
Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans


Abstract
:1. Introduction
2. Materials and Methods
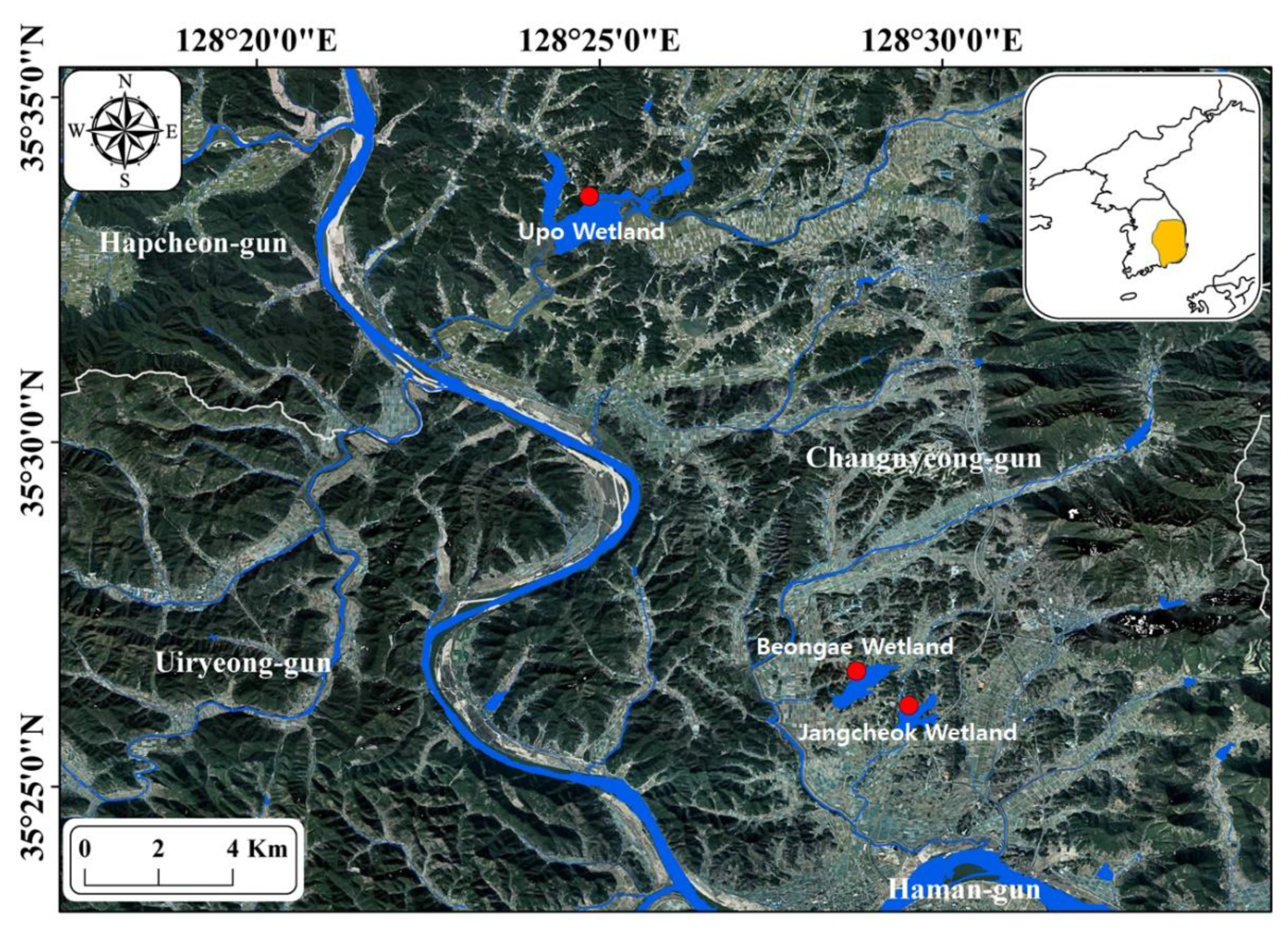
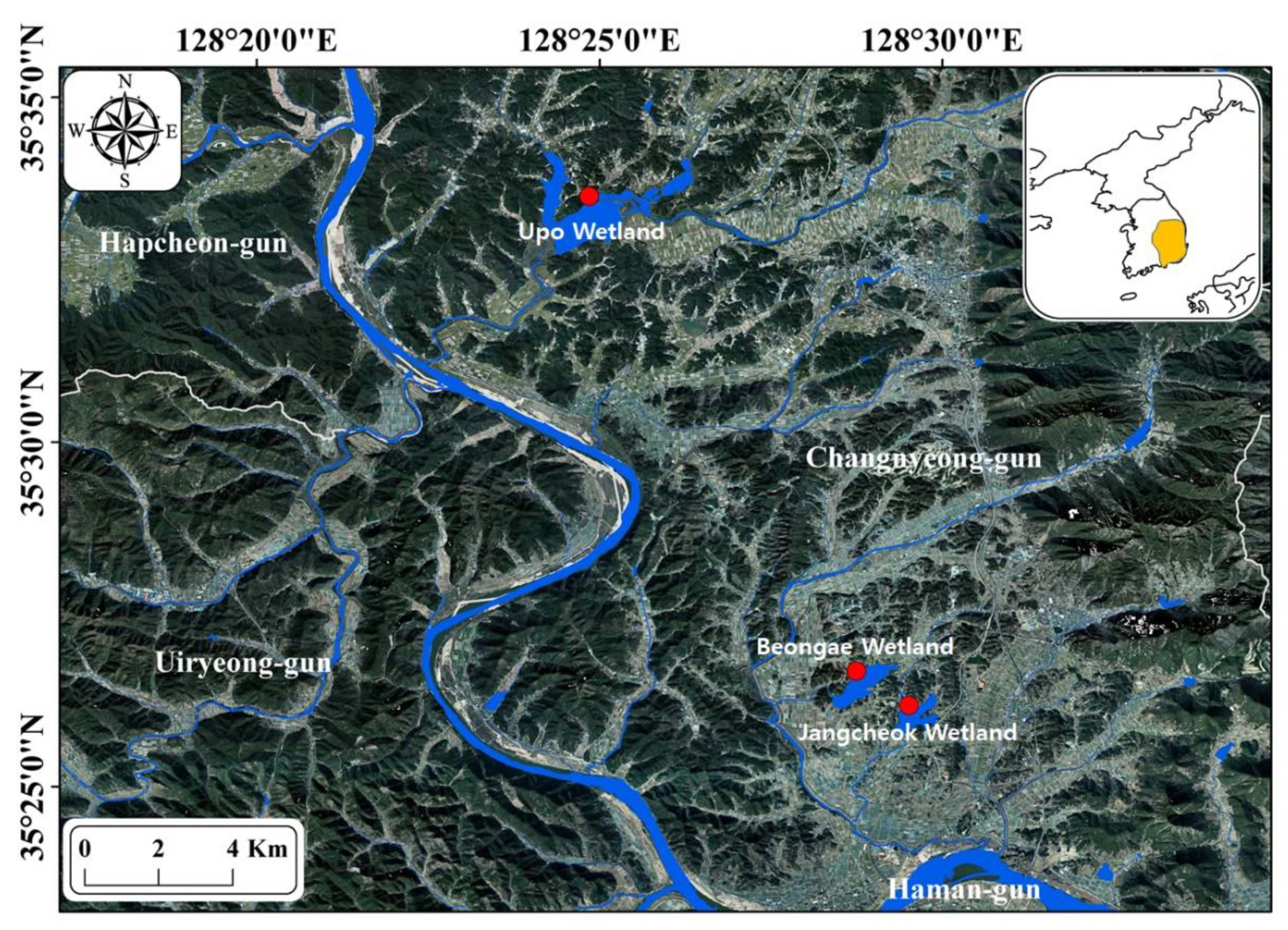
2.1. Study Site
2.2. Monitoring Strategy
2.3. Data Analysis
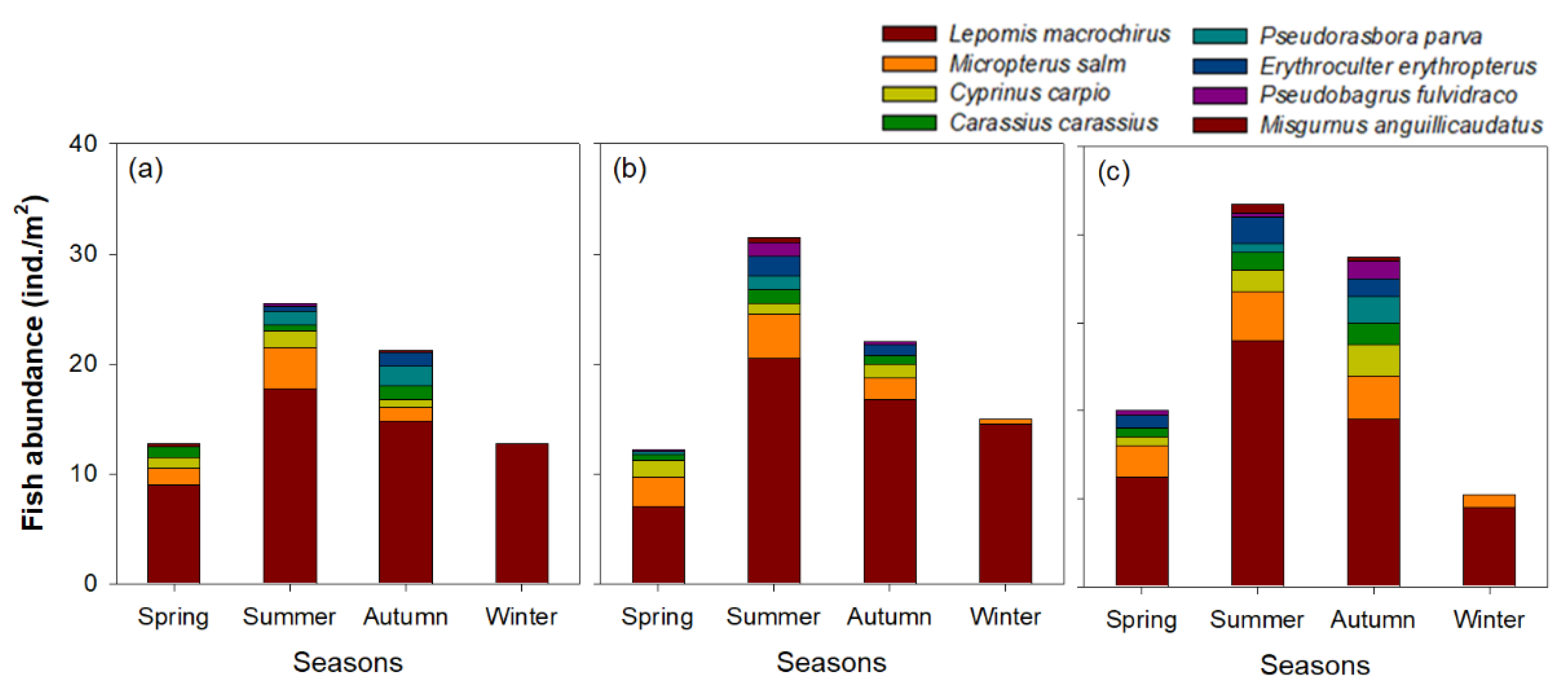
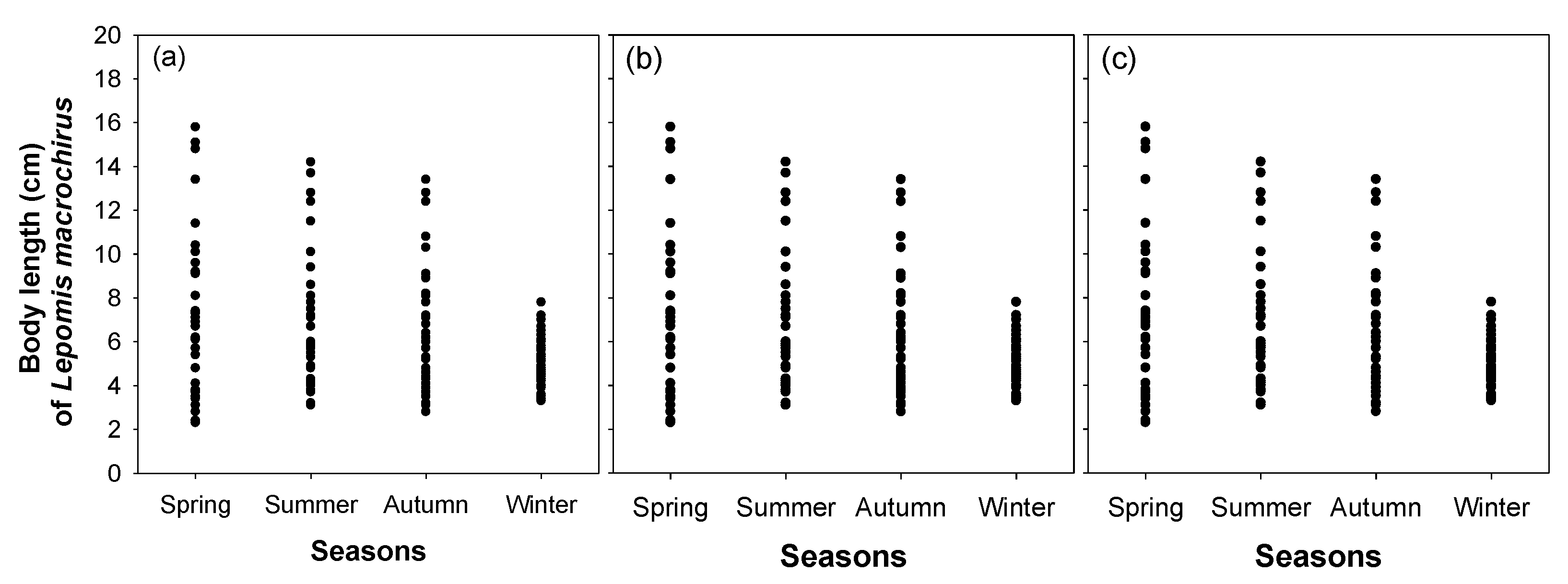
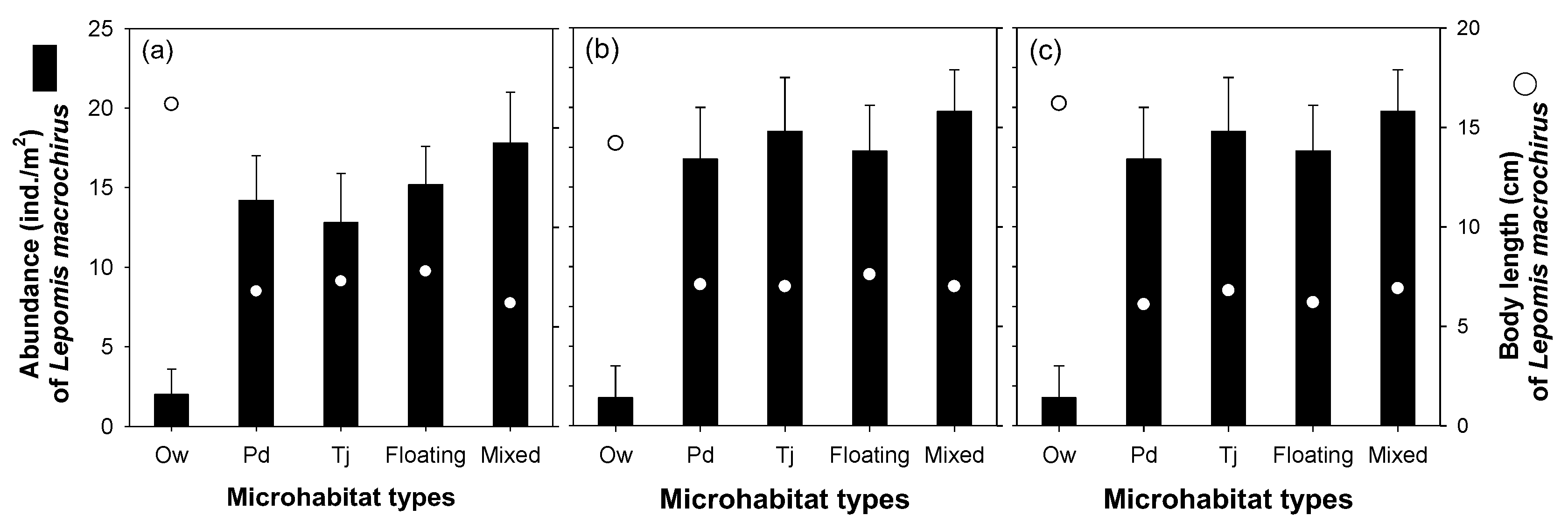
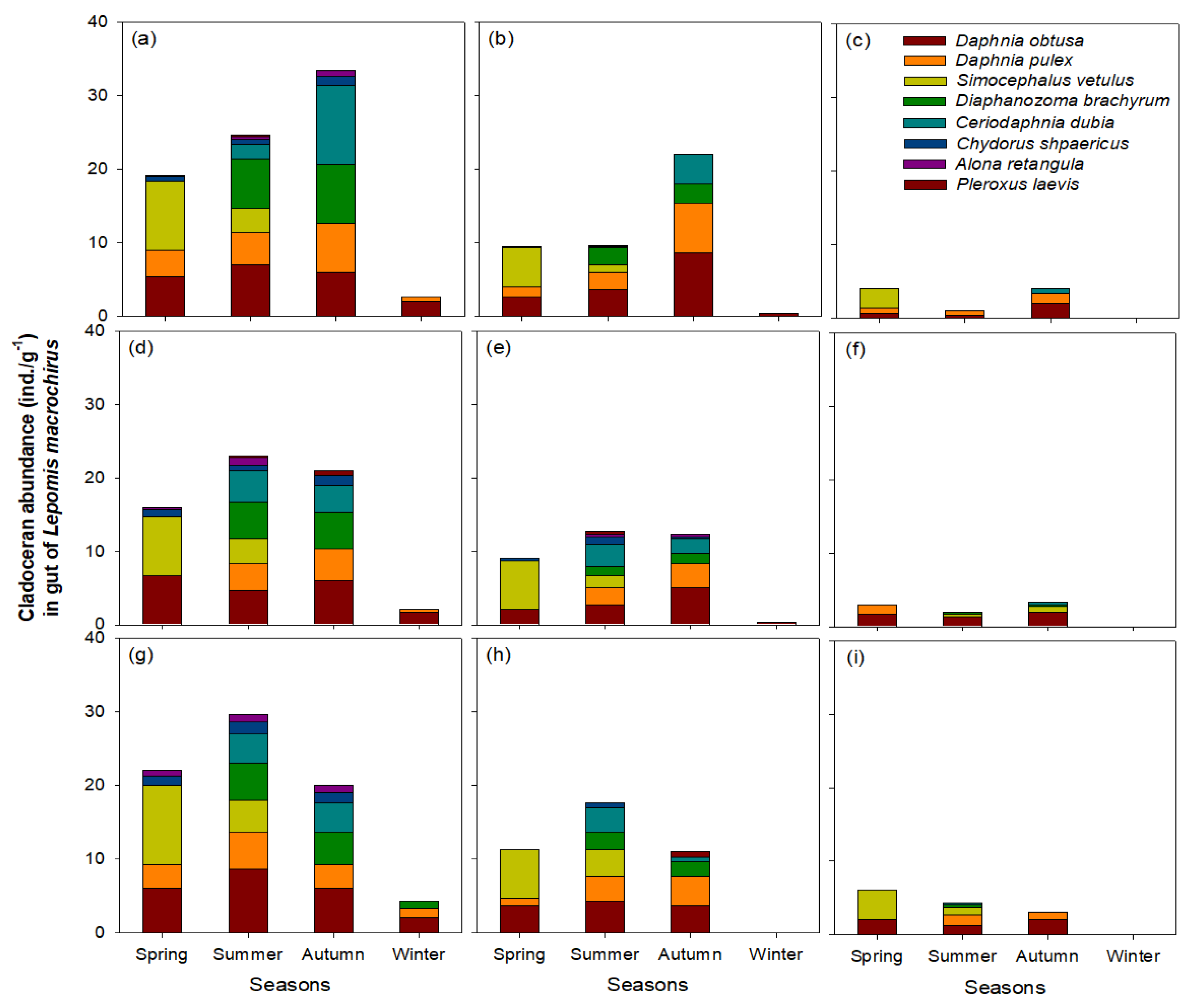
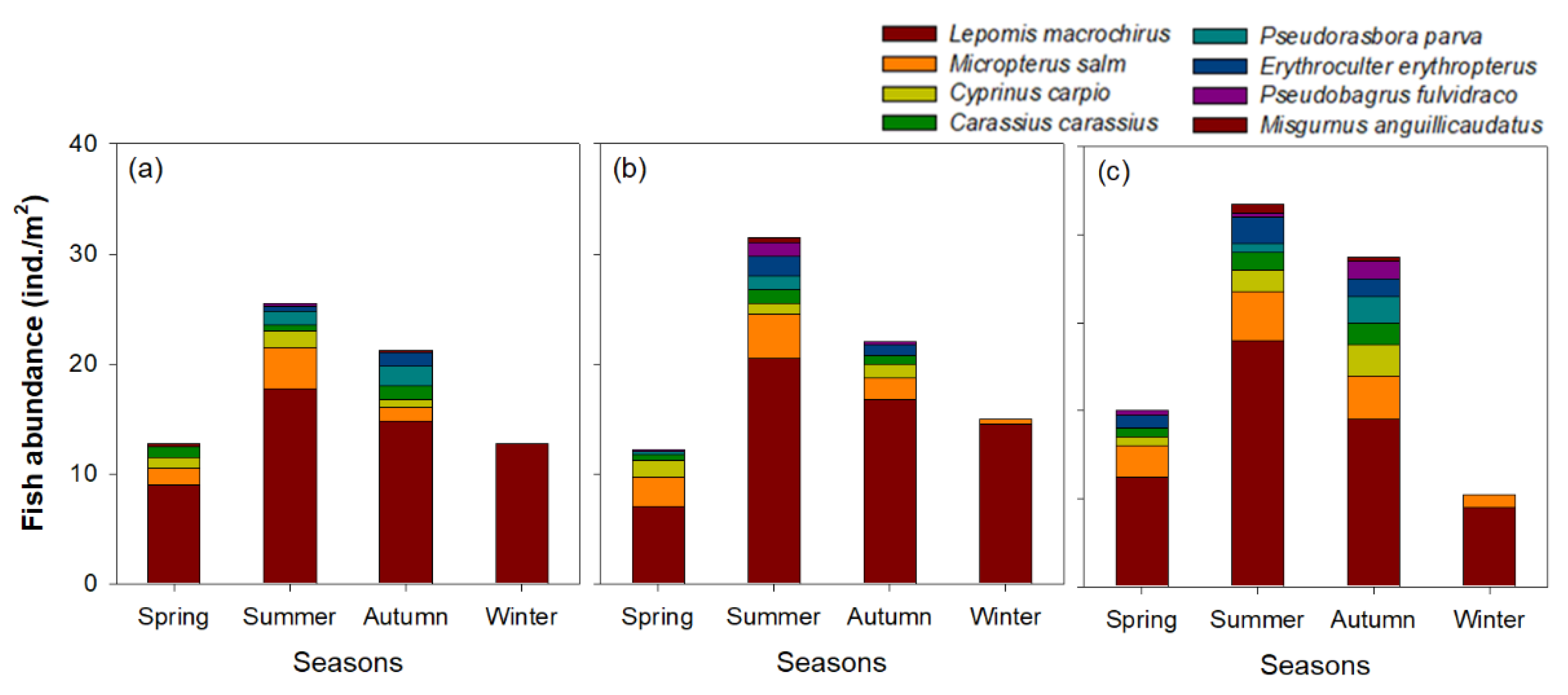
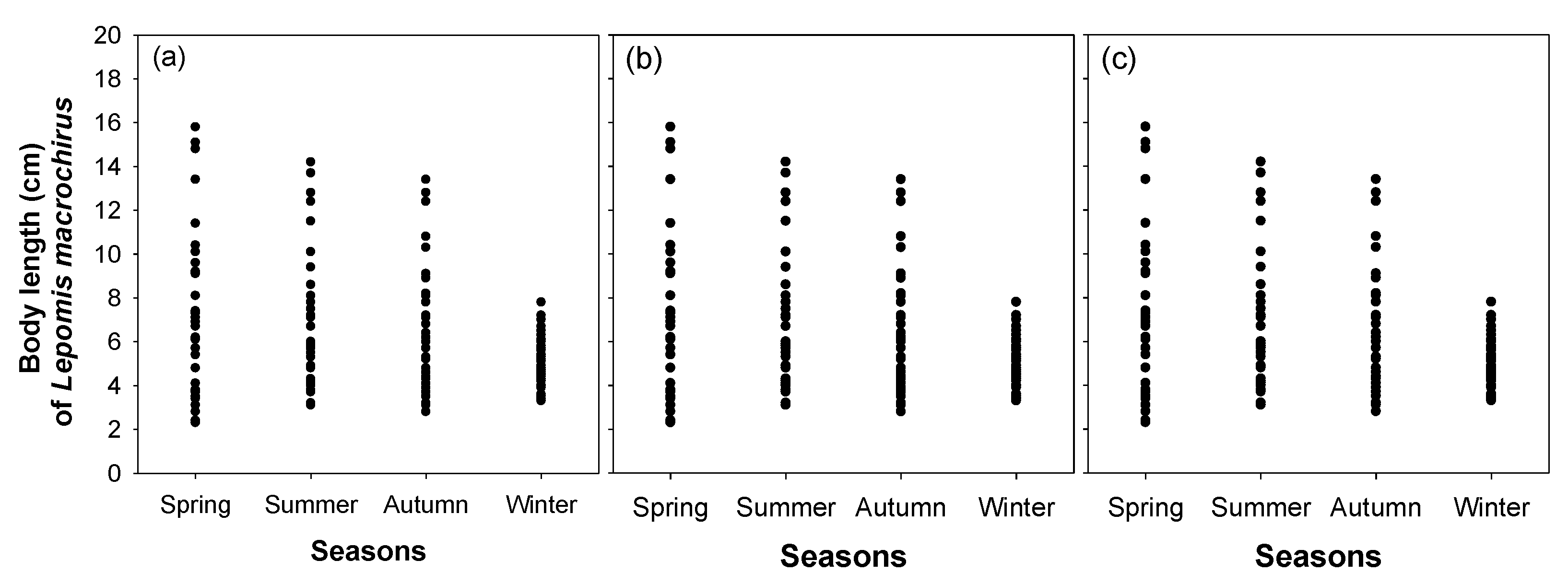
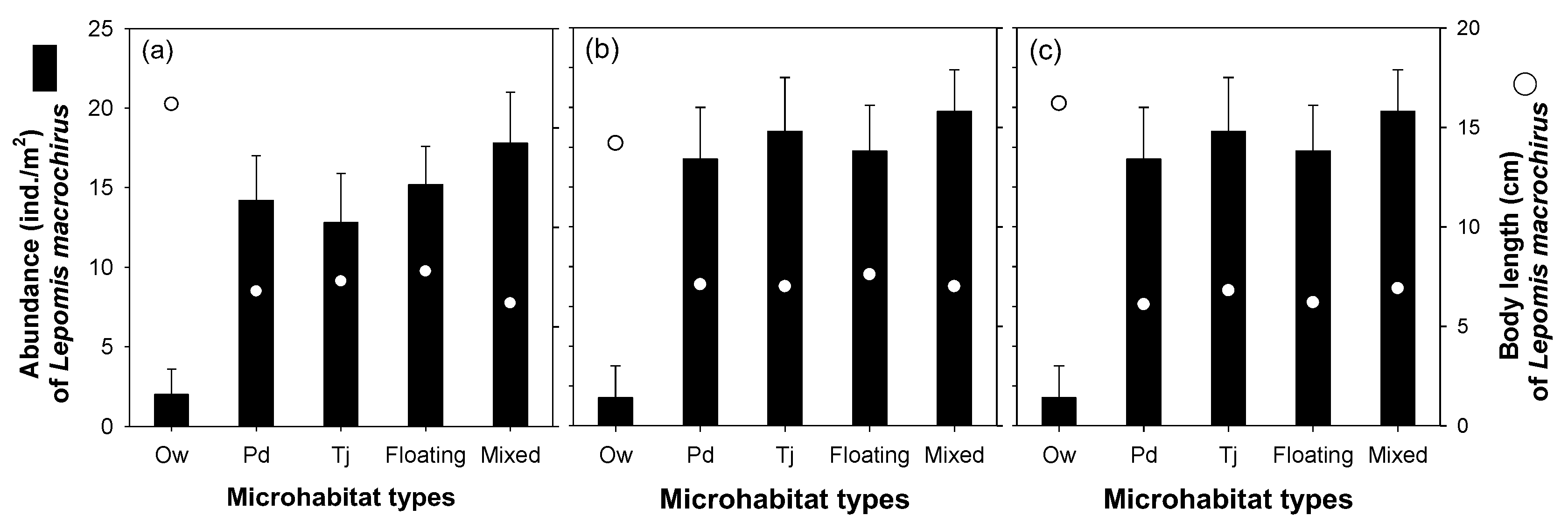
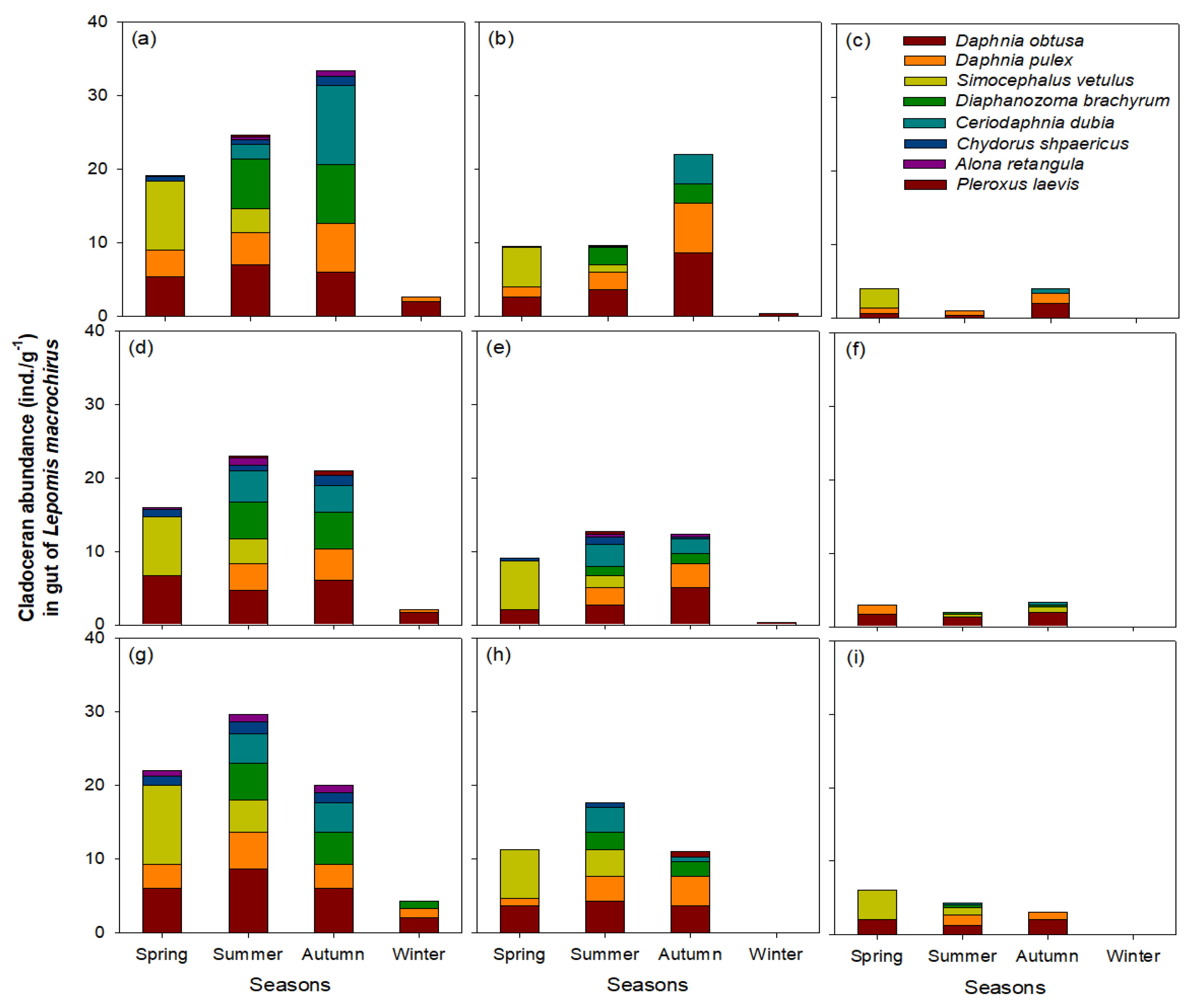
3. Results
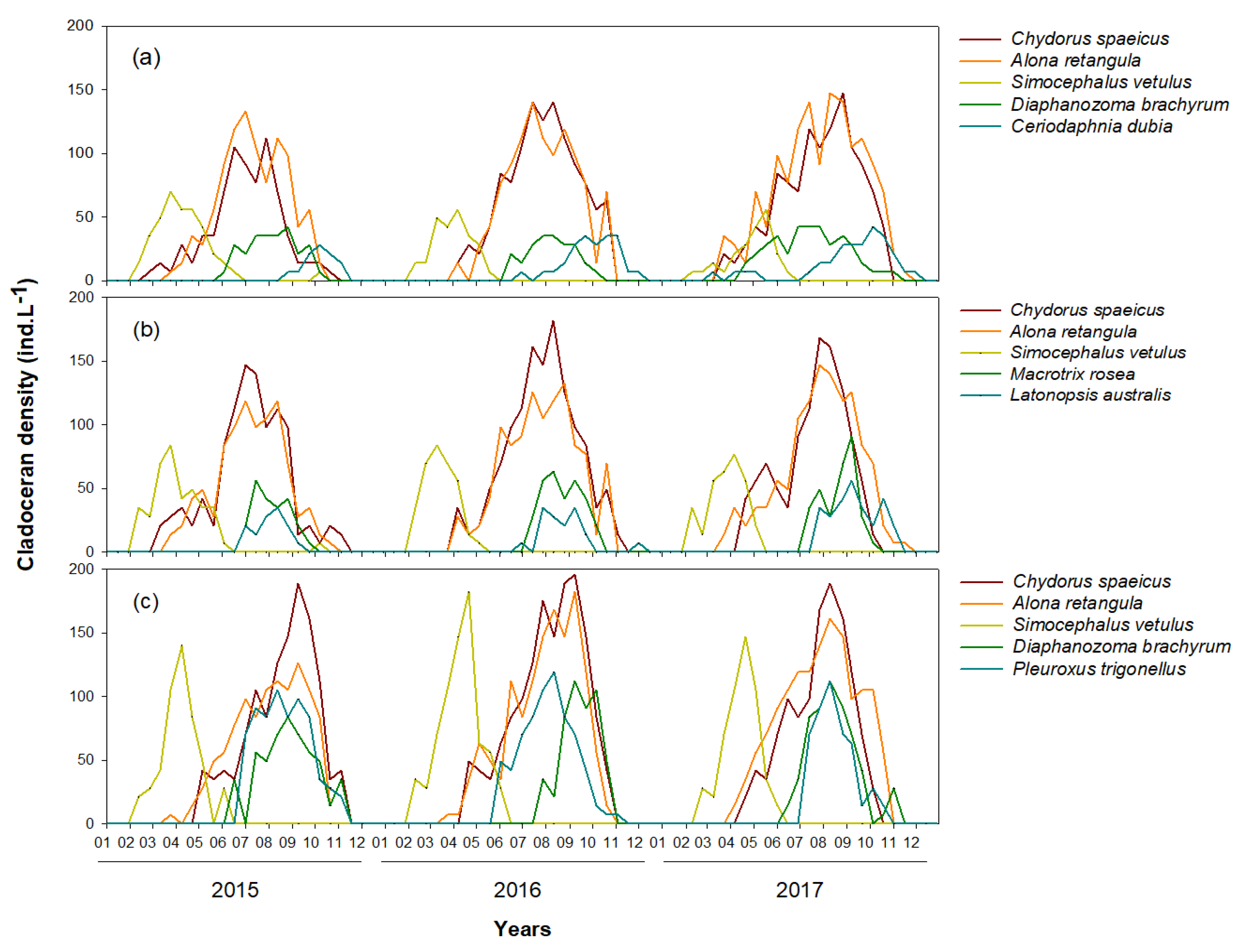
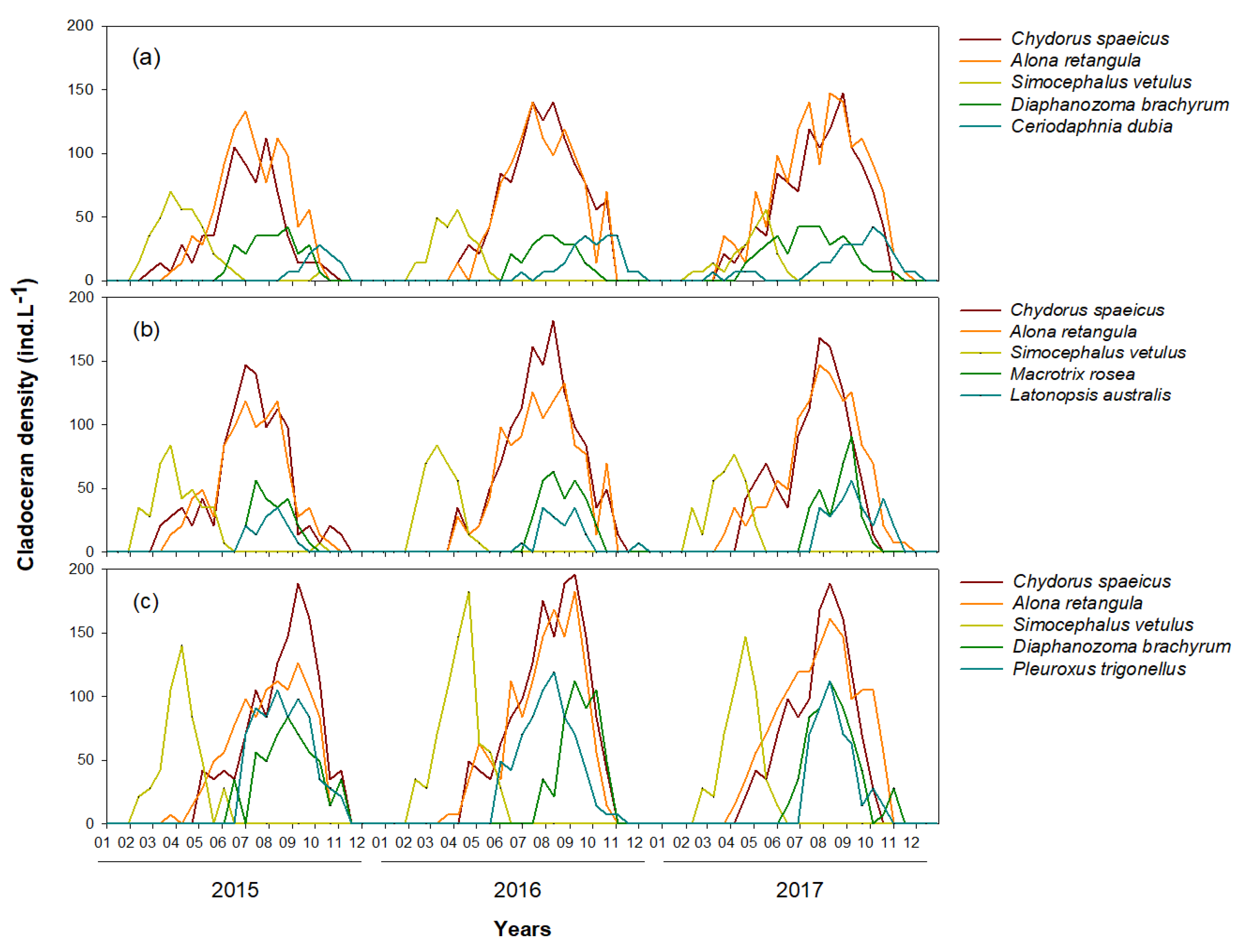
3.1. Environmental Variables and Cladoceran Distributions
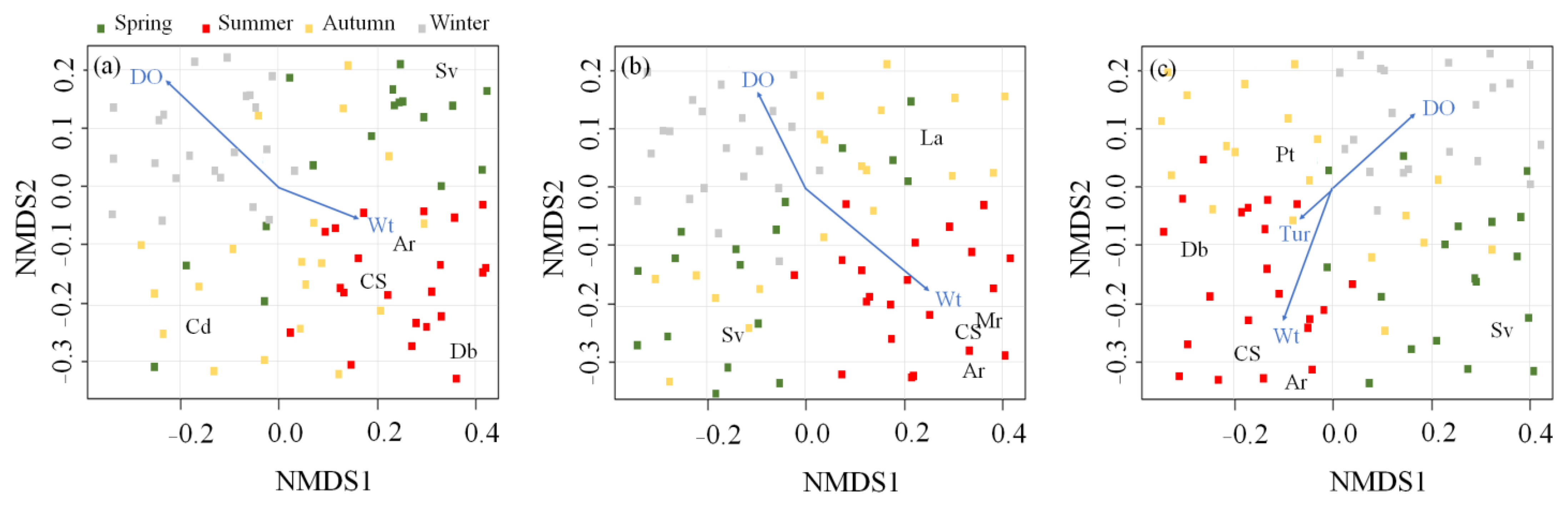
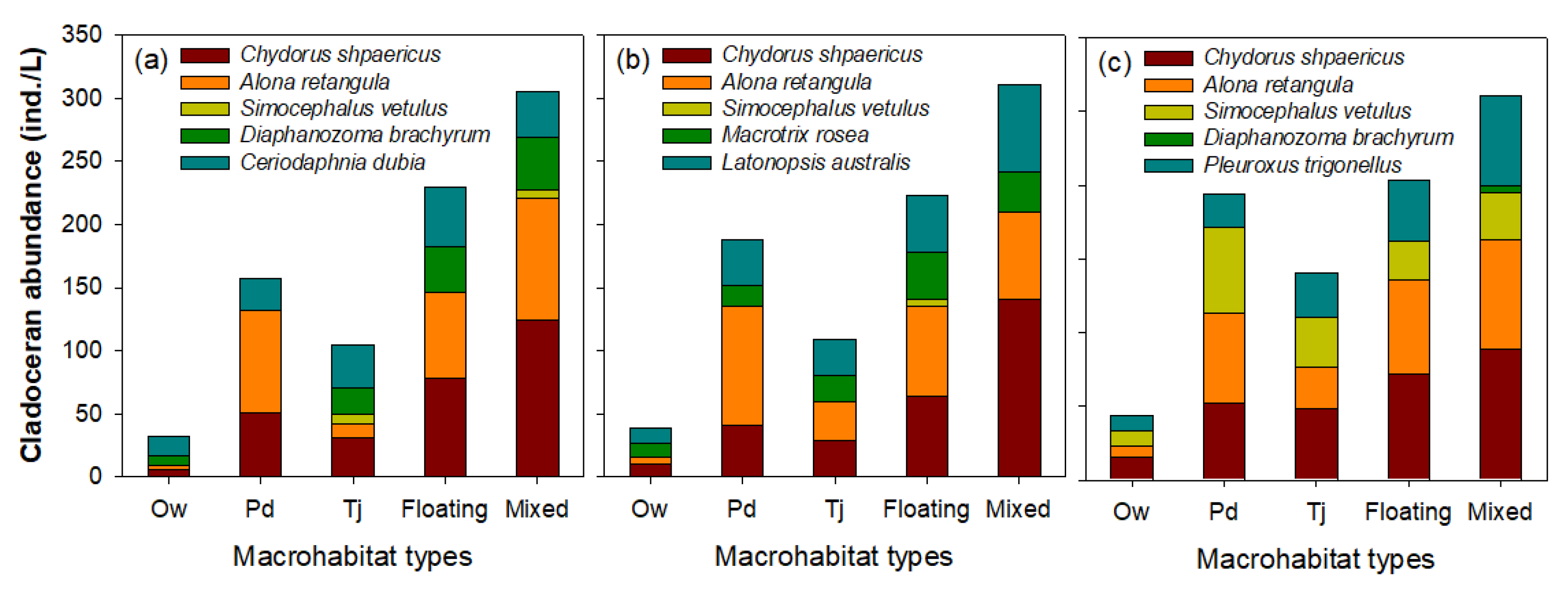
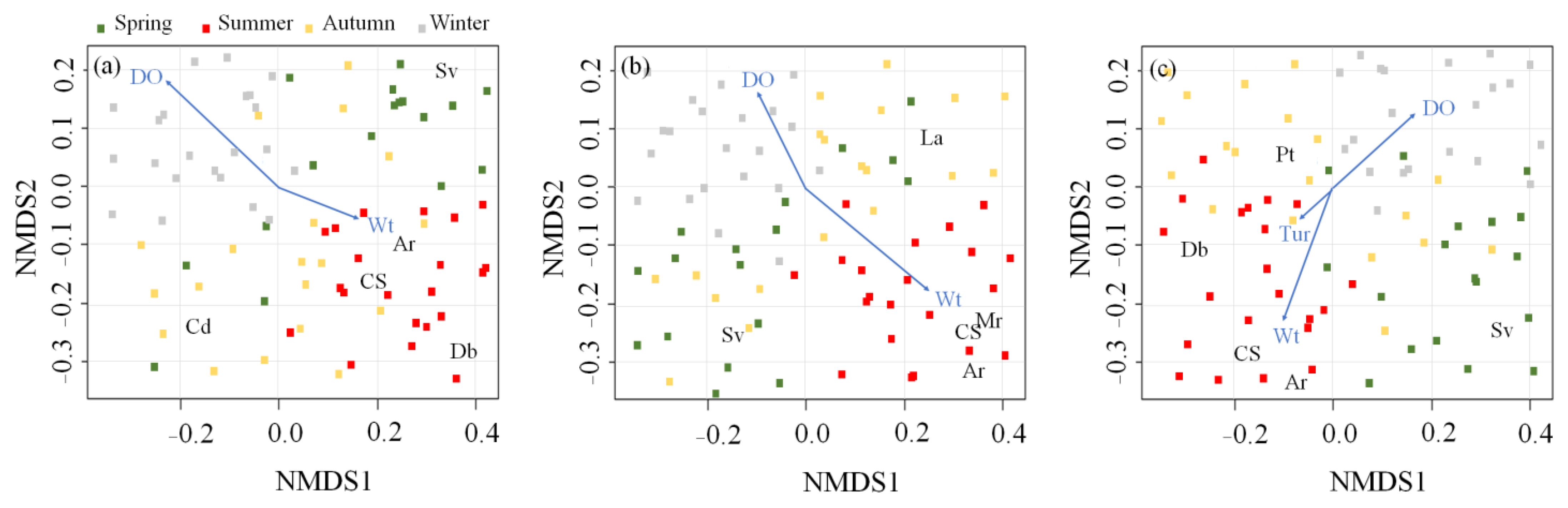
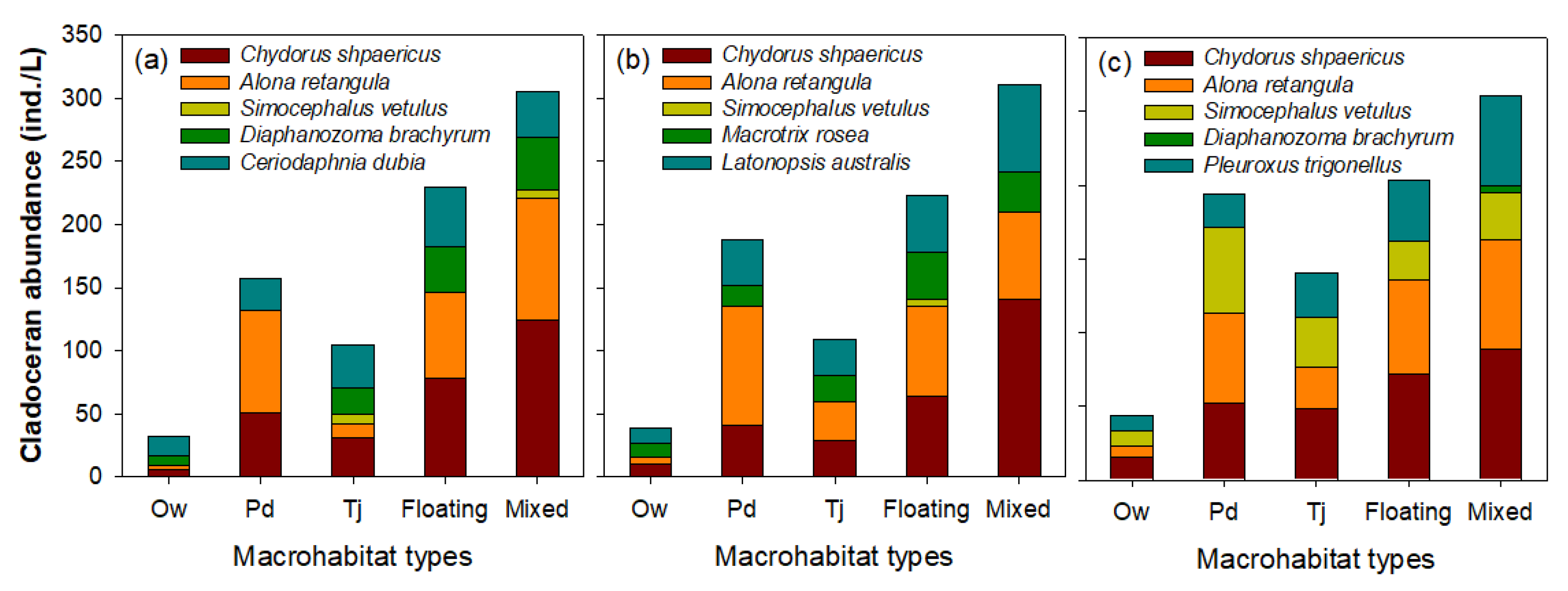
3.2. Effect of Microhabitat Types on Cladoceran Distribution
3.3. Effect of Microhabitat Characteristics on L. macrochirus Distribution
3.4. Utilization of Cladocerans as Food Source for L. macrochirus
4. Discussion
4.1. Distribution Patterns of Cladocerans and Fish Communities
4.2. Influence of L. macrochirus on Cladoceran Community Structure
4.3. Effect of Exotic Fish Introduction on Cladoceran Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caswell, H. Community structure: A neutral model analysis. Ecol. Monogr. 1976, 46, 327–354. [Google Scholar] [CrossRef]
- Devries, P.J.; Walla, T.R. Species diversity and community structure in neotropical fruit-feeding butterflies. Biol. J. Linn. Soc. Lond. 2001, 74, 1–15. [Google Scholar] [CrossRef]
- Heino, J. A macroecological perspective of diversity patterns in the freshwater realm. Freshw. Biol. 2011, 56, 1703–1722. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Violle, C.; Spasojevic, M.J.; McGill, B.; Damschen, E.; Harrison, S.; Enquist, B.J. Intra-specific and inter-specific variation in specific leaf area reveal the importance of abiotic and biotic drivers of species diversity across elevation and latitude. J. Veg. Sci. 2013, 24, 921–931. [Google Scholar] [CrossRef]
- Menge, B.A. Organization of the New England rocky intertidal community: Role of predation, competition, and environmental heterogeneity. Ecol. Monogr. 1976, 46, 355–393. [Google Scholar] [CrossRef]
- Chesson, P.; Kuang, J.J. The interaction between predation and competition. Nature 2008, 456, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Hambright, K.D.; Zohary, T. Phytoplankton species diversity control through competitive exclusion and physical disturbances. Limnol. Oceanogr. 2000, 45, 110–122. [Google Scholar] [CrossRef]
