
Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Ovary Histology
2.3. RNA Isolation, Library Construction and Illumine Sequencing
2.4. Basic Analysis of Sequencing Data
2.5. Differential Expression Genes (DEGs) Analysis and Enrichment Analysis
2.6. RNA-Seq Data Validation by Real–Time Quantitative PCR
3. Results
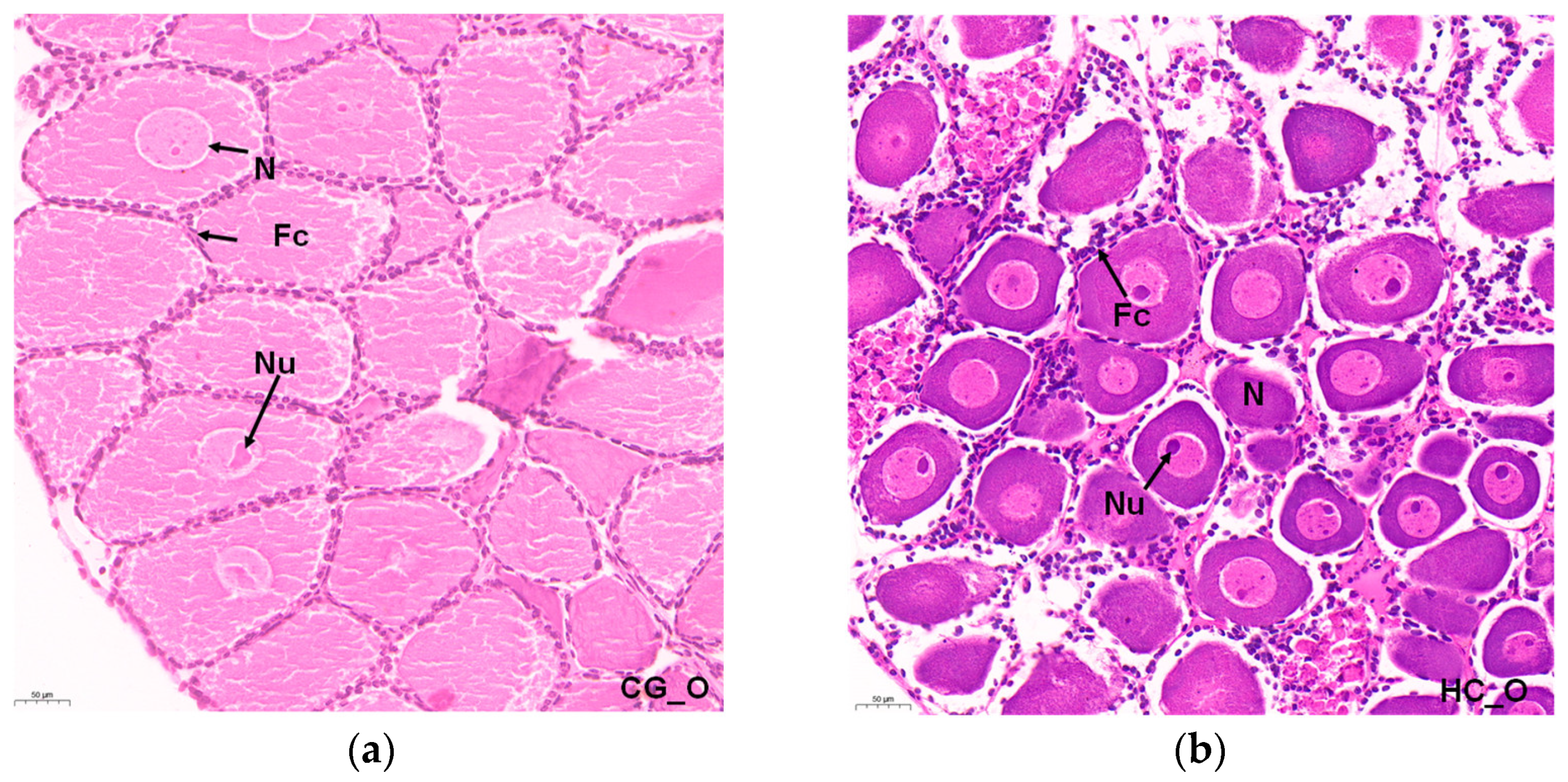
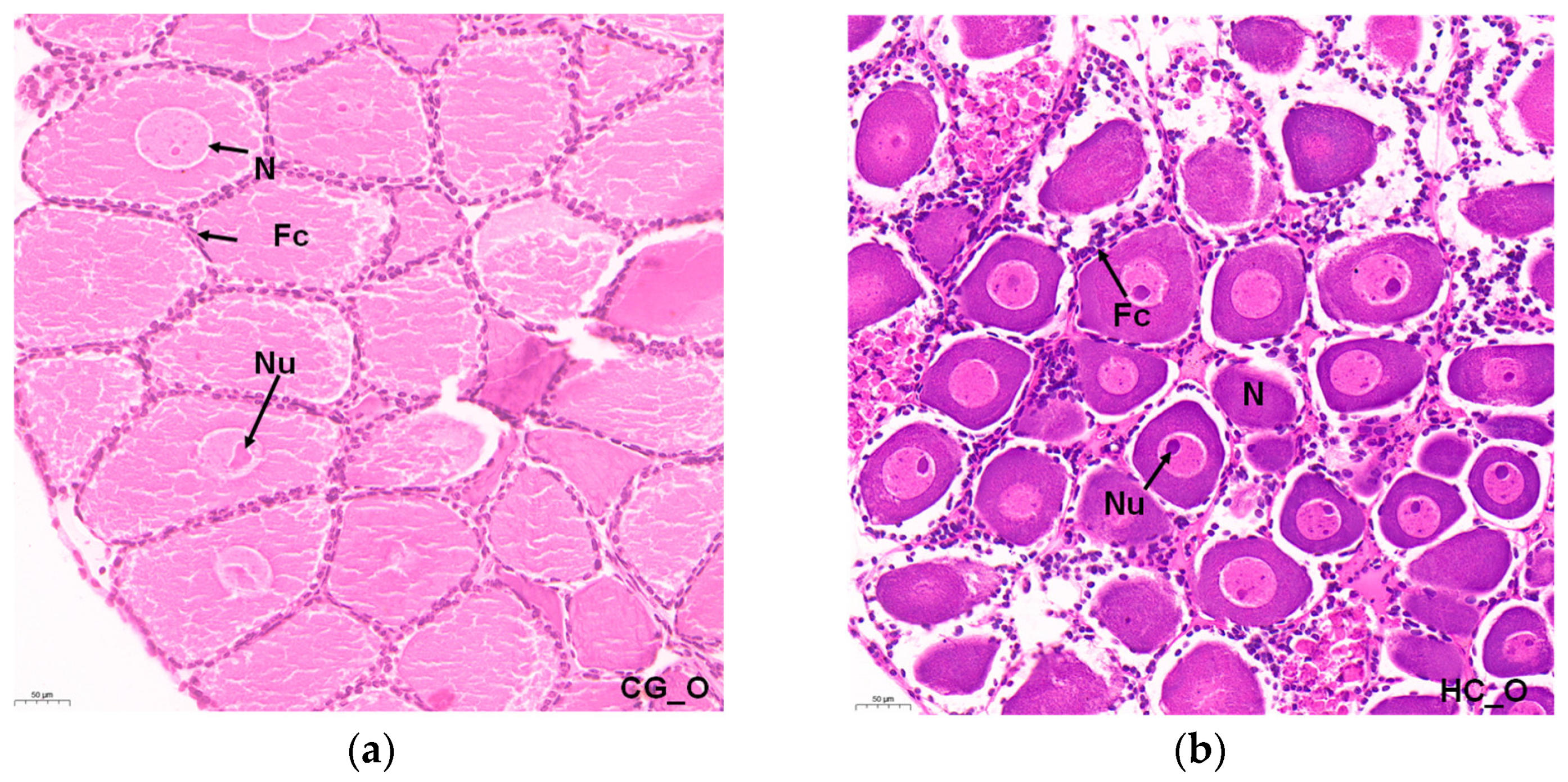
3.1. Histopathology of Ovary
3.2. Summary of the RNA–Sequencing Data
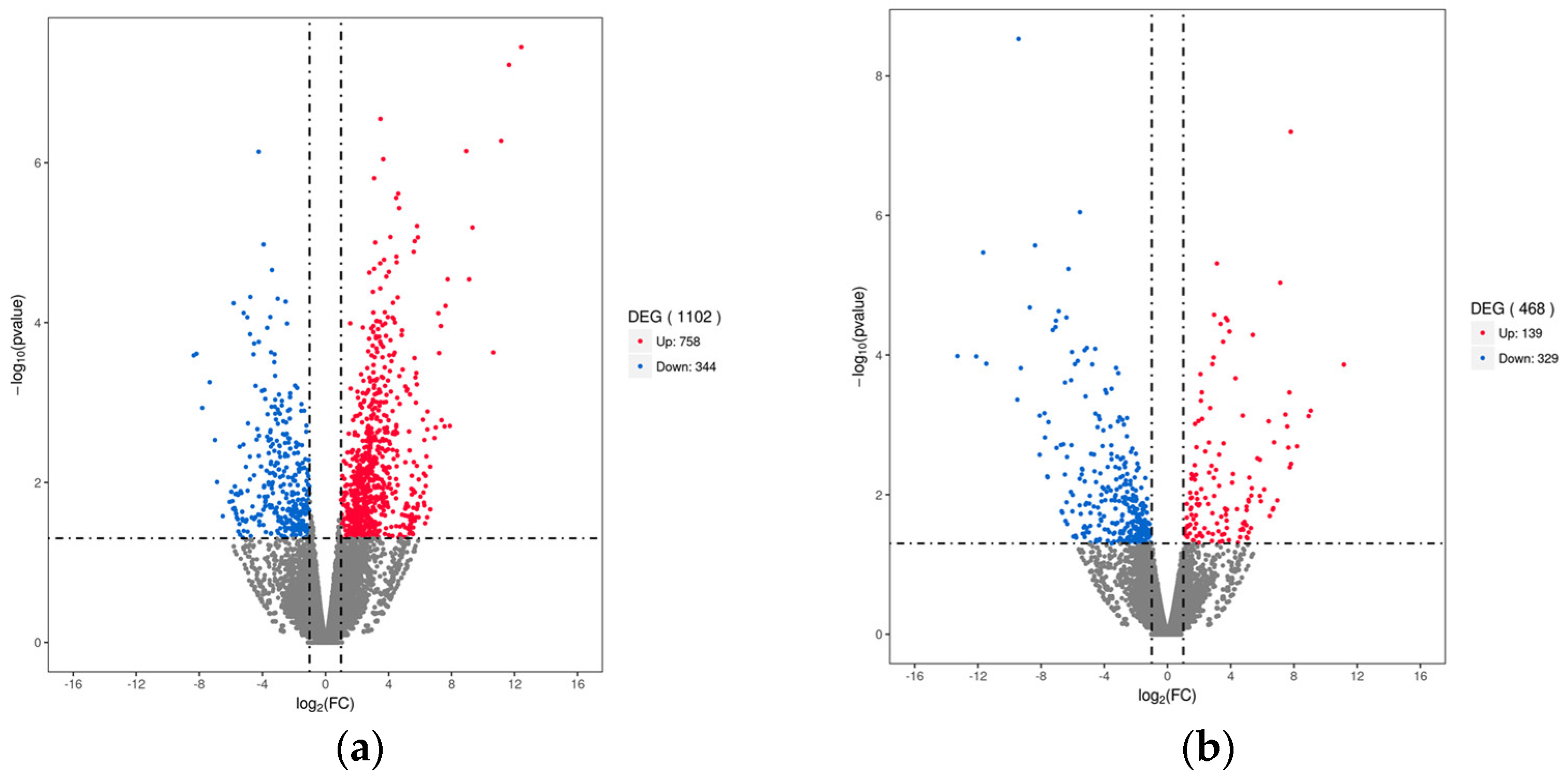
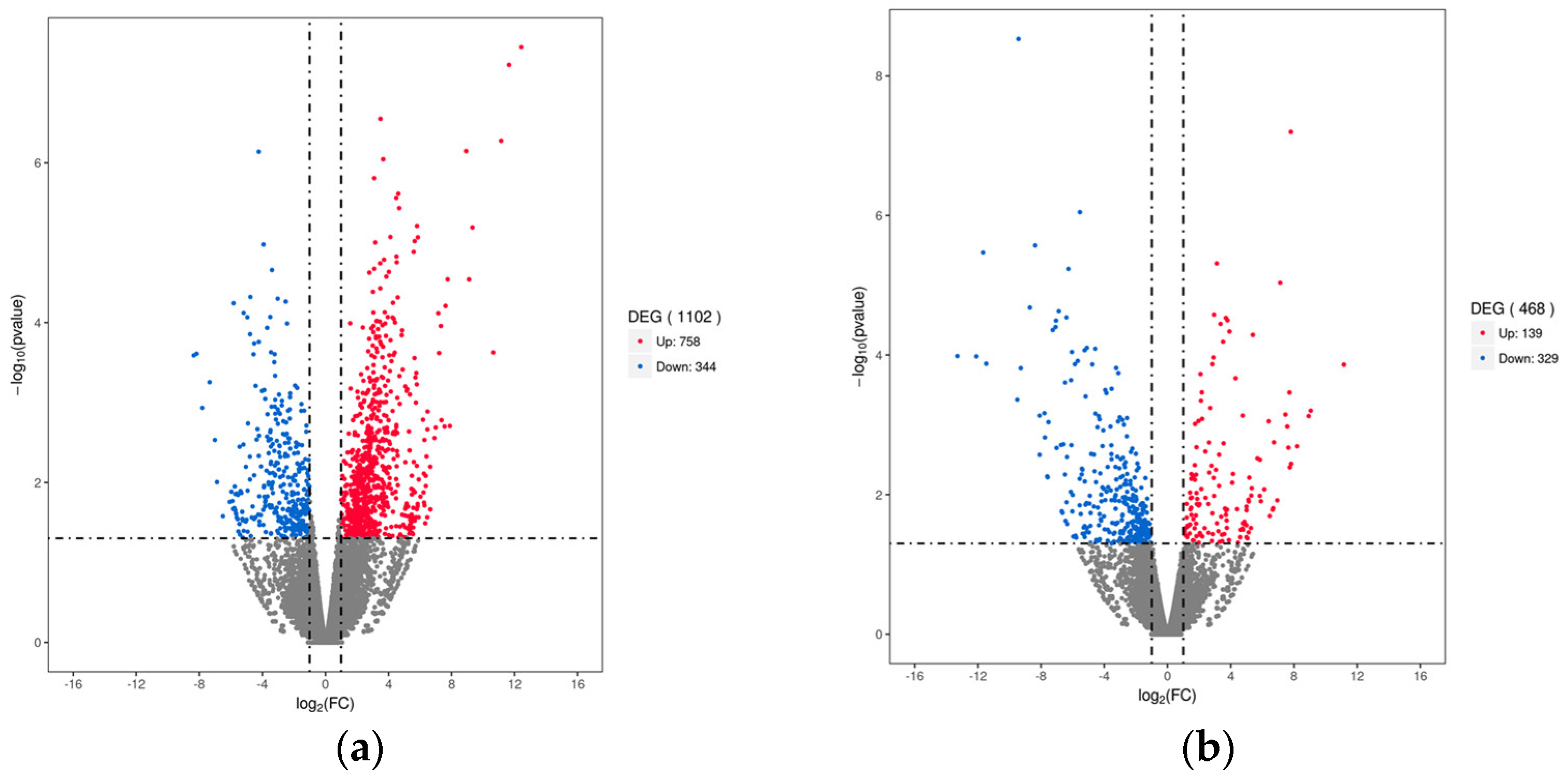
3.3. Differential Gene Expression (DEGs) Analysis
3.4. Gene Ontology (GO)Analysis of Significant DEGs
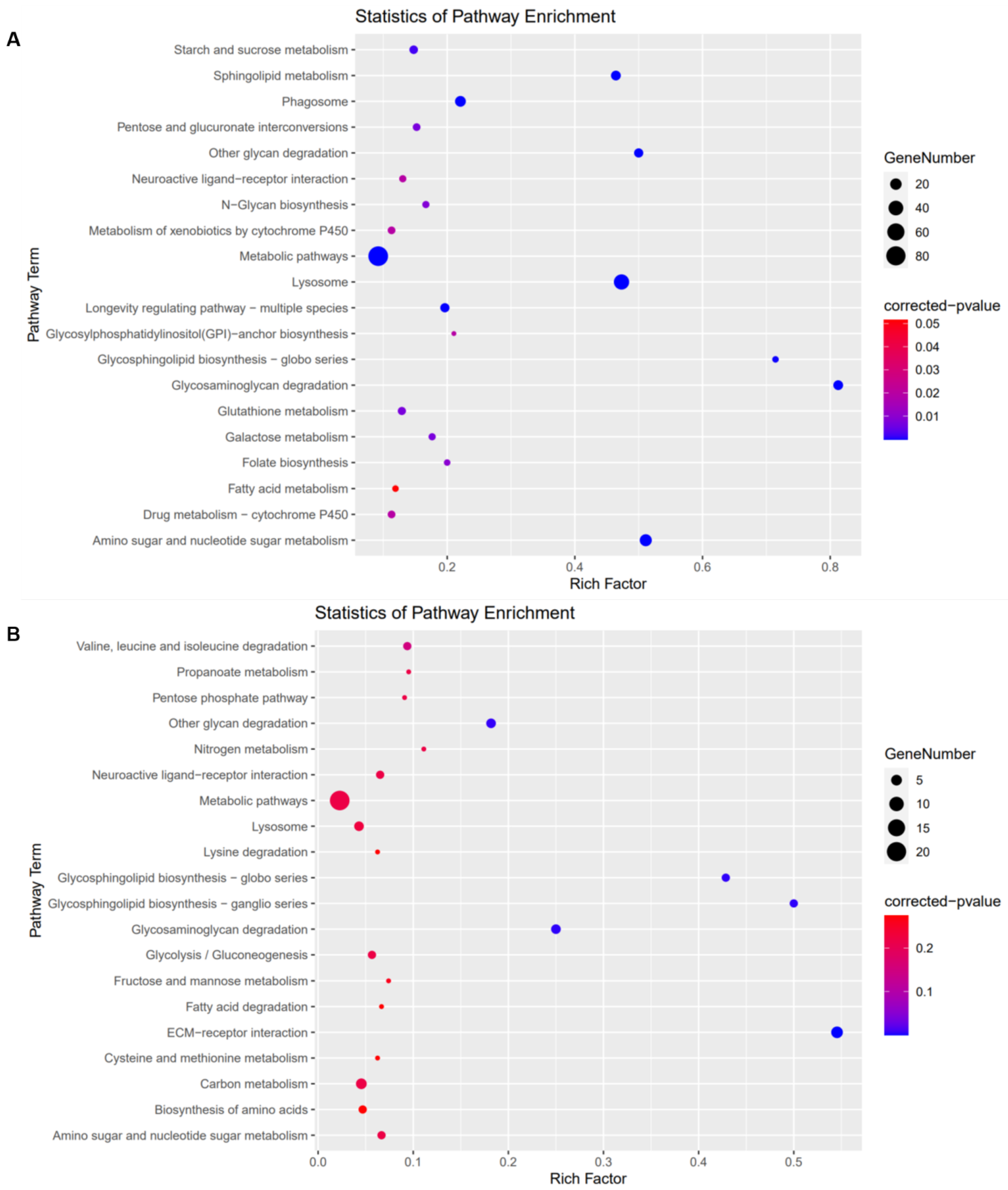
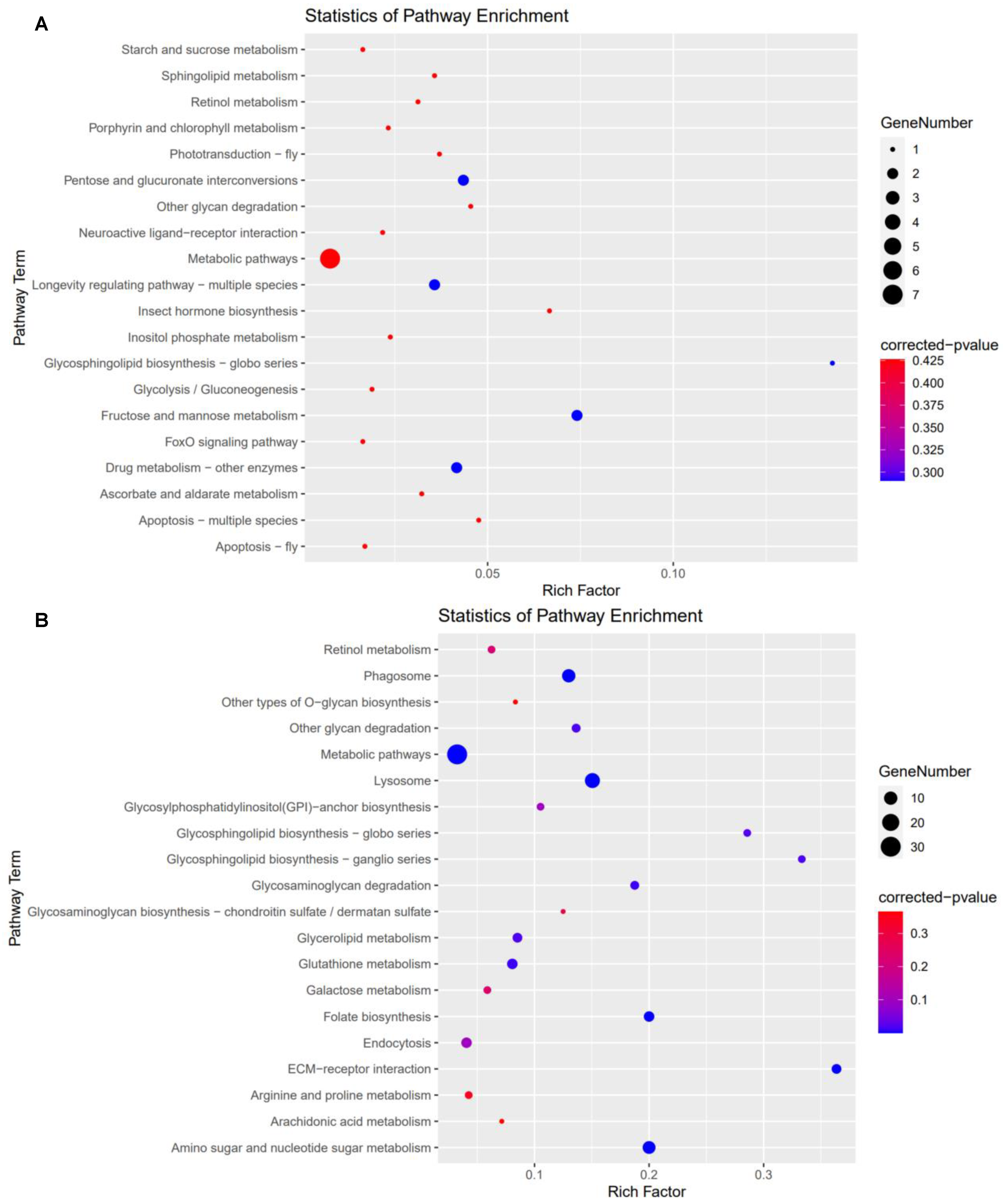
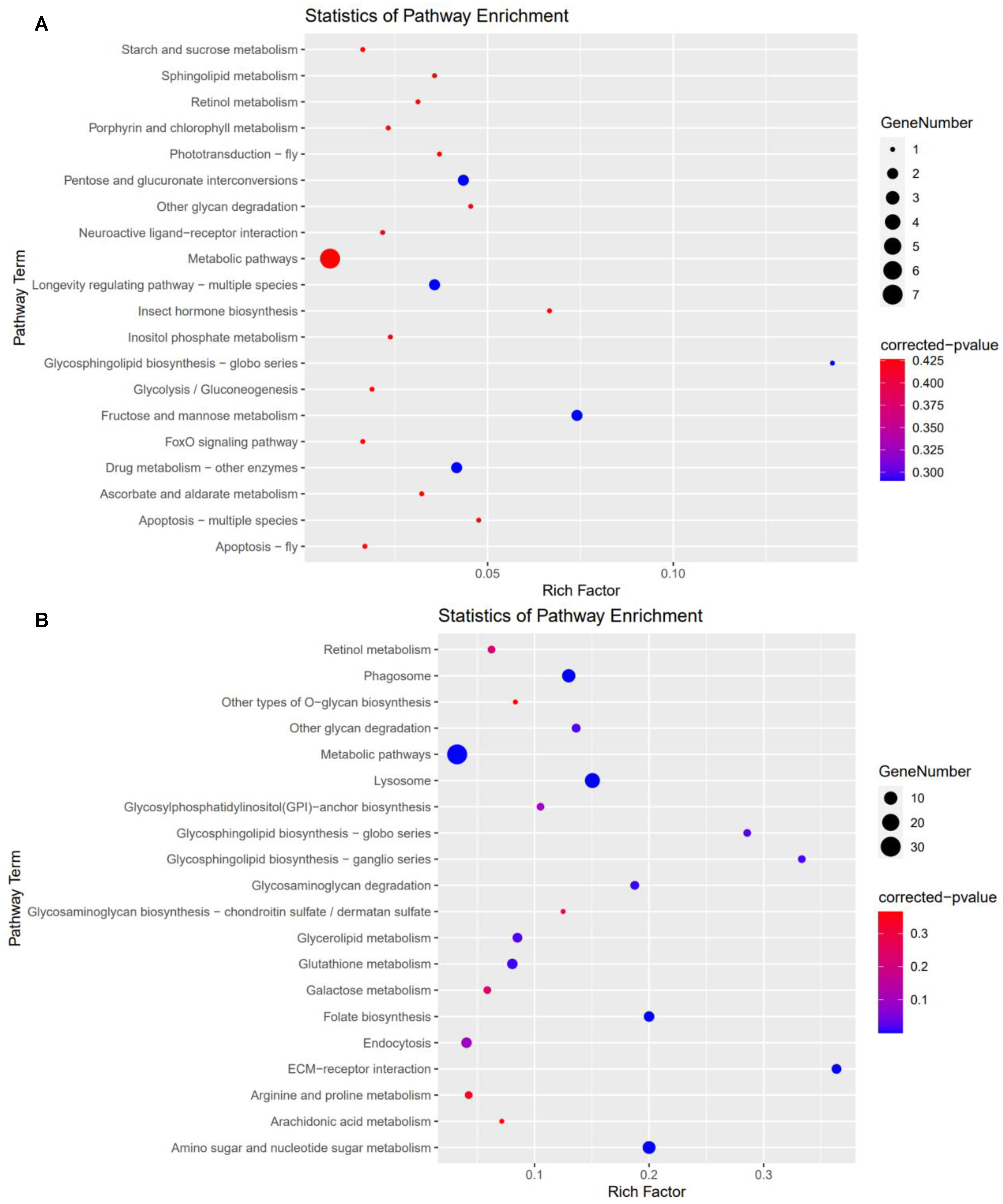
3.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis
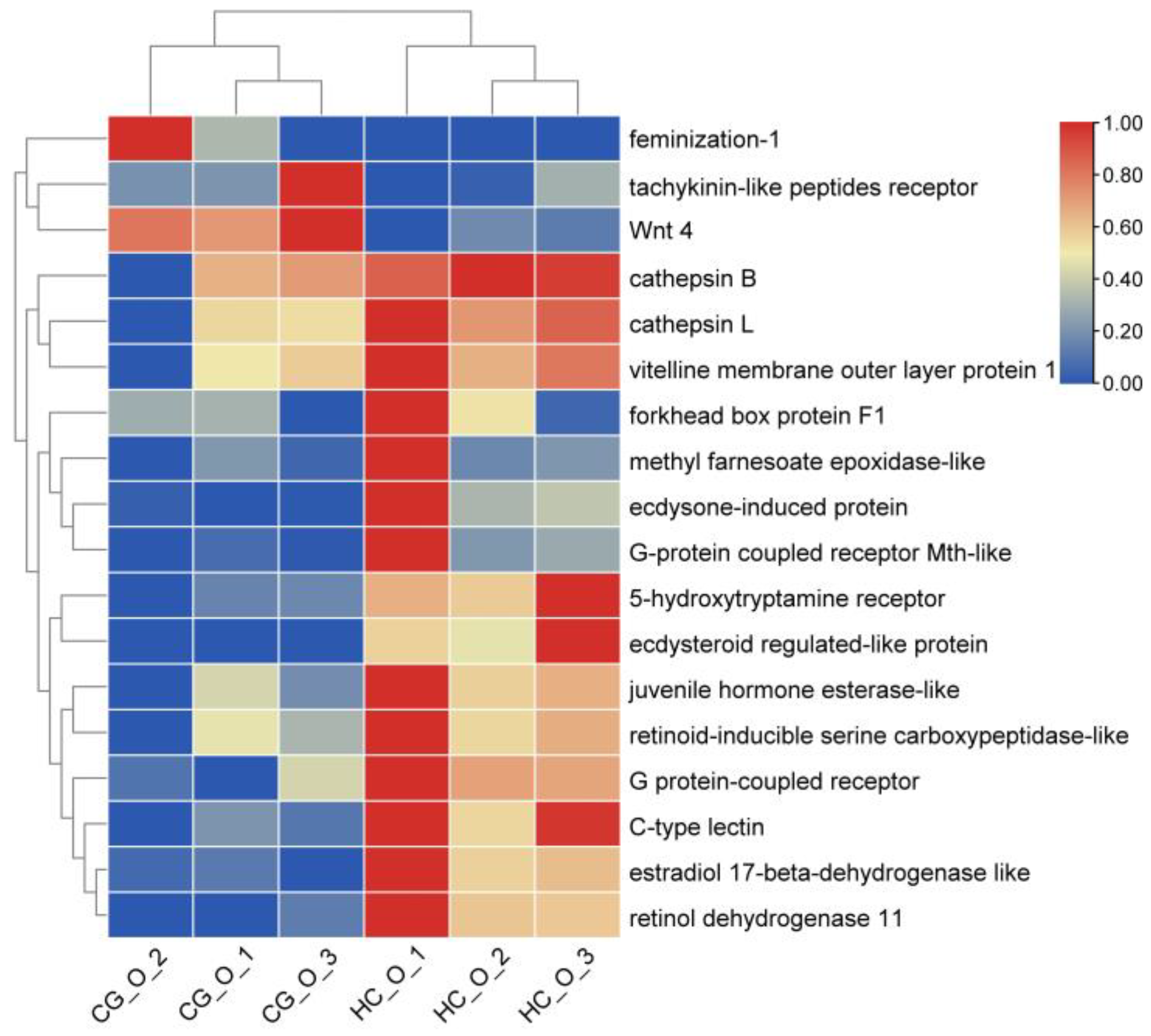
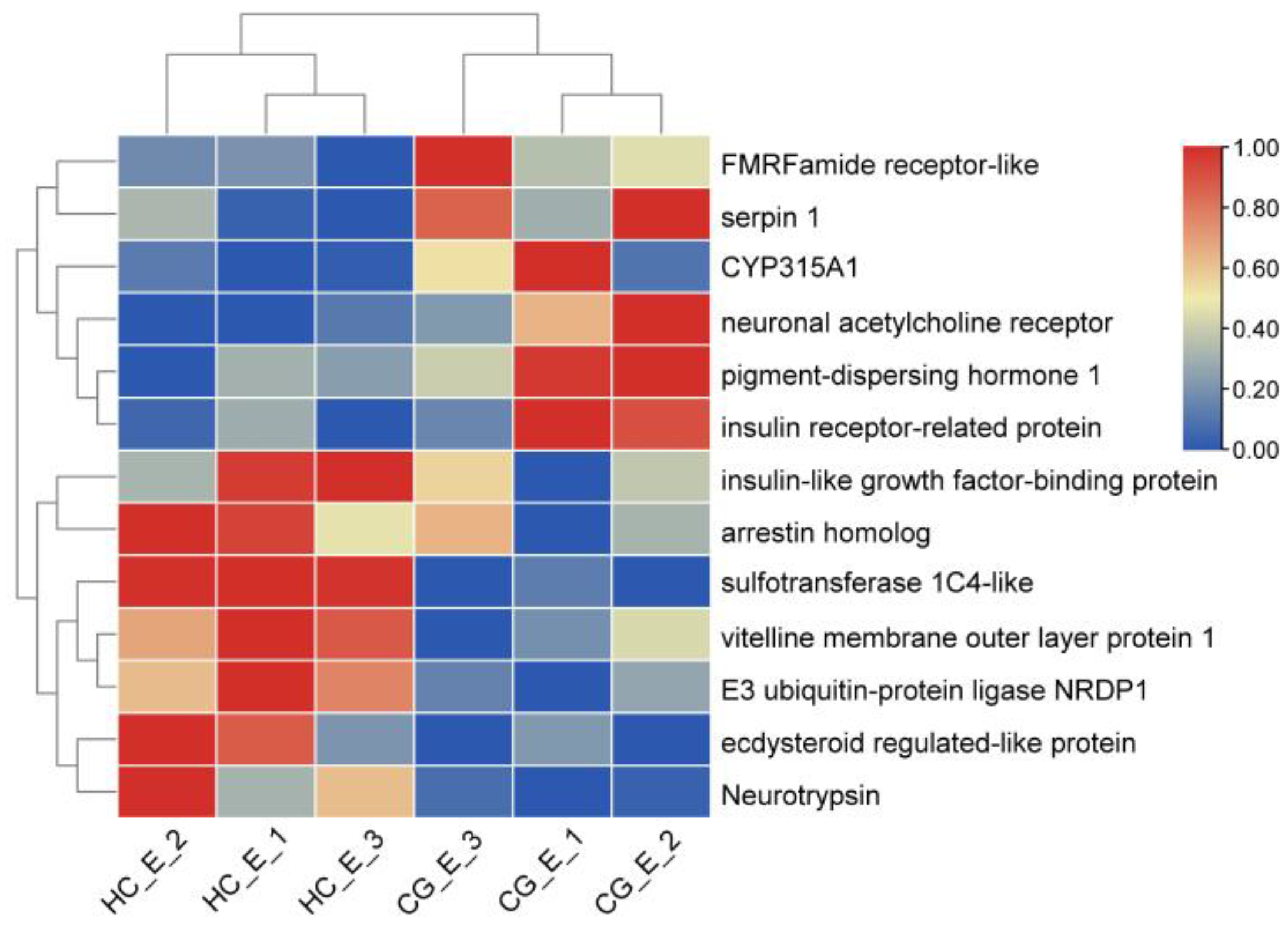
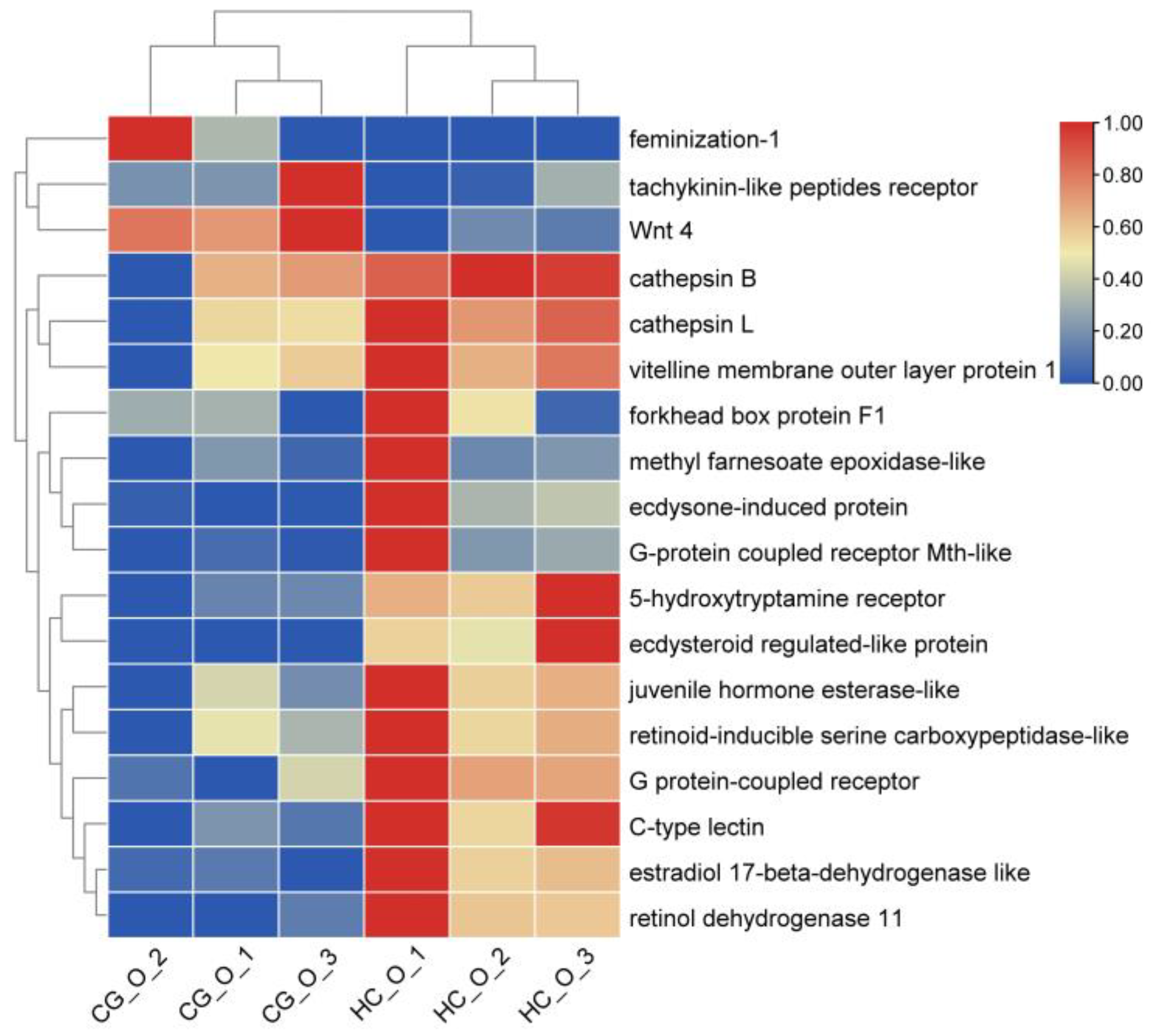
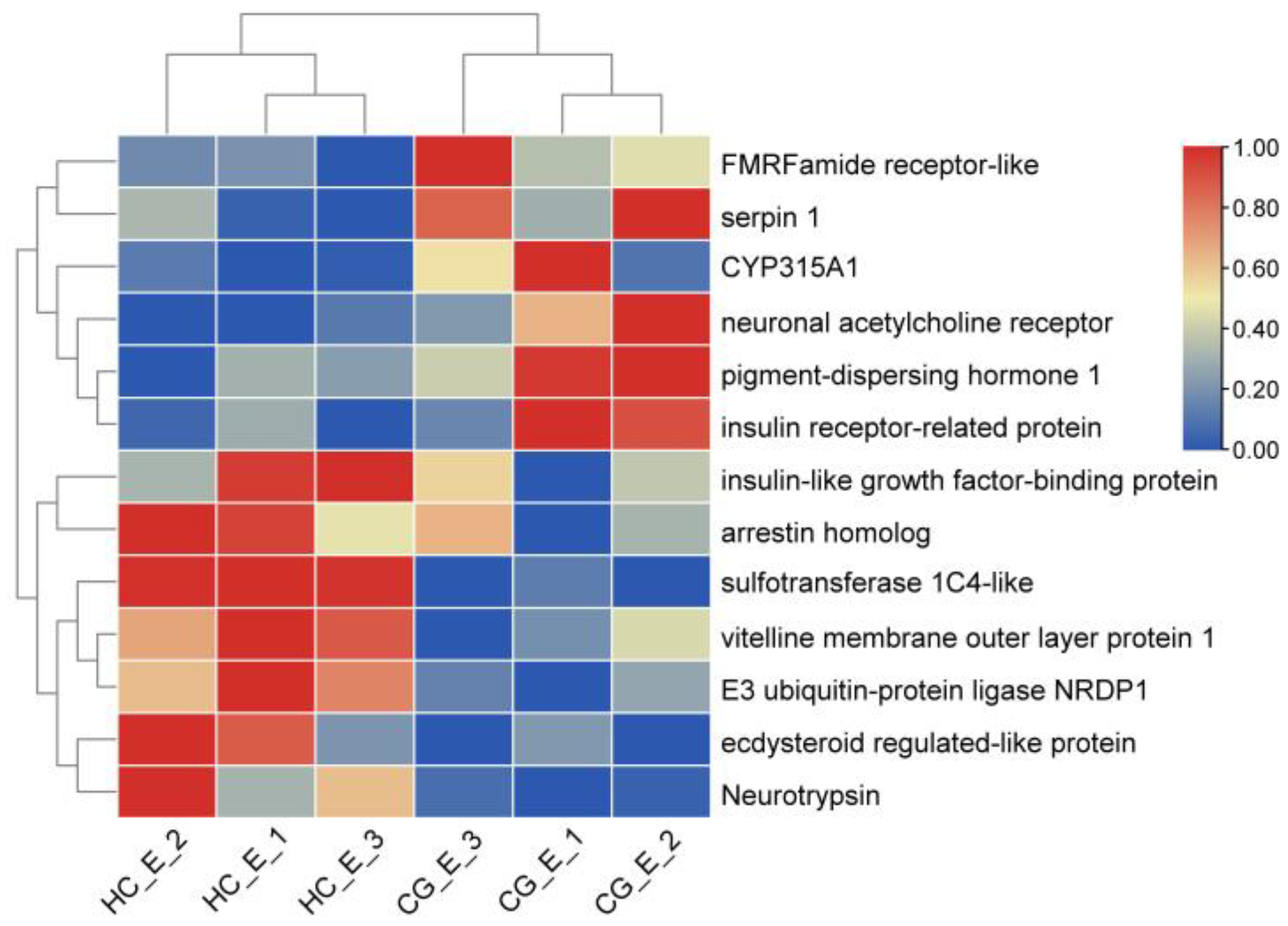
3.6. DEGs Involved in Ovarian Development
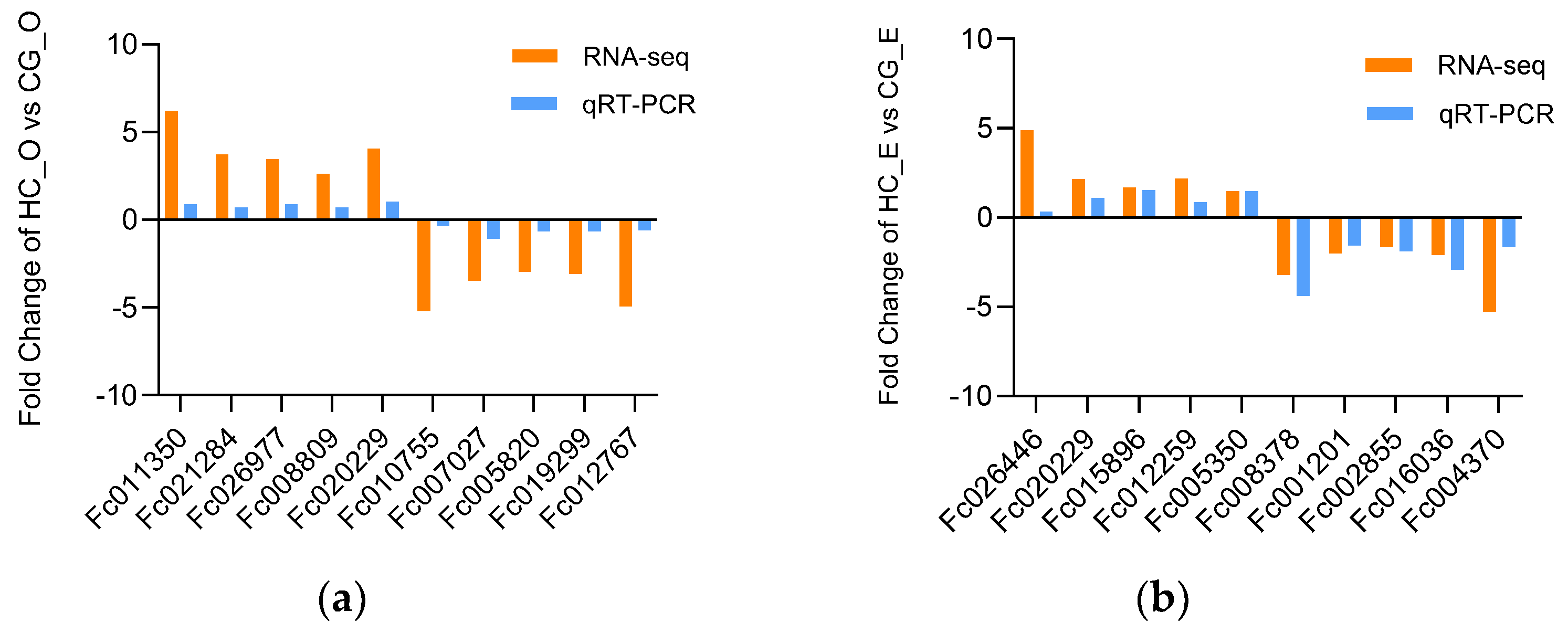
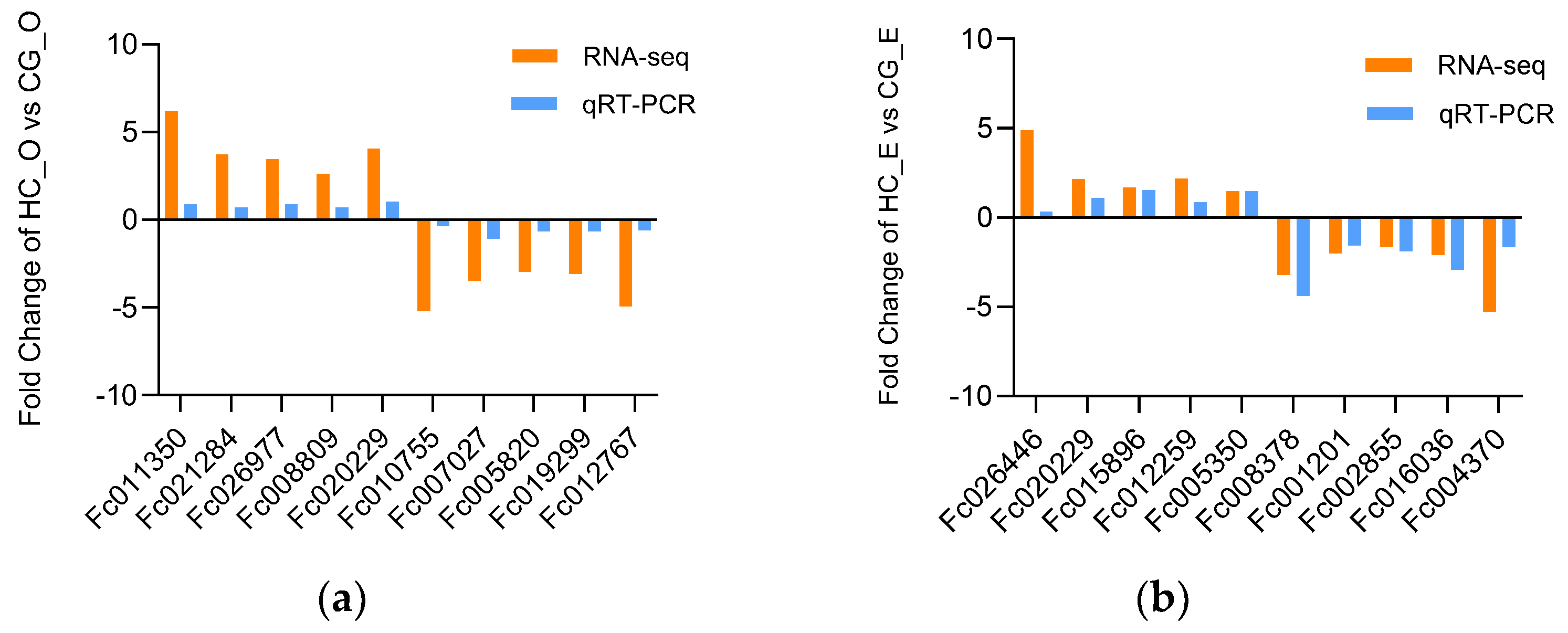
3.7. The Validation of DEGs by qRT−PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, B.R.; Minhas, P.S. Strategies for managing saline/alkali waters for sustainable agricultural production in South Asia. Agric. Water Manag. 2005, 78, 136–151. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, C.; Liu, G.; Jing, J. Development of a cDNA microarray to identify gene expression of Puccinellia tenuiflora under saline-alkali stress. Plant Physiol. Bioch. 2007, 45, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.L.; Lai, Q.F.; Zhou, K.; Rizalita, R.E.; Wang, H. Developmental biology of medaka fish (Oryzias latipes) exposed to alkalinity stress. J. Appl. Ichthyol. 2010, 26, 397–402. [Google Scholar] [CrossRef]
- Fan, Z.; Wu, D.; Zhang, Y.Y.; Li, J.; Xu, Q.Y.; Wang, L.S. Carbonate alkalinity and dietary protein levels affected growth performance, intestinal immune responses and intestinal microflora in Songpu mirror carp (Cyprinus carpio Songpu). Aquaculture 2021, 545, 737135. [Google Scholar] [CrossRef]
- Lin, T.; Lai, Q.; Yao, Z.; Lu, J.; Zhou, K.; Wang, H. Combined effects of carbonate alkalinity and pH on survival, growth and haemocyte parameters of the Venus clam Cyclina sinensis. Fish Shellfish Immun. 2013, 35, 525–531. [Google Scholar] [CrossRef]
- Xu, J.; Ji, P.; Wang, B.; Zhao, L.; Wang, J.; Zhao, Z.; Zhang, Y.; Li, J.; Xu, P.; Sun, X. Transcriptome sequencing and analysis of wild amur ide (Leuciscus waleckii) inhabiting an extreme alkaline–saline lake reveals insights into stress adaptation. PLoS ONE 2013, 8, e59703. [Google Scholar] [CrossRef]
