
Revisiting Soil Water Potential: Towards a Better Understanding of Soil and Plant Interactions





Abstract
:1. Introduction
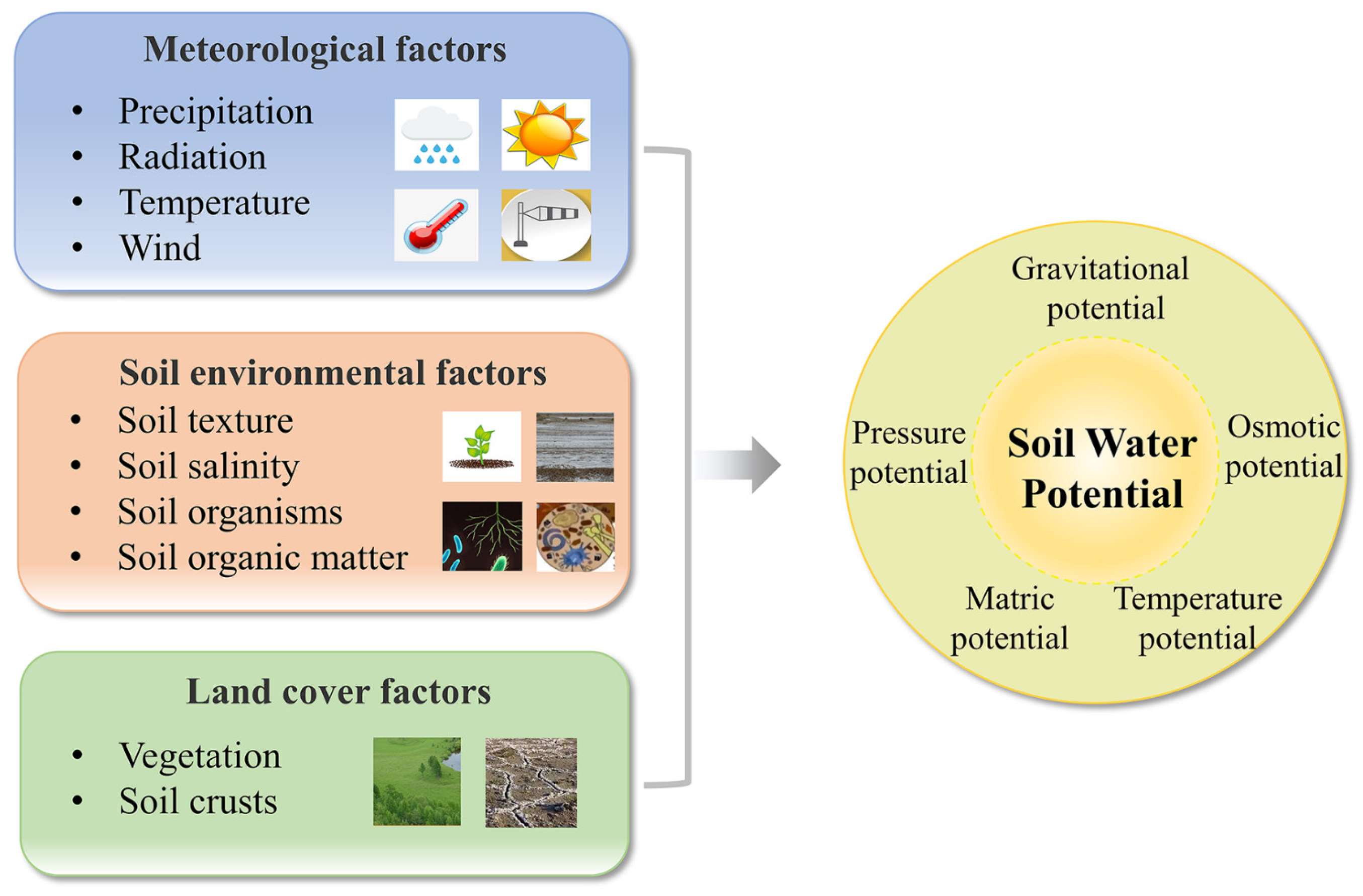
2. Influencing Factors and Evaluation Methods
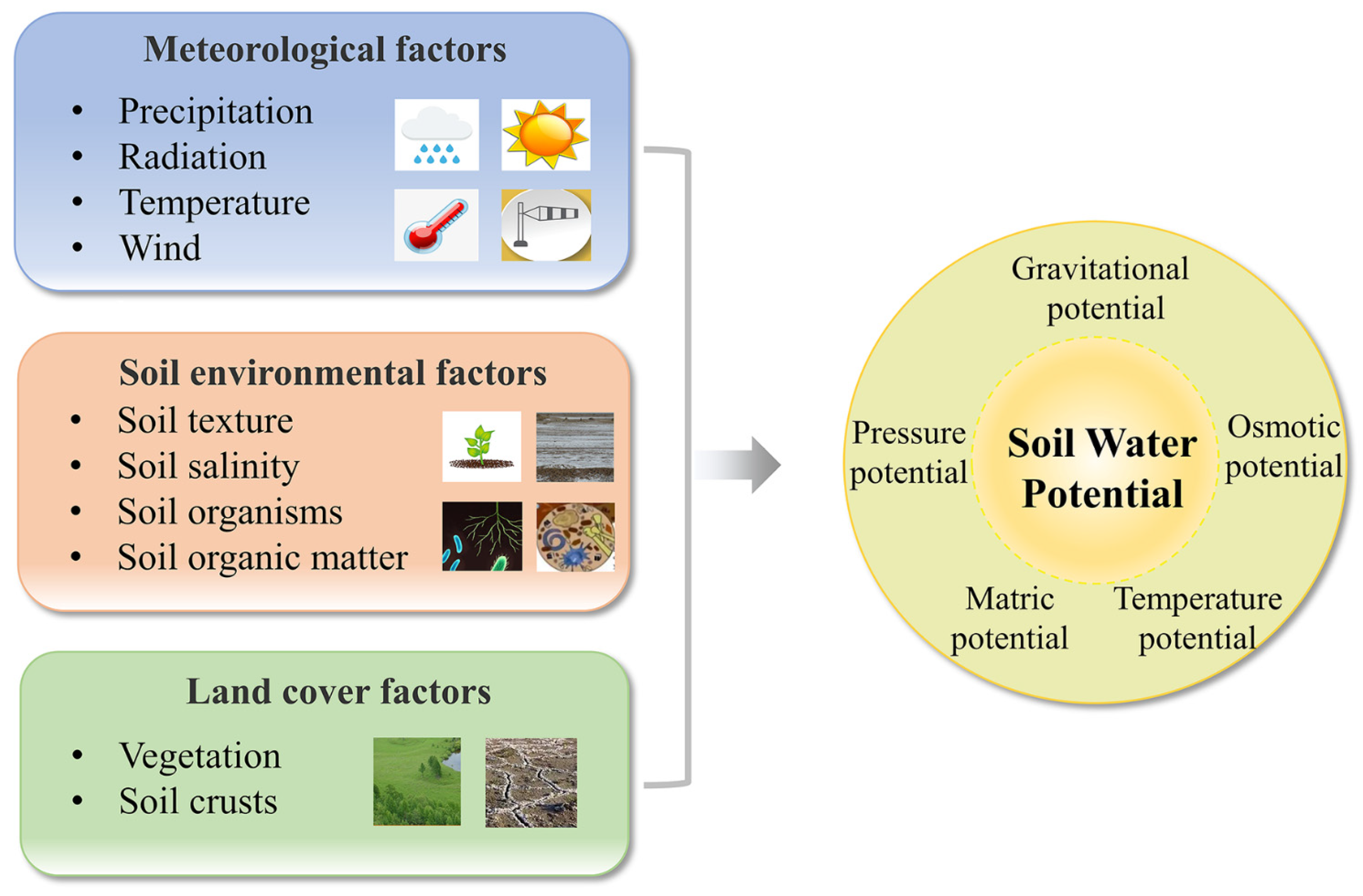
2.1. Influencing Factors
2.2. Evaluation Methods
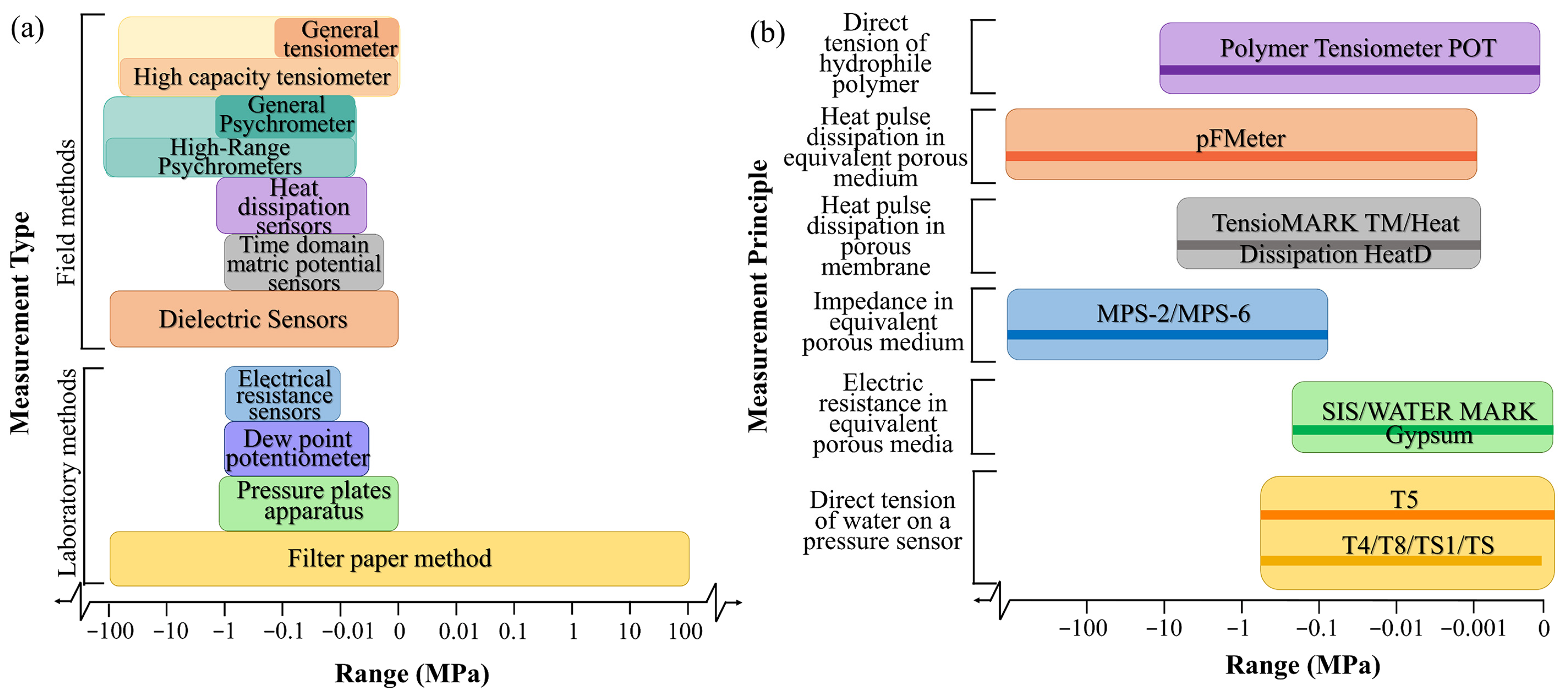
2.2.1. Measurement Methods
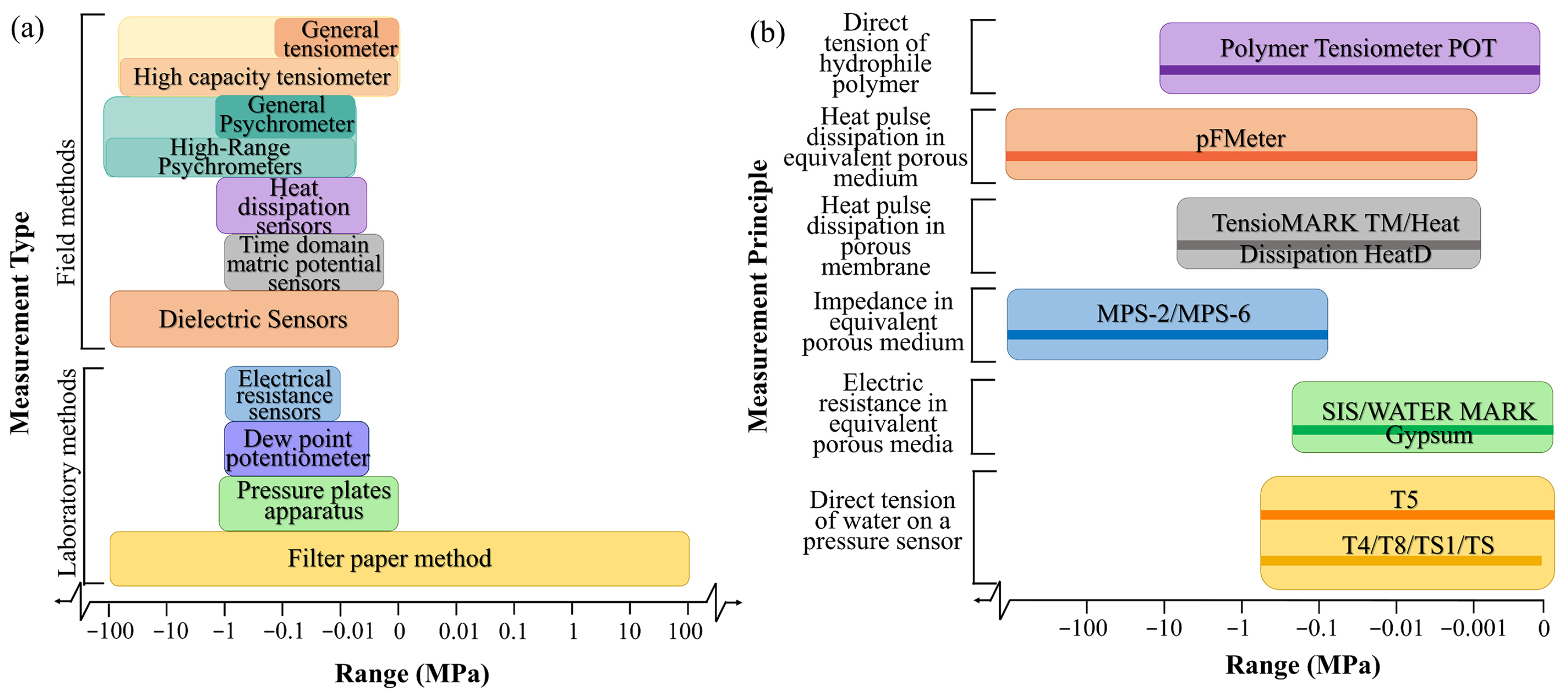

| Measurement | Device | Operational Range/MPa | In Situ Calibration | Measurement Principle | Main Features | References |
|---|---|---|---|---|---|---|
| Field methods | Tensiometer | −0.1~0 | Not required | Equilibrium of the liquid phase | Low range and long response time | [49] |
| Psychrometer | −1.5~−0.08 | Depends on the accuracy | Equilibrium of the vapor phase | Extremely sensitive to temperature | [50] | |
| Piezometer | Depends on the accuracy | Equilibrium of the liquid phase | Used in saturated material | [51,52] | ||
| Dielectric sensors | −100~0 | Depends on the accuracy | Dielectric capacity of the porous cup | Short response time; but subject to hysteresis | [43,44] | |
| Heat dissipation sensors | −1.5~−0.005 | Require separate calibration | Heat pulse dissipationin porousmembrane | Not sensitive to the salt content of the solution | [47] | |
| Frequency Domain and Time Domain Matric Potential Sensors | −1~−0.002 | Depends on the accuracy | Equilibrium of the liquid phase | Subject to hysteresis and very wet range | [53] | |
| Laboratory methods | Filter paper method | Entire range | Required | Equilibrium of the liquid phase | Long equilibration time | [45,46] |