- Del Pozo, R.; Fernández-Aláez, C.; Fernández-Aláez, M. The relative importance of natural and anthropogenic effects on community composition of aquatic macrophytes in Mediterranean ponds. Mar. Freshw. Res. 2011, 62, 101–109. [Google Scholar] [CrossRef]
- Ball, S.L.; Baker, R.L. The non-lethal effects of predators and the influence of food availability on life history of adult Chironomus tentans (Diptera: Chironomidae). Freshw. Biol. 1995, 34, 1–12. [Google Scholar] [CrossRef]
- Casatti, L.; Mendes, H.F.; Ferreira, K.M. Aquatic macrophytes as feeding site for small fishes in the Rosana Reservoir, Paranapanema River, Southeastern Brazil. Bra. J. Biol. 2003, 63, 213–222. [Google Scholar] [CrossRef]
- Nunn, A.D.; Tewson, L.H.; Cowx, I.G. The foraging ecology of larval and juvenile fishes. Rev. Fish Biol. Fish. 2012, 22, 377–408. [Google Scholar] [CrossRef]
- Wipfli, M.S. Terrestrial invertebrates as salmonid prey and nitrogen sources in streams: Contrasting old-growth and young-growth riparian forests in southeastern Alaska, USA. Can. J. Fish Aquat. Sci. 1997, 54, 1259–1269. [Google Scholar] [CrossRef]
- Dodson, S.L.; Cáceres, C.E.; Rogers, D.C. Cladocera and other Branchiopoda. In Ecology and Classification of North American Freshwater Invertebrates; Academic Press: Cambridge, MA, USA, 2010; pp. 773–827. [Google Scholar]
- Goulden, C.E.; Henry, L.L.; Tessier, A.J. Body size, energy reserves, and competitive ability in three species of Cladocera. Ecology 1982, 63, 1780–1789. [Google Scholar] [CrossRef]
- Ramírez-García, P. Life table demography and population growth of Daphnia laevis (Cladocera, Anomopoda) under different densities of Chlorella vulgaris and Microcystis aeruginosa. Crustaceana 2000, 73, 1273–1286. [Google Scholar] [CrossRef]
- Kirk, K.L.; Gilbert, J.J. Suspended clay and the population dynamics of planktonic rotifers and cladocerans. Ecology 1990, 71, 1741–1755. [Google Scholar] [CrossRef]
- Gayosso-Morales, M.A.; Nandini, S.; Martínez-Jeronimo, F.F.; Sarma, S.S.S. Fish-mediated zooplankton community structure in shallow turbid waters: A mesocosm study. Wetl. Ecol. Manag. 2019, 27, 651–661. [Google Scholar] [CrossRef]
- Tollrian, R.; Dodson, S.I. Inducible defenses in Cladocera: Constraints, costs, and multipredator environments. In The Ecology and Evolution of Inducible Defenses; Princeton University Press: Princeton, NJ, USA, 1999; pp. 177–202. [Google Scholar]
- Dzialowski, A.R.; Lennon, J.T.; O’Brien, W.J.; Smith, V.H. Predator-induced phenotypic plasticity in the exotic cladoceran Daphnia lumholtzi. Freshw. Biol. 2003, 48, 1593–1602. [Google Scholar] [CrossRef] [Green Version]
