
Phytoplankton Community Diversity and Its Environmental Driving Factors in the Northern South China Sea
Abstract
:1. Introduction
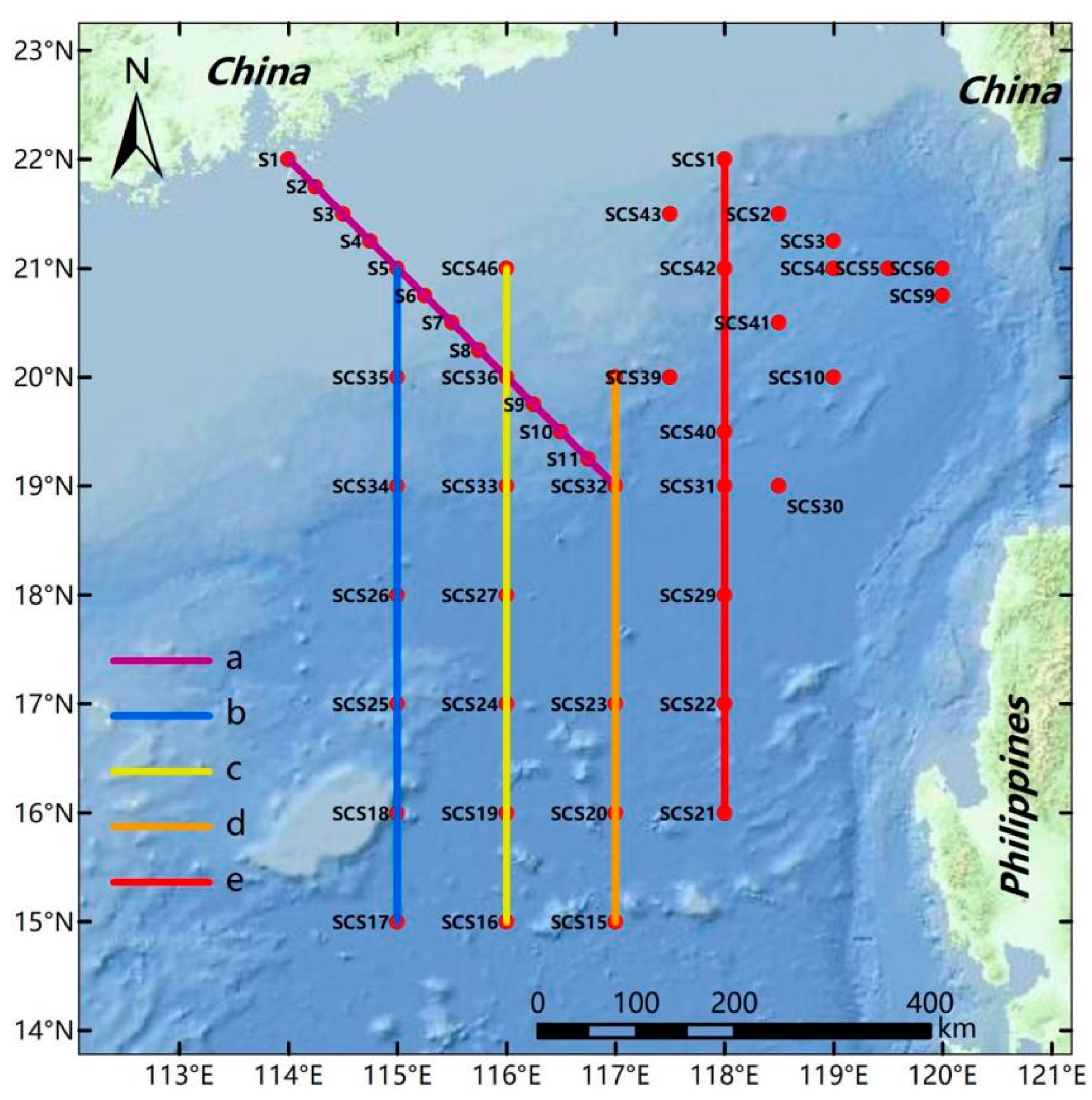
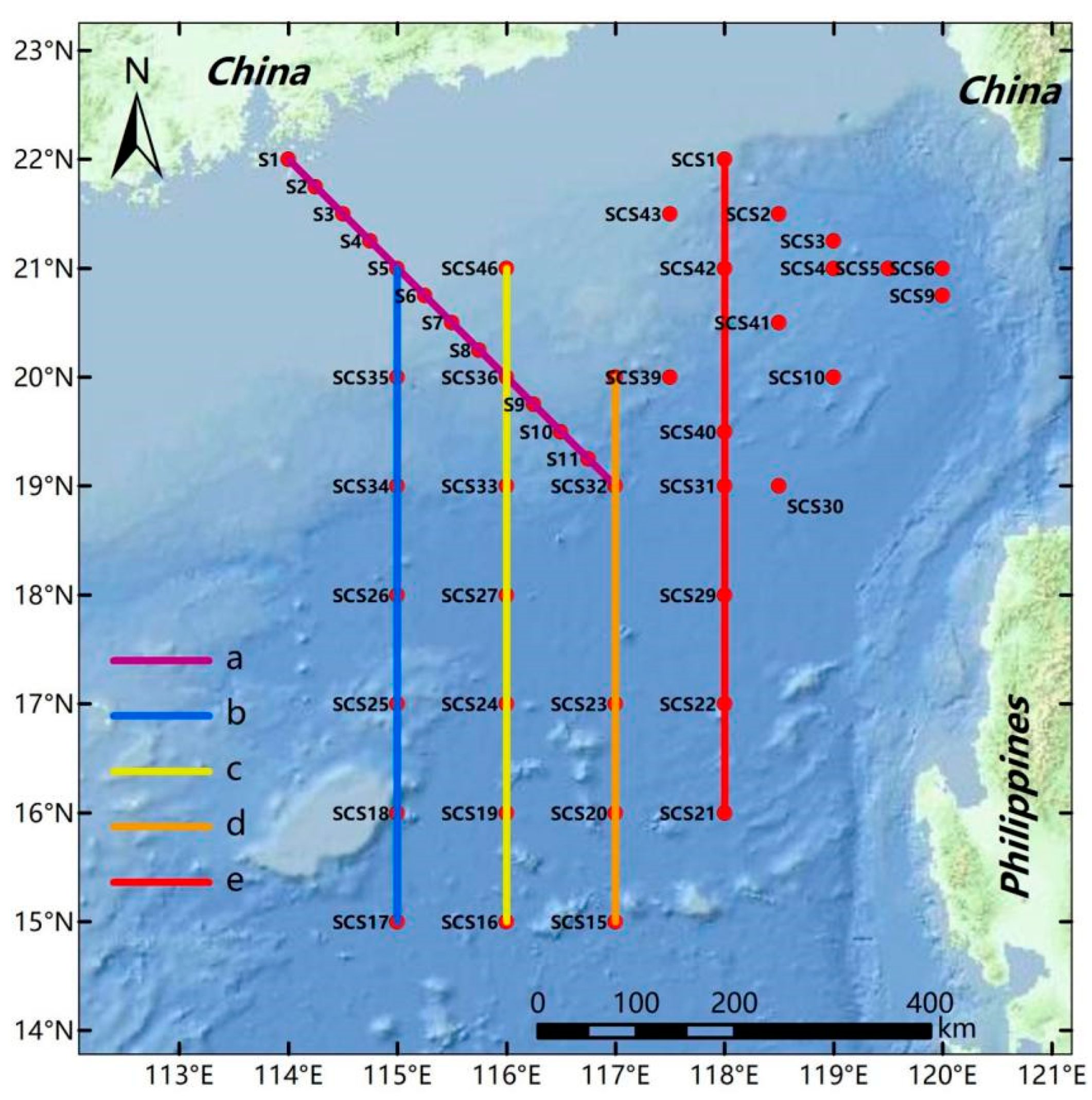
2. Materials and Methods
3. Results
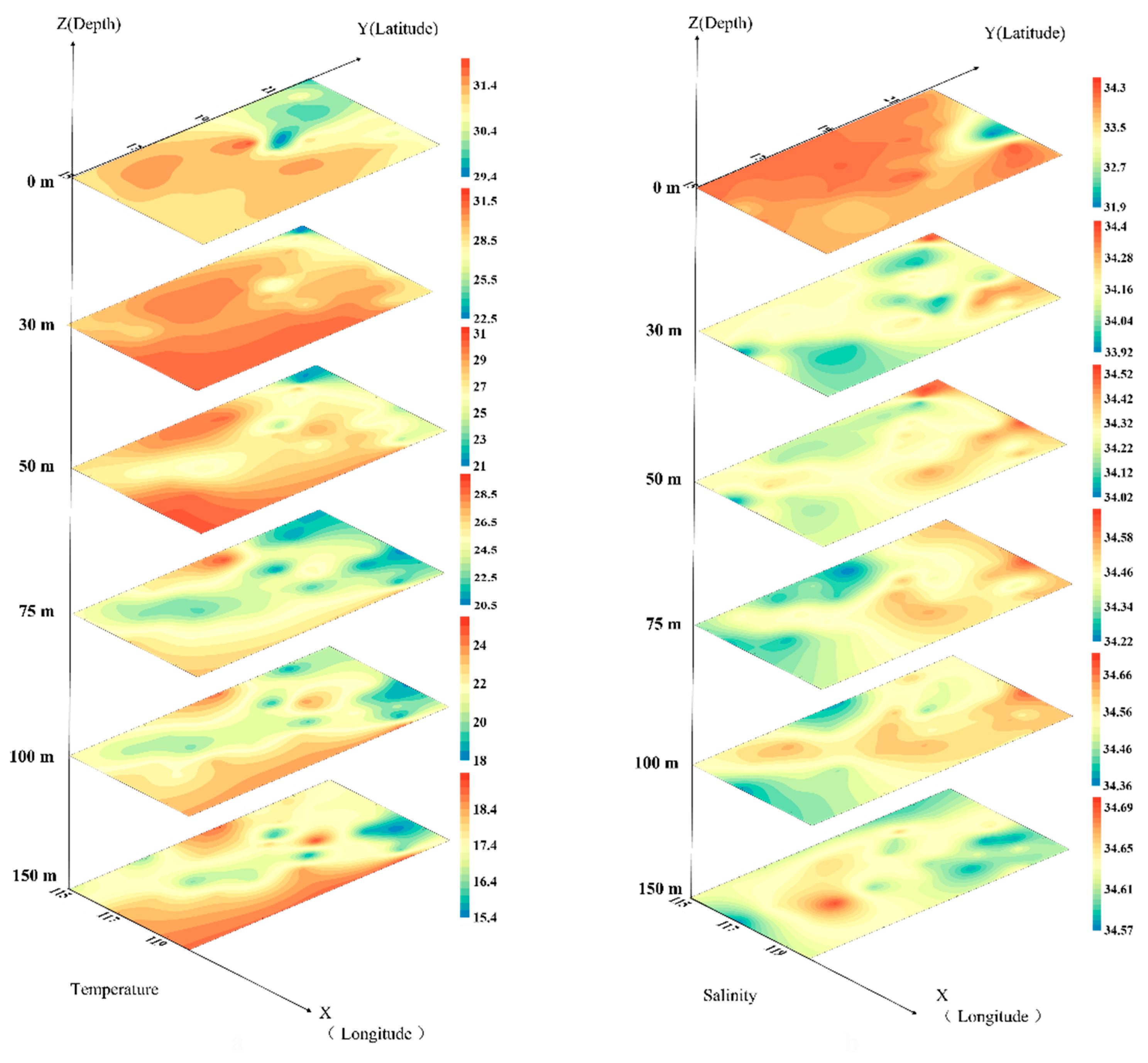
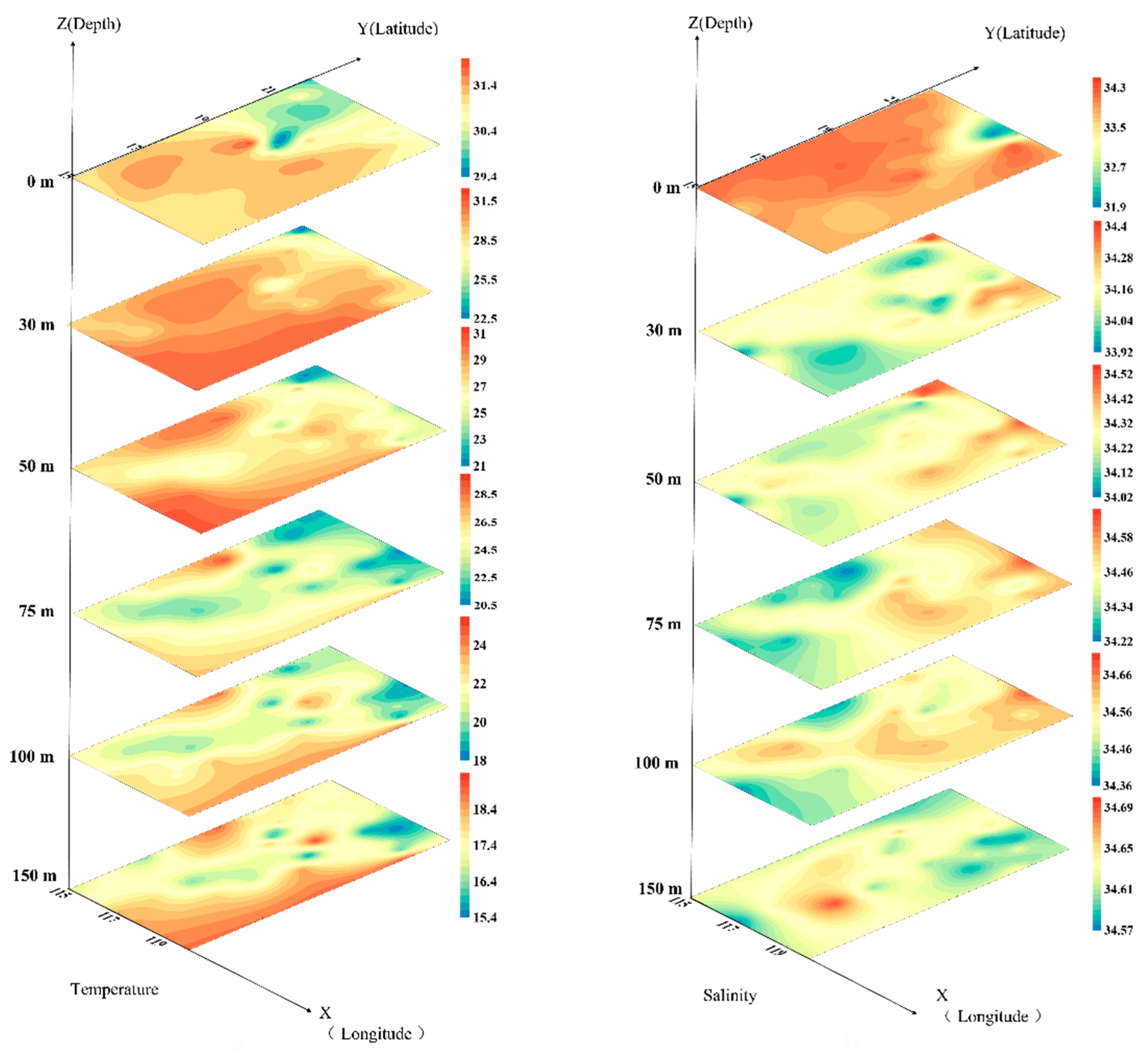
3.1. Environment of the Investigated Sea Area in the Northern South China Sea
3.2. Species Composition, Cell Abundance, and Ecological Type of Phytoplankton
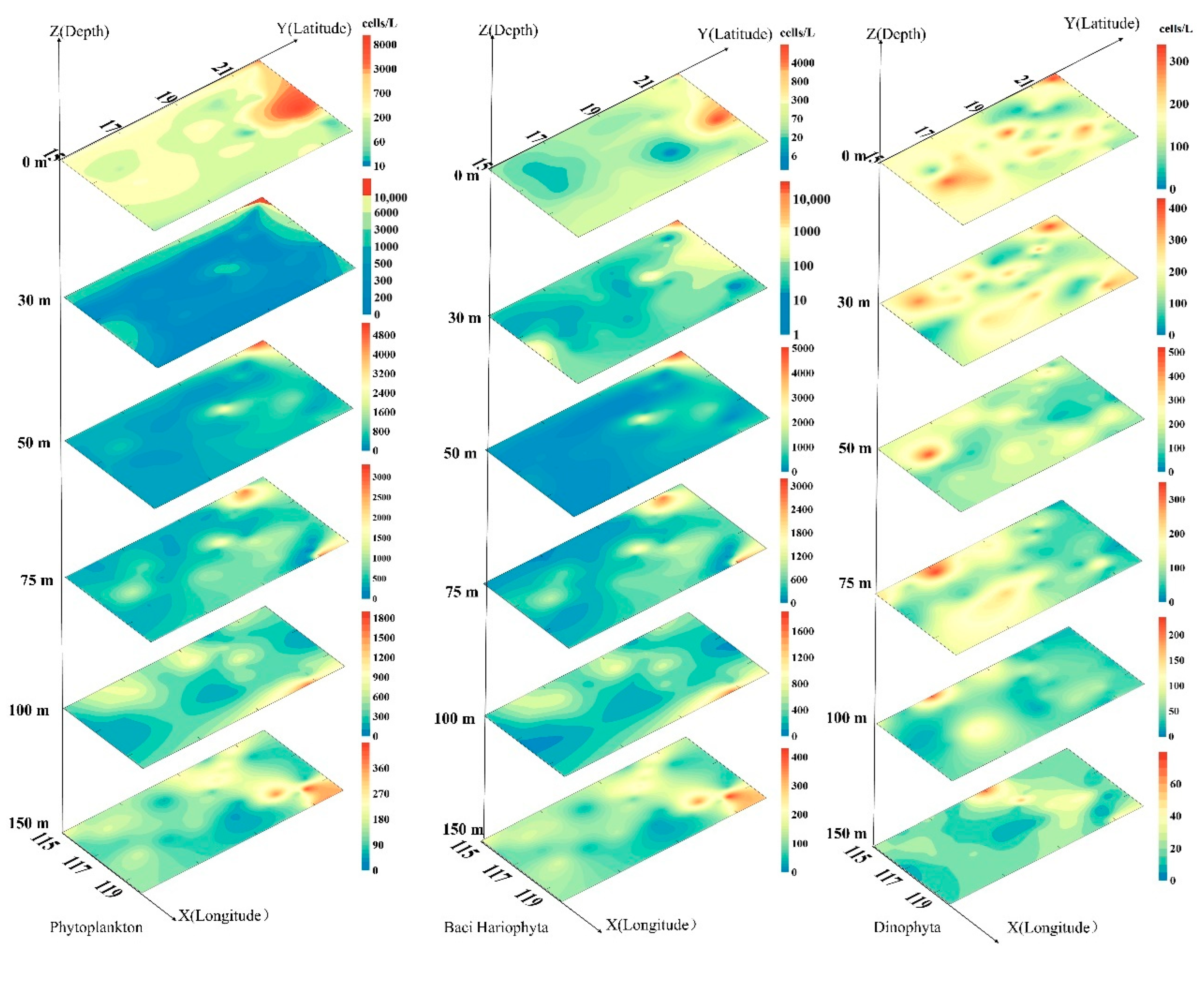
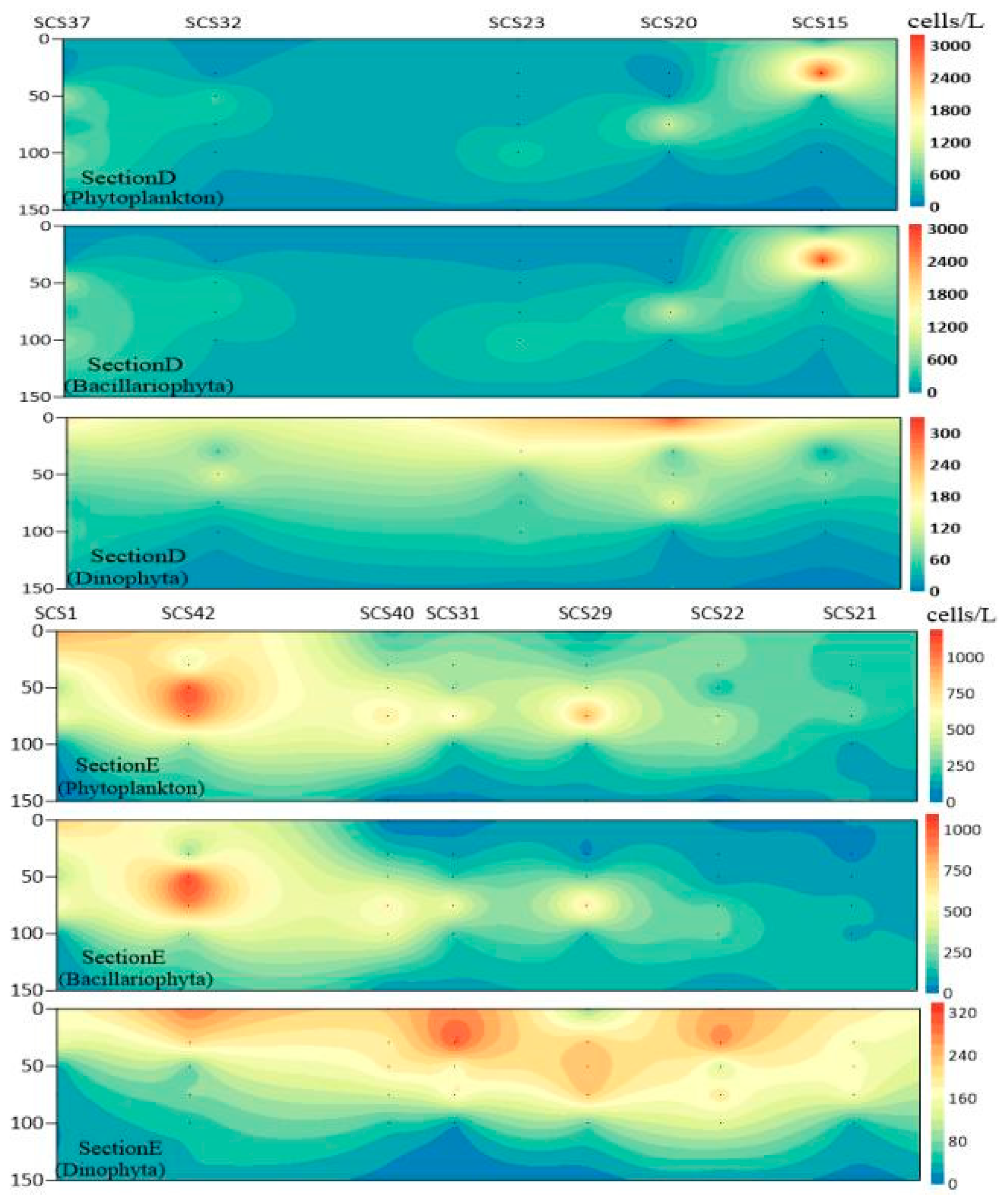
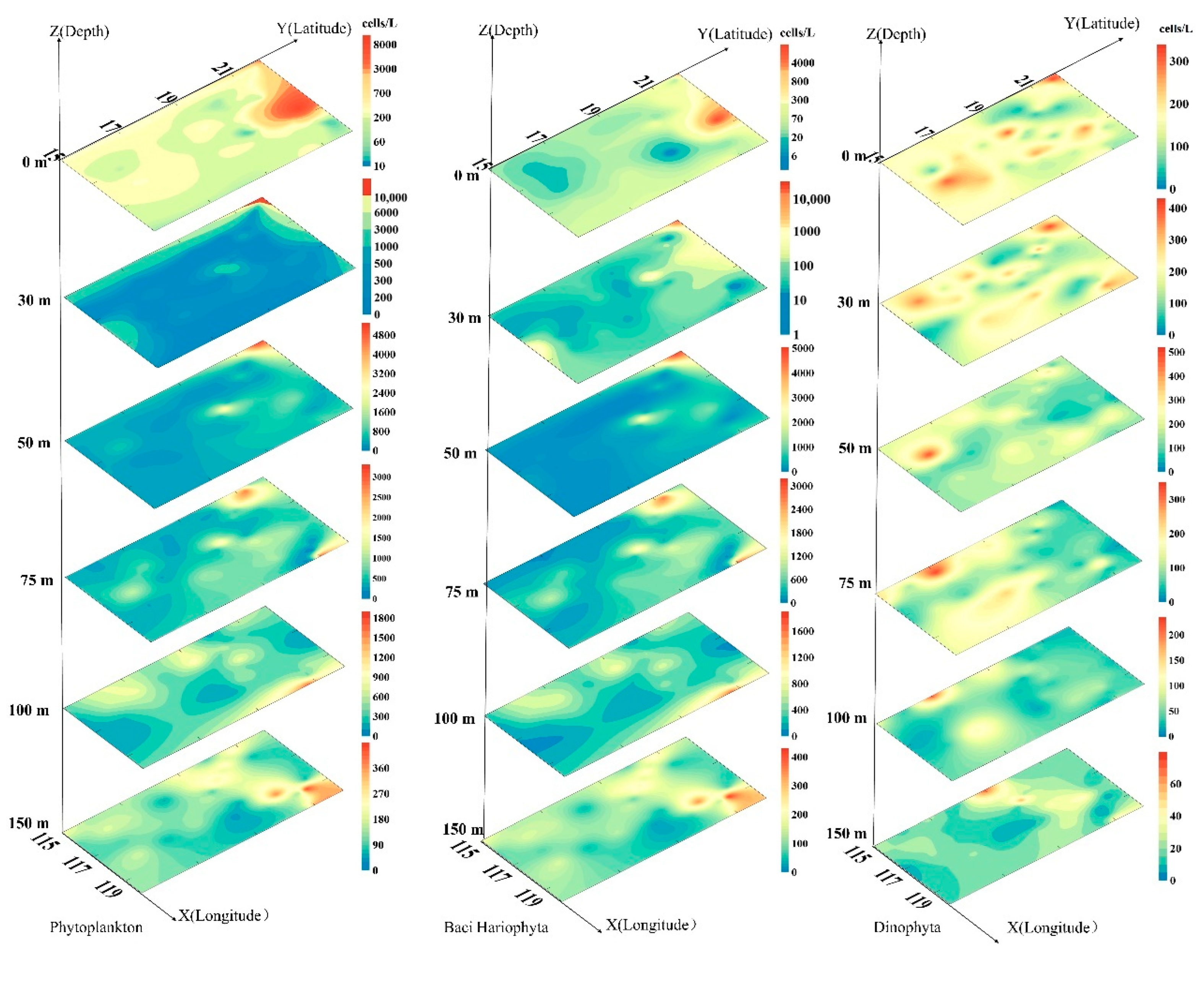
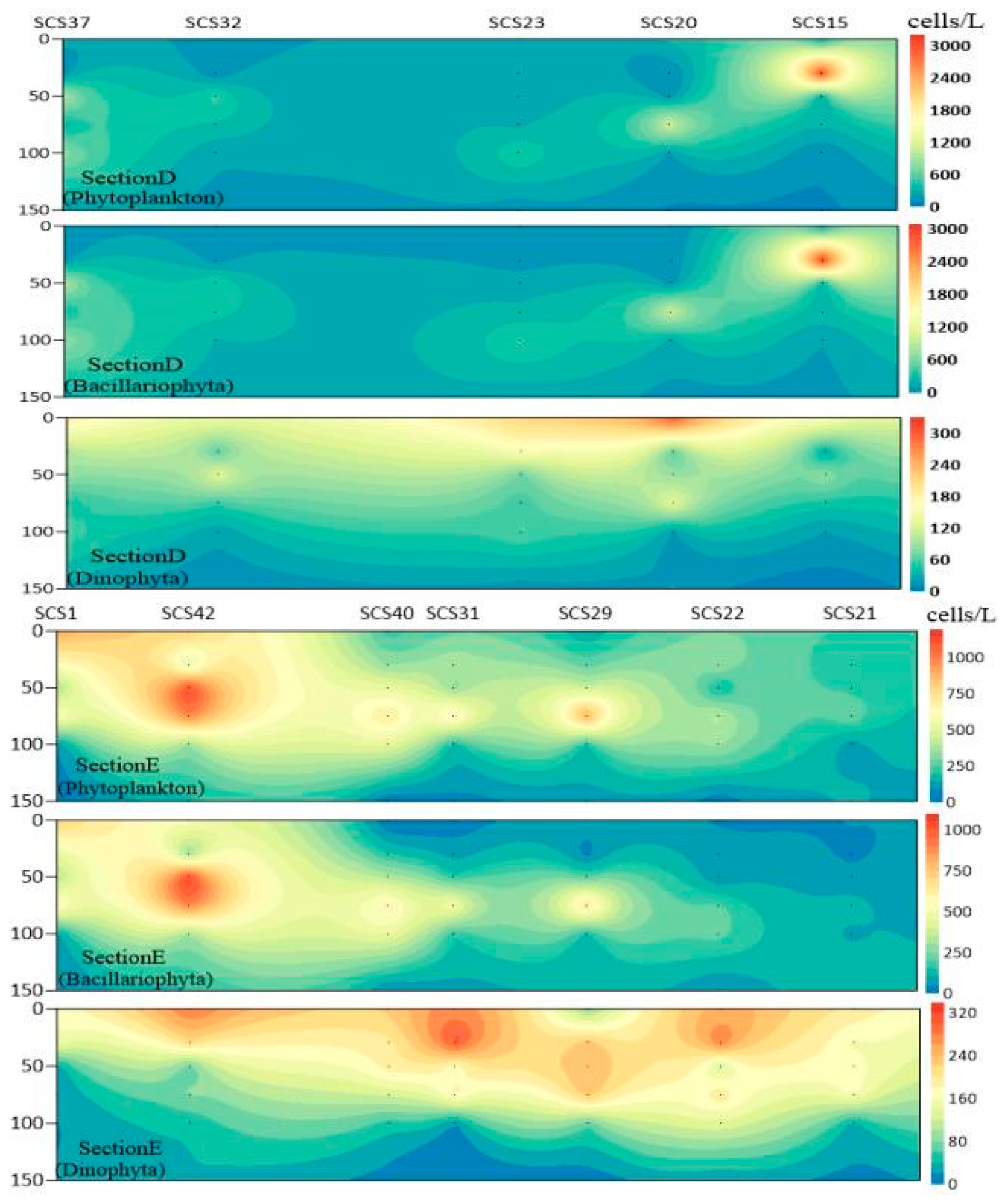
3.3. Plane Distribution of Phytoplankton
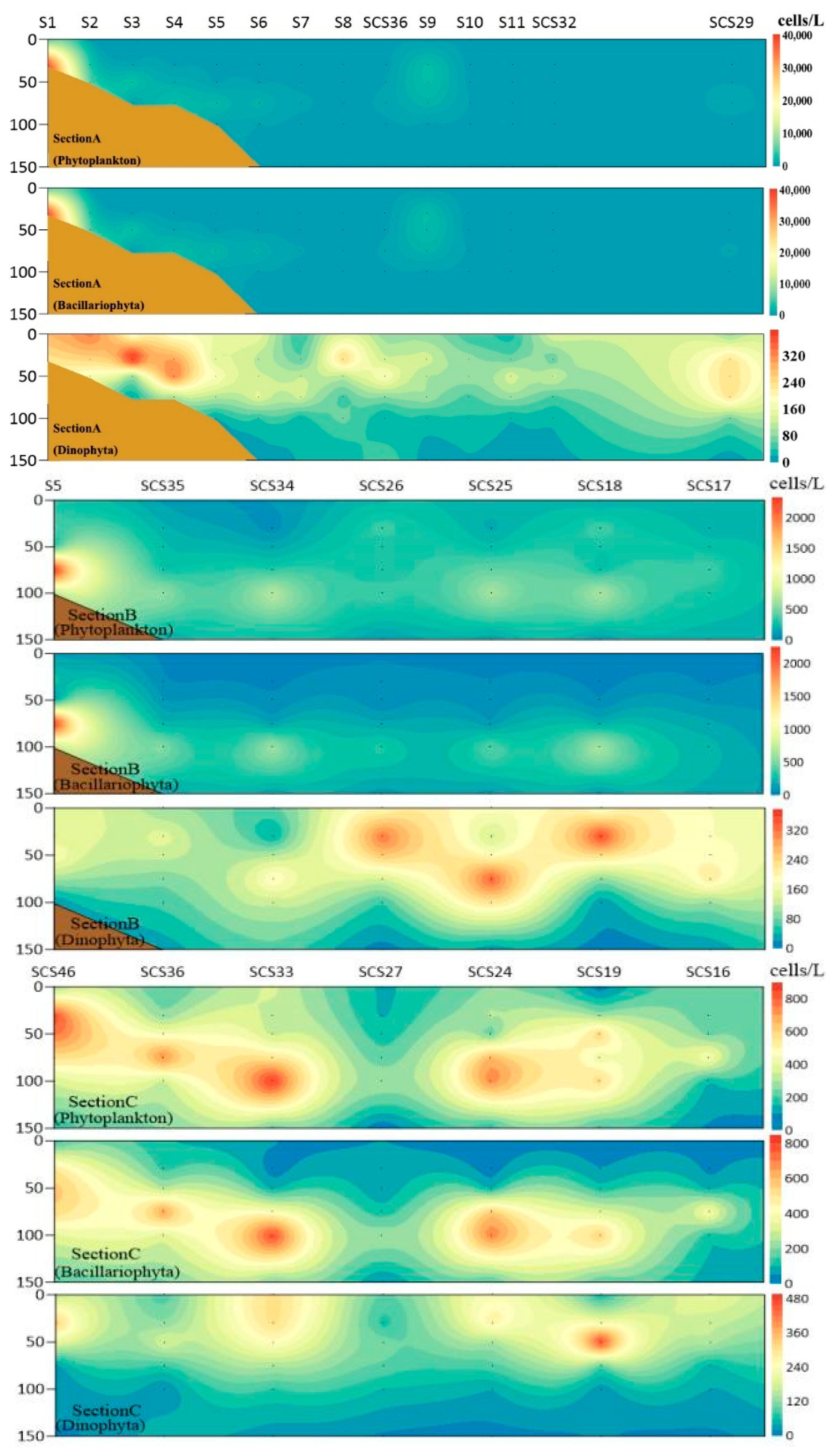
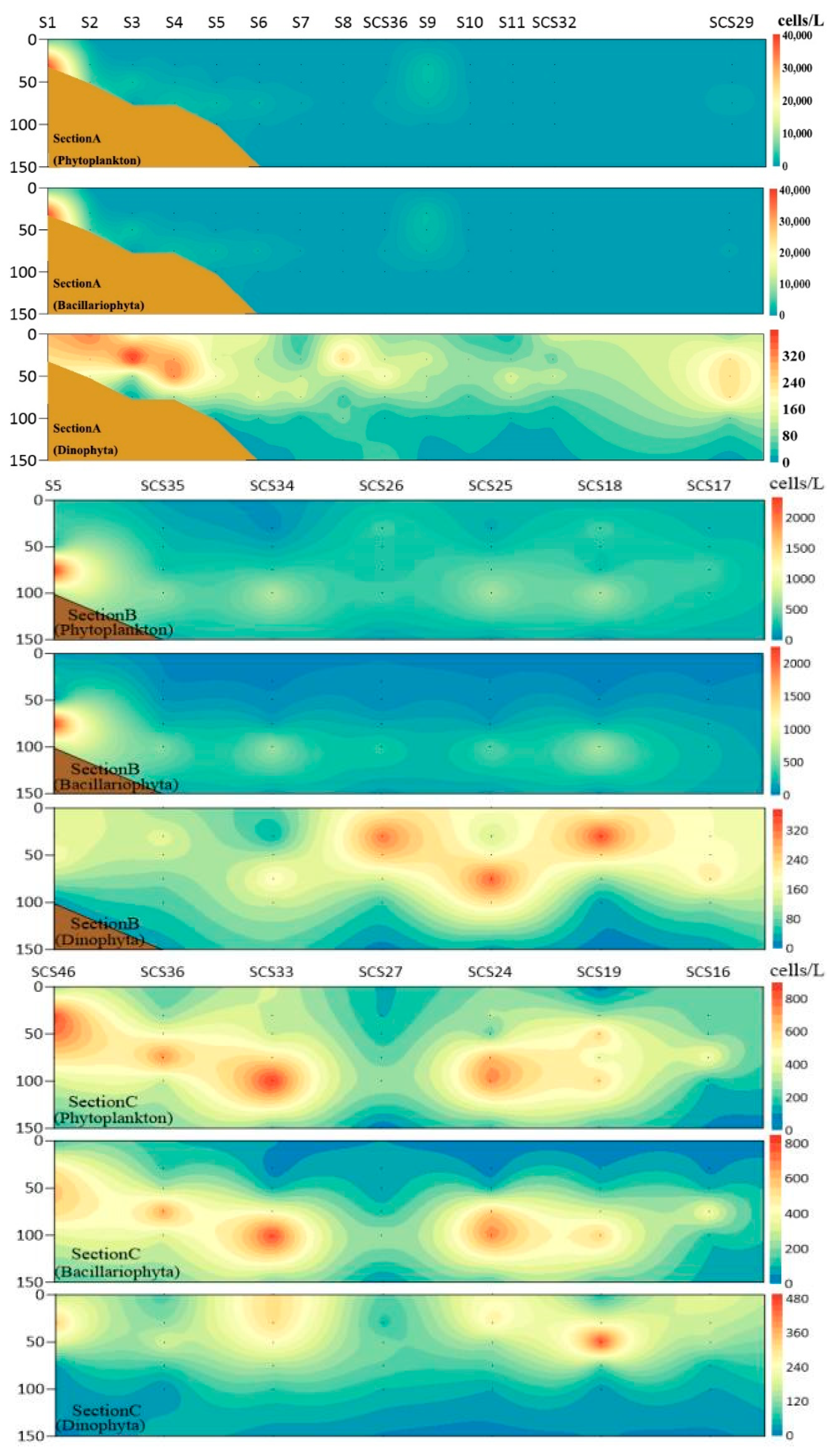
3.4. Vertical Distribution of Phytoplankton
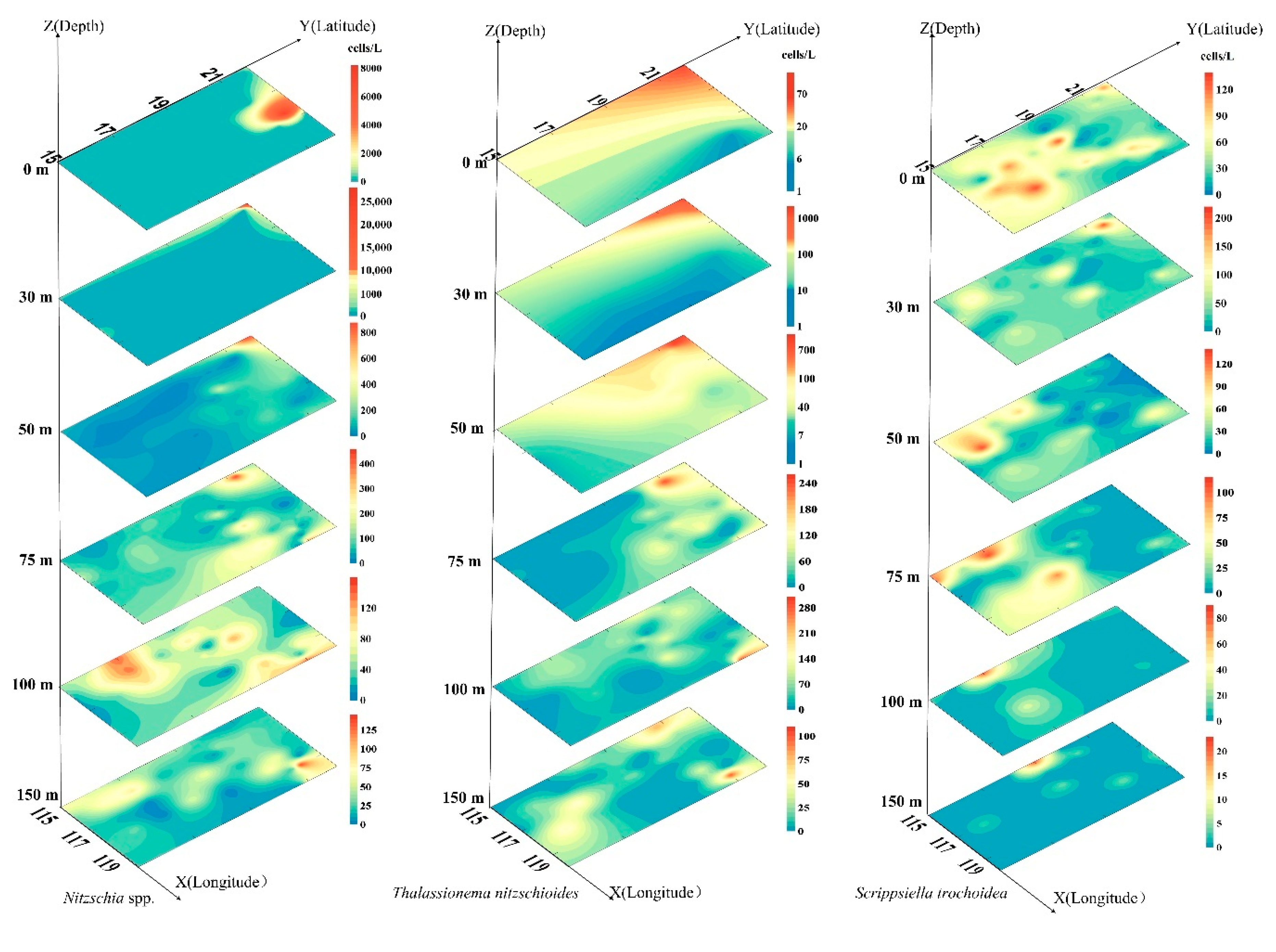
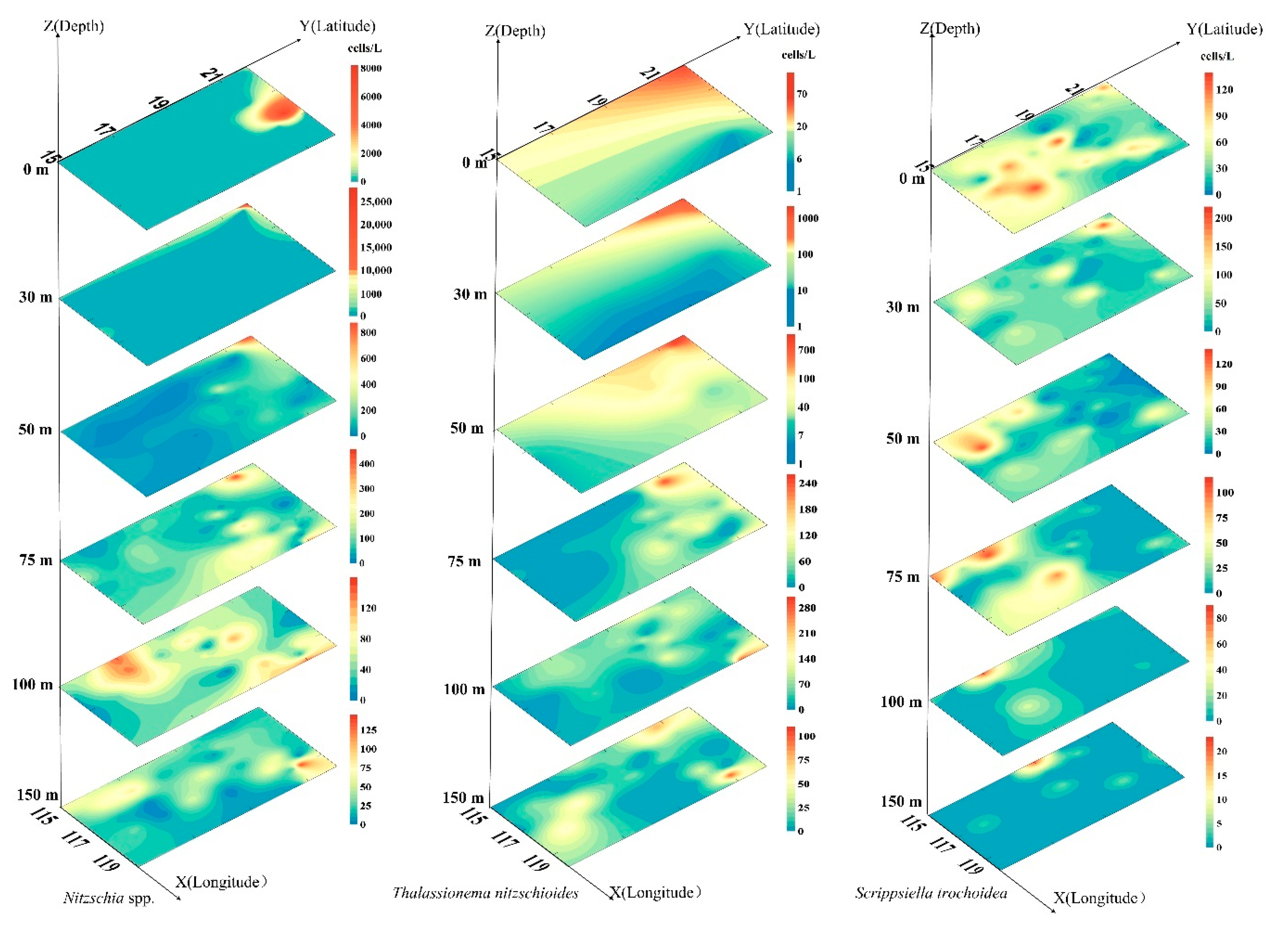
3.5. Dominant Species of Phytoplankton and Their Horizontal Distribution
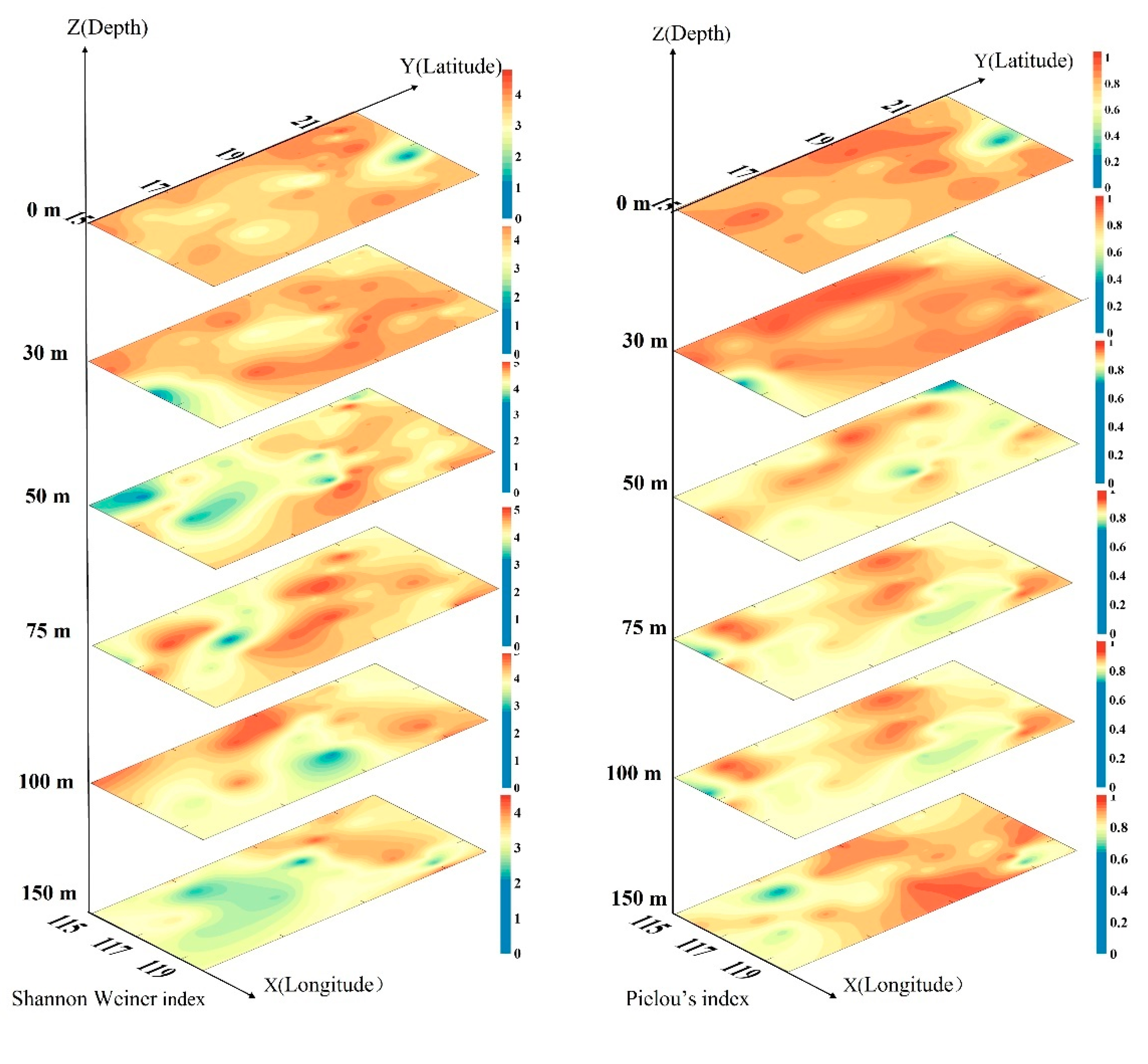
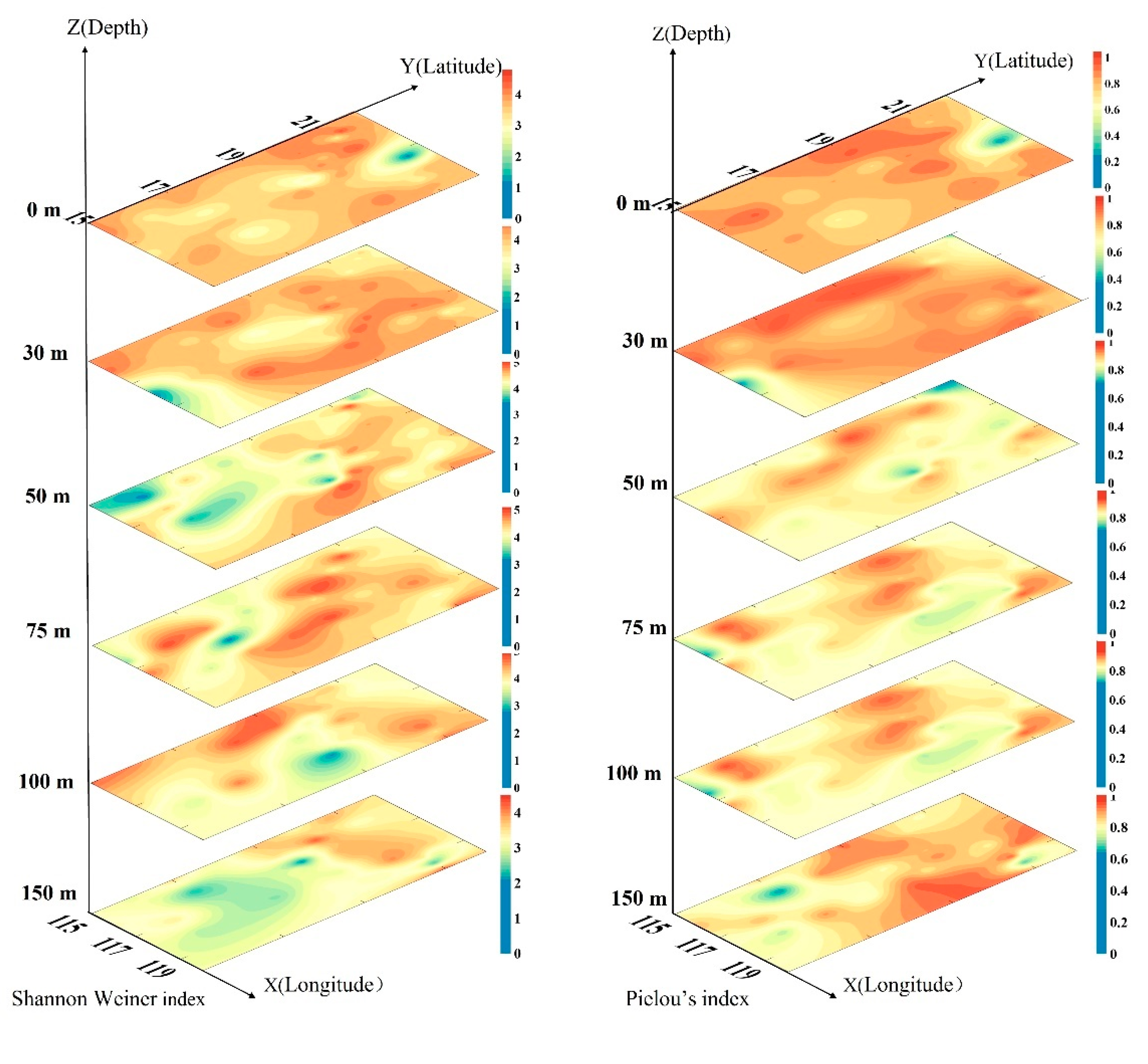
3.6. Diversity and Evenness Index of the Phytoplankton Community
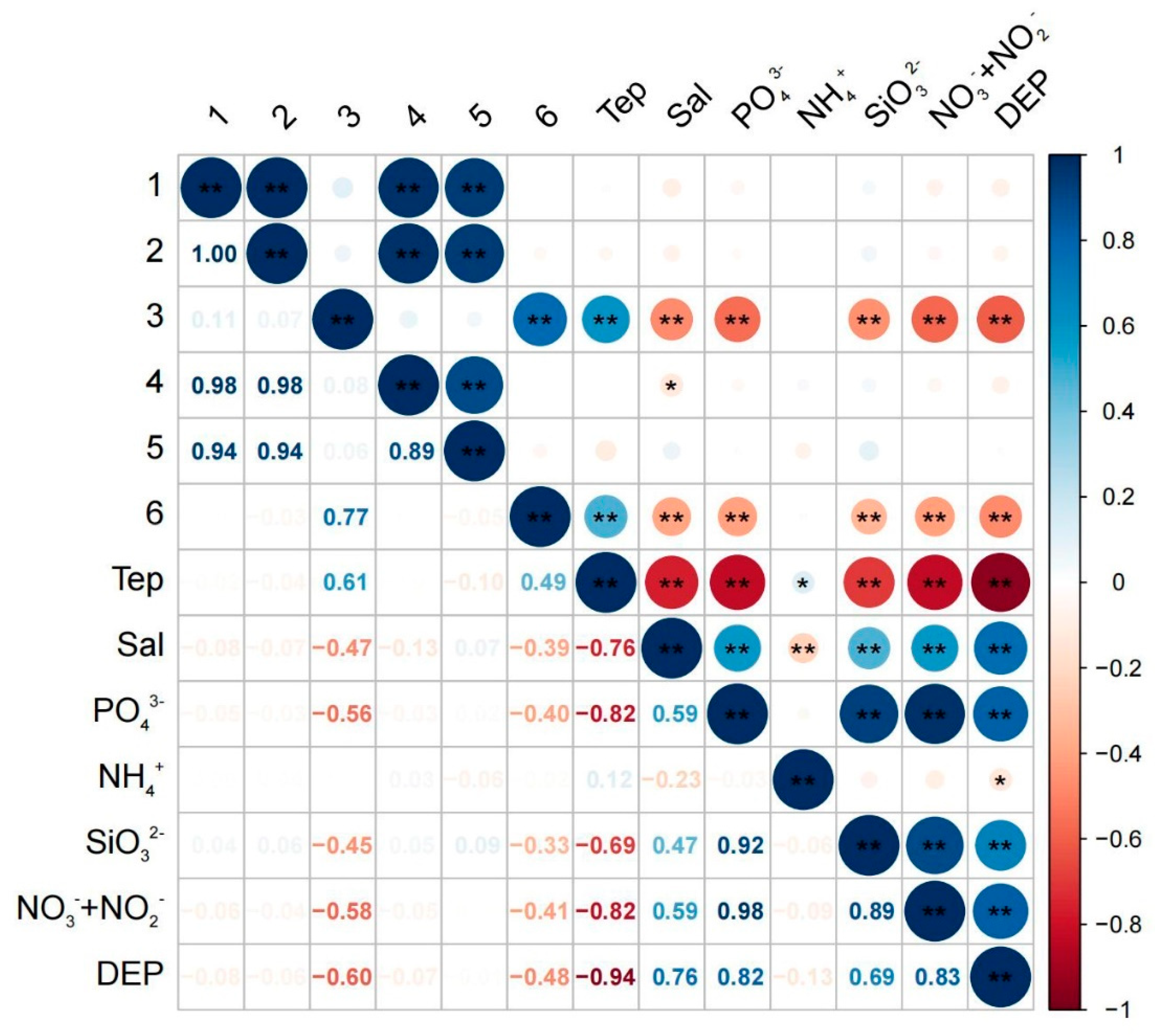
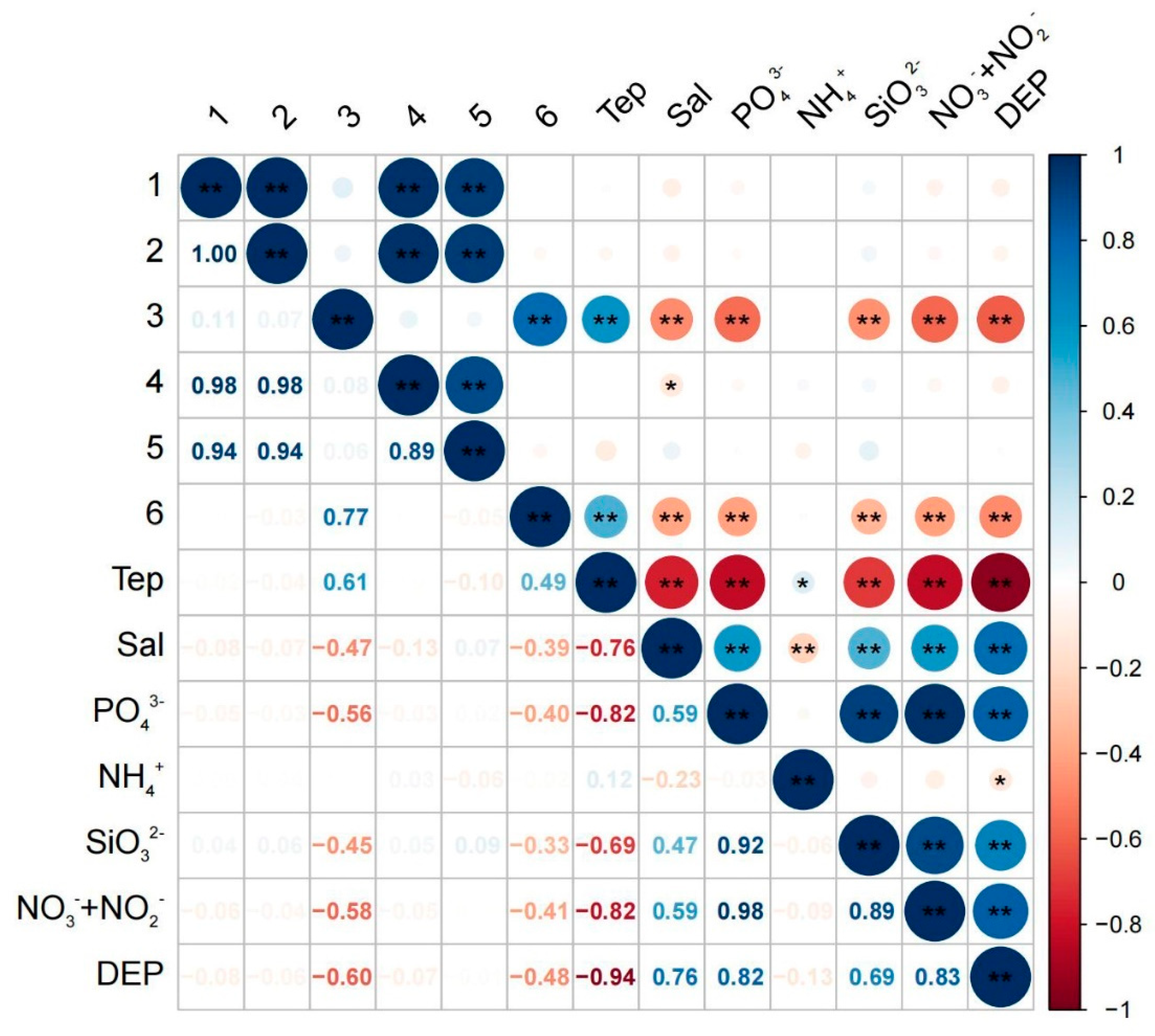
3.7. Correlation Results
3.8. Cluster Analysis of Community Structure
4. Discussion
4.1. Factors Affecting the Distribution of Phytoplankton
4.2. Comparison with Historical Data
4.3. Assessment of Community Structure Stability and Eutrophication in the Northern SCS
4.4. Correlation between Phytoplankton Community Structure and Environmental Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacillariophyta | Chaetoceros distans Cleve |
| Achnanthes brevipes Agardh | Chaetoceros femur Schütt |
| Actinocyclus octonarius Ehrenberg | Chaetoceros hirundinellus Qian |
| Actinocyclus octonarius Ehrenberg | Chaetoceros imbricatus Mangin |
| Actinoptychus hexagonus Grunow in Schmidt | Chaetoceros indicus Karsten |
| Actinoptychus senarius (Ehr.) Ehrenberg | Chaetoceros knipowitschii Henckel |
| Asterolampra marylandica Ehrenberg | Chaetoceros lauderi Ralfs |
| Asterolampra vanheurckii Brun | Chaetoceros messanensis Castracane |
| Asteromphalus cleveanus Grunow | Chaetoceros pelagicus Cleve |
| Asteromphalus elegans Greville | Chaetoceros pendulus Karsten |
| Asteromphalus flabellatus (Brébisson) Greville | Chaetoceros peruvianus Brightwell |
| Asteromphalus heptactis (Breb.) Ralfs | Chaetoceros pseudodichaeta Ikari |
| Asteromphalus rubustus Castracane | Chaetoceros rostratus Lauder |
| Asteromphalus spp. | Chaetoceros rostratus var. glandazi Mangin |
| Bacillaria paxillifera (Müller) Hendey | Chaetoceros spp. |
| Bacteriastrum comosum Pavillard | Chaetoceros teres Cleve |
| Bacteriastrum elongatum Cleve | Chaetoceros tetrastichon Cleve |
| Bacteriastrum furcatum Shadbolt | Chaetoceros tortissimus Gran |
| Bacteriastrum hyalinum Lauder | Climacodium frauenfeldianum Grunow |
| Bacteriastrum mediterraneum Pavillard | Corethron criophilum Castracane |
| Biddulphia sinensis Greville | Coscinodiscus debilis Grove |
| Campylosira cymbelliformis (Schmidt) Grunow ex Van Heurck | Coscinodiscus gigas Ehrenberg |
| Cerataulina bergonii Ostenfeld | Coscinodiscus granii Grough |
| Chaetoceros aequatoriale Cleve | Coscinodiscus jonesianus (Greville) Ostenfeld |
| Chaetoceros affinis Lauder | Coscinodiscus nobilis Grunow |
| Chaetoceros atlanticus Cleve | Coscinodiscus oculus-iridis Ehrenberg |
| Chaetoceros atlanticus var. neapolitana (Schröder) Hustedt | Coscinodiscus spp. |