- Li, H.; Lai, Q.; Yao, Z.; Liu, Y.; Sun, Z. Ammonia excretion and blood gas variation in naked carp (Gymnocypris przewalskii) exposed to acute hypoxia and high alkalinity. Fish Physiol. Biochem. 2020, 46, 1981–1990. [Google Scholar] [CrossRef]
- Su, H.; Ma, D.; Zhu, H.; Liu, Z.; Gao, F. Transcriptomic response to three osmotic stresses in gills of hybrid tilapia (Oreochromis mossambicus female × O. urolepis hornorum male). BMC Genom. 2020, 21, 110. [Google Scholar] [CrossRef] [Green Version]
- Li, J.T.; Li, J.; Duan, Y.F.; Chen, P.; Liu, P. The roles of heat shock proteins 70 and 90 in Exopalaemon carinicauda after WSSV and Vibrio anguillarum challenges. J. Ocean Univ. China 2018, 17, 399–406. [Google Scholar] [CrossRef]
- Feng, Y.Y.; Zhai, Q.Q.; Wang, J.J.; Li, J.T.; Li, J. Comparison of florfenicol pharmacokinetics in Exopalaemon carinicauda at different temperatures and administration routes. J. Vet. Pharmacol. Ther. 2019, 42, 230–238. [Google Scholar] [CrossRef]
- Li, J.T.; Li, J.; Chen, P.; Liu, P.; He, Y.Y. Transcriptome analysis of eyestalk and hemocytes in the ridgetail white prawn Exopalaemon carinicauda: Assembly, annotation and marker discovery. Mol. Biol. Rep. 2015, 42, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Ma, P.; Ping, L.; Chen, P.; Jian, L. The roles of Na+/K+-ATPase α-subunit gene from the ridgetail white prawn Exopalaemon carinicauda in response to salinity stresses. Fish Shellfish Immun. 2015, 42, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.Q.; Li, J.; Wang, J.J.; Li, Z.D.; Li, J.T. Characterization, functional analysis, and expression levels of three carbonic anhydrases in response to pH and saline–alkaline stresses in the ridgetail white prawn Exopalaemon carinicauda. Cell Stress Chaperon. 2019, 24, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.Q.; Neori, A.; He, Y.Y.; Li, J.T.; Li, J. Development and current state of seawater shrimp farming, with an emphasis on integrated multi–trophic pond aquaculture farms, in China–a review. Rev. Aquacult. 2020, 4, 2544–2558. [Google Scholar] [CrossRef]
- González-Vera, C.; Brown, J.H. Effects of alkalinity and total hardness on growth and survival of postlarvae freshwater prawns, Macrobrachium rosenbergii (De Man 1879). Aquaculture 2017, 473, 521–527. [Google Scholar] [CrossRef]
- Yao, Z.L.; Wang, H.; Chen, L.; Zhou, K.; Ying, C.Q.; Lai, Q.F. Transcriptomic profiles of Japanese medaka (Oryzias latipes) in response to alkalinity stress. Genet. Mol. Res. 2012, 11, 2200–2246. [Google Scholar] [CrossRef]
- Mcfarland, K.; Donaghy, L.; Volety, A.K. Effect of acute salinity changes on hemolymph osmolality and clearance rate of the non-native mussel, Perna viridis, and the native oyster, Crassostrea virginica, in Southwest Florida. Aquat. Invasions 2013, 8, 299–310. [Google Scholar] [CrossRef]