| Pressure plate apparatus | −1.5~0 | Depends on the accuracy | Equilibrium of the liquid phase | Only used in the laboratory | [46,54] | |
| Electrical resistance sensors | −1~−0.01 | Depends on the accuracy | Electric resistancein equivalentporousmedium | Interface easily with data loggers; butsubject to hysteresis | [55] | |
| Dew point potentiometer | −1~−0.005 | Not required | Equilibrium of the vapor phase | Needs temperature control | [53] |
2.2.2. Simulation Methods
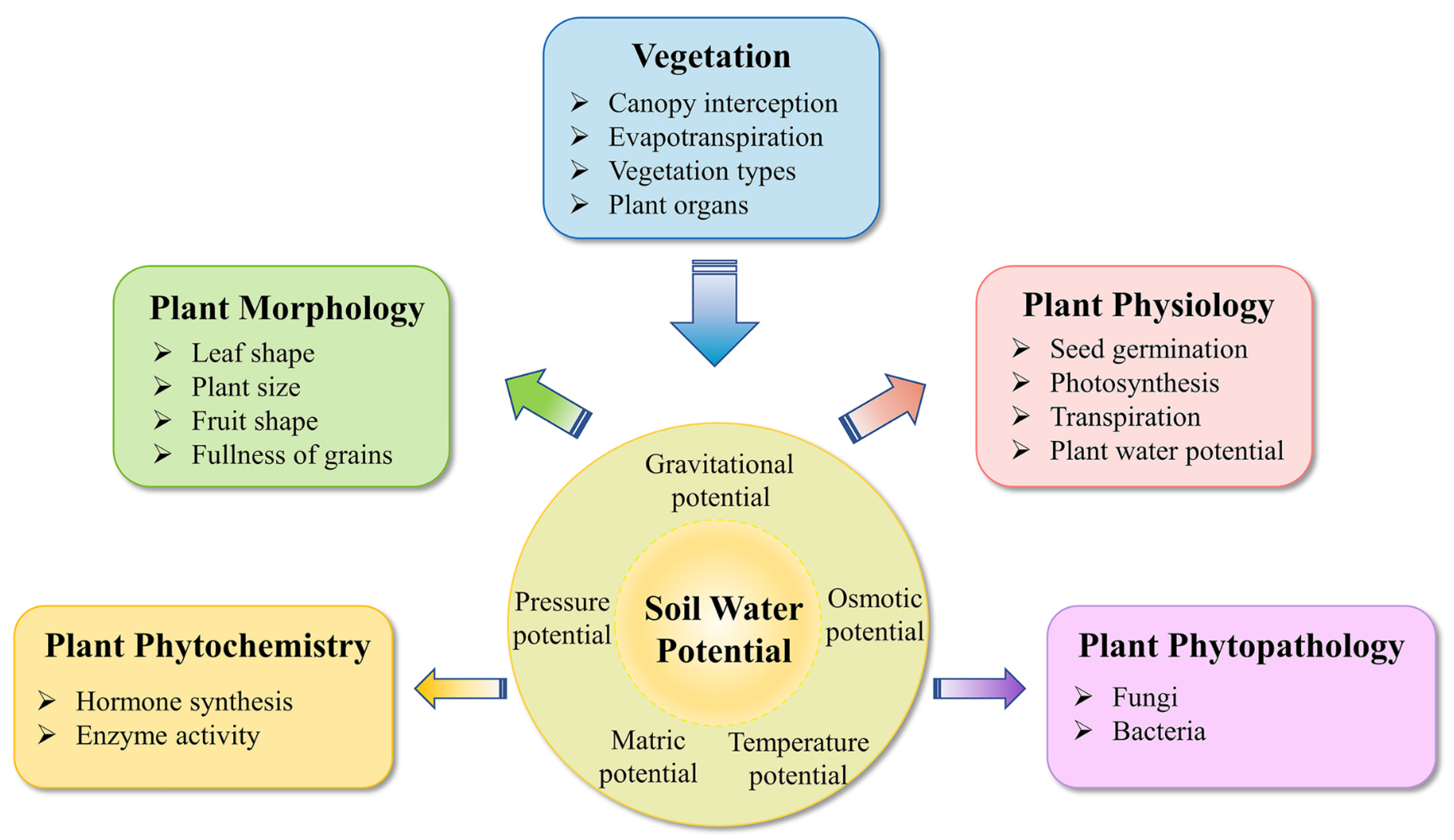
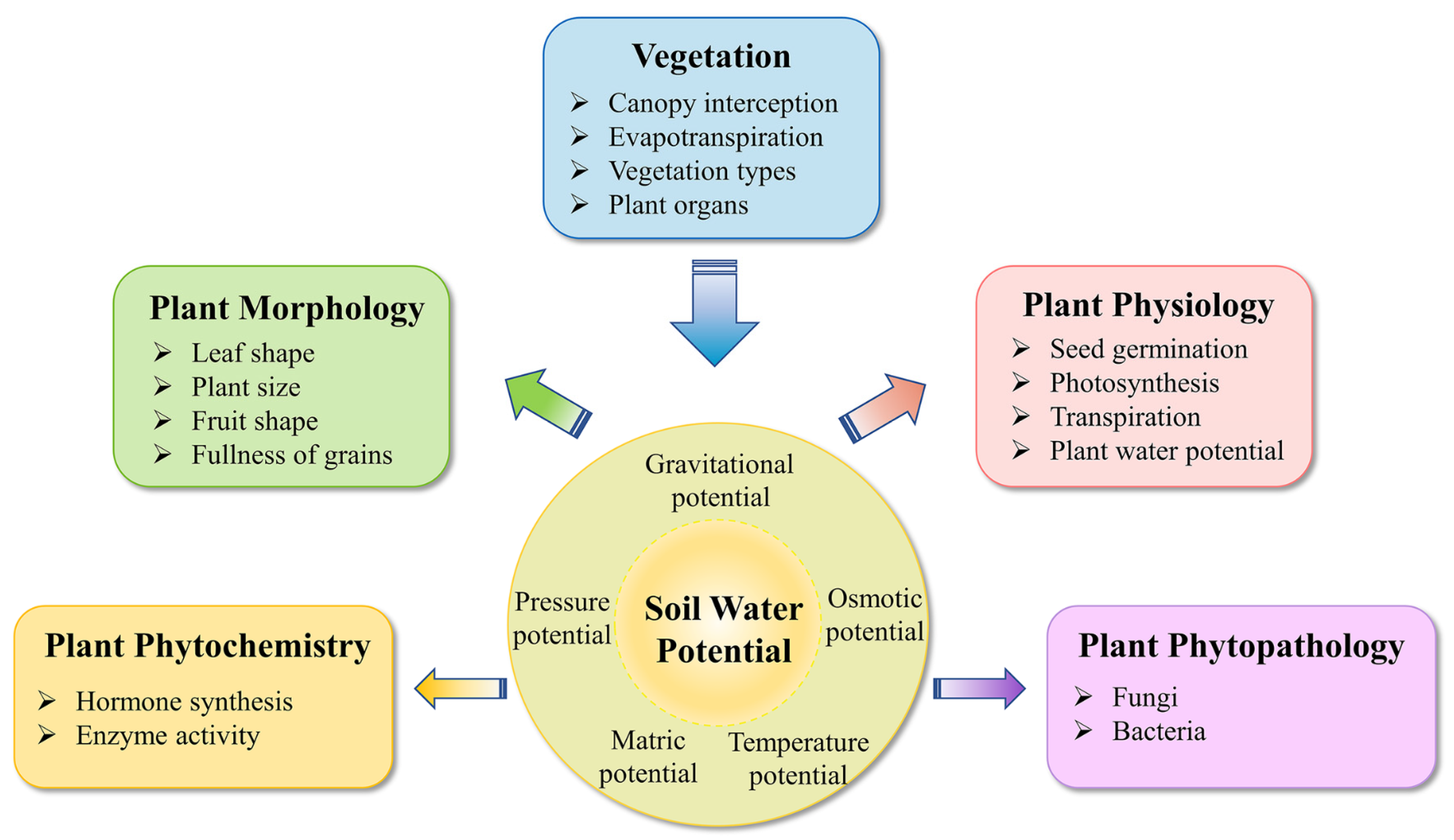
3. Plant Biological Responses to Varied SWP
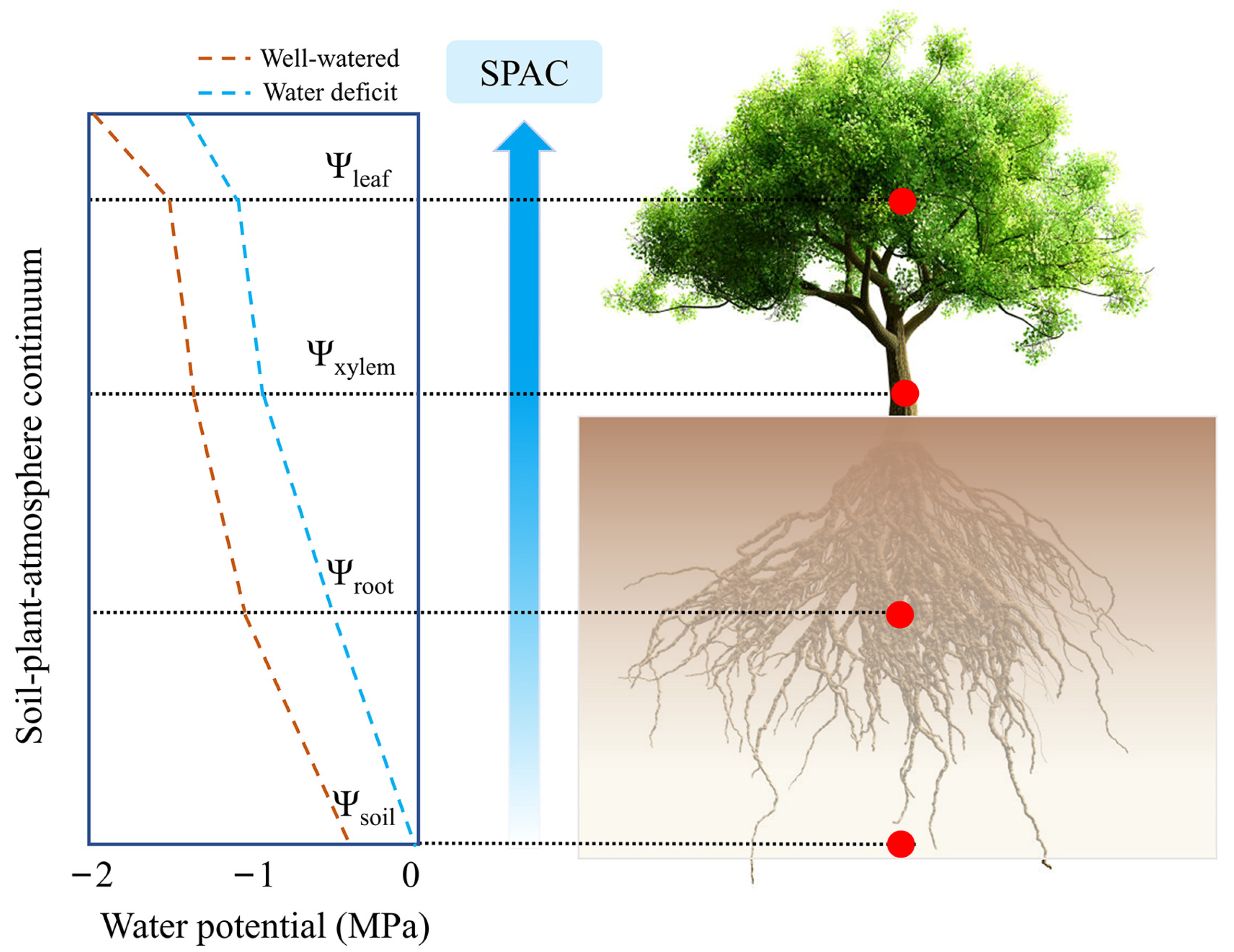
3.1. Physiology
3.2. Morphology
3.3. Phytochemistry and Phytopathology
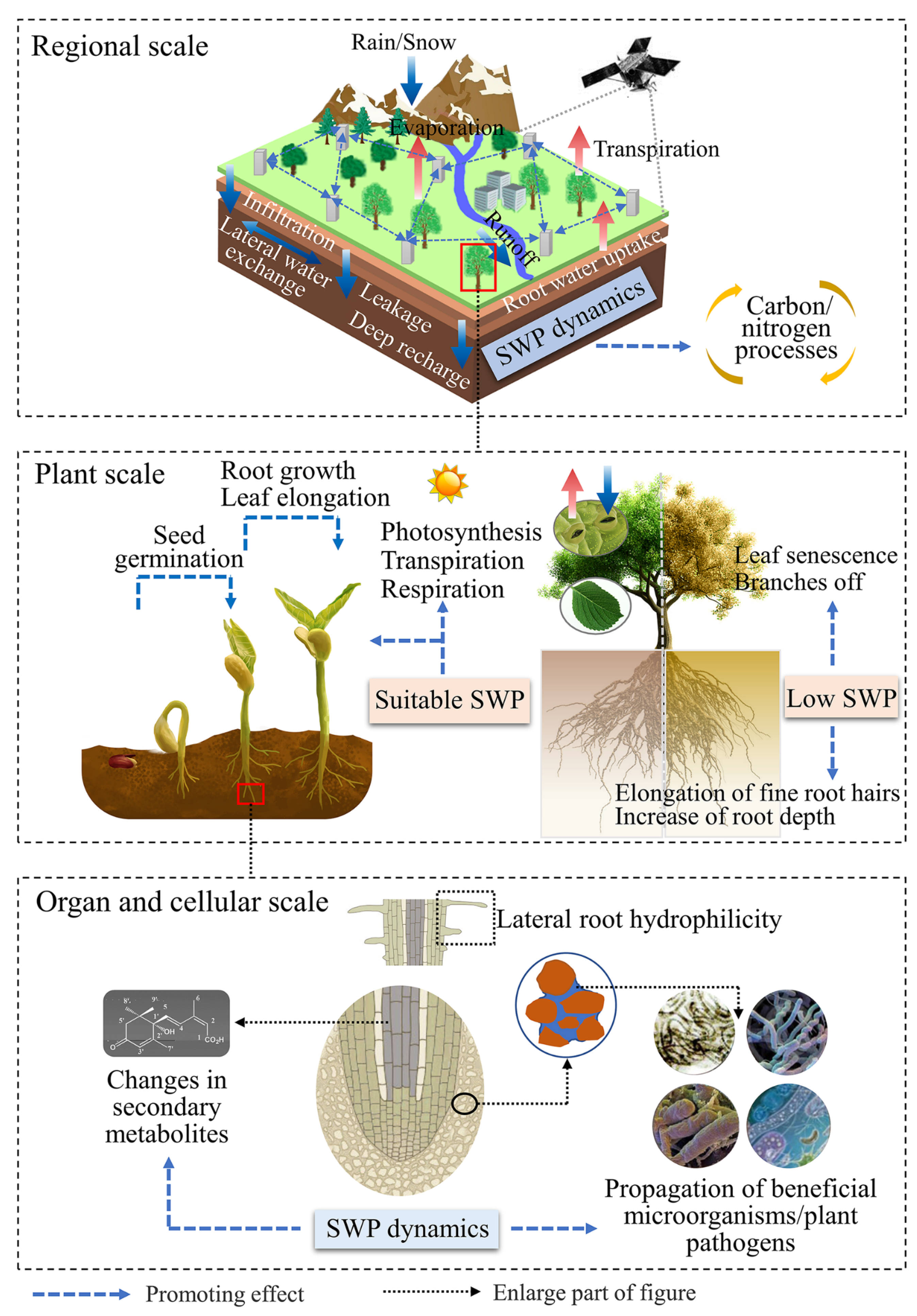
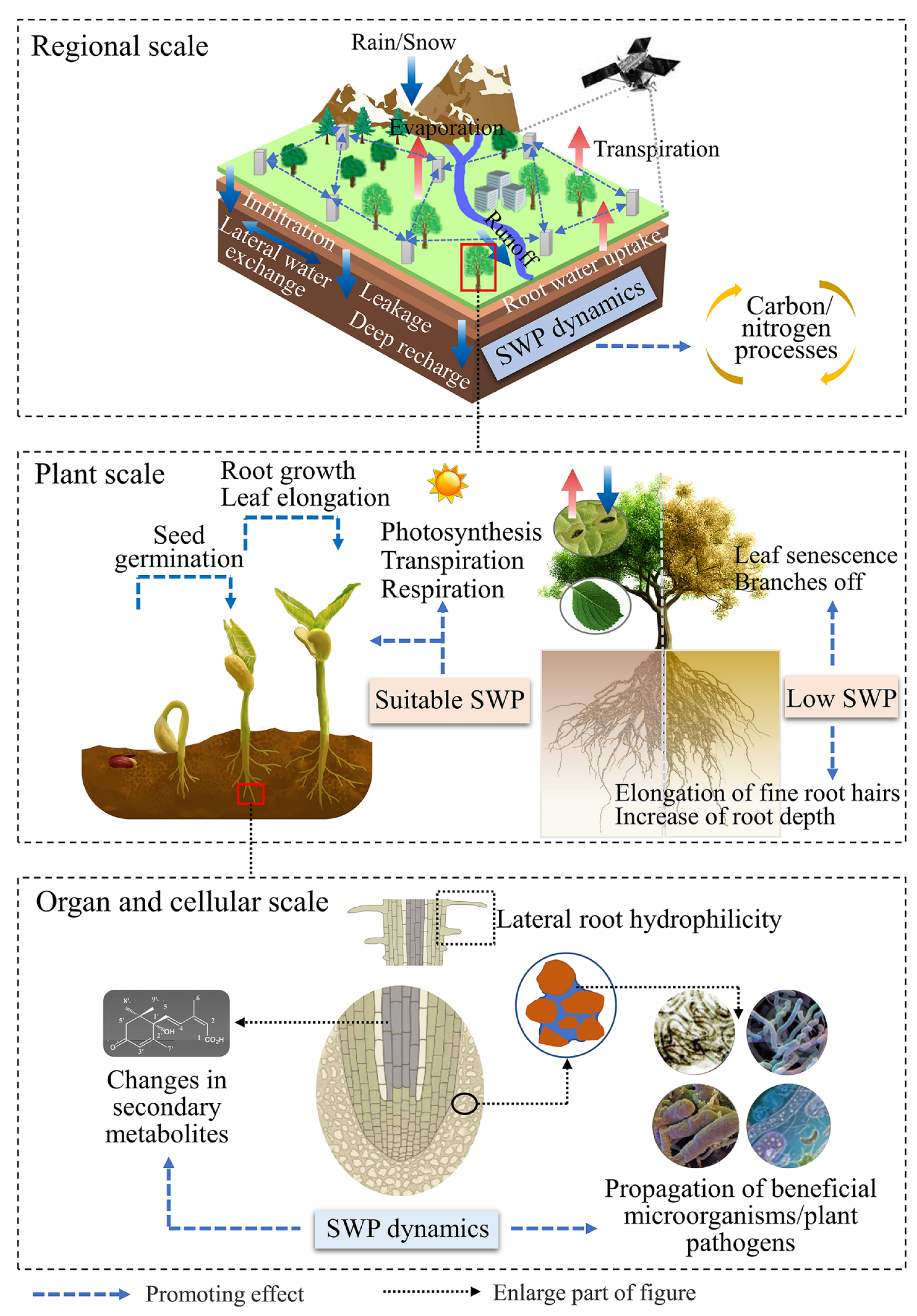
4. The Responses of Ecohydrological Processes to Varied SWP
4.1. Water Processes

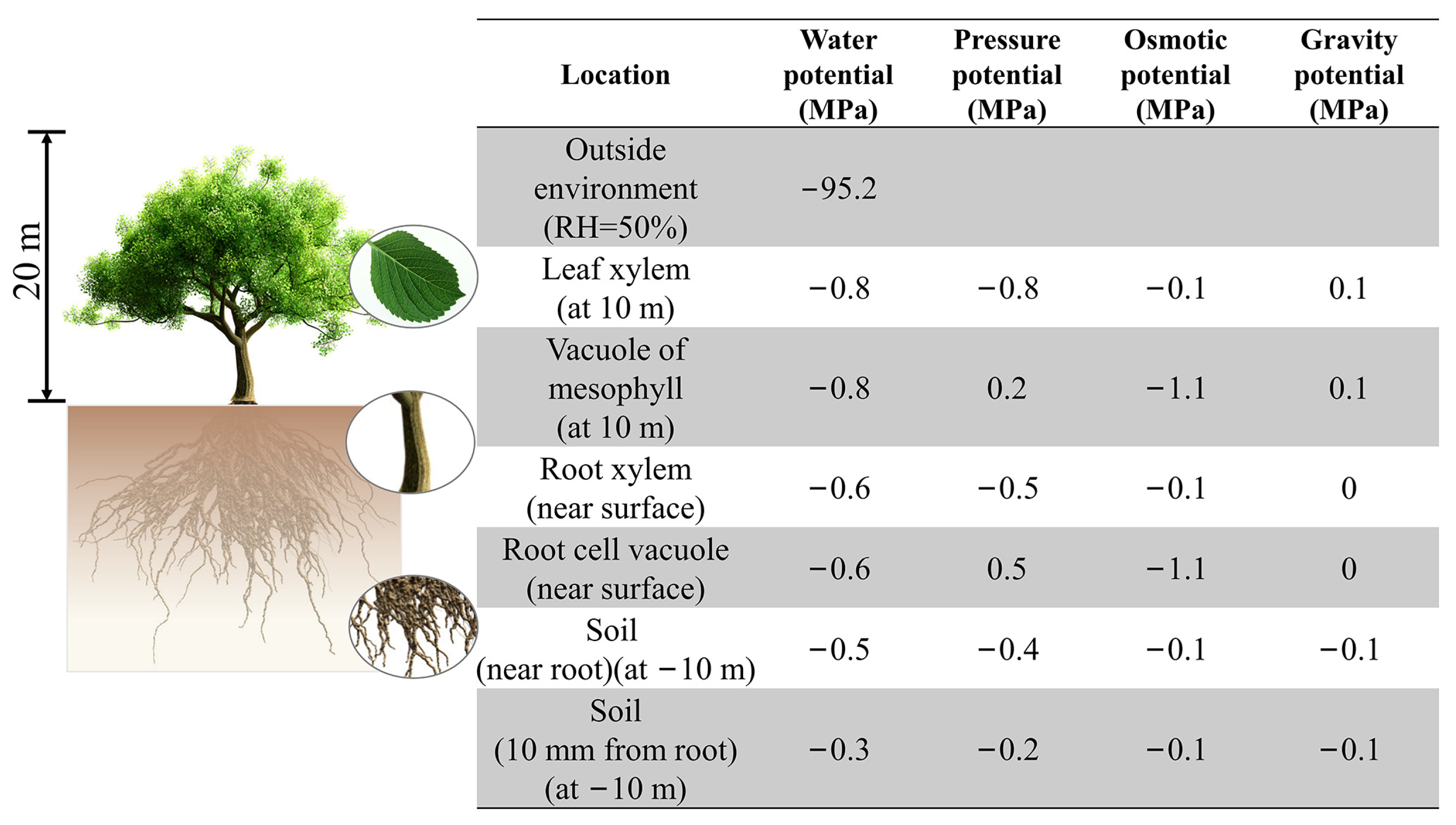
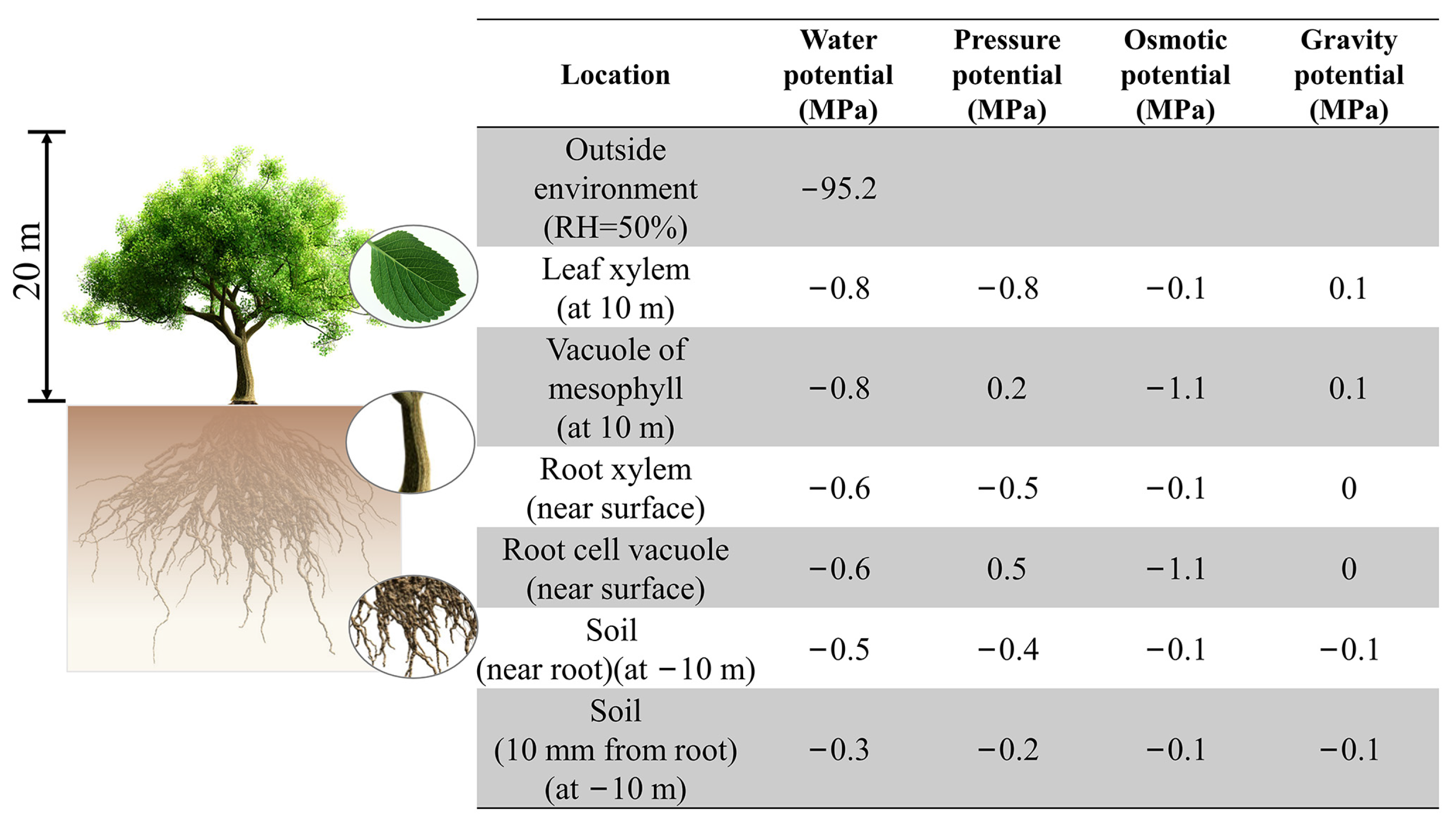{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.2. Carbon Processes
4.3. Nutrient Processes
5. Challenges and Insights for Future Research
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Papendick, R.I.; Campbell, G.S. Theory and measurement of water potential. In Water Potential Relations in Soil Microbiology; Parr, J.F., Gardner, W.R., Elliott, L.F., Eds.; Soil Science Society of America: Madison, WI, USA, 1981; Volume 9, pp. 1–22. [Google Scholar]
- Passioura, J.B. Water in the soil-plant-atmosphere continuum. In Physiological Plant Ecology II: Water Relations and Carbon Assimilation; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; Volume 12, pp. 5–33. [Google Scholar]