- Von Elert, E.; Pohnert, G. Predator specificity of kairomones in diel vertical migration of Daphnia: A chemical approach. Oikos 2000, 88, 119–128. [Google Scholar] [CrossRef]
- Jeppesen, E.; Lauridsen, T.L.; Kairesalo, T.; Perrow, M.R. Impact of submerged macrophytes on fish-zooplankton interactions in lakes. In The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; pp. 91–114. [Google Scholar]
- Choi, J.Y.; Jeong, K.S.; Kim, S.K.; La, G.H.; Chang, K.H.; Joo, G.J. Role of macrophytes as microhabitats for zooplankton community in lentic freshwater ecosystems of South Korea. Ecol. Inform. 2014, 24, 177–185. [Google Scholar] [CrossRef]
- Zeng, L.; He, F.; Dai, Z.; Xu, D.; Liu, B.; Zhou, Q.; Wu, Z. Effect of submerged macrophyte restoration on improving aquatic ecosystem in a subtropical, shallow lake. Ecol. Eng. 2017, 106, 578–587. [Google Scholar] [CrossRef]
- Kovalenko, K.E.; Thomaz, S.M.; Warfe, D.M. Habitat complexity: Approaches and future directions. Hydrobiologia 2012, 685, 1–17. [Google Scholar] [CrossRef]
- Rossa, D.C.; Lansac-Tôha, F.A.; Bonecker, C.C.; Velho, L.F.M. Abundance of cladocerans in the littoral regions of two environments of the upper Paraná river floodplain, Mato Grosso do Sul, Brazil. Braz. J. Biol. 2001, 61, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Dibble, E.D.; Pelicice, F.M. Influence of aquatic plant-specific habitat on an assemblage of small neotropical floodplain fishes. Ecol. Freshw. Fish 2010, 19, 381–389. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.K. Effects of aquatic macrophytes on spatial distribution and feeding habits of exotic fish species Lepomis macrochirus and Micropterus salmoides in shallow reservoirs in South Korea. Sustainability 2020, 12, 1447. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, M.; Hanazato, T.; Saji, A.; Nakazato, R. Migration from plant to plant: An important factor controlling densities of the epiphytic cladoceran Alona (Chydoridae, Anomopoda) on lake vegetation. Limnology 2004, 5, 17–23. [Google Scholar]
- Basińska, A.M.; Antczak, M.; Świdnicki, K.; Jassey, V.E.; Kuczyńska-Kippen, N. Habitat type as strongest predictor of the body size distribution of Chydorus sphaericus (OF Müller) in small water bodies. Int. Rev. Hydrobiol. 2014, 99, 382–392. [Google Scholar] [CrossRef]
- Belyaeva, M.A. Littoral Cladocera (Crustacea: Branchiopoda) from the Altai mountain lakes, with remarks on the taxonomy of Chydorus sphaericus (OF Müller, 1776). Arthropoda Sel. 2003, 12, 171–182. [Google Scholar]
- Adamczuk, M. Niche separation by littoral-benthic Chydoridae (Cladocera, Crustacea) in a deep lake-potential drivers of their distribution and role in littoral-pelagic coupling. J. Limnol. 2014, 73, 490–501. [Google Scholar] [CrossRef] [Green Version]