| Chaetoceros atlanticus var. skeleton (Schütt) Hustedt | Coscinodiscus subtilis Ehrenberg |
| Chaetoceros aurivillii Cleve | Cyclotella striata (Kuetz.) Grunow |
| Chaetoceros bacteriastroides Karsten | Chaetoceros diadema (Ehrenberg) Gran |
| Chaetoceros buceros Karsten | Diploneis bombus Ehrenberg |
| Chaetoceros castracanei Karsten | Ditylum brightwellii (West) Grunow |
| Chaetoceros coarctatus Lauder | Ditylum sol Grunow |
| Chaetoceros compressus Lauder | Donkinia sp. |
| Chaetoceros constrictus Gran | Eucampia cornuta (Cleve) Grunow |
| Chaetoceros dadayi Pavillard | Eucampia zodiacus Ehrenberg |
| Chaetoceros danicus Cleve | Eunotogramma debile Grunow in Van Heurck |
| Chaetoceros debilis Cleve | Fragilaria spp. |
| Chaetoceros decipiens f. singularis Gran | Fragilariopsis doliolus (Wallich) Medlin & Sims |
| Chaetoceros densus (Cleve) Cleve | Gossleriella tropica Schütt |
| Chaetoceros denticulatus f. angusta Hustedt ex Simonsen | Rhizosolenia styliformis var. latissima Brightwell |
| Guinardia delicatula (Cleve) Hasle et al. | Schröderella delicatula f. schröderi (Bergon) Sournia |
| Guinardia striata (Stolterfoth) Hasle et al. | Skeletonema costatum (Greville) Cleve |
| Gyrosigma balticum (Ehrenberg) Cleve | Stephanopyxis turris (Greville) Ralfs |
| Helicotheca tamesis (Shrubsole) Ricard | Streptotheca indica Karsten |
| Hemiaulus hauckii Grunow ex Van Heurck | Synedra spp. |
| Hemiaulus sinensis Greville | Thalassionema frauenfeldii (Grunow) Hallegraeff |
| Hemidiscus cuneiformis var. cuneiformis Wallich | Thalassionema nitzschioides Grunow |
| Lauderia annulata Cleve | Thalassiosira leptopus (Grunow ex Van Heurck) Hasle & G. Fryxell |
| Lauderia mediterranea Peragallo | Thalassiosira nordenskiöldii Cleve |
| Lauderia pumila Castracane | Thalassiosira rotula Meunier |
| Leptocylindrus danicus Cleve | Thalassiosira spp. |
| Mastogloia rostrata (Wallich) Hustedt | Thalassiothrix longissima Cleve et Grunow |
| Meuniera membranacea (Cleve) Silva | Triceratium affine Grunow |
| Navicula spp. | Dinophyta |
| Nitzschia longissima (Brébisson) Ralfs | Akashiwo sanguinea (Hirasaka) Hansen & Moestrup |
| Nitzschia lorenziana Grunow | Alexandrium cohorticula (Balech) Balech |
| Nitzschia spp. | Alexandrium spp. |
| Odontella longicruris (Greville) Hoban | Alexandrium tamiyavanichii Balech |
| Odontella mobiliensis (Bailey) Grunow | Amphidoma nucula Stein |
| Odontella regia (Schultze) Simonsen | Amphisolenia bidentata Schröder |
| Odontella spp. | Amphisolenia brevicauda Kofoid |
| Paralia sulcata (Ehrenberg) Cleve | Amphisolenia globifera Stein |
| Pinnularia spp. | Amphisolenia inflata Murray & Whitting |
| Planktoniella blanda Syvertsen & Hasle | Amphisolenia thrinax Schütt |
| Planktoniella formosa Qian & Wang | Amylax triacantha (Jörgensen) Sournia |
| Pleurosigma acutum Norman | Blepharocysta splendor-maris (Ehrenberg) Ehrenberg |
| Proboscia alata (Brightwell) Sundströn | Ceratium declinatum var. angusticornum Peters |
| Pseudo-nitzschia delicatissima (Cleve) Heiden et al. | Ceratocorys horrida Stein |
| Pseudo-nitzschia pungens (Grunow ex Cleve) Hasle | Ceratocorys magna Kofoid |
| Pseudosolenia calcar-avis (Schultze) Sundström | Citharistes regius Stein |
| Rhizosolenia alata f. indica (Peragallo) Ostenfeld | Cladopyxis brachiolata Stein |
| Rhizosolenia bergonii Peragallo | Corythodinium belgicae (Meunier) F.J.R. Taylor |
| Rhizosolenia castracanei Peragallo | Corythodinium carinatum (Gaarder) F.J.R. Taylor |
| Rhizosolenia clevei Ostenfeld | Corythodinium compressum (Kofoid) Taylor |