- Sun, Y.C.; Han, S.C.; Yao, M.Z.; Liu, H.B.; Wang, Y.M. Exploring the metabolic biomarkers and pathway changes in crucian under carbonate alkalinity exposure using high–throughput metabolomics analysis based on UPLC–ESI–QTOF–MS. RSC Adv. 2020, 10, 1552–1571. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, C.; Zhang, L.; Wei, J.; Yang, W.; Teng, L. Effects of alkalinity and pH on the survival, growth and neonate production of two strains of Moina mongolica Daday. Acta Ecol. Sin. 2009, 2, 589–598, (In Chinese with English Abstract). [Google Scholar]
- Xu, W.; Geng, L.W.; Li, C.T.; Jin, G.X.; Liu, X.Y. The artificial propagation, embryonic development and saline–alkali tolerant experiment of Barbus capito. J. Fish. China 2011, 2, 255–260, (In Chinese with English Abstract). [Google Scholar]
- Liu, F.; Li, J.; Li, J.T.; Ge, Q.Q.; Ge, H.X.; Shen, M.M. Effects of carbonate alkalinity stress on the survival, growth, reproduction, and immune enzyme activities of Exopalaemon carinicauda. J. Fish. Sci. China 2016, 5, 1137–1147, (In Chinese with English Abstract). [Google Scholar]
- Dugas, D.V. Functional annotation of the transcriptome of Sorghum bicolor in response to osmotic stress and abscisic acid. BMC Genom. 2011, 12, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teets, N.M.; Peyton, J.T.; Colinet, H.; Renault, D.; Denlinger, D.L. Gene expression changes governing extreme dehydration tolerance in an Antarctic insect. Proc. Natl. Acad. Sci. USA 2013, 109, 20744–20749. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, E.C.; Li, T.Y.; Xu, C.; Wang, X.D.; Lin, H.Z.; Qin, J.G.; Chen, L.Q. Transcriptome and molecular pathway analysis of the hepatopancreas in the pacific white shrimp Litopenaeus vannamei under chronic low–salinity stress. PLoS ONE 2015, 10, e131503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, E.; Wang, S.; Li, C.; Wang, X.; Chen, K.; Chen, L. Transcriptome sequencing revealed the genes and pathways involved in salinity stress of Chinese mitten crab, Eriocheir sinensis. Physiol. Genom. 2014, 46, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Biogeosciences 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Bu, D.H.; Luo, H.T.; Huo, P.P.; Wang, Z.H.; Zhang, S.; He, Z.H.; Wu, Y.; Zhao, L.H.; Liu, J.J.; Guo, J.C.; et al. KOBAS–i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, 317–325. [Google Scholar] [CrossRef]
- Duan, Y.F.; Liu, P.; Li, J.T.; Li, J.; Gao, B.Q. cDNA cloning, characterization and expression analysis of peroxiredoxin 5 gene in the ridgetail white prawn Exopalaemon carinicauda. Mol. Biol. Rep. 2013, 40, 6569–6577. [Google Scholar] [CrossRef] [Green Version]