- De Eyto, E.; Irvine, K. The response of three chydorid species to temperature, pH and food. Hydrobiologia 2001, 459, 165–172. [Google Scholar] [CrossRef]
- Muro-Cruz, G.; Nandini, S.; Sarma, S.S.S. Comparative life table demography and population growth of Alona rectangula and Macrothrix triserialis (Cladocera: Crustacea) in relation to algal (Chlorella vulgaris) food density. J. Freshw. Ecol. 2002, 17, 1–11. [Google Scholar] [CrossRef]
- Jang, M.H.; Kim, J.G.; Park, S.B.; Jeong, K.S.; Cho, G.I.; Joo, G.J. The current status of the distribution of introduced fish in large river systems of South Korea. Int. Rev. Hydrobiol. 2002, 87, 319–328. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.K. Effect of the human utilization of northern snakehead (Channa argus Cantor, 1842) on the settlement of exotic fish and cladoceran community structure. Sustainability 2021, 13, 2486. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Tsukada, H.; Shiah, F.K. Latitudinal gradient in the ovary weight of bluegill Lepomis macrochirus in Lake Biwa, Japan. Ethol. Ecol. Evol. 2011, 23, 291–299. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Shiah, F.K. Spatial variation in reproductive measures of female bluegill Lepomis macrochirus Rafinesque in a small lake with diverse thermal conditions. Zool. Stud. 2013, 52, 19. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Kim, S.K.; Kim, J.C.; Kwon, S.J. Habitat preferences and trophic position of Brachydiplax chalybea flavovittata Ris, 1911 (Insecta: Odonata) larvae in Youngsan River wetlands of South Korea. Insects 2020, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Jeong, K.S.; Kim, H.W.; Chang, K.H.; Joo, G.J. Inter-annual variability of a zooplankton community: The importance of summer concentrated rainfall in a regulated river ecosystem. J. Ecol. Environ. 2011, 34, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Kim, S.K. Responses of rotifer community to microhabitat changes caused by summer-concentrated rainfall in a shallow reservoir, South Korea. Diversity 2020, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses; Springer: New York, NY, USA, 2000. [Google Scholar]
- Haney, J.F.; Hall, D.J. Sugar-coated Daphnia: A preservation technique for Cladocera. Limnol. Oceanogr. 1973, 18, 331–333. [Google Scholar] [CrossRef]
- Mizuno, T.; Takahashi, E. An Illustrated Guide to Freshwater Zooplankton in Japan; Tokai University Press: Tokyo, Japan, 1999; p. 532. [Google Scholar]
- Kim, I.S.; Park, J.Y. Freshwater Fish of Korea; Kyo Hak Publishing: Seoul, Korea, 2002. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. Community Ecology Package, Version 2. 2013. Available online: http://cran.r-project.org/web/packages/vegan/index.html (accessed on 23 March 2022).
