
Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions

Key Laboratory of Mariculture (Ministry of Education), Fisheries College, Ocean University of China, Qingdao 266003, China
*
Authors to whom correspondence should be addressed.
Water 2022, 14(17), 2664; https://doi.org/10.3390/w14172664
Submission received: 20 July 2022
/
Revised: 25 August 2022
/
Accepted: 26 August 2022
/
Published: 28 August 2022
(This article belongs to the Special Issue Advances in Aquaculture Ecology Research)
Abstract
:The golden tide dominated by Sargassum has become a frequently-occurring marine ecological event that may constitute major biotic threats to seaweed aquaculture. In this study, the interaction between cultivated Gracilariopsis lemaneiformis (GL) and floating Sargassum horneri (SH) was investigated by physiological and biochemical measurements under mono-culture and co-culture with different biomass density ratios of 2:1 (2GL:1SH), 1:1 (1GL:1SH), and 1:2 (1GL:2SH). The relative growth rate, net photosynthetic rate, and NO3-N uptake rate of G. lemaneiformis were significantly greater at the biomass density ratio of 2:1 than at mono-culture. However, these physiological parameters and biochemical composition contents (chlorophyll a and soluble protein) of G. lemaneiformis decreased significantly with increasing biomass of S. horneri. Meanwhile, these physiological and biochemical parameters of S. horneri were greater in all co-culture models than at mono-culture. They decreased significantly with decreasing biomass of G. lemaneiformis. These results indicate that the occurrence of floating S. horneri with low biomass can stimulate the growth of G. lemaneiformis, whereas its outbreak may significantly reduce the production and quality of G. lemaneiformis. G. lemaneiformis cultivation may be beneficial to the increased biomass of floating S. horneri.
1. Introduction
In recent years, multiple large-scale outbreaks of golden tide caused by Sargassum have occurred along the coastal waters of the Pacific and Atlantic Ocean [1,2,3,4]. This phenomenon may be closely correlated with marine ecological deterioration, including seawater eutrophication and warming [5,6]. Along the northwest Pacific coast, Sargassum horneri is a predominant golden-tide-forming seaweed that originally forms extensive underwater forests [7,8]. Sessile populations of S. horneri have continuously decreased in this region [9], whereas a prominent drifting biomass has been frequently detected in the coastal regions of China in the last decade [3,10]. Golden tides have caused considerable disruption to coastal ecosystems, including the death of aquatic organisms by hypoxia, resource competition with native species, and even shifts in biological community structures [11,12]. Furthermore, it has brought considerable damage to local economic industries such as tourism, traditional fisheries, and mariculture [13,14,15].
Positive and negative interactions among plants are widespread in both terrestrial and aquatic ecosystems, and have complicated and changeable effects on population dynamics, community structure, and biodiversity patterns [16,17,18]. In coastal regions, a common interspecific interaction pattern is for marine plants with overlapping niches to compete for diverse limited environmental resources [19,20,21]. Epiphytic Ulva lactuca has been proven to severely interfere with mariculture of Neoporphyra haitanensis, as they compete for carbon and nitrogen resources [22]. Additionally, the growth of Neopyropia yezoensis was apparently restrained by Ulva prolifera proliferation because of competition for space, nutrients, and light for spore attachments [23,24]. Allelopathy is another pattern of interspecific interaction among marine algae, which can stimulate or inhibit each other through the release of biochemical metabolites [25,26,27]. Yuan et al. [28] showed that aqueous extract from Sargassum fusiforme promoted the growth of Karenia mikimotoi at 0.2 g L−1, while its growth was inhibited at 1.6 g L−1. Similarly, Patil et al. [29] documented that the growth of Skeletonema costatum was not significantly decreased by culture filtrates of Pyropia haitanensis at 0.625–10 g FW L−1, while it was greatly inhibited at 15 and 20 g FW L−1. These results imply that interaction through allelopathy is closely associated with the biomass density ratio among species. However, thus far there has been little information produced regarding the influence of the biomass density ratio on the interaction between marine macroalgae.