| Rhizosolenia cochlea Brun | Corythodinium constrictum (Stein) Taylor |
| Rhizosolenia gracillima Cleve | Corythodinium curvicaudatum (Kofoid) F.J.R. Taylor |
| Rhizosolenia hyalina Ostenfeled et Schmidt | Corythodinium elegans (Pavillard) Taylor |
| Rhizosolenia imbricata Brightwell | Corythodinium frenguellii (Rampi) Taylor |
| Rhizosolenia robusta Norman ex Ralfs | Corythodinium tesselatum (Stein) Loeblich Jr. & Loeblich III |
| Rhizosolenia semispina Hensen | Dinophysis acuminata Claparède et Lachmann |
| Rhizosolenia setigera Brightwell | Dinophysis argus (Stein) Abé |
| Rhizosolenia sinensis Qian | Dinophysis caudata Saville-Kent |
| Rhizosolenia styliformis Brightwell | Dinophysis cuneus (Schütt) Abé |
| Dinophysis doryphorum (Stein) Abé | Histioneis costata Kofoid & Michener |
| Dinophysis expulsa Kofoid et Michener | Histioneis cymbalaria Stein |
| Dinophysis fortii Pavillard | Histioneis para Murray & Whitting |
| Dinophysis laevis Claparède & Lachmann | Histioneis parallela Gaarder |
| Dinophysis mitra (Schütt) Abé | Histioneis pulchra Kofoid |
| Dinophysis ovata Claparède & Lachmann | Histioneis biremis Stein |
| Dinophysis oviformis Chen & Ni | Histioneis cleaveri Rampi |
| Dinophysis parvula (Schütt) Balech | Karenia spp. |
| Dinophysis porodictyum (Stein) Abé | Lingulodinium polyedrum (Stein) Dodge |
| Dinophysis rapa (Stein) Abé | Neoceratium arietinum (Cleve) Gómez, Moreira & López-Garcia |
| Dinophysis rotundata Claparède & Lachmann | Neoceratium axiale (Kofoid) Gómez, Moreira & López-Garcia |
| Dinophysis schuettii Murray & Whitting | Neoceratium belone (Cleve) Gómez, Moreira & López-Garcia |
| Dinophysis spp. | Neoceratium biceps (Claparède & Lachmann) Gómez, Moreira & López-Garcia |
| Dinophysis tailisuni Chen & Ni | Neoceratium boehmii (Graham et Bronikovsky) |
| Dinophysis uracantha Stein | Neoceratium carriense (Gourret) Gómez, Moreira & López-Garcia |
| Diplopsalopsis bomba (Stein ex Jorgensen) Dodge & Toriumi | Neoceratium deflexum (Kofoid) Gómez, Moreira & López-Garcia |
| Diplopsalopsis globula Abé | Neoceratium ehrenbergii (Kofoid) |
| Dissodinium lunula Schütt | Neoceratium extensum (Gourret) Gómez, Moreira & López-Garcia |
| Dolichodinium lineatum (Kofoid & Michener) Kofoid & Adamson | Neoceratium falcatum (Kofoid) Gómez, Moreira & López-Garcia |
| Goniodoma polyedricum (Pouchet) Jörgensen | Neoceratium furca (Ehrenberg) Gómez, Moreira & López-Garcia |
| Goniodoma sphaericum Murray & Whitting | Neoceratium furca var. nannofurca (Jörgensen) Yang, Li & Dong |
| Gonyaulax birostris Stein | Neoceratium fusus (Ehrenberg) Gómez, Moreira & López-Garcia |
| Gonyaulax brevisulcata Dangeard | Neoceratium gravidum (Gourret) Gómez, Moreira & López-Garcia |
| Gonyaulax digitale (Pouchet) Kofoid | Neoceratium hexacanthum (Gourret) Gómez, Moreira & López-Garcia |
| Gonyaulax fusiformis Graham | Neoceratium hircus (Schröder) Gómez, Moreira & López-Garcia |
| Gonyaulax hyalina Ostenfeld & Schmidt | Neoceratium horridum (Gran) Gómez, Moreira & López-Garcia |
| Gonyaulax kofoidii Pavillard | Neoceratium karstenii (Pavillard) Gómez, Moreira & López-Garcia |
| Gonyaulax milneri (Murray & Whitting) Kofoid | Neoceratium lineatum (Ehrenberg) Gómez, Moreira & López-Garcia |
| Gonyaulax minuta Kofoid & Michener | Neoceratium macroceros (Ehrenberg) Gómez, Moreira & López-Garcia |
| Gonyaulax monacantha Pavillard | Neoceratium macroceros var. gallicum (Kofoid) Yang, Li & Dong |
| Gonyaulax monospina Rampi | Neoceratium massiliense (Gourret) Gómez, Moreira & López-Garcia |