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Fenger-Grøn, M.; Bramm, M.E.; Sandby, K.; Møller, P.H.; Rasmussen, H.U. Impact of fish predation on cladoceran body weight distribution and zooplankton grazing in lakes during winter. Freshw. Biol. 2004, 49, 432–447. [Google Scholar] [CrossRef]
- Ginter, K.; Blank, K.; Haberman, J.; Kangur, A.; Kangur, K. Fish predation pressure on zooplankton in a large northern temperate lake: Impact of adult predators versus juvenile predators. Proc. Est. Acad. Sci. 2018, 65, 356–367. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.K. A study of the distribution of Daphnia obtusa and Simocephalus vetulus in response to varying environmental conditions using field and microcosm approaches. Water 2020, 12, 815. [Google Scholar] [CrossRef]
- Graeb, B.D.; Dettmers, J.M.; Wahl, D.H.; Cáceres, C.E. Fish size and prey availability affect growth, survival, prey selection, and foraging behavior of larval yellow perch. Trans. Am. Fish. Soc. 2004, 133, 504–514. [Google Scholar] [CrossRef]
- Kim, S.K.; Joo, G.J.; Choi, J.Y. Effect of gated weir opening on the topography and zooplankton community of Geum River, South Korea. Land 2022, 11, 529. [Google Scholar] [CrossRef]
- Kim, H.W.; Choi, J.Y.; La, G.H.; Jeong, K.S.; Jo, G.J. Relationship between rainfall and zooplankton community dynamics in a riverine wetland ecosystem (Upo). Korean J. Ecol. Environ. 2010, 43, 129–135. [Google Scholar]
- Thomaz, S.M.; Cunha, E.R.D. The role of macrophytes in habitat structuring in aquatic ecosystems: Methods of measurement, causes and consequences on animal assemblages’ composition and biodiversity. Acta Limnol. Bras. 2010, 22, 218–236. [Google Scholar] [CrossRef]
- Kaufmann, P.R.; Hughes, R.M.; Van Sickle, J.; Whittier, T.R.; Seeliger, C.W.; Paulsen, S.G. Lakeshore and littoral physical habitat structure: A field survey method and its precision. Lake Reserv. Manag. 2014, 30, 157–176. [Google Scholar] [CrossRef]
- Son, S.H.; Kwon, S.J.; Im, J.H.; Kim, S.K.; Kong, D.; Choi, J.Y. Aquatic macrophytes determine the spatial distribution of invertebrates in a shallow reservoir. Water 2021, 13, 1455. [Google Scholar] [CrossRef]
- Rantala, M.J.; Ilmonen, J.; Koskimäki, J.; Suhonen, J.; Tynkkynen, K. The macrophyte, Stratiotes aloides, protects larvae of dragonfly Aeshna viridis against fish predation. Aquat. Ecol. 2004, 38, 77–82. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; Joo, G.J. Rainfall as dominant driver of rotifer dynamics in shallow wetlands: Evidence from a long-term data record (Upo wetlands, South Korea). Int. Rev. Hydrobiol. 2015, 100, 21–33. [Google Scholar] [CrossRef]
