

Factors Affecting the Interpretation of Online Phycocyanin Fluorescence to Manage Cyanobacteria in Drinking Water Sources

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Microscopic Taxonomic Cell Counts
2.3. Fluorometer Measurements
2.4. NOM Addition
2.5. Water Temperature Effect
2.6. Fluorescence Excitation-Emission Matrix Spectra Analysis
2.7. Field Sampling
2.8. Data Analysis
3. Results and Discussion
3.1. Addition of NOM
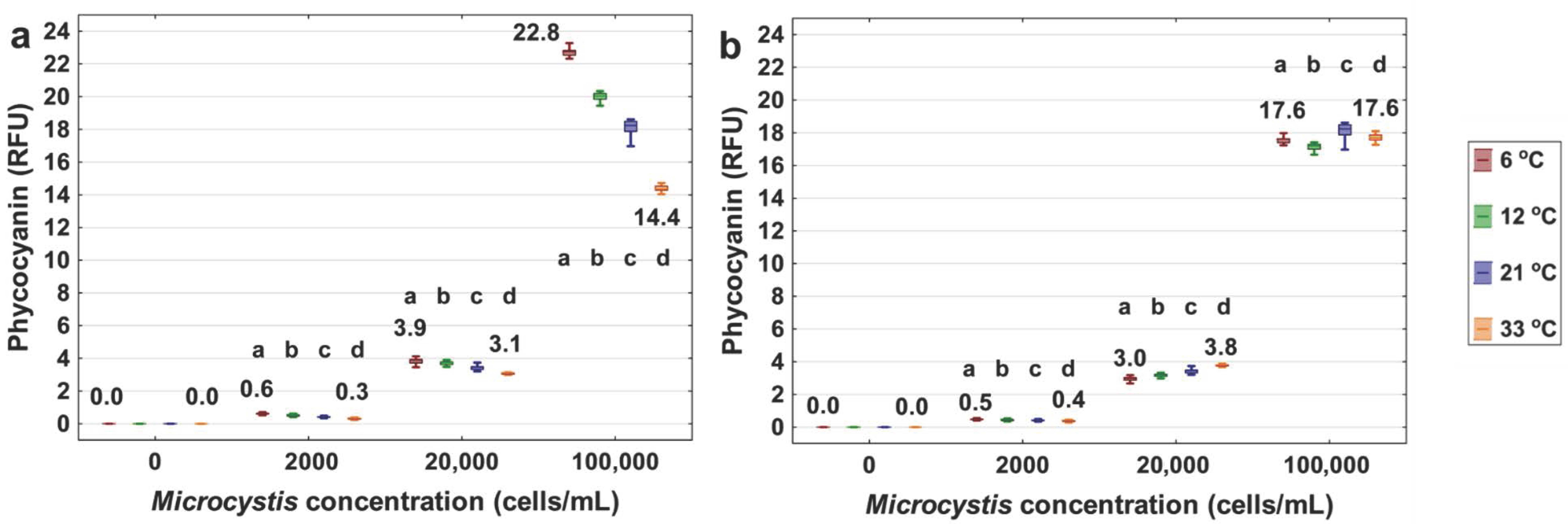
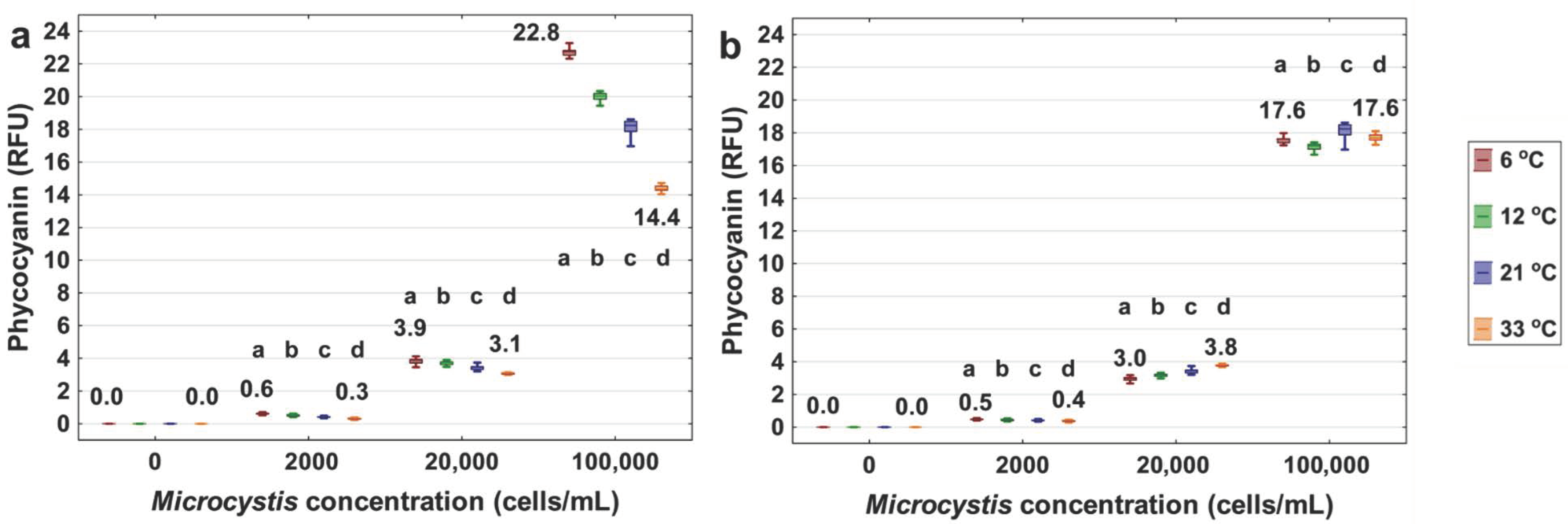
3.1.1. Impact of NOM on Phycocyanin Probe Performance
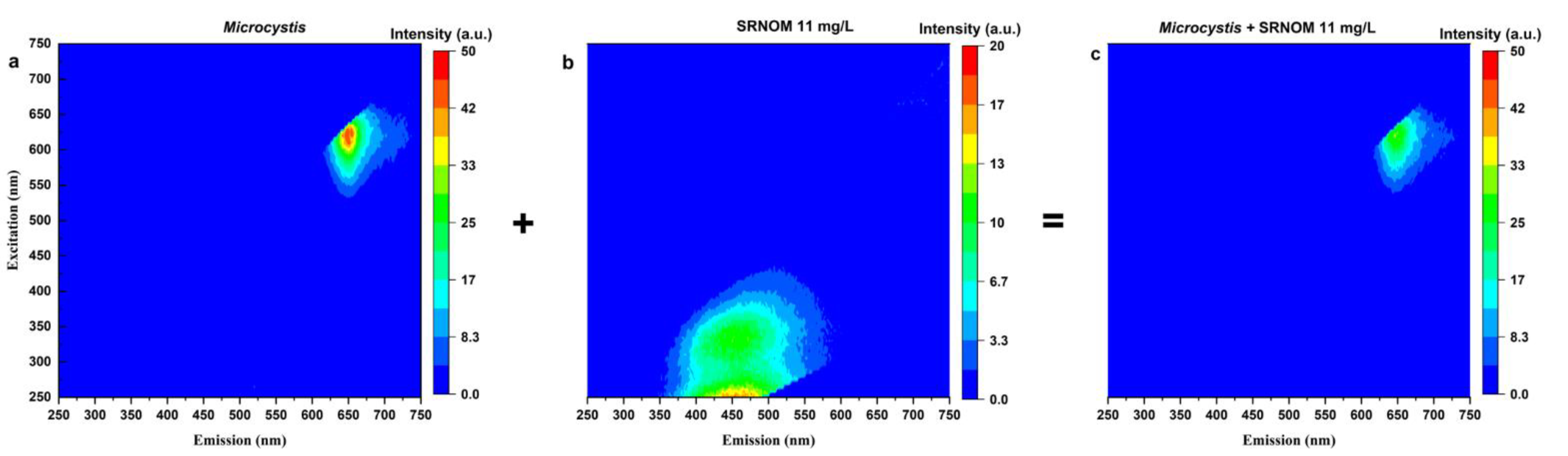
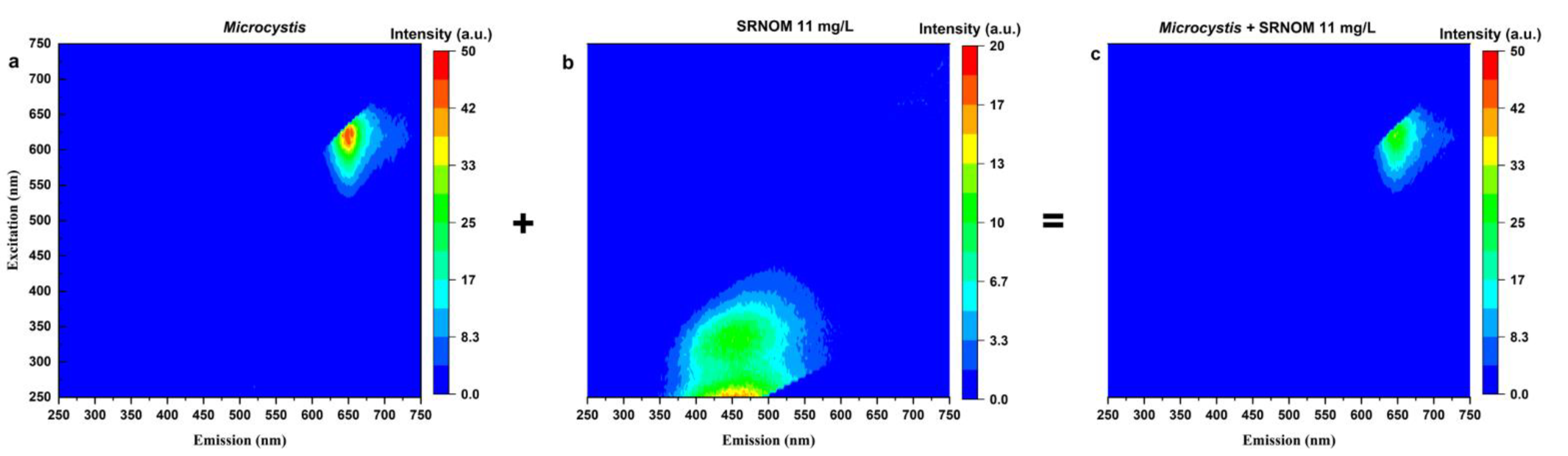
3.1.2. Fluorescence EEM Studies