| Gonyaulax pacifica Kofoid | Neoceratium minutum (Jørgensen) Gómez, Moreira & López-Garcia |
| Gonyaulax polygramma Stein | Neoceratium pentagonum (Gourret) Gómez, Moreira & López-Garcia |
| Gonyaulax sphaeroidea Kofoid | Neoceratium praeolongum (Lemmermann) Gómez, Moreira & López-Garcia |
| Gonyaulax subulata Kofoid & Michener | Neoceratium ranipes (Cleve) Gómez, Moreira & López-Garcia |
| Gonyaulax turbynei Murray & Whitting | Neoceratium seta (Ehrenberg) Yang & Li |
| Gonyaulax verior Sournia | Neoceratium sumatranum (Karsten) Yang & Li |
| Gymnodinium spp. | Neoceratium tenue (Ostenfeld & Schmidt) Gómez, Moreira & López-Garcia |
| Gyrodinium dominans Hulbert | Neoceratium teres (Kofoid) Gómez, Moreira & López-Garcia |
| Gyrodinium fusiform | Neoceratium trichoceros (Ehrenberg) Gómez, Moreira & López-Garcia |
| Gyrodinium spirale (Bergh) Kofoid et Swezy | Neoceratium tripos (O.F. Müller) Gómez, Moreira & López-Garcia |
| Histioneis depressa Schiller | Noctiluca scintillans (Macartney) Ehrenberg |
| Histioneis gregoryi Böhm | Ornithocercus heteroporus Kofoid |
| Histioneis highleyi Murray & Whitting | Ornithocercus magnificus Stein |
| Histioneis oxypteris Schiller | Ornithocercus quadratus v. quadratus Schütt |
| Histioneis panda Kofoid & Michener | Ornithocercus skogsbergii Abé |
| Ornithocercus thumii (Schmidt) Kofoid & Skogsberg | Protoperidinium acutum (Karsten) Balech |
| Oxytoxum crassum Schiller | Protoperidinium asymmetricum (Abé) Balech |
| Oxytoxum curvatum (Kofoid) Kofoid | Protoperidinium breve Paulsen |
| Oxytoxum elongatum Wood | Protoperidinium curtipes (Jörgensen) Balech |
| Oxytoxum laticeps Schiller | Protoperidinium depressum (Bailey) Balech |
| Oxytoxum milneri Murray & Whitting | Protoperidinium divergens (Ehrenberg) Balech |
| Oxytoxum mitra Stein | Protoperidinium elegans (Cleve) Balech |
| Oxytoxum mucronatum Hope | Protoperidinium exageratum Balech |
| Oxytoxum parvum Schiller | Protoperidinium globulus (Stein) Balech |
| Oxytoxum sceptrum (Stein) Schröder | Protoperidinium grande (Kofoid) Balech |
| Oxytoxum scolopax Stein | Protoperidinium heterocanthum (Dangeard) Balech |
| Oxytoxum sphaeroideum Stein | Protoperidinium latispinum (Mangin) Balech |
| Oxytoxum subulatum Kofoid | Protoperidinium leonis (Pavillard) Balech |
| Oxytoxum turbo Kofoid | Protoperidinium melo (Balech) Balech |
| Oxytoxum variabile Schiller | Protoperidinium obtusum (Karsten) Parke & Dodge |
| Palaeophalacroma unicinctum Schiller | Protoperidinium orientale (Matzenauer) Balech |
| Palaeophalacroma verrucosum Schiller | Protoperidinium parvum Abé |
| Podolampas bipes Stein | Protoperidinium porosum Balech |
| Podolampas palmipes Stein | Protoperidinium pyriforme (Paulsen) Balech |
| Podolampas spinifera Okamura | Protoperidinium quarnerense (B. Schröder) Balech |
| Pronoctiluca pelagica Fabre-Domerqne | Protoperidinium rhombiforme (Abé) Balech |
| Prorocentrum compressum (Ostenfeld) Abé | Protoperidinium schilleri (Paulsen) Balech |
| Prorocentrum dentatum Stein | Protoperidinium steinii (Jörgensen) Balech |
| Prorocentrum lenticulatum (Matzenauer) Taylor | Protoperidinium tenuissimum (Kofoid) Balech |
| Prorocentrum micans Ehrenberg | Protoperidinium tuba (Schiller) Balech |
| Prorocentrum minimun (Pavillard) Schiller | Protoperidinium variegatum (Peters) Balech |
| Prorocentrum rostratum Stein | Protoperidinium venustum (Matzenauer) Balech |
| Prorocentrum sigmoides Böhm | Pyrocystis lunula (Schütt) Schütt |