- Sittikankaew, K.; Pootakham, W.; Sonthirod, C.; Sangsrakru, D.; Yoocha, T.; Khudet, J.; Nookaew, I.; Uawisetwathana, U.; Rungrassamee, W.; Karoonuthaisiri, N. Transcriptome analyses reveal the synergistic effects of feeding and eyestalk ablation on ovarian maturation in black tiger shrimp. Sci. Rep. 2020, 10, 3229. [Google Scholar]
- Mak, A.S.C.; Choi, C.L.; Tiu, S.H.K.; Hui, J.H.L.; He, J.G.; Tobe, S.S.; Chan, S.M. Vitellogenesis in the red crab Charybdis feriatus: Hepatopancreas–specific expression and farnesoic acid stimulation of vitellogenin gene expression. Mol. Reprod. Dev. 2005, 70, 288–300. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, H.H.; Huang, H.Y.; Jin, Z.X.; Li, S.J. Cloning, expression and functional analysis of farnesoic acid O-methyltransferase (FAMeT) in the mud crab, Scylla paramamosain. Mar. Freshw. Behav. Phy. 2012, 45, 209–222. [Google Scholar] [CrossRef]
- Miyakawa, H.; Toyota, K.; Sumiya, E.; Iguchi, T. Comparison of JH signaling in insects and crustaceans. Curr. Opin. Insect Sci. 2014, 1, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Veeran, S.; Wang, J.; Li, S.; Liu, S.N. Dual roles of juvenile hormone signaling during early oogenesis in Drosophila. Insect Sci. 2019, 27, 665–674. [Google Scholar] [CrossRef]
- Suang, S.; Hiruma, K.; Yu, K.; Manaboon, M. Diapause hormone directly stimulates the prothoracic glands of diapause larvae under juvenile hormone regulation in the bamboo borer, Omphisa fuscidentalis Hampson. Arch. Insect Biochem. 2019, 102, e21603. [Google Scholar] [CrossRef] [PubMed]
- Tomer, V.; Rivka, M.; Aflalo, E.D.; Vered, C.C.; Simy, W.; Omri, S.; Amir, S.; Yu, J. Post–embryonic transcriptomes of the prawn Macrobrachium rosenbergii: Multigenic succession through metamorphosis. PLoS ONE 2013, 8, e55322. [Google Scholar]
- Ventura, T.; Fitzgibbon, Q.P.; Battaglene, S.C.; Elizur, A. Redefining metamorphosis in spiny lobsters: Molecular analysis of the phyllosoma to puerulus transition in Sagmariasus verreauxi. Sci. Rep. 2015, 5, 13537. [Google Scholar] [CrossRef] [Green Version]
- Lamming, D.W.; Bar-Peled, L. Lysosome: The metabolic signaling hub. Traffic 2019, 20, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, Z.; Andersen, C.L.; Ye, X. Functions of lysosomes in mammalian female reproductive system. Reprodu. Dev. Med. 2020, 4, 109–122. [Google Scholar] [CrossRef]
- Harwood, G.; Amdam, G. Vitellogenin in the honey bee midgut. Apidologie 2021, 52, 837–847. [Google Scholar] [CrossRef]