- Cheruvelil, K.S.; Soranno, P.A.; Madsen, J.D.; Roberson, M.J. Plant architecture and epiphytic macroinvertebrate communities: The role of an exotic dissected macrophyte. J. N. Am. Benthol. Soc. 2002, 21, 261–277. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; La, G.H.; Kim, S.K.; Joo, G.J. Sustainment of epiphytic microinvertebrate assemblage in relation with different aquatic plant microhabitats in freshwater wetlands (South Korea). J. Limnol. 2014, 73, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Kim, S.K.; Jeong, K.S.; Joo, G.J. Distribution pattern of epiphytic microcrustaceans in relation to different macrophyte microhabitats in a shallow wetland (Upo wetlands, South Korea). Oceanol. Hydrobiol. Stud. 2015, 44, 151–163. [Google Scholar] [CrossRef]
- Kemp, W.M.; Batleson, R.; Bergstrom, P.; Carter, V.; Gallegos, C.L.; Hunley, W.; Karrh, L.; Koch, E.W.; Landwehr, J.M.; Moore, K.A.; et al. Habitat requirements for submerged aquatic vegetation in Chesapeake Bay: Water quality, light regime, and physical-chemical factors. Estuaries 2004, 27, 363–377. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Z.; Xing, W.; Liu, G. Effects of substrate and shading on the growth of two submerged macrophytes. Hydrobiologia 2013, 700, 157–167. [Google Scholar] [CrossRef]
- Manatunge, J.; Asaeda, T.; Priyadarshana, T. The influence of structural complexity on fish–zooplankton interactions: A study using artificial submerged macrophytes. Environ. Biol. Fishes 2000, 58, 425–438. [Google Scholar] [CrossRef]
- Deosti, S.; de Fátima Bomfim, F.; Lansac-Tôha, F.M.; Quirino, B.A.; Bonecker, C.C.; Lansac-Tôha, F.A. Zooplankton taxonomic and functional structure is determined by macrophytes and fish predation in a Neotropical river. Hydrobiologia 2021, 848, 1475–1490. [Google Scholar] [CrossRef]
- Post, D.M. Individual variation in the timing of ontogenetic niche shifts in largemouth bass. Ecology 2003, 84, 1298–1310. [Google Scholar] [CrossRef]
- Jang, M.H.; Joo, G.J.; Lucas, M.C. Diet of introduced largemouth bass in Korean rivers and potential interactions with native fishes. Ecol. Freshw. Fish 2006, 15, 315–320. [Google Scholar] [CrossRef]
- Szedlmayer, S.T.; Lee, J.D. Diet shifts of juvenile red snapper (Lutjanus campechanus) with changes in habitat and fish size. Fish. Bull. 2004, 102, 366–375. [Google Scholar]
- Huss, M.; Byström, P.; Persson, L. Resource heterogeneity, diet shifts and intra-cohort competition: Effects on size divergence in YOY fish. Oecologia 2008, 158, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Holmes, T.H.; McCormick, M.I. Size-selectivity of predatory reef fish on juvenile prey. Mar. Ecol. Prog. Ser. 2010, 399, 273–283. [Google Scholar] [CrossRef]