| Prorocentrum spp. | Pyrophacus steinii (Schiller) Wall & Dale |
| Prorocentrum triestinum Schiller | Schuettiella mitra (Schütt) Balech |
| Protoceratium areolatum Kofoid | Scrippsiella trochoidea (Stein) Loeblich III |
| Protoceratium reticulatum (Claparède & Lachmann) Butschli | Triposolenia bicornis Kofoid |
| Protoceratium spinulosum (Murray & Whitting) Schiller | Chrysophyta |
| Protoperidinium acanthophorum (Balech) Balech | Dictyocha fibula Ehrenberg |
| Protoperidinium achromaticum (Levander) Balech | Dictyocha speculum Ehrenberg |
References
- Pan, H.; Li, A.; Cui, Z.; Ding, D.; Qu, K.; Zheng, Y.; Lu, L.; Jiang, T.; Jiang, T. A comparative study of phytoplankton community structure and biomass determined by HPLC-CHEMTAX and microscopic methods during summer and autumn in the central Bohai Sea, China. Mar. Pollut. Bull. 2020, 155, 111172. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Gao, Y.; Zhou, F.; Shi, K.; Johnes, P.J.; Dungait, J.A.J.; Ma, M.; Lu, Y. Identifying the main drivers of change of phytoplankton community structure and gross primary productivity in a river-lake system. J. Hydrol. 2020, 583, 124633. [Google Scholar] [CrossRef]
- McCollin, T.; Shanks, A.M.; Dunn, J. Changes in zooplankton abundance and diversity after ballast water exchange in regional seas. Mar. Pollut. Bull. 2008, 56, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Righetti, D.; Vogt, M.; Gruber, N.; Psomas, A.; Zimmermann, E.N. Global pattern of phytoplankton diversity driven by temperature and environmental variability. Sci. Adv. 2019, 5, eaau6253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, J.P. Fall and rise of the phytoplankton. Nat. Clim. Change 2022, 12, 708–709. [Google Scholar] [CrossRef]
- Toseland, A.; Daines, S.J.; Clark, J.R.; Kirkham, A.; Strauss, J.; Uhlig, C.; Lenton, T.M.; Valentin, K.; Pearson, G.A.; Moulton, V.; et al. The impact of temperature on marine phytoplankton resource allocation and metabolism. Nat. Clim. Change 2013, 3, 979–984. [Google Scholar] [CrossRef]
- Doney, S.C. Plankton in a warmer world. Nature 2006, 444, 695–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.Y.; Wang, X.H. Progress on upwelling studies in the China seas. Rev. Geophys. 2016, 54, 653–673. [Google Scholar] [CrossRef]
- Li, D.; Chou, W.; Shih, Y.; Chen, G.; Chang, Y.; Chow, C.H.; Lin, T.; Hung, C. Elevated particulate organic carbon export flux induced by internal waves in the oligotrophic northern South China Sea. Sci. Rep. 2018, 8, 2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sherif, Z.; Mikhail, S.K. Phytoplankton dynamics in the southwestern part of Abu Qir Bay, Alexandria, Egypt. Egypt. J. Aquat. Biol. Fish 2003, 7, 219–239. [Google Scholar]
- Alprol, A.E.; Ashour, M.; Mansour, A.T.; Alzahrani, O.M.; Mahmoud, S.F.; Gharib, S.M. Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt. J. Mar. Sci. Eng. 2021, 9, 1328. [Google Scholar] [CrossRef]
- Alprol, A.E.; Heneash, A.M.M.; Ashour, M.; El-Kafrawy, S.; Soliman, A.M. Chemical assessment of water quality, heavy metals, and the distribution of zooplankton communities, based on field and GIS data in the drains of Burullus Lake, Egypt. Arab. J. Geosci. 2022, 15, 1511. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommung der quantitativen Phytoplankton-Methodik. Mitt. Inter. Verein. Lim. 1958, 9, 1–38. [Google Scholar]
- Sun, J.; Liu, D.Y.; Qian, S.B. A quantitative research and analysis method for marine phytoplankton: An introduction to Utermöhl method and its modification. J. Oceanogr. Huanghai Bohai Seas 2002, 20, 105–112. [Google Scholar]