- Carnevali, O.; Cionna, C.; Tosti, L.; Lubzens, E.; Maradonna, F. Role of cathepsins in ovarian follicle growth and maturation. Gen. Comp. Endocr. 2006, 146, 195–203. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Jiang, S.F.; Qiao, H.; Xiong, Y.W.; Fu, H.T.; Zhang, W.Y.; Gong, Y.S.; Jin, S.B.; Wu, Y. Transcriptome analysis of five ovarian stages reveals gonad maturation in female updates Macrobrachium nipponense. BMC Genom. 2021, 22, 510. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.Y.; Qian, H.L.; Jiang, H.C.; Xiong, X.Y.; Ye, B.Q.; Liu, X.; Guo, Z.Q.; Ma, K.Y. Transcriptional changes revealed water acidification leads to the immune response and ovary maturation delay in the Chinese mitten crab Eriocheir sinensis. Comp. Biochem. Phys. D 2021, 39, 100868. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Qi, D.; Chen, L.Q.; Zhang, H.; Zhang, X.W.; Qin, J.G.; Hu, S.N. Gene discovery from an ovary cDNA library of oriental river prawn Macrobrachium nipponense by ESTs annotation. Comp. Biochem. Phys. D 2009, 4, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.V.; Rotllant, G.E.; Cummins, S.F.; Elizur, A.; Ventura, T. Insights into sexual maturation and reproduction in the norway lobster (Nephrops norvegicus) via in silico prediction and characterization of neuropeptides and G protein–coupled receptors. Front. Endocrinol. 2018, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Flaherty, P.; Radhakrishnan, M.L.; Dinh, T.; Rebres, R.A.; Roach, T.I.; Jordan, M.I.; Arkin, A.P. A dual receptor crosstalk model of G protein coupled signal transduction. PLoS Comput. Biol. 2008, 4, e1000185. [Google Scholar] [CrossRef]
- Subramoniam, T. Mechanisms and control of vitellogenesis in crustaceans. Fish. Sci. 2011, 77, 1–21. [Google Scholar] [CrossRef]
- Crown, A.; Clifton, D.K.; Steiner, R.A. Neuropeptide signaling in the integration of metabolism and reproduction. Neuroendocrinology 2016, 86, 175–182. [Google Scholar] [CrossRef]
- Sharabi, O.; Manor, R.; Weil, S.; Aflalo, E.D.; Lezer, Y.; Levy, T.; Aizen, J.; Ventura, T.; Mather, P.B.; Khalaila, I. Identification and characterization of an insulin–like receptor involved in crustacean reproduction. Endocrinology 2016, 157, 928–941. [Google Scholar] [CrossRef] [Green Version]
- Christie, A.E.; Stemmler, E.A.; Dickinson, P.S. Crustacean neuropeptides. Cell. Mol. Life Sci. 2010, 67, 4135–4169. [Google Scholar] [CrossRef]
- Tateno, H.; Saneyoshi, A.; Ogawa, T.; Muramoto, K.; Kamiya, H.; Saneyoshi, M. Isolation and characterization of rhamnose-binding lectins from eggs of steelhead trout (Oncorhynchus mykiss) homologous to low density lipoprotein receptor superfamily. J. Biol. Chem. 1998, 273, 19190–19197. [Google Scholar] [CrossRef] [Green Version]