Gracilariopsis lemaneiformis is a common intertidal red seaweed that is naturally distributed across temperate regions worldwide [30,31,32]. In China, G. lemaneiformis is extensively cultivated due to its widespread applications in agar production, the food industry, and abalone aquaculture [33,34,35]. It is the seaweed species with the third highest production rate in terms of mass, with 368,967 tons of dry weight produced in 2020 [36]. Additionally, the cultivation of this species is beneficial to coastal ecosystems in many ways, including alleviation of harmful microalgal blooms, increasing the dissolved oxygen in the seawater, and maintaining coastal ecological balance [37]. In recent years, the deterioration of the marine environment caused by climate change and human activities has posed a severe threat to the mariculture production of seaweeds [38,39,40]. Due to the economic and ecological importance of G. lemaneiformis, a considerable number of studies about the impacts of diverse abiotic factors on its physiological and biochemical performance have been conducted [41,42,43,44]. Nevertheless, the effects of biotic stresses such as macroalgal blooms on G. lemaneiformis have rarely been reported.
In the current study, we performed a short-term culture experiment to examine the interactive influences between cultivated G. lemaneiformis and floating S. horneri. Changes in growth, photosynthesis, NO3-N uptake, and biochemical compositions were estimated. Our results provide significant information to improve the mariculture management of this valuable species and to address marine ecological hazards.
2. Materials and Methods
2.1. Sampling and Maintenance
G. lemaneiformis thalli were sampled from farmed populations on Lidao Island (36°26′ N, 122°56′ E), China, in June 2020. Meanwhile, S. horneri thalli were sampled from floating populations in the cultivation area of G. lemaneiformis. Healthy samples were chosen and completely rinsed using sterilized seawater to remove epiphytes and detritus. Algal fragments of G. lemaneiformis (6 cm in length) and S. horneri (3 cm in length) were respectively excised from the apical position of branches for the experiments. Then, they were incubated in tanks containing 6 L of 25% PESI medium [45]. These fragments were kept for three days at 20 °C, 90 μmol photons m−2 s−1, and a 12:12 L:D cycle in order to minimize the impacts of excision.
2.2. Culture Experiment
A twelve-day culture experiment was performed under mono-culture conditions of G. lemaneiformis and S. horneri and co-culture at different biomass density ratios (BDRs). For the mono-culture, the initial biomass density was set at 6 g for both species. For the co-culture, the initial BDR was set at 2:1 (4 g G. lemaneiformis cultured with 2 g S. horneri), 1:1 (3 g G. lemaneiformis cultured with 3 g S. horneri), and 1:2 (2 g G. lemaneiformis cultured with 4 g S. horneri). A total of five treatments were set up, and each treatment was conducted in three replicates. In this experiment, 20 °C, 90 μmol photons m−2 s−1, and a 12:12 L:D cycle were maintained. This experiment used fifteen tanks, with each tank containing 6 L of sterilized seawater enriched with 25% PESI medium. Gentle aeration was conducted during this experiment. The medium was changed every three days. The fresh weights (FWs) of all fragments before and after incubation were determined after each fragment was blotted dry. The relative growth rate (RGR; % day−1) was calculated by Equation (1):
where W0 is the initial FW, Wt is the final FW, and t is the culture time in days.
RGR = 100 × (lnWt − lnW0) / t
2.3. Measurement of Photosynthesis
Following the culture experiment, we determined the net photosynthetic rates of G. lemaneiformis and S. horneri using a manual FireStingO2II oxygen meter (Pyro Science GmbH, Aachen, Germany). For each species, 0.33 g (FW) of samples from each treatment were moved to the oxygen electrode cuvette containing 330 mL of 25% PESI medium. Next, the medium was continuously stirred to ensure homogenous oxygen diffusion. The culture conditions were the same as for the experiment above. The oxygen increase in the seawater was regarded as the net photosynthetic rate after an increase in the light density. Before measurements, these samples were placed into the cuvette for 5 min acclimation. The oxygen concentration in the medium was documented every 1 min for 15 min.