- Cowan, I. Transport of water in the soil-plant-atmosphere system. J. Appl. Ecol. 1965, 2, 221–239. [Google Scholar] [CrossRef]
- Hillel, D. Introduction to Soil Physics; Academic Press: San Diego, CA, USA, 1982. [Google Scholar]
- McKenzie, N.; Coughlan, K.; Cresswell, H. Soil Physical Measurement and Interpretation for Land Evaluation; CSIRO Publishing: Melbourne, Australia, 2002; Volume 5. [Google Scholar]
- Novick, K.A.; Ficklin, D.L.; Baldocchi, D.; Davis, K.J.; Ghezzehei, T.A.; Konings, A.G.; MacBean, N.; Raoult, N.; Scott, R.L.; Shi, Y.; et al. Confronting the water potential information gap. Nat. Geosci. 2022, 15, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Iwata, S.; Tabuchi, T.; Warkentin, B.P. Soil-Water Interactions: Mechanisms and Applications; CRC Press: Abingdon, UK, 2020. [Google Scholar]
- Bianchi, A.; Masseroni, D.; Thalheimer, M.; Medici, L.; Facchi, A. Field irrigation management through soil water potential measurements: A review. Ital. J. Agrometeorol. 2017, 22, 25–38. [Google Scholar] [CrossRef]
- Jackisch, C.; Germer, K.; Graeff, T.; Andrä, I.; Schulz, K.; Schiedung, M.; Haller-Jans, J.; Schneider, J.; Jaquemotte, J.; Helmer, P.; et al. Soil moisture and matric potential–an open field comparison of sensor systems. Earth Syst. Sci. Data 2020, 12, 683–697. [Google Scholar] [CrossRef] [Green Version]
- Philip, J. Fifty years progress in soil physics. Geoderma 1974, 12, 265–280. [Google Scholar] [CrossRef]
- Gardner, W.; Israelsen, O.; Edlefsen, N. The capillary potential function and its relation to irrigation practice. Phys. Rev. 1922, 20, 196. [Google Scholar]
- Richards, L.A. Capillary conduction of liquids through porous mediums. Physics 1931, 1, 318–333. [Google Scholar] [CrossRef]