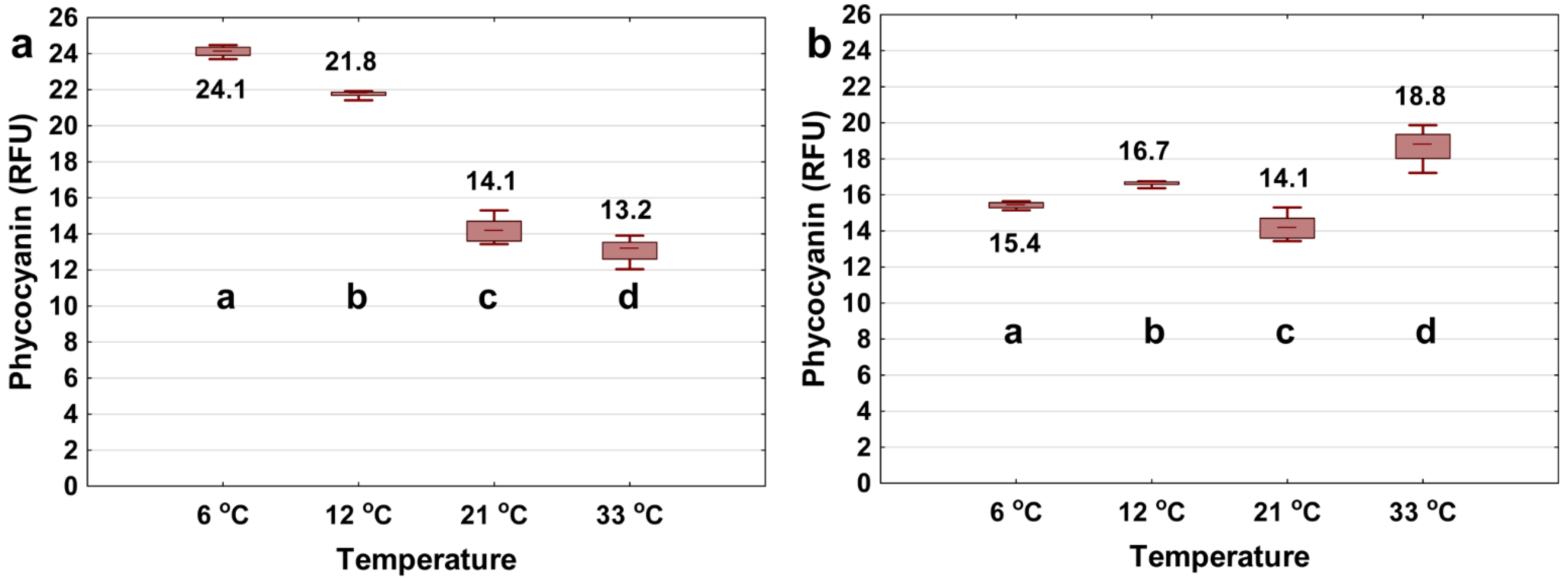
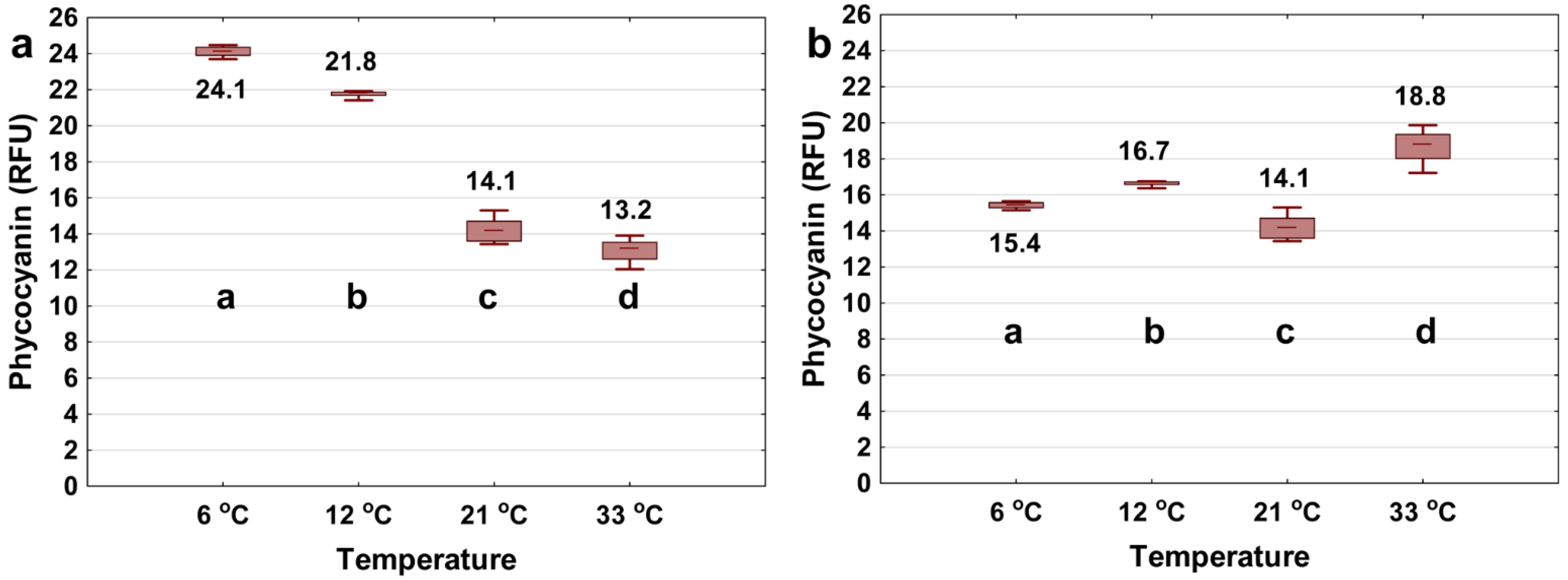
3.2. Varying Water Matrix Temperature
3.3. Validation of Probes with Field Samples
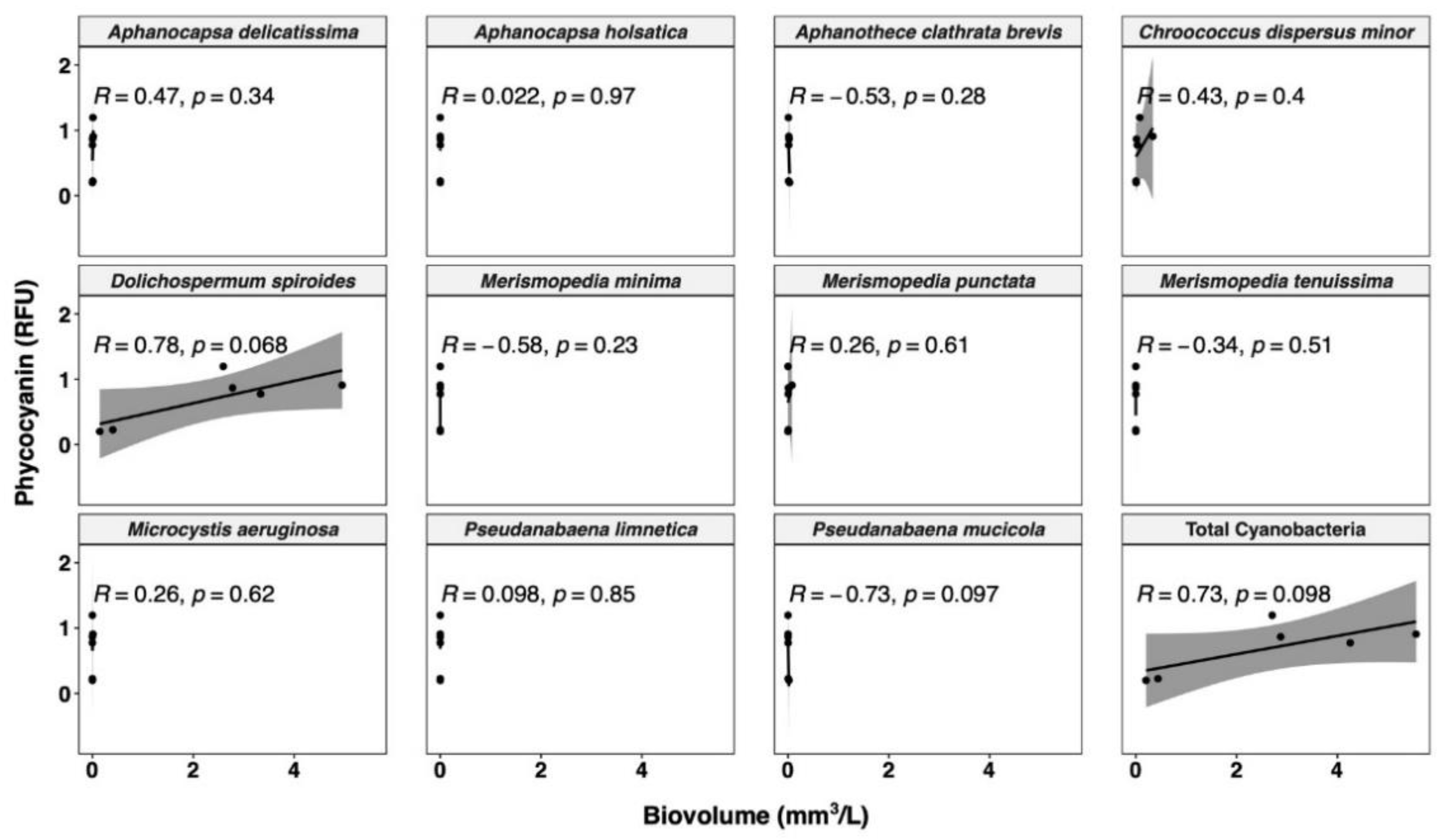
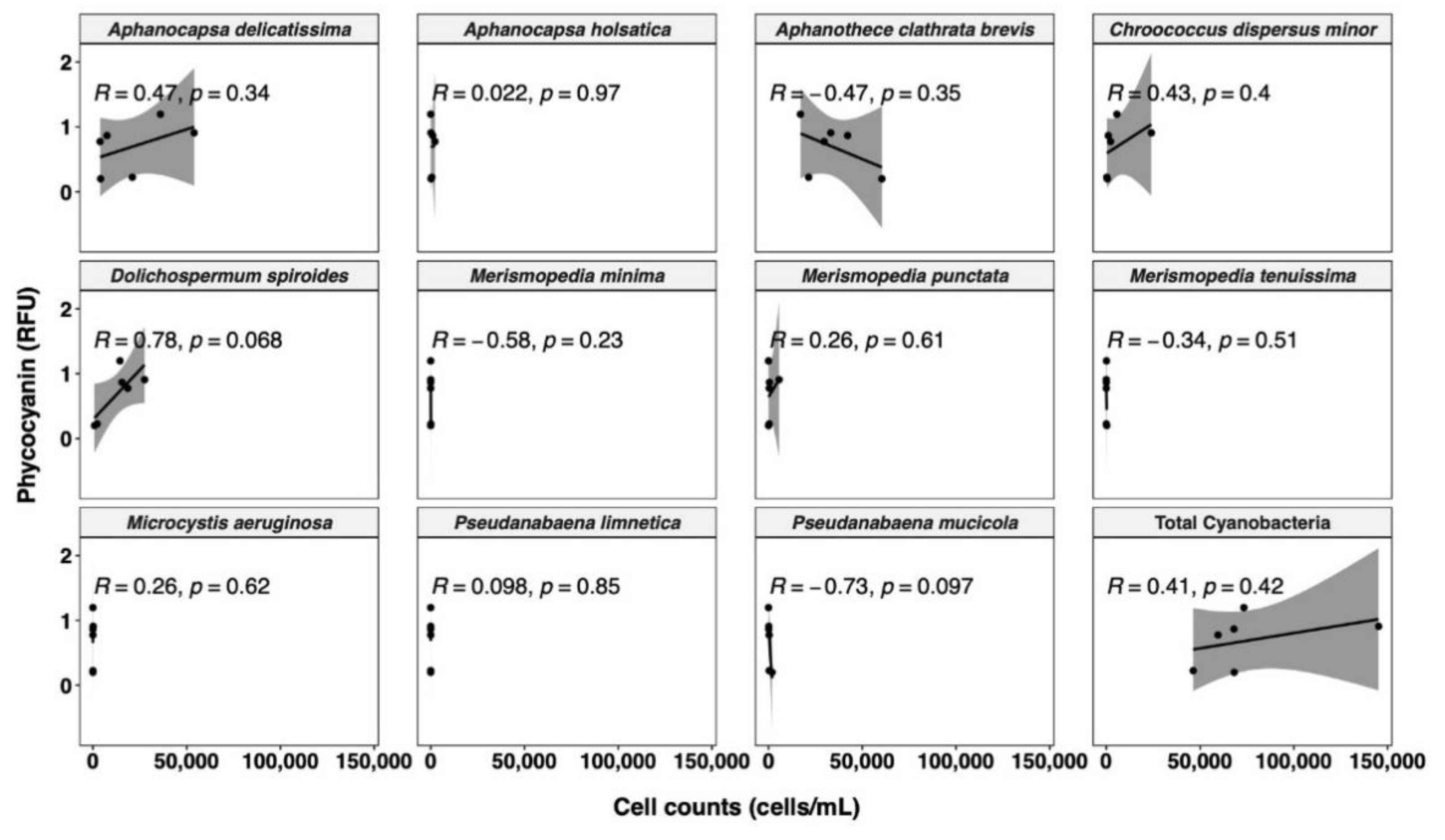
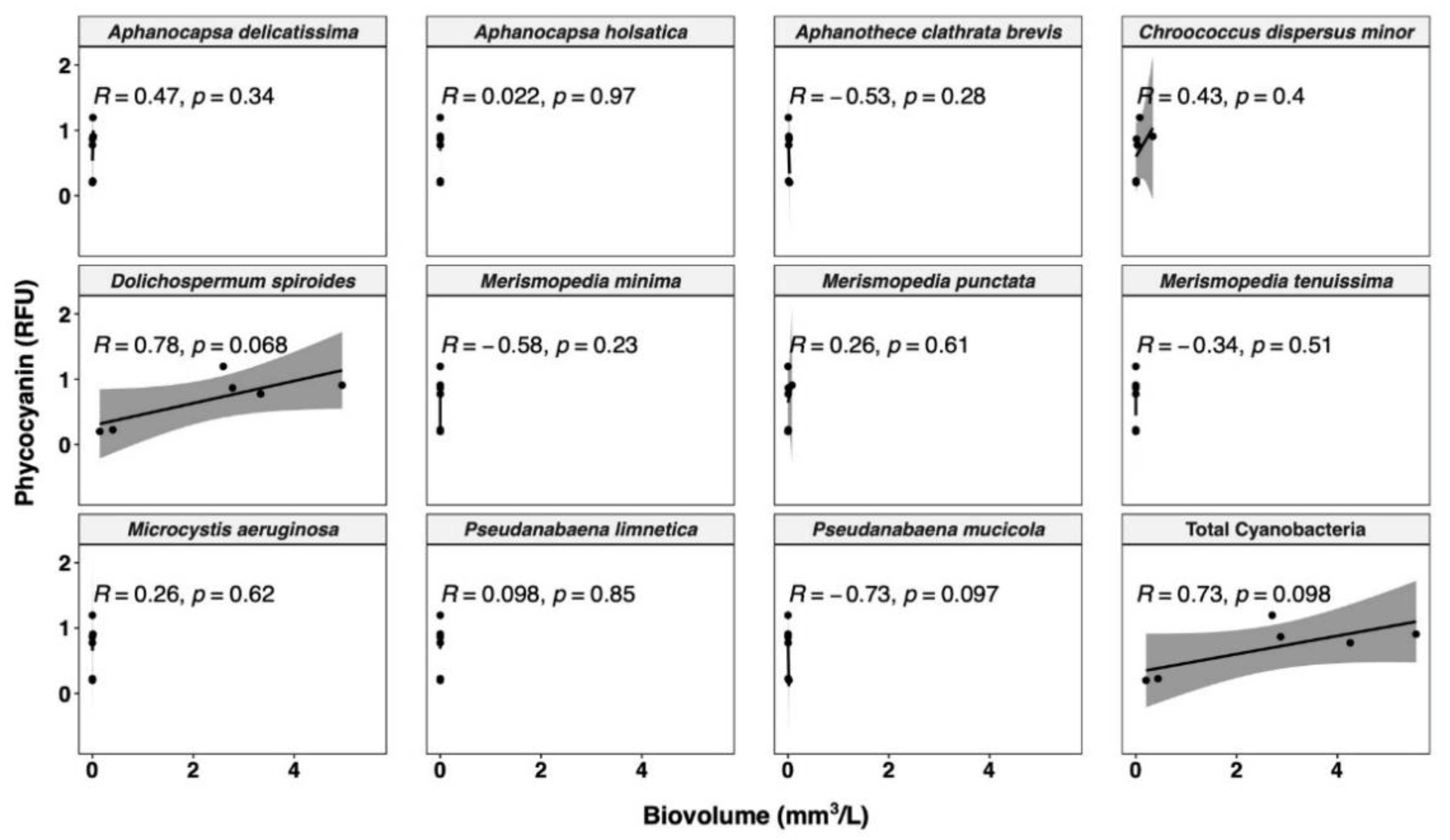
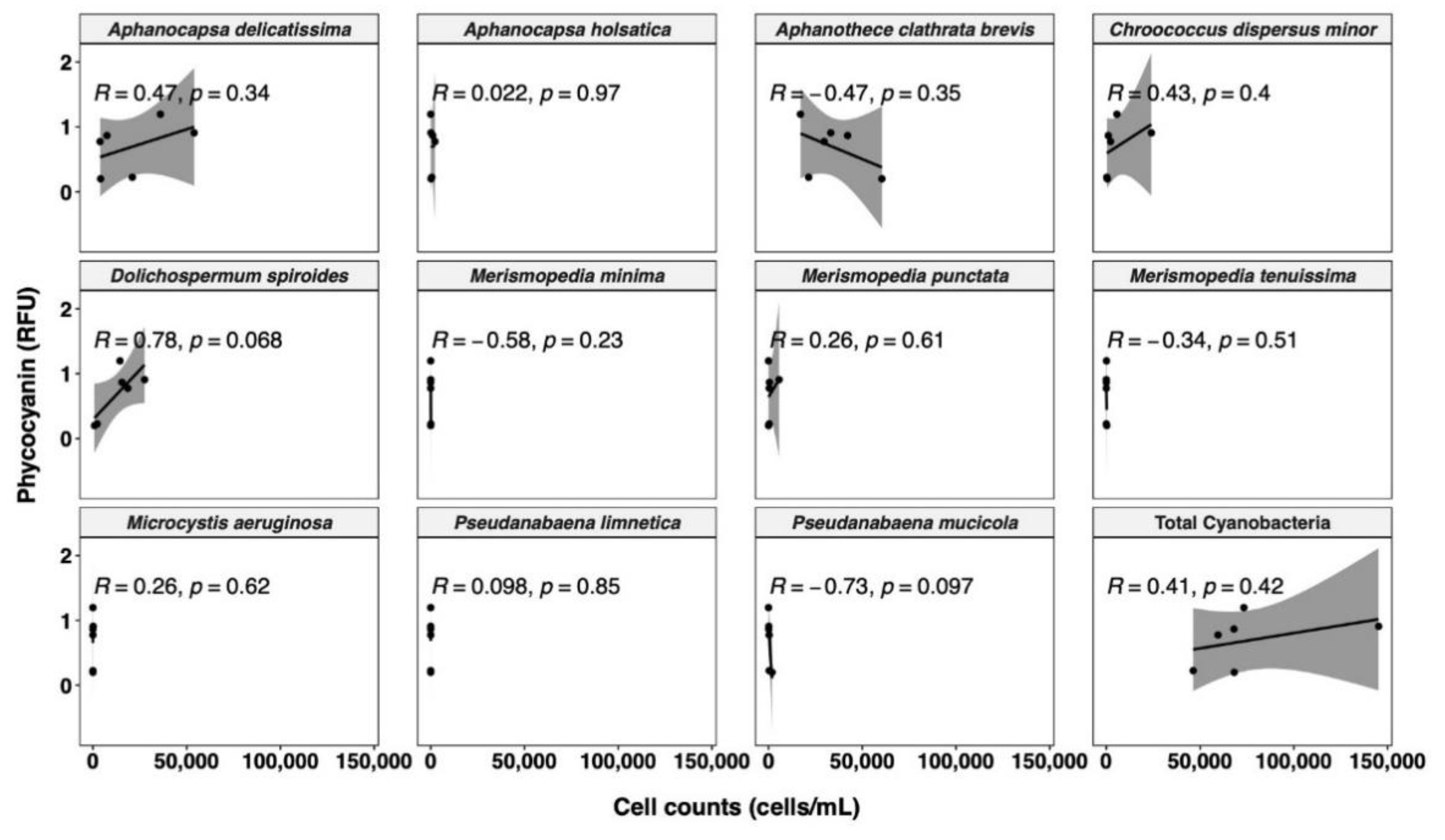