- Niu, H.Y.; Chen, C.; Han, B.P. Quality and reliability of quantifying phytoplankton abundance and biomass data based upon the concentrated water sample method. J. Lake Sci. 2015, 05, 776–782. [Google Scholar]
- Yang, S.M.; Dong, S.G. Atlas of Common Planktonic Diatoms in China Sea Area; Ocean University of China Press: Qingdao, China, 2006; pp. 1–267. [Google Scholar]
- Yang, S.M.; Li, R.X.; Dong, S.G. Dinoflagellates I (Prorocentrales, Dinophysiaoes) in Chinese Waters; Ocean Press: Beijing, China, 2014; pp. 1–156. [Google Scholar]
- Yang, S.M.; Li, R.X.; Dong, S.G. Dinoflagellates Ⅱ (Goneaulacales) in Chinese Waters; Ocean Press: Beijing, China, 2016; pp. 1–248. [Google Scholar]
- Yang, S.M.; Li, R.X.; Dong, S.G. Dinoflagellates Ⅲ (Peridiniales) in Chinese Waters; Ocean Press: Beijing, China, 2019; pp. 1–211. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; pp. 1–125. [Google Scholar]
- Pielou, E.G. An introduction to mathematical ecology. J. Ecol. 1970, 58, 1–286. [Google Scholar]
- Lampitt, R.S.; Wishner, K.F.; Turley, C.M.; Angel, M.V. Marine snow studies in the Northeast Atlantic Ocean: Distribution, composition and role as a food source for migrating plankton. Mar. Biol. 1993, 116, 689–702. [Google Scholar] [CrossRef]
- Taylor, F. Scanning electron microscopy of thecae of the dinoflagellate genus ornithocercus. J. Phycol. 2010, 7, 249–258. [Google Scholar] [CrossRef]
- Sha, L.B.; Huang, Y.; Wang, L. Paleoenvironmental significance of thalassionema nitzschioides and its varieties of core 17940 in the South China Sea during the latest pleistocene. J. Meteorol. Enviroment 2008, 24, 5. [Google Scholar]
- Xu, Z.T.; Li, S.Y.; Hu, J.T.; Wang, S.Y.; Wang, B.; Guo, M.X. Summer phytoplankton responses to upwelling and river plume in northern South China Sea. J. Trop. Oceanogr. 2018, 37, 12. [Google Scholar]
- Shu, Y.Q.; Wang, Q.; Zu, T.T. Progress on shelf and slope circulation in the northern South China Sea. Sci. Sin. (Terrae) 2018, 48, 12. [Google Scholar] [CrossRef]
- Xiao, W.; Laws, E.A.; Xie, Y.; Wang, L.; Huang, B. Responses of marine phytoplankton communities to environmental changes: New insights from a niche classification scheme. Water Res. 2019, 166, 115070. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, M.; Zhong, Y.; Zhang, G.; Jiang, S.; Gao, Y.; Zhang, R.; Smith, W.O., Jr. Spatial variations of phytoplankton biomass controlled by river plume dynamics over the lower changjiang estuary and adjacent shelf based on high-resolution observations. Front. Mar. Sci. 2020, 7, 587539. [Google Scholar] [CrossRef]
- Zheng, M.L.; Li, M.M.; Xie, L.L.; Hong, Y.B.; He, Y.K.; Zong, X.L. Observation and analysis of winter hydrological characteristics of Northwest continental shelf of South China Sea in 2012. Oceanol. Et Limnol. Sin. 2018, 49, 12. [Google Scholar]
- Murrell, M.C.; Lores, E.M. Phytoplankton and zooplankton seasonal dynamics in a subtropical estuary: Importance of cyanobacteria. J. Plank. Res. 2004, 26, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Glibert, P.; Burkholder, J.; Kana, T.; Alexander, J.; Skelton, H.; Shilling, C. Grazing by karenia brevis on synechococcus enhances its growth rate and may help to sustain blooms. Aquat. Microb. Ecol. 2009, 55, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Ou, L.J.; Wang, D.; Huang, B.Q.; Hong, H.S.; Qi, Y.Z.; Lu, S.H. Comparative study of phosphorus strategies of three typical harmful algae in Chinese coastal waters. J. Plankton Res. 2008, 30, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Ou, L.J.; Huang, B.Q.; Hong, H.S.; Qi, Y.Z.; Lu, S.H. Comparative alkaline phosphatase characteristics of the algal bloom dinoflagellates Prorocentrum donghaiense and Alexandrium catenella, and the diatom Skeletonema costatum. J. Phycol. 2010, 46, 260–265. [Google Scholar] [CrossRef]