- Klute, A. A numerical method for solving the flow equation for water in unsaturated materials. Soil Sci. 1952, 73, 105–116. [Google Scholar] [CrossRef]
- Jones, H.G. Estimation of an effective soil water potential at the root surface of transpiring plants. Plant Cell Environ. 1983, 6, 671–674. [Google Scholar] [CrossRef]
- Young, D.R.; Nobel, P.S. Predictions of soil-water potentials in the north-western Sonoran Desert. J. Ecol. 1986, 74, 143–154. [Google Scholar] [CrossRef]
- García-Tejera, O.; López-Bernal, Á.; Testi, L.; Villalobos, F.J. A soil-plant-atmosphere continuum (SPAC) model for simulating tree transpiration with a soil multi-compartment solution. Plant Soil 2016, 412, 215–233. [Google Scholar] [CrossRef]
- Brodribb, T.J.; McAdam, S.A.; Carins Murphy, M.R. Xylem and stomata, coordinated through time and space. Plant Cell Environ. 2017, 40, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Lu, N.; Zhang, C.; Likos, W. Soil water potential: A historical perspective and recent breakthroughs. Vadose Zone J. 2022, 21, e20203. [Google Scholar] [CrossRef]
- Campbell, G.S. Soil water potential measurement: An overview. Irrig. Sci. 1988, 9, 265–273. [Google Scholar] [CrossRef]
- Clark, G.A. Measurement of soil water potential. HortScience 1990, 25, 1548–1551. [Google Scholar] [CrossRef] [Green Version]
- Bittelli, M. Measuring soil water potential for water management in agriculture: A review. Sustainability 2010, 2, 1226–1251. [Google Scholar] [CrossRef] [Green Version]
- Malazian, A.; Hartsough, P.; Kamai, T.; Campbell, G.S.; Cobos, D.R.; Hopmans, J.W. Evaluation of MPS-1 soil water potential sensor. J. Hydrol. 2011, 402, 126–134. [Google Scholar] [CrossRef]