3.3.1. Relationships between Cyanobacterial Biovolume and Phycocyanin Probe Measurements
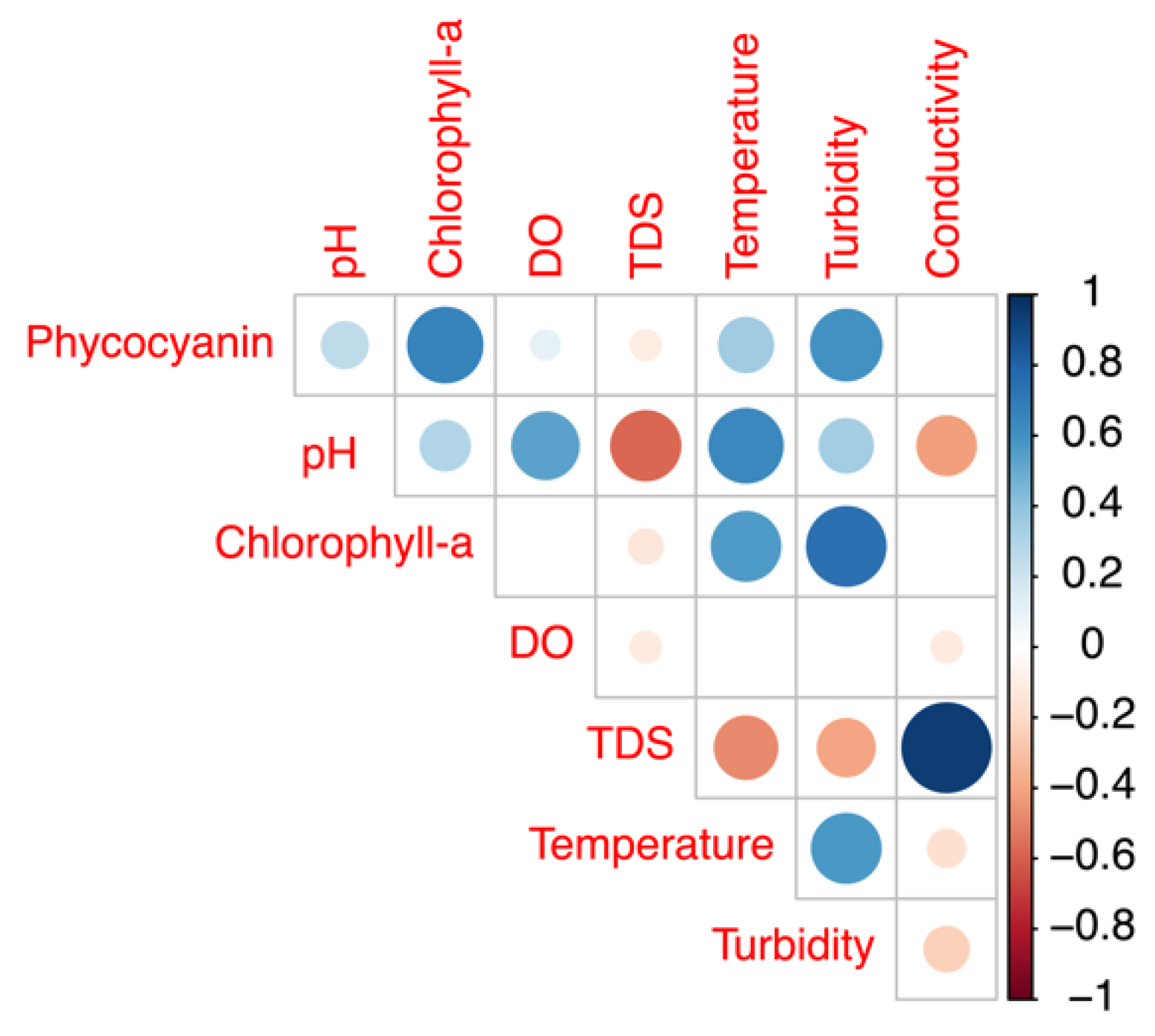
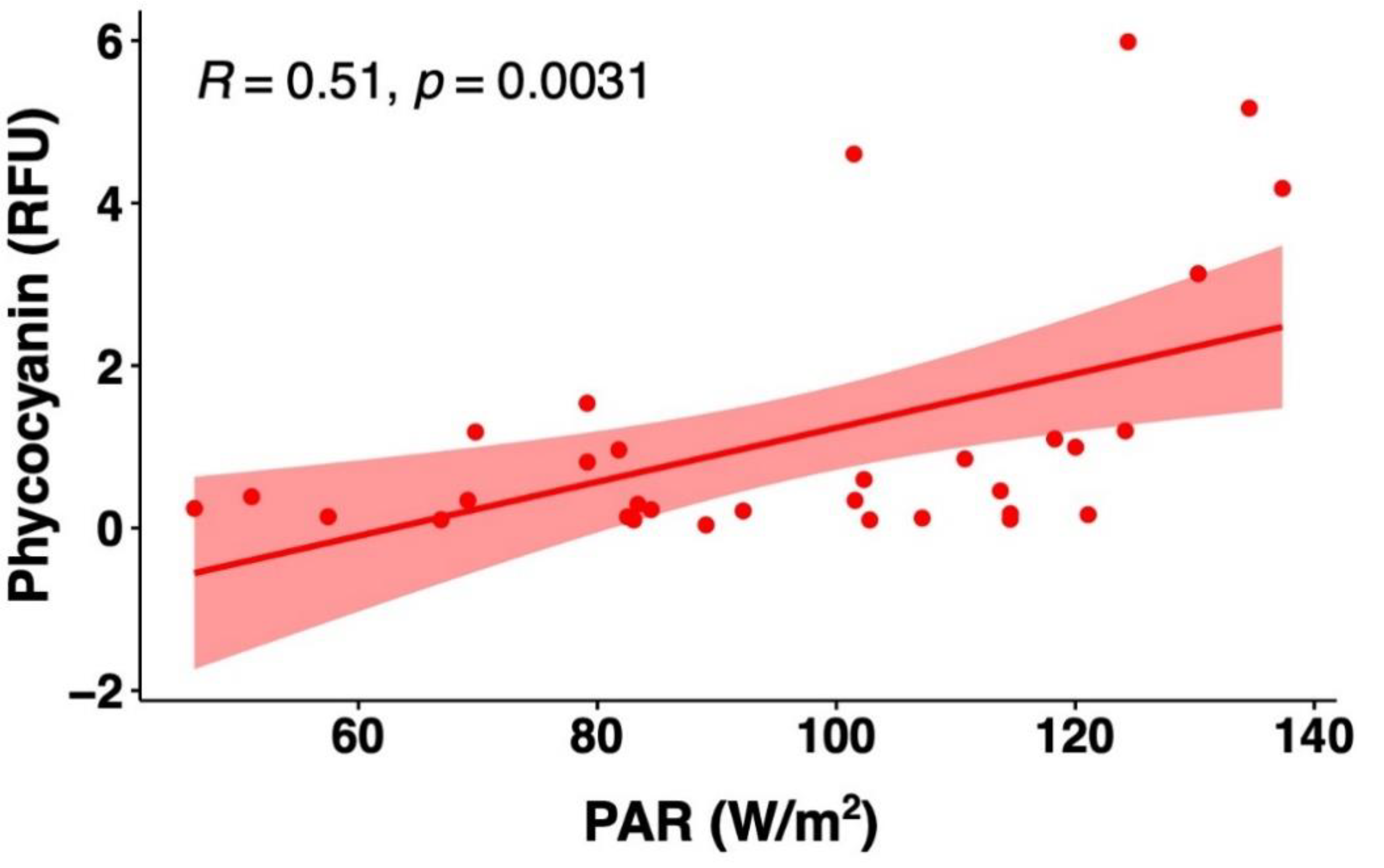
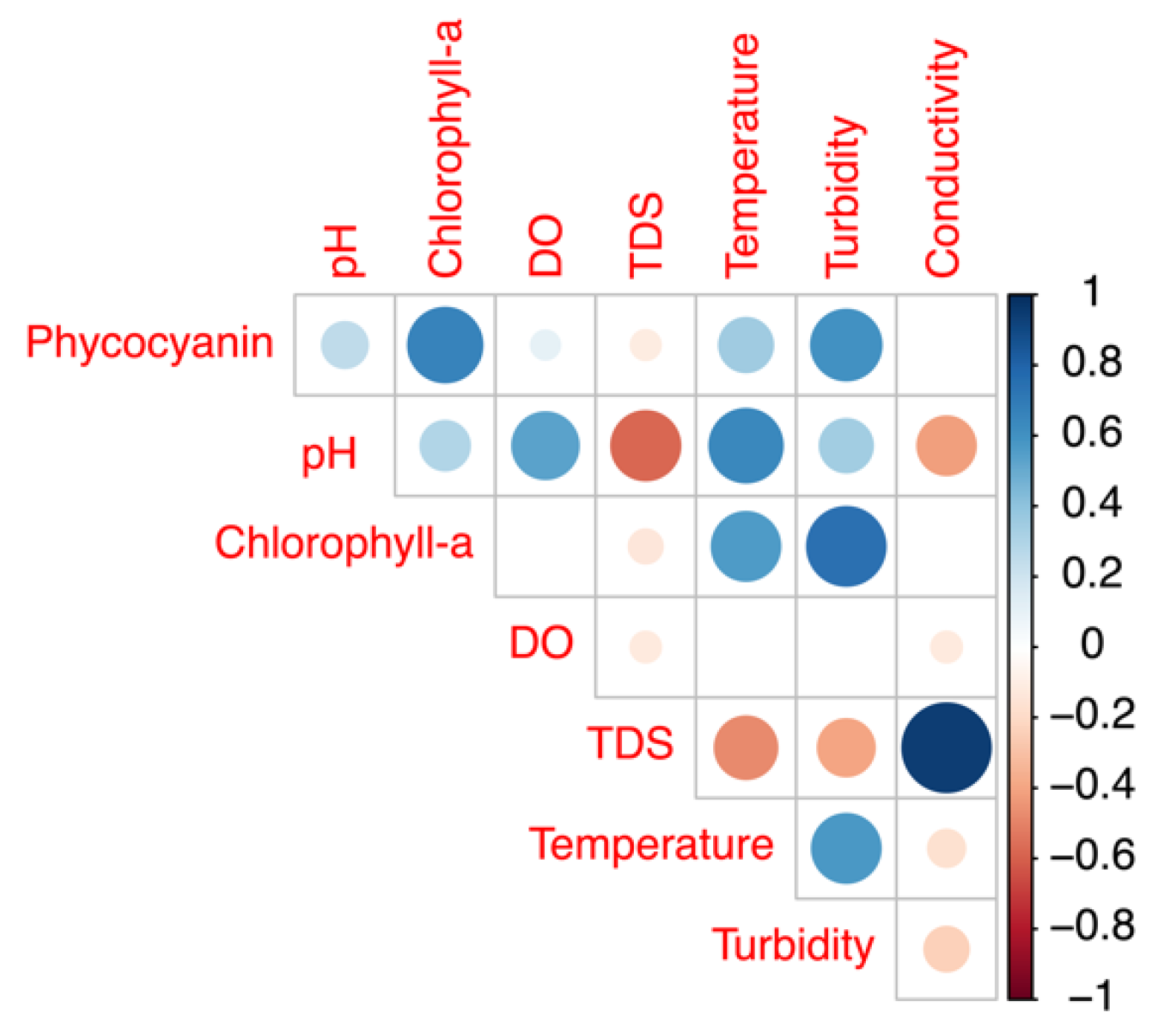
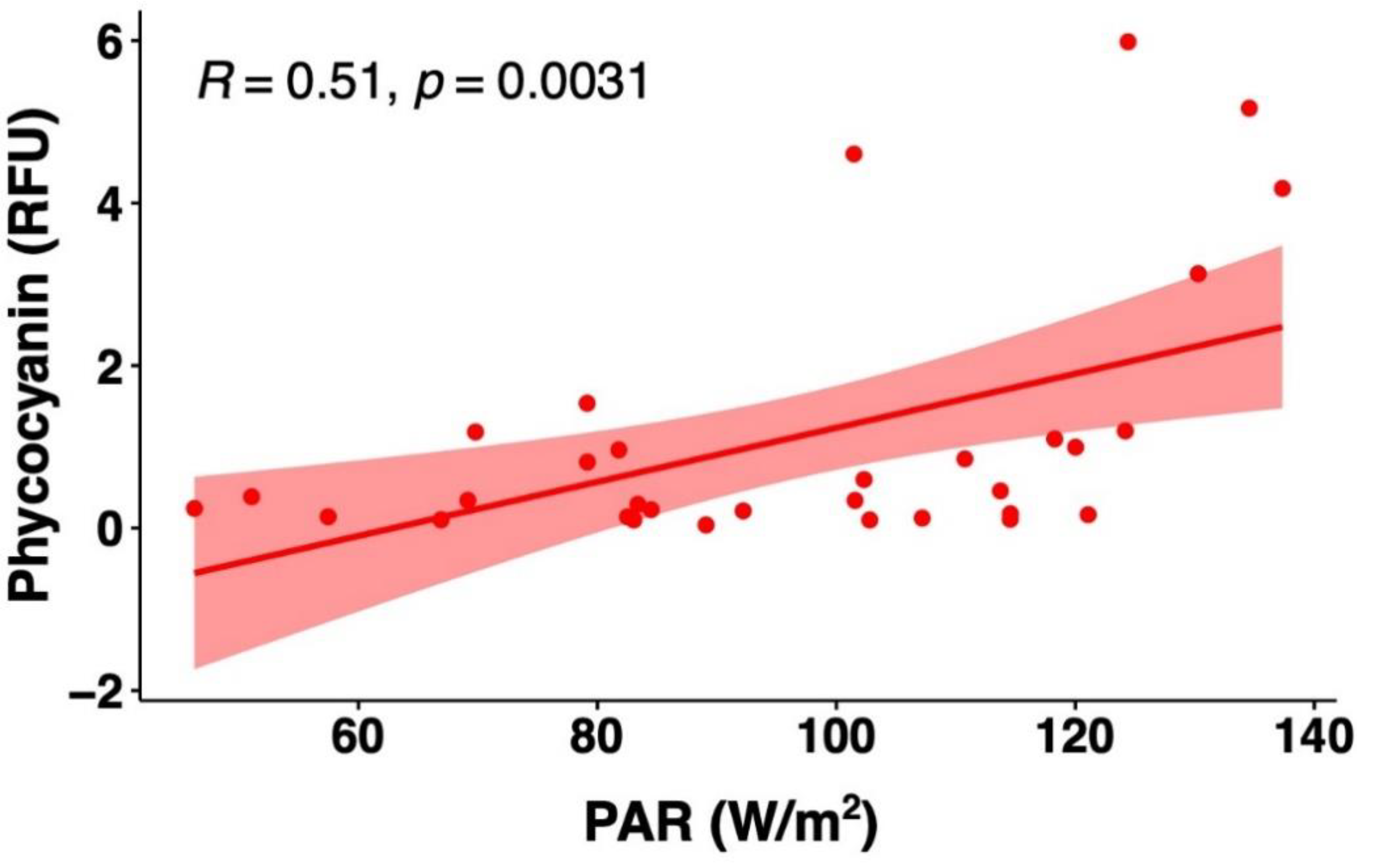
3.3.2. Comparison of Field Phycocyanin Measurements and Other Water Quality Parameters