- Dodd, R.B.; Kurt, D. Lectin-like proteins in model organisms: Implications for evolution of carbohydrate–binding activity. Glycobiology 2001, 11, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Zhang, J.Q.; Li, F.H.; Liu, C.Z.; Xiang, J.H. Cloning of a novel C–type lectin LvLec2 from the shrimp Litopenaeus vannamei and its immune response to different challenges. Mar. Sci. 2010, 34, 103–110. [Google Scholar]
- Qin, Y.; Jiang, S.; Huang, J.; Zhou, F.; Yang, Q.; Jiang, S.; Yang, L. C–type lectin response to bacterial infection and ammonia nitrogen stress in tiger shrimp (Penaeus monodon). Fish Shellfish Immun. 2019, 90, 188–198. [Google Scholar] [CrossRef]
- Kido, S.; Morimoto, A.; Kim, F.; Doi, Y. Isolation of a novel protein from the outer layer of the vitelline membrane. Biochem. J. 1992, 286, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Back, J.F.; Bain, J.M.; Vadehra, D.V.; Burley, R.W. Proteins of the outer layer of the vitelline membrane of hen’s eggs. Biochim. Biophys. Acta. 1982, 705, 12–19. [Google Scholar] [CrossRef]
- Rahman, M.A.; Akihiko, M.; Atsushi, I.; Norio, Y. VMO–II mediates the binding of the chalaziferous layer with the vitelline membrane in quail eggs. J. Poult. Sci. 2009, 46, 240–248. [Google Scholar] [CrossRef]
- Gismondi, E.; Thome, J.P.; Urien, N.; Uher, E.; Baiwir, D.; Mazzucchelli, G.; De Pauw, E.; Fechner, L.C.; Lebrun, J.D. Ecotoxicoproteomic assessment of the functional alterations caused by chronic metallic exposures in gammarids. Environ. Pollut. 2017, 225, 428–438. [Google Scholar] [CrossRef]
- Sricharoen, S.; Kim, J.J.; Tunkijjanukij, S.; Soderhall, I. Exocytosis and proteomic analysis of the vesicle content of granular hemocytes from a crayfish. Dev. Comp. Immunol. 2005, 29, 1017–1031. [Google Scholar] [CrossRef]
- Philip, B.; Abigail, E.; Richard, W.; Cummins, S.F.; Wayne, K. Gene expression profiling of the cephalothorax and eyestalk in penaeus monodon during ovarian maturation. Int. J. Biol. Sci. 2012, 8, 328–343. [Google Scholar]
- Jung, H.T.; Lyons, R.E.; Hurwood, D.A.; Mather, P. Genes and growth performance in crustacean species: A review of relevant genomic studies in crustaceans and other taxa. Rev. Aquacult. 2013, 5, 77–110. [Google Scholar] [CrossRef]
- Koshio, S.; Teshima, S.I.; Kanazawa, A. Effects of unilateral eyestalk ablation and feeding frequencies on growth, survival, and body compositions of juvenile freshwater prawn Macrobrachium rosenbergii. Nippon Suisan Gakk. 1992, 58, 1419–1425. [Google Scholar] [CrossRef]