- Cassab, G.I.; Eapen, D.; Campos, M.E. Root hydrotropism: An update. Am. J. Bot. 2013, 100, 14–24. [Google Scholar] [CrossRef]
- Feng, W.; Lindner, H.; Robbins, N.E., 2nd; Dinneny, J.R. Growing out of stress: The role of cell- and organ-scale growth control in plant water-stress responses. Plant Cell 2016, 28, 1769–1782. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Prieto, I.; Armas, C.; Pugnaire, F.I. Water release through plant roots: New insights into its consequences at the plant and ecosystem level. New Phytol. 2012, 193, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Nadezhdina, N.; David, T.S.; David, J.S.; Ferreira, M.I.; Dohnal, M.; Tesař, M.; Gartner, K.; Leitgeb, E.; Nadezhdin, V.; Cermak, J.; et al. Trees never rest: The multiple facets of hydraulic redistribution. Ecohydrology 2010, 3, 431–444. [Google Scholar] [CrossRef]
- Johnson, M.S.; Couto, E.G.; Pinto, O.B., Jr.; Milesi, J.; Santos Amorim, R.S.; Messias, I.A.; Biudes, M.S. Soil CO2 dynamics in a tree island soil of the Pantanal: The role of soil water potential. PLoS ONE 2013, 8, e64874. [Google Scholar] [CrossRef] [PubMed]
- Serraj, R.; Sinclair, T.R.; Purcell, L.C. Symbiotic N2 fixation response to drought. J. Exp. Bot. 1999, 50, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Kroeckel, L.; Stolp, H. Influence of soil water potential on respiration and nitrogen fixation of Azotobacter vinelandii. Plant Soil 1984, 79, 37–49. [Google Scholar] [CrossRef]
- Nishida, K.; Shiozawa, S. Modeling and experimental determination of salt accumulation induced by root water uptake. Soil Sci. Soc. Am. J. 2010, 74, 774–786. [Google Scholar] [CrossRef]