4. Conclusions
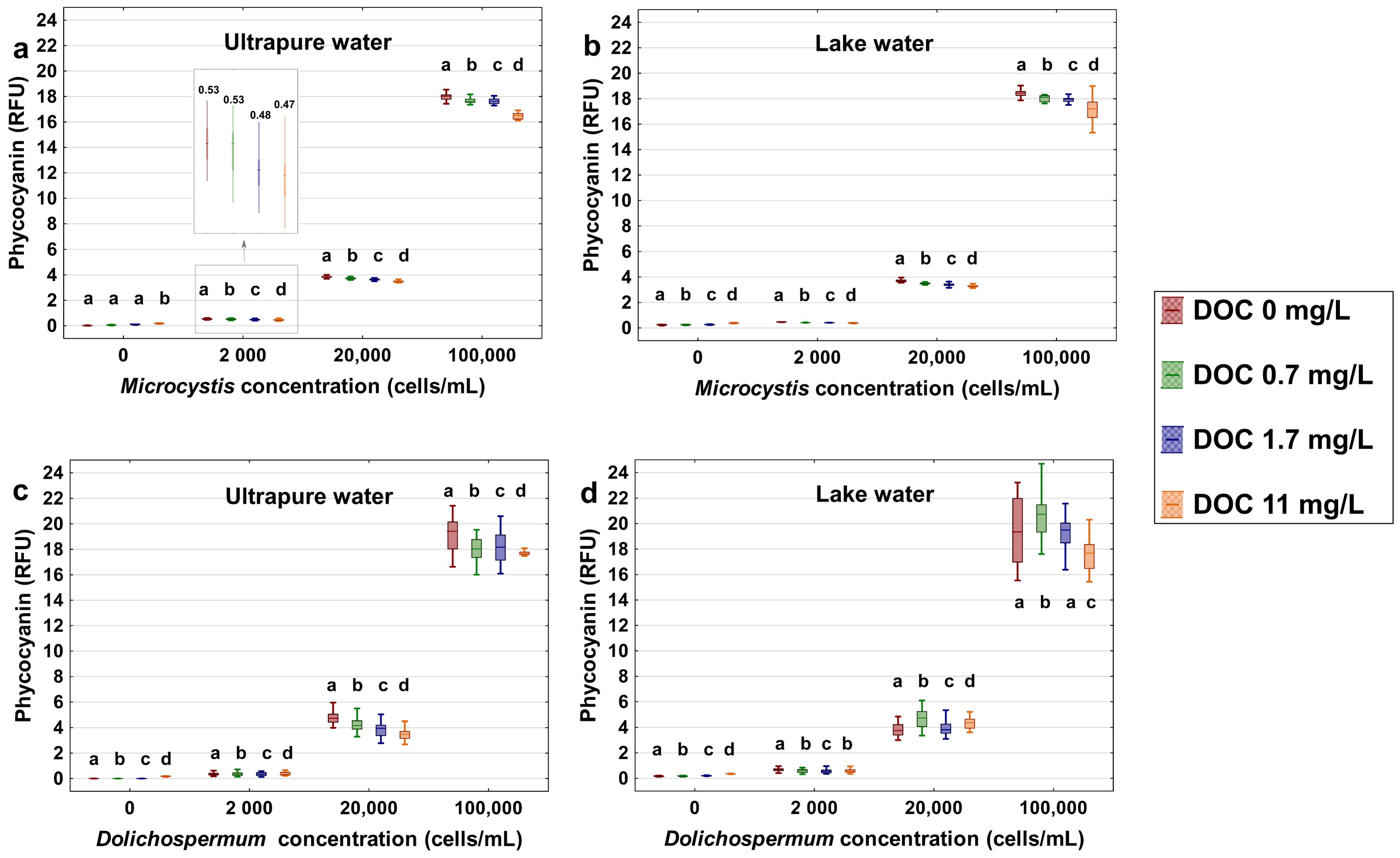
- SRNOM can quench and interfere with the reliability of the cell-bound phycocyanin readings. High SRNOM content in the water matrix can lead to an underestimation of cyanobacterial quantification measured by a phycocyanin probe, especially for high cyanobacterial cell concentrations.