2.4. Measurement of NO3-N Uptake Rate
Following the culture experiment, 0.2 g (FW) of G. lemaneiformis and/or S. horneri were randomly selected from each tank for NO3-N uptake measurements. They were transferred into conical flasks containing 200 mL of sterilized seawater enriched with 25% PESI medium and gently shaken for 2 h using a horizontal oscillator. The light and temperature conditions were the same as those described for the culture experiment. For each treatment, the media before and after the culture experiment were separately collected and the NO3-N concentrations were determined by the cadmium column reduction method and molybdenum blue method with ultraviolet absorption spectrophotometer, respectively [46,47]. The NO3-N uptake rate was calculated by Equation (2):
where UN is the NO3-N uptake rate, C0 is the initial concentration of NO3-N (mg L−1), Ct is the final concentration of NO3-N (mg L−1) following the experiment, V is the medium volume (mL), T is the experiment period, and W is the FW of samples (g).
UN = (C0 − Ct) × V / (T × W)
2.5. Measurement of Chlorophyll a (Chl a)
0.1 g (FW) of G. lemaneiformis was used to extract the Chl a in each replicate of all the experimental treatments. These samples were ground using liquid nitrogen, then 3 mL of phosphate buffer solution (0.1 M, pH = 6.8) was added. After transferring the samples into tubes, they were centrifuged for 30 min at 4000 rpm at 4 °C. Subsequently, 8 mL of dimethylformamide (DMF) was added to the precipitates obtained following centrifugation and maintained at 4 °C for 1 d. Extracts were then centrifuged for 10 min at 6000 rpm at 4 °C. The supernatant was collected and the absorption was determined at 750, 664, 647, and 625 nm. The Chl a content (mg g−1) was calculated by Equation (3):
where A750–625 is the absorption of extracts at 750, 664, 647, and 625 nm, Ve is the DMF volume (mL), I is the optical path in the cuvette (cm), and W is the FW of the samples (g).
Chl a = [12.65 × (A664 − A750) − 2.99 × (A647 − A750) − 0.04 × (A625 − A750)] × Ve / (I × W × 1000)
0.25 g (FW) of S. horneri was used to extract the Chl a in each replicate of all the experimental treatments. These samples were placed into 2 mL of dimethyl sulfoxide for 5 min, and the supernatant absorption was determined at 665, 631, 582, and 480 nm. Subsequently, the same samples were placed into 3 mL of acetone for 2 h. The supernatant was moved to a 10 mL tube, then 1 mL of methanol and 1 mL of distilled water were added. The supernatant absorption was determined at 664, 631, 581, and 470 nm. The Chl a content (mg g−1) was calculated by Equation (4):
where A665 and A664 are the absorption rates of the extracts at 665 and 664 nm, respectively, V1 is the volume of the extract at the first extraction process (mL), V2 is the volume of extract at the second extraction process (mL), I is the optical path in the cuvette (cm), and W is the FW of the samples (g).
Chl a = [(A665 / 72.8) × V1 + (A664 / 73.6) × V2] / (I × W)
2.6. Measurements of Soluble Protein and Carbohydrate
For all the experimental treatments, 0.1 g (FW) of samples from each replicate were homogenized with a pestle and mortar and 0.9 mL of phosphate buffer solution (0.1 M, pH = 7.4). The extract was centrifuged for 30 min at 12,000 rpm at 4 °C. The supernatant absorption at 595 nm was recorded using an ultraviolet spectrophotometer. The soluble proteins of samples were evaluated using Coomassie Brilliant Blue G-250 dye (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) and bovine albumin [48].
An amount (0.1 g FW) of the samples from each replicate were ground in 2 mL of distilled water and diluted to 10 mL after adding 2 mL of MgCO3 suspension liquid. The mixture was centrifuged for 5 min at 4000 rpm at 4 °C. Then, 1 mL of supernatant was moved to a tube and diluted to 2 mL. After that, 8 mL of anthrone reagent was added. The mixture was bathed in boiled water for 10 min and cooled to room temperature. The absorption at 620 nm was recorded and the mixture was standardized. The soluble carbohydrate content was measured according to [49].
2.7. Statistical Analysis
For both species, one-way ANOVA was applied to analyze the RGR, net photosynthetic rate, NO3-N uptake rate, and the contents of biochemical compositions among different culture models (mono- or co-cultures with three BDRs). Prior to these analyses, the variances of the data were subjected to homogeneity tests. Duncan’s multiple range test was used if significant differences were detected (p < 0.05). Statistical analyses were carried out using SPSS 26.0 software.