- Lal, R.; Shukla, M.K. Principles of Soil Physics; CRC Press: New York, NY, USA, 2004. [Google Scholar]
- Saxton, K.E.; Rawls, W.J.; Romberger, J.S.; Papendick, R.I. Estimating generalized soil-water characteristics from texture. Soil Sci. Soc. Am. J. 1986, 50, 1031–1036. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, J.; Zhang, J. A study of the temperature effect on soil water potential. Acta Pedol. Sin. 1990, 27, 454–458. [Google Scholar]
- Sheldon, A.R.; Dalal, R.C.; Kirchhof, G.; Kopittke, P.M.; Menzies, N.W. The effect of salinity on plant-available water. Plant Soil 2017, 418, 477–491. [Google Scholar] [CrossRef]
- Bachmann, J.; Horton, R.; Grant, S.A.; Van der Ploeg, R. Temperature dependence of water retention curves for wettable and water-repellent soils. Soil Sci. Soc. Am. J. 2002, 66, 44–52. [Google Scholar] [CrossRef]
- Hohmann, M. Soil freezing—The concept of soil water potential. State of the art. Cold Reg. Sci. Technol. 1997, 25, 101–110. [Google Scholar] [CrossRef]
- Ernst, G.; Felten, D.; Vohland, M.; Emmerling, C. Impact of ecologically different earthworm species on soil water characteristics. Eur. J. Soil Biol. 2009, 45, 207–213. [Google Scholar] [CrossRef]
- Sun, F.; Xiao, B.; Li, S.; Kidron, G.J. Towards moss biocrust effects on surface soil water holding capacity: Soil water retention curve analysis and modeling. Geoderma 2021, 399, 115120. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015. [Google Scholar]
- Buckingham, E. Studies on the Movement of Soil Moisture; Govt. Print. Off.: Washington, DC, USA, 1907; Volume 38.