- The concentration of NOM added, the aromaticity of NOM, the species present, and the background DOC in the test water define the extent and the trends for fluorescence quenching.
- In the case of a source with high naturally occurring NOM, DWTP managers need to assess the interference caused by NOM, and the phycocyanin RFU-based thresholds can be adjusted accordingly.
- Phycocyanin probe readings can be significantly affected by temperature in the range of their typical application (6–33 °C). Applying the correction algorithms can yield high success rates to decrease the interference. The relative importance of such interference varies between commercially available probes.
- The correlation between probe readings and taxonomic cell counts is unreliable, and only biovolume-based estimations are a relevant option for management purposes.
- A simultaneous measurement of the other physical-chemical parameters may help increase the accuracy of phycocyanin readings.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baures, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G. Harmful algal blooms and public health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Alex, E.J. Is the future blue-green? A review of the current model predictions of how climate change could affect pelagic freshwater cyanobacteria. Water Res. 2012, 46, 1364–1371. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef]
- Beaulieu, M.; Pick, F.; Gregory-Eaves, I. Nutrients and water temperature are significant predictors of cyanobacterial biomass in a 1147 lakes data set. Limnol. Oceanogr. 2013, 58, 1736–1746. [Google Scholar] [CrossRef]
- Richardson, J.; Feuchtmayr, H.; Miller, C.; Hunter, P.D.; Maberly, S.C.; Carvalho, L. Response of cyanobacteria and phytoplankton abundance to warming, extreme rainfall events and nutrient enrichment. Glob. Change Biol. 2019, 25, 3365–3380. [Google Scholar] [CrossRef] [Green Version]
- Zamyadi, A.; McQuaid, N.; Prévost, M.; Dorner, S. Monitoring of potentially toxic cyanobacteria using an online multi-probe in drinking water sources. J. Environ. Monit. 2012, 14, 579–588. [Google Scholar] [CrossRef]
- Bertone, E.; Burford, M.A.; Hamilton, D.P. Fluorescence probes for real-time remote cyanobacteria monitoring: A review of challenges and opportunities. Water Res. 2018, 141, 152–162. [Google Scholar] [CrossRef]
- Zamyadi, A.; Choo, F.; Newcombe, G.; Stuetz, R.; Henderson, R.K. A review of monitoring technologies for real-time management of cyanobacteria: Recent advances and future direction. TrAC Trends Anal. Chem. 2016, 85, 83–96. [Google Scholar] [CrossRef]
- Lou, I.; Han, B.; Zhang, W. Advances in Monitoring and Modelling Algal Blooms in Freshwater Reservoirs: General principles and a Case Study of Macau; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Hawkins, P.R.; Holliday, J.; Kathuria, A.; Bowling, L. Change in cyanobacterial biovolume due to preservation by Lugol’s Iodine. Harmful Algae 2005, 4, 1033–1043. [Google Scholar] [CrossRef]
- Newcombe, G. International Guidance Manual for the Management of Toxic Cyanobacteria; Global Water Research Coalition and Water Quality Research Australia: London, UK, 2009; p. 44. [Google Scholar]
- America Water Works Association (AWWA). Algae Source to Treatment. Manual of Water Supply Practices - M57, 1st ed.; America Water Works Association (AWWA): Denver, CO, USA, 2010; p. 481. [Google Scholar]
- Sanseverino, I.; Conduto António, D.; Loos, R.; Lettieri, T. Cyanotoxins: Methods and Approaches for Their Analysis and Detection; EUR 28624; European Commission: Brussels, Belgium, 2017; p. 70. [Google Scholar]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Taylor & Francis: Abingdon, UK, 2021; p. 859. [Google Scholar]
- Serrano-Silva, N.; Calderón-Ezquerro, M.C. Metagenomic survey of bacterial diversity in the atmosphere of Mexico City using different sampling methods. Environ. Pollut. 2018, 235, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Singh, S.; Ahn, C.-Y.; Oh, H.-M.; Asthana, R.K. Monitoring approaches for a toxic cyanobacterial Bloom. Environ. Sci. Technol. 2013, 47, 8999–9013. [Google Scholar] [CrossRef]
- Romanis, C.S.; Pearson, L.A.; Neilan, B.A. Cyanobacterial blooms in wastewater treatment facilities: Significance and emerging monitoring strategies. J. Microbiol. Methods 2021, 180, 106123. [Google Scholar] [CrossRef] [PubMed]
- John, N.; Koehler, A.V.; Ansell, B.R.E.; Baker, L.; Crosbie, N.D.; Jex, A.R. An improved method for PCR-based detection and routine monitoring of geosmin-producing cyanobacterial blooms. Water Res. 2018, 136, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Te, S.H.; Chen, E.Y.; Gin, K.Y.-H. Comparison of quantitative PCR and droplet digital PCR multiplex assays for two genera of bloom-forming cyanobacteria, Cylindrospermopsis and Microcystis. Appl. Environ. Microbiol. 2015, 81, 5203–5211. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.T.; Chen, Y.H.; Wang, T.S.; Yen, H.K.; Lin, T.F. A qPCR-based tool to diagnose the presence of harmful cyanobacteria and cyanotoxins in drinking water sources. Int. J. Environ. Res. Public Health 2017, 14, 547. [Google Scholar] [CrossRef] [Green Version]
- Tromas, N.; Fortin, N.; Bedrani, L.; Terrat, Y.; Cardoso, P.; Bird, D.; Greer, C.W.; Shapiro, B.J. Characterising and predicting cyanobacterial blooms in an 8-year amplicon sequencing time course. ISME J. 2017, 11, 1746–1763. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Zhang, J.; Wang, X.; Yang, Y.; Chen, N.; Lu, Z.; Ge, Q.; Jiang, R.; Zhang, X.; Yang, Y.; et al. Responses of cyanobacterial aggregate microbial communities to algal blooms. Water Res. 2021, 196, 117014. [Google Scholar] [CrossRef]
- Moradinejad, S.; Trigui, H.; Guerra Maldonado, J.F.; Shapiro, J.; Terrat, Y.; Zamyadi, A.; Dorner, S.; Prévost, M. Diversity assessment of toxic cyanobacterial blooms during oxidation. J. Toxins 2020, 12, 728. [Google Scholar] [CrossRef]
- Zamyadi, A.; Romanis, C.; Mills, T.; Neilan, B.; Choo, F.; Coral, L.A.; Gale, D.; Newcombe, G.; Crosbie, N.; Stuetz, R.; et al. Diagnosing water treatment critical control points for cyanobacterial removal: Exploring benefits of combined microscopy, next-generation sequencing, and cell integrity methods. Water Res. 2019, 152, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Jalili, F.; Trigui, H.; Guerra Maldonado, J.F.; Dorner, S.; Zamyadi, A.; Shapiro, B.J.; Terrat, Y.; Fortin, N.; Sauvé, S.; Prévost, M. Can cyanobacterial diversity in the source predict the diversity in sludge and the risk of toxin release in a drinking water treatment plant? Toxins 2021, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Bharti, R.; Grimm, D.G. Current challenges and best-practice protocols for microbiome analysis. Brief Bioinform 2021, 22, 178–193. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Ng, C.; Nshimyimana, J.; Loh, L.-L.; Gin, K.; Thompson, J. Next-generation sequencing (NGS) for assessment of microbial water quality: Current progress, challenges, and future opportunities. Front. Microbiol. 2015, 6, 1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, D.A. The cyanobacterial photosynthetic apparatus: Comparisosn to those of higher plants and photosynthetic bacteria. Can. Bull. Fish. Aquat. Sci. 1986, 214, 423–500. [Google Scholar]
- Carr, N.G.; Whitton, B.A. The Biology of Cyanobacteria; University of California Press: Berkeley, CA, USA, 1982; p. 688. [Google Scholar]
- Prescott, L.M.; Harley, J.P.; Klein, D.A. Microbiology, 6th ed.; The McGraw Hill Companies Inc.: New York, NY, USA, 2005; p. 1152. [Google Scholar]
- McQuaid, N.; Zamyadi, A.; Prévost, M.; Bird, D.F.; Dorner, S. Use of in vivo phycocyanin fluorescence to monitor potential microcystin-producing cyanobacterial biovolume in a drinking water source. J. Environ. Monit. 2011, 13, 455–463. [Google Scholar] [CrossRef]
- Pazouki, P.; Dorner, S.; Bouchard, R.; Zamyadi, A. Applying an online multi-probe to monitor the potentially toxic cyanobacteria in Lake Erie. In Proceedings of the 29th Eastern Canadian Symposium on Water Quality Research, Montréal, QC, Canada, 17 October 2014. [Google Scholar]