3. Results
3.1. Growth
The RGRs of G. lemaneiformis were significantly affected by culture model (Figure 1A and Table 1). The RGR at BDR of 2:1 was significantly greater than that at mono-culture. However, the RGR decreased significantly with increasing biomass of S. horneri, from BDR of 2:1 to 1:2. The RGRs of S. horneri differed significantly among culture models (Figure 1B and Table 1). The RGRs of all co-culture treatments were greater than that at mono-culture, and significantly higher values were found at BDRs of 2:1 and 1:1. Additionally, the RGR at BDR of 2:1 was significantly greater than that at BDR of 1:1.
3.2. Photosynthesis
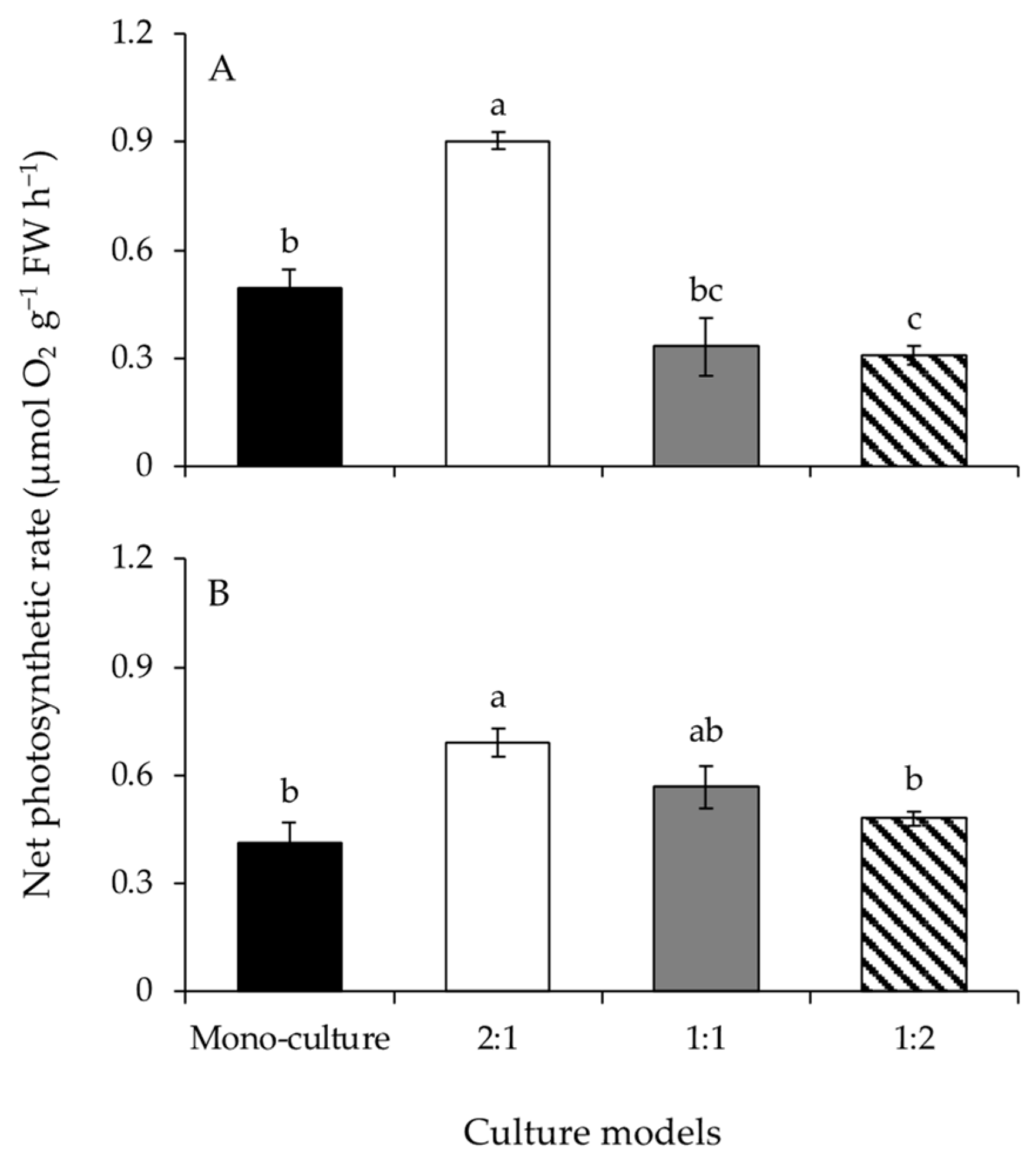
The net photosynthetic rates of G. lemaneiformis differed significantly among culture models (Figure 2A and Table 1). The net photosynthetic rate at BDR of 2:1 was significantly higher than those of the other culture models. Additionally, the net photosynthetic rate at BDR of 1:2 was significantly lower than that at mono-culture. The net photosynthetic rates of S. horneri were significantly affected by culture model (Figure 2B and Table 1). The net photosynthetic rate at BDR of 2:1 was significantly higher than that at mono-culture. However, the net photosynthetic rates decreased significantly with increasing biomass of S. horneri, from BDR of 2:1 to 1:2.
