651 |
| Ammonium nitrogen (NH4-N) (mg/L) | 0.96 ± 3.11 | 1.19 ± 1.64 | 2.05 ± 8.78 | <0.001 | 11.931 |
| Nitrate nitrogen (NO3-N) (mg/L) | 2.2 ± 1.95 | 3.81 ± 3.73 | 1.35 ± 1.72 | <0.001 | 27.563 |
| Chemical oxygen demand (COD) (mg/L) | 20.56 ± 17.89 | 21.88 ± 14.28 | 32.18 ± 31.45 | <0.001 | 27.522 |
| Biochemical oxygen demand (BOD) (mg/L) | 2.97 ± 4.25 | 3.58 ± 3.76 | 5.48 ± 7.53 | <0.001 | 14.85 |
| Total phosphorus (TP) (mg/L) | 0.28 ± 0.69 | 0.33 ± 0.58 | 0.39 ± 1.02 | <0.001 | 24.121 |
| Farmland (%) | 34.31 ± 33.67 | 24.52 ± 24.33 | 59.90 ± 28.38 | <0.001 | 9.293 |
| Forest land (%) | 4.86 ± 16.36 | 6.074 | |||
| Grassland (%) | 2.91 ± 8.82 | 6.074 | |||
| Water area (%) | 24.50 ± 24.57 | 17.39 ± 28.76 | 12.35 ± 23.55 | 0.190 | 3.319 |
| Urban land (%) | 30.40 ± 28.05 | 58.03 ± 41.09 | 26.52 ± 26.75 | 0.120 | 4.233 |
| Axes1 | Axes2 | |
|---|---|---|
| Eigenvalue | 18.12 | 3.10 |
| % of total co-inertia | 78.47% | 13.43% |
| Covariation | 4.26 | 1.76 |
| Correlation | 0.66 | 0.46 |
| Cumulative inertia (environment) | 3.70 | 6.53 |
| Ratio (environment) | 84.61% | 91.26% |
| Cumulative inertia (traits) | 11.35 | 16.47 |
| Ratio (traits) | 99.08% | 83.98% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, A.; Li, J.; Zheng, B.; Yin, X. Effect of Different Land Use Types on the Taxonomic and Functional Diversity of Macroinvertebrates in an Urban Area of Northern China. Water 2022, 14, 3793. https://doi.org/10.3390/w14233793
Lu A, Li J, Zheng B, Yin X. Effect of Different Land Use Types on the Taxonomic and Functional Diversity of Macroinvertebrates in an Urban Area of Northern China. Water. 2022; 14(23):3793. https://doi.org/10.3390/w14233793
Chicago/Turabian StyleLu, Aoran, Jiaxin Li, Biao Zheng, and Xuwang Yin. 2022. "Effect of Different Land Use Types on the Taxonomic and Functional Diversity of Macroinvertebrates in an Urban Area of Northern China" Water 14, no. 23: 3793. https://doi.org/10.3390/w14233793