- Zamyadi, A.; Dorner, S.; Ndong, M.; Bolduc, A.; Bastien, C.; Prévost, M. Application of in vivo measurements for the management of cyanobacterial cell breakthrough into drinking water treatment plants. Environ. Sci. Process. Impacts 2014, 16, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.l.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Korak, J.A.; Wert, E.C.; Rosario-Ortiz, F.L. Evaluating fluorescence spectroscopy as a tool to characterize cyanobacteria intracellular organic matter upon simulated release and oxidation in natural water. Water Res. 2015, 68, 432–443. [Google Scholar] [CrossRef]
- Chang, D.-W.; Hobson, P.; Burch, M.; Lin, T.-F. Measurement of cyanobacteria using in-vivo fluoroscopy-Effect of cyanobacterial species, pigments, and colonies. Water Res. 2012, 46, 5037–5048. [Google Scholar] [CrossRef]
- Hodges, C.M.; Wood, S.A.; Puddick, J.; McBride, C.G.; Hamilton, D.P. Sensor manufacturer, temperature, and cyanobacteria morphology affect phycocyanin fluorescence measurements. Environ. Sci. Pollut. Res. 2018, 25, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Thomson-Laing, G.; Puddick, J.; Wood, S.A. Predicting cyanobacterial biovolumes from phycocyanin fluorescence using a handheld fluorometer in the field. Harmful Algae 2020, 97, 101869. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; McQuaid, N.; Dorner, S.; Bird, D.F.; Burch, M.; Baker, P.; Hobson, P.; Prévost, M. Cyanobacterial detection using in vivo fluorescence probes: Managing interferences for improved decision-making. J. Am. Water Work. Assoc. 2012, 104, E466–E479. [Google Scholar] [CrossRef]
- Rousso, B.Z.; Bertone, E.; Stewart, R.A.; Rinke, K.; Hamilton, D.P. Light-induced fluorescence quenching leads to errors in sensor measurements of phytoplankton chlorophyll and phycocyanin. Water Res. 2021, 198, 117133. [Google Scholar] [CrossRef] [PubMed]
- Misumi, M.; Katoh, H.; Tomo, T.; Sonoike, K. Relationship between photochemical quenching and non-photochemical quenching in six species of cyanobacteria reveals species difference in redox state and species commonality in energy dissipation. Plant Cell Physiol. 2016, 57, 1510–1517. [Google Scholar] [CrossRef]
- Brient, L.; Lengronne, M.; Bertrand, E.; Rolland, D.; Sipel, A.; Steinmann, D.; Baudin, I.; Legeas, M.; Le Rouzic, B.; Bormans, M. A phycocyanin probe as a tool for monitoring cyanobacteria in freshwater bodies. J. Environ. Monit. 2008, 10, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Choo, F.; Zamyadi, A.; Newton, K.; Newcombe, G.; Bowling, L.; Stuetzb, R.; Henderson, R.K. Performance evaluation of in situ fluorometers for real-time cyanobacterial monitoring. H2Open J. 2018, 1, 26–46. [Google Scholar] [CrossRef]
- Choo, F.; Zamyadi, A.; Stuetz, R.M.; Newcombe, G.; Newton, K.; Henderson, R.K. Enhanced real-time cyanobacterial fluorescence monitoring through chlorophyll-a interference compensation corrections. Water Res. 2019, 148, 86–96. [Google Scholar] [CrossRef]
- Bowling, L.C.; Merrick, C.; Swann, J.; Green, D.; Smith, G.; Neilan, B.A. Effects of hydrology and river management on the distribution, abundance and persistence of cyanobacterial blooms in the Murray River, Australia. Harmful Algae 2013, 30, 27–36. [Google Scholar] [CrossRef]
- Symes, E.; Van Ogtrop, F. Determining the Efficacy of a Submersible in situ Fluorometric Device for Cyanobacteria Monitoring Coalesced with Total Suspended Solids Characteristic of Lowland Reservoirs. River Res. Appl. 2016, 32, 1632–1641. [Google Scholar] [CrossRef]
- Turner Designs. CyanoFluor Handheld HAB Indicator User Manual; Turner Designs: San Jose, CA, USA, 14 March 2017; p. 19. [Google Scholar]
- Bertone, E.; Chuang, A.; Burford, M.A.; Hamilton, D.P. In-situ fluorescence monitoring of cyanobacteria: Laboratory-based quantification of species-specific measurement accuracy. Harmful Algae 2019, 87, 101625. [Google Scholar] [CrossRef] [PubMed]
- Courtois, S.; Steinmann, D.; Cajon, A.; van der Linden, L. Continuous monitoring of cyanobacterial blooms: Benefits and conditions for using fluorescence probes. Rev. Des Sci. Eau/J. Water Sci. 2017, 30, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Watras, C.J.; Morrison, K.A.; Rubsam, J.L.; Hanson, P.C.; Watras, A.J.; LaLiberte, G.D.; Milewski, P. A temperature compensation method for chlorophyll and phycocyanin fluorescence sensors in freshwater. Limnol Ocean.-Meth 2017, 15, 642–652. [Google Scholar] [CrossRef]
- Pachauri, R.K. Intergovernmental Panel on Climate Change (IPPC). Working Group Impacts. In Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2015. [Google Scholar]
- Wauthy, M.; Rautio, M.; Christoffersen, K.S.; Forsström, L.; Laurion, I.; Mariash, H.L.; Peura, S.; Vincent, W.F. Increasing dominance of terrigenous organic matter in circumpolar freshwaters due to permafrost thaw. Limnol. Oceanogr. Lett. 2018, 3, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.G.; DeSellas, A.M.; Fletcher, R.; Heintsch, L.; Morley, A.; Nakamoto, L.; Utsumi, K. Algal blooms in Ontario, Canada: Increases in reports since 1994. Lake Reserv. Manag. 2011, 27, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Qu, F.; Liang, H.; Wang, Z.; Wang, H.; Yu, H.; Li, G. Ultrafiltration membrane fouling by extracellular organic matters (EOM) of Microcystis aeruginosa in stationary phase: Influences of interfacial characteristics of foulants and fouling mechanisms. Water Res. 2012, 46, 1490–1500. [Google Scholar] [CrossRef]
- Zamyadi, A.; Coral, L.A.; Barbeau, B.; Dorner, S.; Lapolli, F.R.; Prévost, M. Fate of toxic cyanobacterial genera from natural bloom events during ozonation. Water Res. 2015, 73, 204–215. [Google Scholar] [CrossRef]
- Bowling, L.C.; Zamyadi, A.; Henderson, R.K. Assessment of in situ fluorometry to measure cyanobacterial presence in water bodies with diverse cyanobacterial populations. Water Res. 2016, 105, 22–33. [Google Scholar] [CrossRef]
- Genzoli, L.; Kann, J. Evaluation of Phycocyanin Probes as a Monitoring Tool for Toxigenic Cyanobacteria in the Klamath River below Iron Gate Dam; Aquatic Ecosystems LLC: The Woodlands, TX, USA, November 2016. [Google Scholar]
- Bastien, C.; Cardin, R.; Veilleux, E.; Deblois, C.; Warren, A.; Laurion, I. Performance evaluation of phycocyanin probes for the monitoring of cyanobacteria. J. Environ. Monit. 2011, 13, 110–118. [Google Scholar] [CrossRef]
- YSI Incorporated. EXO User Manual; YSI Incorporated: Yellow Springs, OH, USA, 2019. [Google Scholar]
- Armbruster, D.A.; Pry, T. Limit of blank, limit of detection and limit of quantitation. Clin. Biochem. Rev. 2008, 29 (Suppl. 1), S49–S52. [Google Scholar]
- Bowling, L.C.; Blais, S.; Sinotte, M. Heterogeneous spatial and temporal cyanobacterial distributions in Missisquoi Bay, Lake Champlain: An analysis of a 9 year data set. J. Great Lakes Res. 2015, 41, 164–179. [Google Scholar] [CrossRef]
- Stedmon, C.; Bro, R. Characterizing dissolved organic matter fluorescence with parallel factor analysis: A tutorial: Fluorescence-PARAFAC analysis of DOM. Limnol. Oceanogr. Methods 2008, 6, 572–579. [Google Scholar] [CrossRef]
- Kring, S.; Figary, S.E.; Boyer, G.; Watson, S.; Twiss, M. Rapid in situ measures of phytoplankton communities using the bbe FluoroProbe: Evaluation of spectral calibration, instrument intercompatibility, and performance range. Can. J. Fish. Aquat. Sci. 2014, 71, 1087–1095. [Google Scholar] [CrossRef]
- De Oliveira, F.G.; Bertone, E.; Stewart, A.R.; Awad, J.; Holland, A.; O’Halloran, K.; Bird, S. Multi-parameter compensation method for accurate in situ fluorescent dissolved organic matter monitoring and properties characterization. Water 2018, 10, 1146. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.M.; Lee, S.Y. Optical biosensors for the detection of pathogenic microorganisms. Trends Biotechnol. 2016, 34, 7–25. [Google Scholar] [CrossRef]
- Downing, B.D.; Pellerin, B.A.; Bergamaschi, B.A.; Saraceno, J.F.; Kraus, T.E.C. Seeing the light: The effects of particles, dissolved materials, and temperature on in situ measurements of DOM fluorescence in rivers and streams. Limnol. Oceanogr. Methods 2012, 10, 767–775. [Google Scholar] [CrossRef]