3.3. NO3-N Uptake Rate
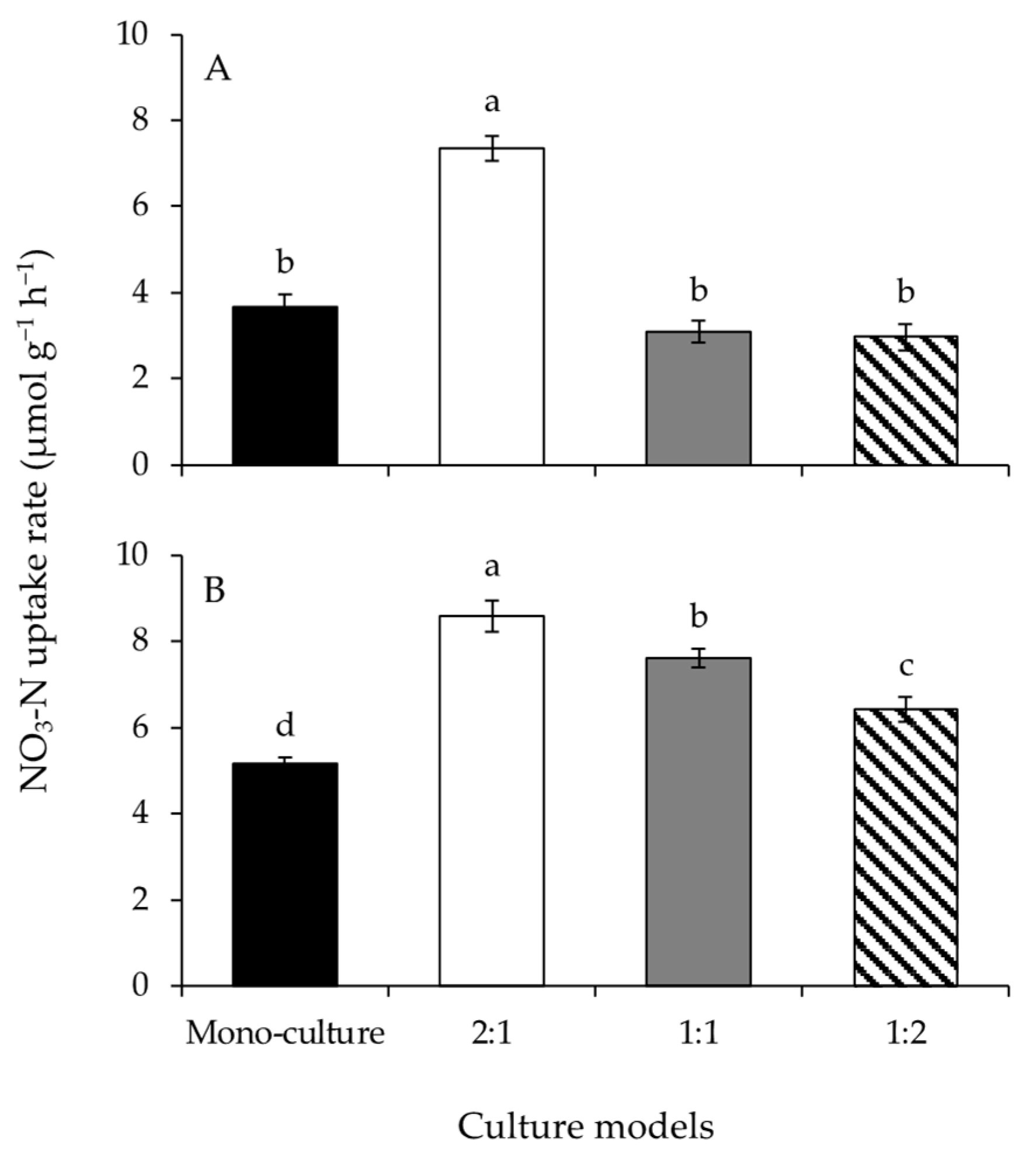
A significant effect of the culture model on the NO3-N uptake rates of G. lemaneiformis was detected (Figure 3A and Table 1). The NO3-N uptake rate at BDR of 2:1 was significantly greater than those of the other culture models. The NO3-N uptake rates of S. horneri were significantly different among culture models (Figure 3B and Table 1). The NO3-N uptake rates of the co-culture treatments were significantly greater than that at mono-culture. The NO3-N uptake rates decreased significantly with increasing biomass of S. horneri, from BDR of 2:1 to 1:2.
3.4. Chl a
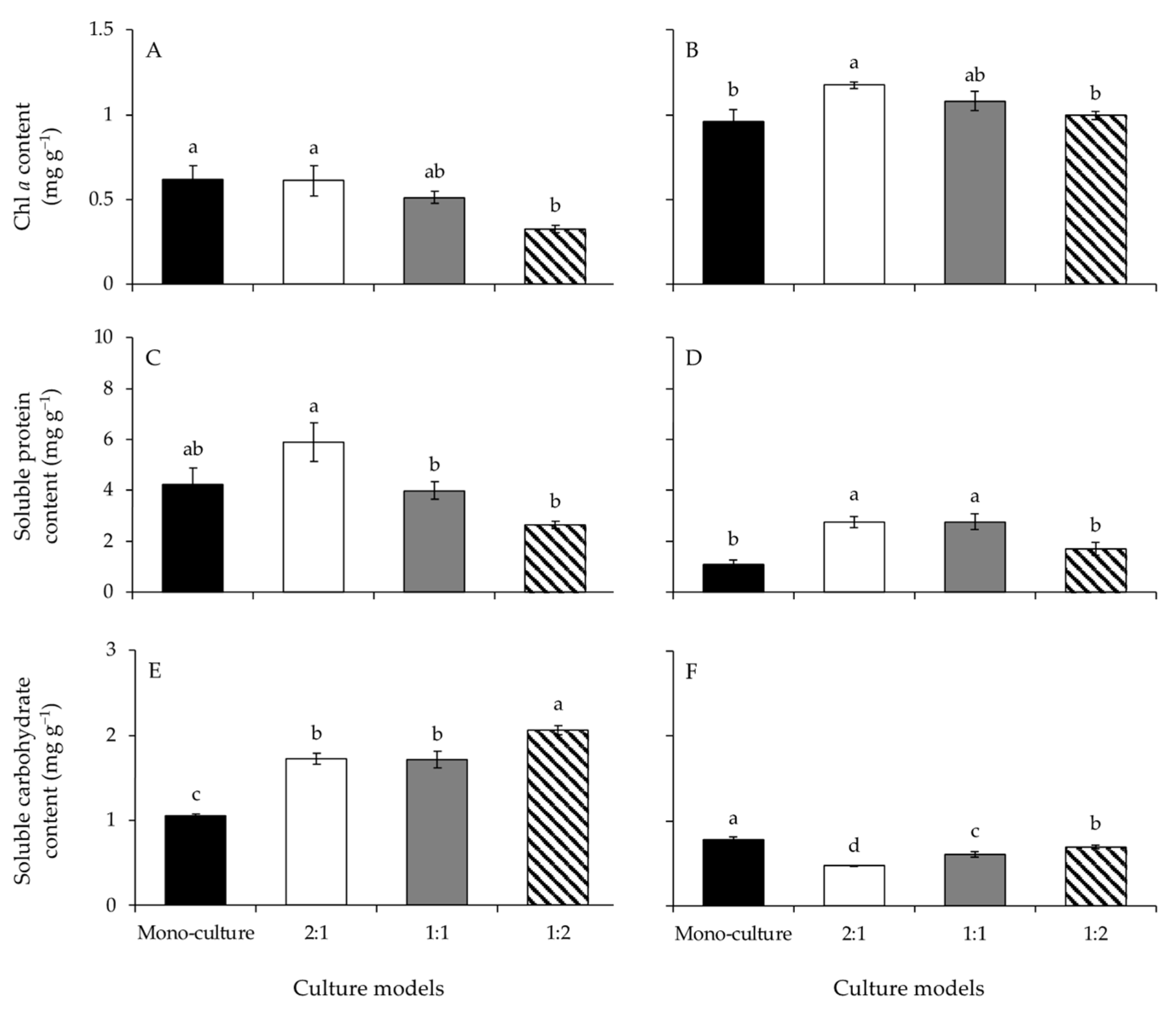
The Chl a contents of G. lemaneiformis differed significantly among culture models (Figure 4A and Table 1). The Chl a content at BDR of 1:2 was significantly lower than at mono-culture and BDR of 2:1. Similarly, the Chl a contents of S. horneri differed significantly among culture models (Figure 4B and Table 1). The Chl a content at BDR of 2:1 was significantly greater than at mono-culture and at BDR of 1:2.
3.5. Soluble Protein and Carbohydrate
The one-way ANOVA results showed a significant difference among culture models on the soluble protein contents of G. lemaneiformis (Figure 4C and Table 1). The soluble protein content at BDR of 2:1 was significantly greater than at BDRs of 1:1 and 1:2. The soluble protein contents of S. horneri differed significantly among culture models (Figure 4D and Table 1). The soluble protein contents at BDRs of 2:1 and 1:1 were significantly greater than at mono-culture and BDR of 1:2.