- Qiao, H.; Xiong, Y.; Zhang, W.; Fu, H.; Jiang, S.; Sun, S.; Bai, H.; Jin, S.; Gong, Y. Characterization, expression, and function analysis of gonad–inhibiting hormone in Oriental River prawn, Macrobrachium nipponense and its induced expression by temperature. Comp. Biochem. Phys. A 2015, 185, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rotllant, G.; Nguyen, T.V.; Aizen, J.; Suwansa-ard, S.; Ventura, T. Toward the identification of female gonad–stimulating factors in crustaceans. Hydrobiologia 2018, 825, 91–119. [Google Scholar] [CrossRef]
- Huang, X.; Ye, H.; Huang, H.; Yu, K.; Huang, Y. Two beta-pigment–dispersing hormone (β–PDH) isoforms in the mud crab, Scylla paramamosain: Implication for regulation of ovarian maturation and a photoperiod-related daily rhythmicity. Anim. Reprod. Sci. 2014, 150, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Bao, C.; Yang, Y.; Huang, H.; Ye, H. Neuropeptides in the cerebral ganglia of the mud crab, Scylla paramamosain: Transcriptomic analysis and expression profiles during vitellogenesis. Sci. Rep. 2015, 5, 17055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.L.; Chen, T.T.; Luo, B.Y.; Qiu, G.F. Evidences for Red Pigment Concentrating Hormone (RPCH) and Beta-Pigment Dispersing Hormone (beta–PDH) Inducing Oocyte Meiotic Maturation in the Chinese Mitten Crab, Eriocheir sinensis. Front. Endocrinol. 2021, 12, 802768. [Google Scholar] [CrossRef]
- Jiang, Y.W.; Li, C.J.; Chen, L.; Wang, F.G.; Zhou, X. Potential role of retinoids in ovarian physiology and pathogenesis of polycystic ovary syndrome. Clin. Chim. Acta 2017, 469, 87–93. [Google Scholar] [CrossRef]
- Kawai, T.; Yanaka, N.; Richards, J.S.; Shimada, M. De novo–synthesized retinoic acid in ovarian antral follicles enhances FSH–mediated ovarian follicular cell differentiation and female fertility. Endocrinology 2016, 157, 2160–2172. [Google Scholar] [CrossRef] [Green Version]
- Damdimopoulou, P.; Chiang, C.; Flaws, J.A. Retinoic acid signaling in ovarian folliculogenesis and steroidogenesis. Reprod. Toxicol. 2019, 87, 32–41. [Google Scholar] [CrossRef]
- Tahaei, L.S.; Eimani, H.; Yazdi, P.E.; Ebrahimi, B.; Fathi, R. Effects of retinoic acid on maturation of immature mouse oocytes in the presence and absence of a granulosa cell co–culture system. J. Assist. Reprod. Gen. 2011, 28, 553–558. [Google Scholar] [CrossRef] [Green Version]
- Levi, L.; Ziv, T.; Admon, A.; Levavi-Sivan, B.; Lubzens, E. Insight into molecular pathways of retinal metabolism, associated with vitellogenesis in zebrafish. Am. J. Physiol.-Endoc. Metab. 2012, 302, 626–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, P.F.; Mao, B.; Fan, C.; Wang, Y.F. Transcriptomic information from the ovaries of red swamp crayfish (Procambarus clarkii) provides new insights into development of ovaries and embryos. Aquaculture 2019, 505, 333–343. [Google Scholar] [CrossRef]