- Dabrowski, K.; Portella, M.C. Feeding plasticity and nutritional physiology in tropical fishes. Fish. Physiol. 2005, 21, 155–224. [Google Scholar]
- Yamada, S.; Urabe, J. Role of sediment in determining the vulnerability of three littoral cladoceran species to odonate larvae predation. Inland Waters 2021, 11, 154–161. [Google Scholar] [CrossRef]
- Theel, H.J.; Dibble, E.D. An experimental simulation of an exotic aquatic macrophyte invasion and its influence on foraging behavior of bluegill. J. Freshw. Ecol. 2008, 23, 79–89. [Google Scholar] [CrossRef]
- Nolfi, S.; Floreano, D. Coevolving predator and prey robots: Do “arms races” arise in artificial evolution? Artif. life 1998, 4, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Boeing, W.J.; Wissel, B.; Ramcharan, C.W. Can costs and benefits of Daphnia defense against fish be measured in a field experiment? Fundam. Appl. Limnol. 2010, 176, 1. [Google Scholar] [CrossRef]
- Berge, J.; Cottier, F.; Varpe, Ø.; Renaud, P.E.; Falk-Petersen, S.; Kwasniewski, S.; Griffiths, C.; Søreide, J.E.; Johnsen, G.; Aubert, A.; et al. Arctic complexity: A case study on diel vertical migration of zooplankton. J. Plankton Res. 2014, 36, 1279–1297. [Google Scholar] [CrossRef]
- Kim, S.K.; Choi, J.Y. Differences in the vertical distribution of two cladoceran species in the Nakdong River estuary, South Korea. Water 2020, 12, 2154. [Google Scholar] [CrossRef]
- Iglesias, C.; Goyenola, G.; Mazzeo, N.; Meerhoff, M.; Rodo, E.; Jeppesen, E. Horizontal dynamics of zooplankton in subtropical Lake Bianca (Uruguay) hosting multiple zooplankton predators and aquatic plant refuges. In Shallow Lakes in a Changing World; Springer: Dordrecht, The Netherlands, 2007; pp. 179–189. [Google Scholar]
- Diel, P.; Kiene, M.; Martin-Creuzburg, D.; Laforsch, C. Knowing the enemy: Inducible defences in freshwater zooplankton. Diversity 2020, 12, 147. [Google Scholar] [CrossRef] [Green Version]
- Kuczyńska-Kippen, N.M.; Nagengast, B. The influence of the spatial structure of hydromacrophytes and differentiating habitat on the structure of rotifer and cladoceran communities. Hydrobiologia 2006, 559, 203–212. [Google Scholar] [CrossRef]
- Ali, M.M.; Mageed, A.A.; Heikal, M. Importance of aquatic macrophyte for invertebrate diversity in large subtropical reservoir. Limnologica 2007, 37, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Bucak, T.; SaraoĞLu, E.C.E.; Levi, E.E.; Nihan Tavşanoğlu, Ü.; Çakiroğlu, A.I.; Jeppesen, E.; Beklioğlu, M. The influence of water level on macrophyte growth and trophic interactions in eutrophic Mediterranean shallow lakes: A mesocosm experiment with and without fish. Freshw. Biol. 2012, 57, 1631–1642. [Google Scholar] [CrossRef]