A significant effect of the culture model on the soluble carbohydrate contents of G. lemaneiformis was detected (Figure 4E and Table 1). Compared to the mono-culture, the soluble carbohydrate content had significantly higher values at co-culture with different BDRs. Moreover, the soluble carbohydrate content at BDR of 1:2 was significantly greater than at BDRs of 2:1 and 1:1. Similarly, the soluble carbohydrate contents of S. horneri varied significantly among culture models (Figure 4F and Table 1). The soluble carbohydrate content was significantly higher at mono-culture than co-culture with different BDRs. The soluble carbohydrate contents increased significantly with increasing biomass of S. horneri, from BDR of 2:1 to 1:2.
4. Discussion
Diverse positive and negative interspecific interactions exist among plants in natural communities [50]. The principal mechanisms of these interactions, such as resource competition or allelopathy, are often species-specific and environment-dependent [51,52]. In this study, the RGR of G. lemaneiformis was significantly enhanced under co-culture with BDR of 2:1 compared to under mono-culture, indicating that the presence of S. horneri with a relatively lower biomass had a stimulating impact on G. lemaneiformis. A similar finding was documented for G. lemaneiformis and epiphytic U. prolifera, showing that the RGR of U. prolifera was greatly increased under co-culture with BDR of 1:1 [53]. This ‘hormesis’ effect may have resulted from the division and expansion of cells through the enzymatic or non-enzymatic processes following low-dose phytotoxin stimulation, thereby further increasing the growth of thalli [54,55]. Additionally, this may be due to an overcompensation response following the initial damage caused by low concentrations of phytotoxins [56,57]. Therefore, we speculated that the secretion of allelochemicals played a more critical role in interspecific interactions than resource competition when G. lemaneiformis suffered from a low biomass value of S. horneri. Moreover, allelopathy presents complex mechanisms that can influence a variety of physiological and biochemical processes [58]. In this study, we observed a significant increase in the net photosynthetic rate, NO3-N uptake rate, and soluble protein content of G. lemaneiformis under co-culture with BDR of 2:1. Similarly, Pan et al. [53] showed that, compared to mono-culture, the Chl a content and photosynthetic rate of G. lemaneiformis were significantly enhanced under co-culture with 0.5 g L−1 of U. prolifera. Further experiments are required to clarify the correlative mechanisms of these findings.