- So, P.T.C.; Dong, C.Y. Fluorescence Spectrophotometry; eLS: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Kong, Y.; Lou, I.; Zhang, Y.; Lou, C.U.; Mok, K.M. Using an online phycocyanin fluorescence probe for rapid monitoring of cyanobacteria in Macau freshwater reservoir. Hydrobiologia 2014, 741, 33–49. [Google Scholar] [CrossRef]
- Almuhtaram, H.; Cui, Y.; Zamyadi, A.; Hofmann, R. Cyanotoxins and cyanobacteria cell accumulations in drinking water treatment plants with a low risk of bloom formation at the source. Toxins 2018, 10, 430. [Google Scholar] [CrossRef] [Green Version]
- Zamyadi, A.; MacLeod, S.; Fan, Y.; McQuaid, N.; Dorner, S.; Sauvé, S.; Prévost, M. Toxic cyanobacterial breakthrough and accumulation in a drinking water plant: A monitoring and treatment challenge. Water Res. 2012, 46, 1511–1523. [Google Scholar] [CrossRef]
- Cotterill, V.; Hamilton, D.P.; Puddick, J.; Suren, A.; Wood, S.A. Phycocyanin sensors as an early warning system for cyanobacteria blooms concentrations: A case study in the Rotorua lakes. N. Z. J. Mar. Freshw. Res. 2019, 53, 555–570. [Google Scholar] [CrossRef]
- Song, K.; Li, L.; Tedesco, L.P.; Clercin, N.; Hall, R.; Li, S.; Shi, K.; Liu, D.; Sun, Y. Remote estimation of phycocyanin (PC) for inland waters coupled with YSI PC fluorescence probe. Environ. Sci. Pollut. Res. 2013, 20, 5330–5340. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; Henderson, R.K.; Stuetz, R.; Newcombe, G.; Newtown, K.; Gladman, B. Cyanobacterial management in full-scale water treatment and recycling processes: Reactive dosing following intensive monitoring. Environ. Sci. Water Res. Technol. 2016, 2, 362–375. [Google Scholar] [CrossRef]
- Ziegmann, M.; Abert, M.; Muller, M.; Frimmel, F.H. Use of fluorescence fingerprints for the estimation of bloom formation and toxin production of Microcystis aeruginosa. Water Res. 2010, 44, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Rousso, B.Z.; Bertone, E.; Stewart, R.; Aguiar, A.; Chuang, A.; Hamilton, D.P.; Burford, M.A. Chlorophyll and phycocyanin in-situ fluorescence in mixed cyanobacterial species assemblages: Effects of morphology, cell size and growth phase. Water Res. 2022, 212, 118127. [Google Scholar] [CrossRef]
- Kasinak, J.-M.E.; Holt, B.M.; Chislock, M.F.; Wilson, A.E. Benchtop fluorometry of phycocyanin as a rapid approach for estimating cyanobacterial biovolume. J. Plankton Res. 2015, 37, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Hodges, C.M. A Validation Study of Phycocyanin Sensors for Monitoring Cyanobacteria in Cultures and Field Samples. Master’s Thesis, University of Waikato, Hamilton, New Zealand, 2016. [Google Scholar]
- Macário, I.P.E.; Castro, B.B.; Nunes, M.I.S.; Antunes, S.C.; Pizarro, C.; Coelho, C.; Gonçalves, F.; de Figueiredo, D.R. New insights towards the establishment of phycocyanin concentration thresholds considering species-specific variability of bloom-forming cyanobacteria. Hydrobiologia 2015, 757, 155–165. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; WHO: London, UK; New York, NY, USA, 1999. [Google Scholar]
- Miller, T.R.; Beversdorf, L.; Chaston, S.D.; McMahon, K.D. Spatiotemporal molecular analysis of cyanobacteria blooms reveals Microcystis—Aphanizomenon interactions. PLoS ONE 2013, 8, e74933. [Google Scholar] [CrossRef]
- Donkor, V.; Häder, D.-P. Effects of ultraviolet irradiation on photosynthetic pigments in some filamentous cyanobacteria. Aquat. Microb. Ecol. 1996, 11, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Mulders, K.J.M.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic pigment production with microalgae: Biological constraints and opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef]
- Ma, R.; Fan, L.; Bi, Y.; Hu, Z. Effects of light intensity and quality on phycobiliprotein accumulation in the cyanobacterium Nostoc sphaeroides Kützing. Biotechnol. Lett. 2015, 37, 1663–1669. [Google Scholar] [CrossRef] [Green Version]
- Thimijan, R.; Heins, R. Photometric, radiometric, and quantum light units of measure: A review of procedures for interconversion. Hortic. Sci. 1983, 18, 818–822. [Google Scholar] [CrossRef]
- Dervaux, J.; Mejean, A.; Brunet, P. Irreversible Collective Migration of Cyanobacteria in Eutrophic Conditions. PLoS ONE 2015, 10, e0120906. [Google Scholar] [CrossRef] [PubMed]
- Aparicio Medrano, E.; Uittenbogaard, R.E.; van de Wiel, B.J.H.; Dionisio Pires, L.M.; Clercx, H.J.H. An alternative explanation for cyanobacterial scum formation and persistence by oxygenic photosynthesis. Harmful Algae 2016, 60, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Hu, C.; Visser, P.M.; Ma, R. Diurnal changes of cyanobacteria blooms in Taihu Lake as derived from GOCI observations. Limnol. Oceanogr. 2018, 63, 1711–1726. [Google Scholar] [CrossRef] [Green Version]
- Rome, M.; Beighley, R.E.; Faber, T. Sensor-based detection of algal blooms for public health advisories and long-term monitoring. Sci. Total Environ. 2021, 767, 144984. [Google Scholar] [CrossRef]
- Guo, D.; Robinson, C.; Herrera, J.E. Mechanism of dissolution of minium (Pb3O4) in water under depleting chlorine conditions. Corros. Sci. 2016, 103, 42–49. [Google Scholar] [CrossRef]
- Zhao, L.; Huang, W. Models for identifying significant environmental factors associated with cyanobacterial bloom occurrence and for predicting cyanobacterial blooms. J. Great Lakes Res. 2014, 40, 265–273. [Google Scholar] [CrossRef]
- Ndong, M.; Bird, D.; Nguyen-Quang, T.; de Boutray, M.L.; Zamyadi, A.; Vincon-Leite, B.; Lemaire, B.J.; Prevost, M.; Dorner, S. Estimating the risk of cyanobacterial occurrence using an index integrating meteorological factors: Application to drinking water production. Water Res. 2014, 56, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Newcombe, G.; House, J.; Ho, L.; Baker, P.; Burch, M. Management Strategies for Cyanobacteria (Blue-Green Algae): A Guide for Water Utilities; The Cooperative Research Centre for Water Quality and Treatment: Adelaïde, South Australia, June 2010; p. 112. [Google Scholar]
- Gao, J.; Zhu, J.; Wang, M.; Dong, W. Dominance and Growth Factors of Pseudanabaena sp. in Drinking Water Source Reservoirs, Southern China. Sustainability 2018, 10, 3936. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Units | Instruments | Data Value | Mean | Min | Max |
|---|---|---|---|---|---|---|
| Phycocyanin | RFU | YSI EXO2 probe | 2-h average | 1.1 | 0 | 38.2 |
| Chlorophyll-a | RFU | 1.3 | 0.2 | 4.5 | ||
| DO | mg/L | 5.8 | 0.5 | 14.5 | ||
| pH | 7.3 | 6.5 | 9.4 | |||
| TDS | mg/L | 91 | 84 | 106 | ||
| Temperature | °C | 25.7 | 22.9 | 28.2 | ||
| Turbidity | NTU | 7.3 | 1.6 | 63.6 | ||
| Conductivity | µS/cm | 141.8 | 132 | 165.5 | ||
| DOC | mg/L | Total organic carbon analyzer | Data collect on certain days | 6.9 | 5.3 | 7.9 |
| PAR | W/m2 | NASA POWER open source | Daily | 96.7 | 46.3 | 137.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Moradinejad, S.; Guerra Maldonado, J.F.; Zamyadi, A.; Dorner, S.; Prévost, M. Factors Affecting the Interpretation of Online Phycocyanin Fluorescence to Manage Cyanobacteria in Drinking Water Sources. Water 2022, 14, 3749. https://doi.org/10.3390/w14223749
Ma L, Moradinejad S, Guerra Maldonado JF, Zamyadi A, Dorner S, Prévost M. Factors Affecting the Interpretation of Online Phycocyanin Fluorescence to Manage Cyanobacteria in Drinking Water Sources. Water. 2022; 14(22):3749. https://doi.org/10.3390/w14223749
Chicago/Turabian StyleMa, Liya, Saber Moradinejad, Juan Francisco Guerra Maldonado, Arash Zamyadi, Sarah Dorner, and Michèle Prévost. 2022. "Factors Affecting the Interpretation of Online Phycocyanin Fluorescence to Manage Cyanobacteria in Drinking Water Sources" Water 14, no. 22: 3749. https://doi.org/10.3390/w14223749
APA StyleMa, L., Moradinejad, S., Guerra Maldonado, J. F., Zamyadi, A., Dorner, S., & Prévost, M. (2022). Factors Affecting the Interpretation of Online Phycocyanin Fluorescence to Manage Cyanobacteria in Drinking Water Sources. Water, 14(22), 3749. https://doi.org/10.3390/w14223749