- Van Onsem, S.; De Backer, S.; Triest, L. Microhabitat–zooplankton relationship in extensive macrophyte vegetations of eutrophic clear-water ponds. Hydrobiologia 2010, 656, 67–81. [Google Scholar] [CrossRef]
- Musil, M.; Novotná, K.; Potužák, J.; Hůda, J.; Pechar, L. Impact of topmouth gudgeon (Pseudorasbora parva) on production of common carp (Cyprinus carpio)-question of natural food structure. Biologia 2014, 69, 1757–1769. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Variable | Spring (n = 18) | Summer (n = 21) | Autumn (n = 19) | Winter (n = 21) |
|---|---|---|---|---|---|
| Upo | Dwm | 16.2 ± 4.8 | 31.5 ± 10.4 | 48.2 ± 11.6 | 5.1 ± 1.8 |
| WT | 16.3 ± 5.1 | 25.6 ± 2.4 | 16.1 ± 5.8 | 4.9 ± 1.7 | |
| DO | 101 ± 38 | 38 ± 27.8 | 34 ± 26.2 | 84 ± 31.8 | |
| Cond. | 408 ± 134 | 420 ± 152 | 361 ± 81 | 483 ± 131 | |
| pH | 7.8 ± 0.4 | 7.1 ± 0.5 | 7.2 ± 0.5 | 7.7 ± 0.3 | |
| Tur. | 36 ± 14.6 | 27 ± 14.8 | 56 ± 36.8 | 39 ± 25.6 | |
| Chl a | 13.8 ± 4.6 | 18 ± 5.4 | 18 ± 3.2 | 32 ± 2.6 | |
| Jangcheok | Dwm | 12.4 ± 3.7 | 34.8 ± 11.9 | 43.7 ± 14.2 | 8.2 ± 4.7 |
| WT | 16.7 ± 5.7 | 25.9 ± 1.6 | 16.9 ± 5.8 | 3.4 ± 2.6 | |
| DO | 96 ± 40.3 | 26 ± 11.6 | 35 ± 17.1 | 73 ± 37.4 | |
| Cond. | 341 ± 66.7 | 320 ± 243 | 275 ± 156 | 384 ± 246 | |
| pH | 7.6 ± 0.4 | 6.8 ± 0.4 | 6.9 ± 0.6 | 7.8 ± 0.4 | |
| Tur. | 26 ± 14.6 | 28 ± 20.6 | 31 ± 21.2 | 28 ± 13.4 | |
| Chl a | 25 ± 2.3 | 36 ± 4.1 | 31 ± 2.7 | 43 ± 4.1 | |
| Beongae | Dwm | 15.1 ± 7.3 | 37.4 ± 11.4 | 41.8 ± 13.7 | 8.0 ± 3.7 |
| WT | 17.3 ± 5.6 | 26.8 ± 1.8 | 18.2 ± 5.9 | 4.7 ± 2.7 | |
| DO | 97 ± 62.3 | 20 ± 31.2 | 23 ± 14.3 | 90 ± 43.2 | |
| Cond. | 289 ± 117 | 238 ± 118 | 269 ± 184 | 348 ± 216 | |
| pH | 7.4 ± 0.6 | 6.9 ± 0.4 | 7.3 ± 0.3 | 7.7 ± 0.4 | |
| Tur. | 29 ± 17.1 | 27 ± 14.8 | 40 ± 14.7 | 31 ± 24.7 | |
| Chl a | 14 ± 3.4 | 32 ± 3.5 | 27 ± 4.8 | 33 ± 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-K.; Choi, J.-Y. Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans. Water 2022, 14, 3781. https://doi.org/10.3390/w14223781
Kim S-K, Choi J-Y. Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans. Water. 2022; 14(22):3781. https://doi.org/10.3390/w14223781
Chicago/Turabian StyleKim, Seong-Ki, and Jong-Yun Choi. 2022. "Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans" Water 14, no. 22: 3781. https://doi.org/10.3390/w14223781
APA StyleKim, S.-K., & Choi, J.-Y. (2022). Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans. Water, 14(22), 3781. https://doi.org/10.3390/w14223781