- Wang, J.J.; Li, J.T.; Ge, Q.Q.; Li, W.Y.; Li, J. Full–Length transcriptome sequencing and comparative transcriptomic analysis provide insights into the ovarian maturation of Exopalaemon carinicauda. Front. Mar. Sci. 2022, 9, 906730. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Significant | Up/Down | ID | p Value |
|---|---|---|---|---|
| Steroid metabolic process | 31 | up | 0008202 | 5.70 × 10−7 |
| Sterol homeostasis | 11 | up | 0055092 | 9.50 × 10−5 |
| Molting cycle | 22 | up | 0042303 | 0.00016 |
| Follicle cell microvillus organization | 3 | up | 0032529 | 0.00031 |
| Retinoid metabolic process | 12 | up | 0001523 | 0.00052 |
| Steroid biosynthetic process | 16 | up | 0006694 | 0.00076 |
| Sterol transport | 11 | up | 0015918 | 0.00486 |
| Response to estrogen | 9 | up | 0043627 | 0.00875 |
| Estrogen secretion | 2 | up | 0035937 | 0.01058 |
| Positive regulation of ovulation | 3 | down | 0060279 | 0.00099 |
| Regulation of ovulation | 3 | down | 0060278 | 0.00176 |
| Sterol transport | 6 | down | 0015918 | 0.02138 |
| Ovulation | 3 | down | 0030728 | 0.03153 |
| Sterol homeostasis | 4 | down | 0055092 | 0.04769 |
| Term | Significant | Up/Down | ID | p Value |
|---|---|---|---|---|
| Positive regulation of sterol transport | 2 | up | 0032373 | 0.00482 |
| Regulation of sterol transport | 2 | up | 0032371 | 0.01030 |
| Embryonic morphogenesis | 10 | up | 0048598 | 0.02561 |
| Embryonic organ morphogenesis | 5 | up | 0048562 | 0.02572 |
| Embryo development | 16 | up | 0009790 | 0.02894 |
| Entry into reproductive diapause | 1 | up | 0055116 | 0.04250 |
| Sterol metabolic process | 5 | up | 0008202 | 0.02501 |
| Sterol transport | 7 | down | 0015918 | 0.00226 |
| Regulation of metaphase/anaphase transition of meiosis I | 1 | down | 1905186 | 0.01765 |
| Ecdysteroid metabolic process | 3 | down | 0045455 | 0.02708 |
| Sterol homeostasis | 4 | down | 0055092 | 0.02759 |
| Maternal placenta development | 2 | down | 0001893 | 0.02798 |
| Metaphase/anaphase transition of meiosis I | 1 | down | 1905186 | 0.01765 |
| Molting cycle | 8 | down | 0042303 | 0.03756 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, J.; Wang, C.; Li, W.; Ge, Q.; Qin, Z.; Li, J.; Li, J. Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda. Water 2022, 14, 3690. https://doi.org/10.3390/w14223690
Zhang X, Wang J, Wang C, Li W, Ge Q, Qin Z, Li J, Li J. Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda. Water. 2022; 14(22):3690. https://doi.org/10.3390/w14223690
Chicago/Turabian StyleZhang, Xiuhong, Jiajia Wang, Chengwei Wang, Wenyang Li, Qianqian Ge, Zhen Qin, Jian Li, and Jitao Li. 2022. "Effects of Long-Term High Carbonate Alkalinity Stress on the Ovarian Development in Exopalaemon carinicauda" Water 14, no. 22: 3690. https://doi.org/10.3390/w14223690