- Campbell, G.S.; Gardner, W.H. Psychrometric measurement of soil water potential: Temperature and bulk density effects. Soil Sci. Soc. Am. J. 1971, 35, 8–12. [Google Scholar] [CrossRef]
- Or, D.; Wraith, J.M. A new soil metric potential sensor based on time domain reflectometry. Water Resour. Res. 1999, 35, 3399–3407. [Google Scholar] [CrossRef]
- Novák, V.; Hlaváčiková, H. Soil-water potential and its measurement. In Applied Soil Hydrology; Theory and applications of transport in porous media; Springer: Cham, Switzerland, 2019; pp. 63–76. [Google Scholar]
- Gardner, R. A method of measuring the capillary tension of soil moisture over a wide moisture range. Soil Sci. 1937, 43, 277–284. [Google Scholar] [CrossRef]
- Fondjo, A.A.; Theron, E.; Ray, R.P. Assessment of various methods to measure the soil suction. Int. J. Eng. Technol. Explor. Eng. 2020, 9, 171–184. [Google Scholar] [CrossRef]
- Durner, W.; Or, D. Soil water potential measurement. In Encyclopedia of Hydrological Sciences; J. Wiley: New York, NY, USA, 2005. [Google Scholar]
- Vereecken, H.; Huisman, J.A.; Bogena, H.; Vanderborght, J.; Vrugt, J.A.; Hopmans, J.W. On the value of soil moisture measurements in vadose zone hydrology: A review. Water Resour. Res. 2008, 44, W00D06. [Google Scholar] [CrossRef] [Green Version]
- Klute, A.; Gardner, W. Tensiometer response time. Soil Sci. 1962, 93, 204–207. [Google Scholar] [CrossRef]
- Rawlins, S.L.; Campbell, G.S. Water potential: Thermocouple psychrometry. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods; American Society of Agronomy: Madison, WI, USA, 1986; Volume 5, pp. 597–618. [Google Scholar]
- Reeve, R. Water potential: Piezometry. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods; American Society of Agronomy: Madison, WI, USA, 1986; Volume 5, pp. 545–561. [Google Scholar]
- Warrick, A.W. Soil Physics Companion; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Scanlon, B.R.; Andraski, B.J.; Bilskie, J. 3.2.4 Miscellaneous methods for measuring matric or water potential. In Methods of Soil Analysis; Dane, J.H., Topp, G.C., Eds.; Soil Science Society of America: Madison, WI, USA, 2002; Volume 5, pp. 643–670. [Google Scholar]
- Richards, L.; Fireman, M. Pressure-plate apparatus for measuring moisture sorption and transmission by soils. Soil Sci. 1943, 56, 395–404. [Google Scholar] [CrossRef]
- Davis, D.; Hughes, J.E. A new approach to recording the wetting parameter by the use of electrical resistance sensors. Plant Dis. Rep. 1970, 54, 474–479. [Google Scholar]
- Toll, D.G.; Augarde, C.; Gallipoli, D.; Wheeler, S. Unsaturated Soils: Advances in Geo-Engineering; CRC Press: London, UK, 2008. [Google Scholar]
- Calbo, A.G. Dihedral Sensor for Evaluating Tension, Potential and Activity of Liquids. Patent No. 9588030; Patent and Trademark Office: Washington, DC, USA, 2017. [Google Scholar]
- Herron, P.M.; Gage, D.J.; Cardon, Z.G. Micro-scale water potential gradients visualized in soil around plant root tips using microbiosensors. Plant Cell Environ. 2010, 33, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Sabatier, J.M. Effects of soil water potential and moisture content on sound speed. Soil Sci. Soc. Am. J. 2009, 73, 1614–1625. [Google Scholar] [CrossRef]
- Fatichi, S.; Pappas, C.; Ivanov, V.Y. Modeling plant–water interactions: An ecohydrological overview from the cell to the global scale. Wiley Interdiscip. Rev. Water 2015, 3, 327–368. [Google Scholar] [CrossRef]
- Leong, E.C.; Rahardjo, H. Review of soil-water characteristic curve equations. J. Geotech. Geoenviron. 1997, 123, 1106–1117. [Google Scholar] [CrossRef] [Green Version]
- Philip, J.R.; De Vries, D.A. Moisture movement in porous materials under temperature gradients. Eos. Trans. Amer. Geophys. Union 1957, 38, 222–232. [Google Scholar] [CrossRef]
- Nimmo, J.R.; Miller, E.E. The temperature dependence of isothermal moisture vs. potential characteristics of soils. Soil Sci. Soc. Am. J. 1986, 50, 1105–1113. [Google Scholar] [CrossRef]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; LWW: Washington, DC, USA, 1954; Volume 78. [Google Scholar]
- Orchard, V.A.; Cook, F. Relationship between soil respiration and soil moisture. Soil Biol. Biochem. 1983, 15, 447–453. [Google Scholar] [CrossRef]
- Moyano, F.E.; Manzoni, S.; Chenu, C. Responses of soil heterotrophic respiration to moisture availability: An exploration of processes and models. Soil Biol. Biochem. 2013, 59, 72–85. [Google Scholar] [CrossRef]
- Chapman, N.; Miller, A.J.; Lindsey, K.; Whalley, W.R. Roots, water, and nutrient acquisition: Let’s get physical. Trends Plant Sci. 2012, 17, 701–710. [Google Scholar] [CrossRef]
- Bengough, A.G. Water dynamics of the root zone: Rhizosphere biophysics and its control on soil hydrology. Vadose Zone J. 2012, 11, vzj2011-0111. [Google Scholar] [CrossRef] [Green Version]
- Arnold, S.; Kailichova, Y.; Knauer, J.; Ruthsatz, A.D.; Baumgartl, T. Effects of soil water potential on germination of co-dominant Brigalow species: Implications for rehabilitation of water-limited ecosystems in the Brigalow Belt bioregion. Ecol. Eng. 2014, 70, 35–42. [Google Scholar] [CrossRef]
- Doneen, L.; MacGillivray, J.H. Germination (emergence) of vegetable seed as affected by different soil moisture conditions. Plant Physiol. 1943, 18, 524. [Google Scholar] [CrossRef] [PubMed]
- Gurvich, D.E.; Pérez-Sánchez, R.; Bauk, K.; Jurado, E.; Ferrero, M.C.; Funes, G.; Flores, J. Combined effect of water potential and temperature on seed germination and seedling development of cacti from a mesic Argentine ecosystem. Flora 2017, 227, 18–24. [Google Scholar] [CrossRef]
- Evans, C.E.; Etherington, J.R. The effect of soil water potential on seed germination of some British plants. New Phytol. 1990, 115, 539–548. [Google Scholar] [CrossRef]
- Gardarin, A.; Guillemin, J.P.; Munier-Jolain, N.M.; Colbach, N. Estimation of key parameters for weed population dynamics models: Base temperature and base water potential for germination. Eur. J. Agron. 2010, 32, 162–168. [Google Scholar] [CrossRef]
- Roberta, M.; Donato, L.; Stefano, B.; Maria Clara, Z.; Mario, M.; Giuseppe, Z. Temperature and water potential as parameters for modeling weed emergence in central-northern Italy. Weed Sci. 2010, 58, 216–222. [Google Scholar] [CrossRef]
- Dietrich, D. Hydrotropism: How roots search for water. J. Exp. Bot. 2018, 69, 2759–2771. [Google Scholar] [CrossRef]
- Cai, G.; Ahmed, M.A.; Abdalla, M.; Carminati, A. Root hydraulic phenotypes impacting water uptake in drying soils. Plant Cell Environ. 2022, 45, 650–663. [Google Scholar] [CrossRef]
- Takano, M.; Takahashi, H.; Hirasawa, T.; Suge, H. Hydrotropism in roots: Sensing of a gradient in water potential by the root cap. Planta 1995, 197, 410–413. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Xu, L.; Kiang, N. How plant functional-type, weather, seasonal drought, and soil physical properties alter water and energy fluxes of an oak–grass savanna and an annual grassland. Agric. For. Meteorol. 2004, 123, 13–39. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response mechanism of plants to drought stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Ahmed, M.; Stockle, C.O. Quantification of Climate Variability, Adaptation and Mitigation for Agricultural Sustainability; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Rodriguez-Dominguez, C.M.; Brodribb, T.J. Declining root water transport drives stomatal closure in olive under moderate water stress. New Phytol. 2020, 225, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Carminati, A.; Ahmed, M.A.; Zarebanadkouki, M.; Cai, G.; Goran, L.; Javaux, M. Stomatal closure prevents the drop in soil water potential around roots. New Phytol. 2020, 226, 1541–1543. [Google Scholar] [CrossRef] [Green Version]
- Flores, J.; Briones, O. Plant life-form and germination in a Mexican inter-tropical desert: Effects of soil water potential and temperature. J. Arid Environ. 2001, 47, 485–497. [Google Scholar] [CrossRef]
- Shrestha, A.; Roman, E.S.; Thomas, A.G.; Swanton, C.J. Modeling germination and shoot-radicle elongation of Ambrosia artemisiifolia. Weed Sci. 1999, 47, 557–562. [Google Scholar] [CrossRef]