- Coble, G.P. Marine Optical Biogeochemistry: The Chemistry of Ocean Color. Chem. Rev. 2007, 107, 402–418. [Google Scholar] [CrossRef]
- Xue, B.; Sun, J.; Li, T.T. Phytoplankton community structure of northern South China Sea in summer of 2014. Acta Oceanol. Sin. 2016, 38, 54–65. [Google Scholar]
- Zhuang, W.; Hu, J.Y.; He, Z.G.; Zeng, G.N.; Chen, Z.Z. An analysis on surface temperature and salinity from southern Taiwan Strait to ZhuJiang River Estuary during July-August. J. Trop. Oceanogr. 2003, 22, 68–76. [Google Scholar]
- Gan, J.P.; Lu, Z.M.; Cheung, A.; Dai, M.H.; Liang, L.L.; Harrison, P.J.; Zhao, X.Z. Assessing ecosystem response to phosphorus and nitrogen limitation in the Pearl River plume using the Regional Ocean Modeling System (ROMS). J. Geophys. Res. Ocean. 2014, 119, 8858–8877. [Google Scholar] [CrossRef]
- Van Mooy, B.A.; Fredricks, H.F.; Pedler, B.E.; Dyhrman, S.T.; Karl, D.M.; Koblízek, M.; Lomas, M.W.; Mincer, T.J.; Moore, L.R.; Moutin, T.; et al. Phytoplankton in the ocean use non-phosphorus lipids in response to phosphorus scarcity. Nature 2009, 458, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.P.; Zhou, X.J.; Song, Y.Q. Effects of nitrogen and phosphorus supplies on the growth and intracellular composition for Chaetoceros gracilis. J. Aquac. 2020, 41, 24–29. [Google Scholar]
- Li, L.; Chen, Z.Z.; Huang, Z.R.; Xu, Y.W. Net-collected phytoplankton communitystructure in relation to environmental factors in the continental shelf of the northern South China Sea in spring 2015. J. Mar. Sci. 2019, 3, 86–96. [Google Scholar]
- Li, D.R.; Dai, X.F.; Lu, D.D.; Xia, P. Community structure of net-phytoplankton in the northwest South China Sea in the summer of 2012. J. Mar. Sci. 2014, 32, 87–96. [Google Scholar]
- Ma, W.; Sun, J.; Xue, B.; Dai, M.H.; Hu, J.Y. Study on phytoplankton community structure in northern South China Sea in autumn 2010. J. Mar. Sci. 2016, 38, 43–53. [Google Scholar]
- Li, X.; Sun, J.; Tian, W.; Wang, M. Phytoplankton communities in the northern South China Sea in summer 2009. J. Mar. Sci. 2012, 36, 33–39. [Google Scholar]
- Ling, J.; Dong, J.D.; Zhang, Y.Y.; Wang, Y.S.; Deng, C.; Lin, L.; Chen, L.; Li, T.; Long, L.J. Structure characteristics of phytoplankton community in Northern South China Sea in the summer of 2008. J. Biol. 2012, 29, 42–46. [Google Scholar]
- Ke, Z.X.; Huang, L.M.; Tan, Y.H.; Yin, J.Q. Species composition and abundance of phytoplankton in the northern South China Sea in summer 2007. J. Trop. Oceanogr. 2011, 30, 131–143. [Google Scholar]
- Le, F.F.; Sun, J.; Ning, X.R.; Song, S.Q.; Cai, Y.M.; Liu, C.G. Phytoplankton in the northern South China Sea in summer 2004. Oceanol. Limnol. Sin. 2006, 03, 238–248. [Google Scholar]
- Dam, H.V.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Neth. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar] [CrossRef]
- Tudesque, L.; Rimet, F.; Ector, L. A new taxon of the section Nitzschiae Lanceolatae Grunow: Nitzschia costei sp. nov. Compared to N. fonticola Grunow, N. macedonica Hustedt, N. tropica Hustedt and related species. Diatom Res. 2008, 23, 483–501. [Google Scholar] [CrossRef]