- Dorsainvil, F.; Dürr, C.; Justes, E.; Carrera, A. Characterisation and modelling of white mustard (Sinapis alba L.) emergence under several sowing conditions. Eur. J. Agron. 2005, 23, 146–158. [Google Scholar] [CrossRef]
- Fyfield, T.; Gregory, P.J. Effects of temperature and water potential on germination, radicle elongation and emergence of mungbean. J. Exp. Bot. 1989, 40, 667–674. [Google Scholar] [CrossRef]
- Finch-Savage, W.; Phelps, K. Onion (Allium cepa L.) seedling emergence patterns can be explained by the influence of soil temperature and water potential on seed germination. J. Exp. Bot. 1993, 44, 407–414. [Google Scholar] [CrossRef]
- Rucker, K.; Kvien, C.; Holbrook, C.; Hook, J. Identification of peanut genotypes with improved drought avoidance traits. Peanut Sci. 1995, 22, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Liu, L.; Zhang, Y.; Sun, H.; Zhang, K.; Bai, Z.; Dong, H.; Li, C. Fine root and root hair morphology of cotton under drought stress revealed with RhizoPot. J. Agron. Crop Sci. 2020, 206, 679–693. [Google Scholar] [CrossRef]
- Ichsan, C.N. Morphological and physiological change of rice (Oryza sativa L.) under water stress at early season. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Banda Aceh, Indonesia, 21–22 September 2020; p. 012030. [Google Scholar]
- Rood, S.B.; Patiño, S.; Coombs, K.; Tyree, M.T. Branch sacrifice: Cavitation-associated drought adaptation of riparian cottonwoods. Trees 2000, 14, 248–257. [Google Scholar] [CrossRef]
- Jacobs, M.; Rais, A.; Pretzsch, H. How drought stress becomes visible upon detecting tree shape using terrestrial laser scanning (TLS). For. Ecol. Manag. 2021, 489, 118975. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental stress and secondary metabolites in plants. In Plant Metabolites and Regulation under Environmental Stress; Elsevier: Amsterdam, The Netherlands, 2018; pp. 153–167. [Google Scholar]
- Salem, N.; Msaada, K.; Dhifi, W.; Sriti, J.; Mejri, H.; Limam, F.; Marzouk, B. Effect of drought on safflower natural dyes and their biological activities. EXCLI J. 2014, 13, 1. [Google Scholar]
- Griffin, D.M. Soil water in the ecology of fungi. Annu. Rev. Phytopathol. 1969, 7, 289–310. [Google Scholar] [CrossRef]
- Lewis, B. Effects of water potential on the infection of potato tubers by Streptomyces scabies in soil. Ann. Appl. Biol. 1970, 66, 83–88. [Google Scholar] [CrossRef]
- Sharghi, A.; Badi, H.N.; Bolandnazar, S.; Mehrafarin, A.; Sarikhani, M.R. Morphophysiological and phytochemical responses of fenugreek to plant growth promoting rhizobacteria (PGPR) under different soil water levels. Folia Hortic. 2018, 30, 215–228. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.H.; Caldwell, M.M. Hydraulic lift: Substantial nocturnal water transport between soil layers by Artemisia tridentata roots. Oecologia 1987, 73, 486–489. [Google Scholar] [CrossRef]
- Horton, J.L.; Hart, S.C. Hydraulic lift: A potentially important ecosystem process. Trends Ecol. Evol. 1998, 13, 232–235. [Google Scholar] [CrossRef]
- Burgess, S.S.O.; Adams, M.A.; Turner, N.C.; Ong, C.K. The redistribution of soil water by tree root systems. Oecologia 1998, 115, 306–311. [Google Scholar] [CrossRef]
- Scharwies, J.D.; Dinneny, J.R. Water transport, perception, and response in plants. J. Plant Res. 2019, 132, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Boanares, D.; Oliveira, R.S.; Isaias, R.M.S.; Franca, M.G.C.; Penuelas, J. The neglected reverse water pathway: Atmosphere-plant-soil continuum. Trends Plant Sci. 2020, 25, 1073–1075. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Feng, Y.; Boersma, L. Partition of photosynthates between shoot and root in spring wheat (Triticum aestivum L.) as a function of soil water potential and root temperature. Plant Soil 1994, 164, 43–50. [Google Scholar] [CrossRef]
- Harris, R.F. Effect of water potential on microbial growth and activity. In Water Potential Relations in Soil Microbiology; Parr, J.F., Gardner, W.R., Elliott, L.F., Eds.; Soil Science Society of America: Madison, WI, USA, 1981; Volume 9, pp. 23–95. [Google Scholar]
- Borken, W.; Matzner, E. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob. Chang. Biol. 2009, 15, 808–824. [Google Scholar] [CrossRef]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecol. 2012, 93, 930–938. [Google Scholar] [CrossRef]
- Birch, H.F. The effect of soil drying on humus decomposition and nitrogen availability. Plant Soil 1958, 10, 9–31. [Google Scholar] [CrossRef]
- Manzoni, S.; Chakrawal, A.; Fischer, T.; Schimel, J.P.; Porporato, A.; Vico, G. Rainfall intensification increases the contribution of rewetting pulses to soil respiration. Biogeosciences 2020, 17, 4007–4023. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Augustine, D.J.; Brewer, P.; von Fischer, J.C. Nitrogen cycling and water pulses in semiarid grasslands: Are microbial and plant processes temporally asynchronous? Oecologia 2012, 170, 799–808. [Google Scholar] [CrossRef]
- Plante, A.F. Soil biogeochemical cycling of inorganic nutrients and metals. In Soil Microbiology, Ecology and Biochemistry; Elsevier: New York, NY, USA, 2007; pp. 389–432. [Google Scholar]
- Sinegani, A.A.S.; Mahohi, A. Temporal variability of available P, microbial P and some phosphomonoesterase activities in a sewage sludge treated soil: The effect of soil water potential. Afr. J. Biotechnol. 2009, 8, 6888–6895. [Google Scholar]
- Wells, J.M.; Thomas, J.; Boddy, L. Soil water potential shifts: Developmental responses and dependence on phosphorus translocation by the saprotrophic, cord-forming basidiomycete Phanerochaete velutina. Mycol. Res. 2001, 105, 859–867. [Google Scholar] [CrossRef]
- Grierson, P.F.; Comerford, N.B.; Jokela, E.J. Phosphorus mineralization and microbial biomass in a Florida Spodosol: Effects of water potential, temperature and fertilizer application. Biol. Fertil. Soils 1999, 28, 244–252. [Google Scholar] [CrossRef]
- Couvreur, V.; Vanderborght, J.; Beff, L.; Javaux, M. Horizontal soil water potential heterogeneity: Simplifying approaches for crop water dynamics models. Hydrol. Earth Syst. Sci. 2014, 18, 1723–1743. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.; Li, S.; Yu, X.; Yang, P.; Yu, G.; Feng, R.; Zhuang, X. Chinese ecosystem research network: Progress and perspectives. Ecol. Complex. 2010, 7, 225–233. [Google Scholar] [CrossRef]
- Brantley, S.L.; McDowell, W.H.; Dietrich, W.E.; White, T.S.; Kumar, P.; Anderson, S.P.; Chorover, J.; Lohse, K.A.; Bales, R.C.; Richter, D.D. Designing a network of critical zone observatories to explore the living skin of the terrestrial Earth. Earth Surf. Dyn. 2017, 5, 841–860. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, F.Y.; IEEE. Towards blockchain-based intelligent transportation systems. In Proceedings of the 2016 IEEE 19th international conference on intelligent transportation systems (ITSC), Rio de Janeiro, Brazil, 1–4 November 2016; pp. 2663–2668. [Google Scholar]
- Vereecken, H.; Amelung, W.; Bauke, S.L.; Bogena, H.; Brüggemann, N.; Montzka, C.; Vanderborght, J.; Bechtold, M.; Blöschl, G.; Carminati, A.; et al. Soil hydrology in the earth system. Nat. Rev. Earth Environ. 2022, 3, 573–587. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Liu, H.; Yu, Y.; Guo, L.; Zhao, W.; Yetemen, O. Revisiting Soil Water Potential: Towards a Better Understanding of Soil and Plant Interactions. Water 2022, 14, 3721. https://doi.org/10.3390/w14223721
Ma Y, Liu H, Yu Y, Guo L, Zhao W, Yetemen O. Revisiting Soil Water Potential: Towards a Better Understanding of Soil and Plant Interactions. Water. 2022; 14(22):3721. https://doi.org/10.3390/w14223721
Chicago/Turabian StyleMa, Yuanyuan, Hu Liu, Yang Yu, Li Guo, Wenzhi Zhao, and Omer Yetemen. 2022. "Revisiting Soil Water Potential: Towards a Better Understanding of Soil and Plant Interactions" Water 14, no. 22: 3721. https://doi.org/10.3390/w14223721
APA StyleMa, Y., Liu, H., Yu, Y., Guo, L., Zhao, W., & Yetemen, O. (2022). Revisiting Soil Water Potential: Towards a Better Understanding of Soil and Plant Interactions. Water, 14(22), 3721. https://doi.org/10.3390/w14223721