- Mukherjee, R.; Kumar, S.; Muduli, P.R. Spatial variation of nitrogen uptake rates in the largest brackish water lagoon of Asia (Chilika, India). Estuar. Coast. Shelf Sci. 2019, 216, 87–97. [Google Scholar] [CrossRef]
- Li, J.J.; Tan, Y.H.; Zhou, L.B.; Jiang, X.; Zhao, C.Y. Effects of nutrients on phytoplankton growth in the oligotrophic waters of north-eastern South China Sea. Mar. Sci. Bull. 2016, 35, 9. [Google Scholar]
- Jiang, Z.B.; Chen, J.F.; Zhou, F.S.; Shou, L.; Chen, Q.Z.; Tao, B.Y.; Yan, X.J.; Wang, K. Controlling factors of summer phytoplankton community in the Changjiang (Yangtze River) Estuary and adjacent East China Sea shelf. Cont. Shelf Res. 2015, 101, 71–84. [Google Scholar] [CrossRef]

| Sampling Data | Survey Area | Sampling Method | Number of Species | Average Cell Abundance (cells/L) | Average Diversity Index | Dominant Species | Reference |
|---|---|---|---|---|---|---|---|
| August 2020 | 15°–22° N 114°–120° E | Water sample | 332 | 649.97 | 3.95 | Nitzschia spp. Thalassionema nitzschioides Scrippsiella spp. | This study |
| May 2015 | 19°–23.5° N 110.5°–117.5° E | Net sample | 378 | 762.00 | 3.99 | Rhizosolenia alata Thalassiothrix frauenfeldii Pseudo-nitzschia pungens Eucampia zodiacus Nitzschia lorenziana Rhizosolenia gracillima | [40] |
| August 2014 | 18°–22° N 114°–116° E | Water sample | 229 | 16318 | 2.37 | Skeletonema costatum Fragilariopsis spp. Chaetoceros brevis T. nitzschioides Pseudo-nitzschia delicatissima Pseudo-nitzschia pungens Chaetoceros compressus Chaetoceros lorenzianus Chaetoceros pelagicus | [35] |
| August 2012 | 11°–22° N 110°–116.5° E | Net sample | 206 | 666.70 | 2.67 | Thalassiothrix frauenfeldii Rhizosolenia alata Thalassionema T. nitzschioides | [41] |
| October 2010~November 2010 | 18°–23.5° N 110.5°–118° E | Water sample | 204 | 500.00 | 3.14 | Thalassionema T. nitzschioides Navicula spp. Skeletonema costatum Chaetoceros curvisetus Rhizosolenia stolterfothii Paralia sulcata | [42] |
| August 2009 | 18°–22° N 110°–117° E | Water sample | 109 | 819.70 | — | Pseudo-nitzschia delicatissima Thalassiothrix frauenfeldii Pseudo-nitzschia pungens Detonula pumila Protoperidinium spp. Asterionella glacialis | [43] |
| August 2008 | 18°–23° N 110°–120° E | Net sample | 169 | 180.60 | — | Chaetoceros lorenzianus Pseudo-nitzschia delicatissima Thalassionema T. nitzschioides | [44] |
| August 2007 | 18°–23° N 110°–120° E | Water sample | 216 | 11,220 | 2.62 | Thalassionema T. nitzschioides Thalassiosira spp. Skeletonema costatum Prorocentrum minimun Gymnodinium spp. | [45] |
| August 2004–September 2004 | 18°–22° N 110°–117° E | Water sample | 159 | 115,050 | 2.08 | Pseudo-nitzschia delicatissima Chaetoceros curvisetus Chaetoceros diadema Prorocentrum dentatum Asterionellopsis glacialis Thalassionema T. nitzschioides Chaetoceros lorenzianus Bacteriastrum comosum Emiliania huxleyi | [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Gao, J.; Xu, Z.; Yan, Y.; Yang, S. Phytoplankton Community Diversity and Its Environmental Driving Factors in the Northern South China Sea. Water 2022, 14, 3777. https://doi.org/10.3390/w14223777
Chen W, Gao J, Xu Z, Yan Y, Yang S. Phytoplankton Community Diversity and Its Environmental Driving Factors in the Northern South China Sea. Water. 2022; 14(22):3777. https://doi.org/10.3390/w14223777
Chicago/Turabian StyleChen, Wenqing, Jie Gao, Zongjun Xu, Yantao Yan, and Shimin Yang. 2022. "Phytoplankton Community Diversity and Its Environmental Driving Factors in the Northern South China Sea" Water 14, no. 22: 3777. https://doi.org/10.3390/w14223777