Compared to the stimulating effect of S. horneri on G. lemaneiformis with a relatively lower biomass, the RGR at BDR of 1:2 was significantly lower than at mono-culture. Similarly, Xie et al. [59] demonstrated that the initial population density rate greatly influences the interaction of Heterosigma akashiwo (H) and Prorocentrum donghaiense (P). The growth of H. akashiwo was restrained by P. donghaiense at the inoculation proportion of 1H:4P, whereas H. akashiwo possessed a higher growth rate at the inoculation proportion of 4H:1P. Both nutrient competition and allelopathy have been demonstrated to play a critical role in interspecific interaction between these two microalgal species. In addition, the net photosynthetic rate, Chl a content, and soluble protein content of G. lemaneiformis significantly decrease at BDR of 1:2. A similar result was presented in [24], where the growth, photosynthetic activity, and biochemical metabolism of N. yezoensis were significantly inhibited by epiphytic U. prolifera, mainly as a result of nutrition and light competition. On the other hand, high concentrations of allelochemicals may lead to cell expansion and rupture, while cells can maintain osmotic pressure and intracellular homeostasis by accumulating soluble carbohydrates [60,61,62]. In the current study, the soluble carbohydrate content of G. lemaneiformis presented a significant increase at BDR of 1:2, indicating that S. horneri may have exerted a strong allelopathic effect on G. lemaneiformis under this condition. Therefore, the inhibitory effect of S. horneri with a relatively higher biomass on G. lemaneiformis may be associated with a combination of resource competition and allelopathy.
Compared to the mono-culture, the RGR, net photosynthetic rate, NO3-N uptake rate, Chl a content, and soluble protein content of S. horneri were enhanced under co-culture conditions with G. lemaneiformis at different BDRs. Macroalgal species with a stronger growth capacity usually have advantages in interspecific competition scenarios [51,63]. Our results showed that S. horneri (7.23%) has a better growth rate than G. lemaneiformis (3.53%) under mono-culture conditions. The outstanding growth performance of S. horneri may be correlated with its adaptability to frequent marine environment changes during the long-distance floating processes [64]. Additionally, U. lactuca showed greater growth and maximal photosynthetic ability than G. lemaneiformis under competitive conditions, benefiting from its larger frond surface area and higher nutrient uptake rate [65]. All these results indicate that S. horneri has a dominant role in its competition with G. lemaneiformis because of its physiological and morphological advantages. In addition, we observed that all physiological and biochemical parameters of S. horneri increased significantly with increasing biomass of G. lemaneiformis under co-culture conditions. Wang et al. [66] reported that co-culture with a small quantity of G. lemaneiformis thalli was significantly beneficial to the growth of the red tide species H. akashiwo. Similarly, G. lemaneiformis can stimulate the growth of S. costatum by secreting various types of allelochemicals [67]. Therefore, we suggest that certain allelochemicals produced by G. lemaneiformis may partially contribute to the increased growth rate of S. horneri.
The golden tide caused by Sargassum species has become a major ecological threat to the coastal environment and aquaculture production. According to the present data, although the occurrence of small quantities of S. horneri can stimulate the growth of G. lemaneiformis, its outbreak has a greatly harmful impact on the growth and biochemical metabolism of G. lemaneiformis, resulting in reduced yield and quality. Inversely, the growth of S. horneri is enhanced by the occurrence of G. lemaneiformis, regardless of BDR. This observation suggests that G. lemaneiformis cultivation has no inhibitory effect on golden tide, rather favoring its further expansion. Hence, S. horneri should be removed or intercepted in time to minimize economic and ecological losses. Due to the limited availability of data in this study, more studies involving related physiological and molecular mechanisms are required to evaluate the coping strategies of G. lemaneiformis against golden tide outbreaks.
Author Contributions
Investigation, data curation, visualization, writing—original draft, H.S.; conceptualization, methodology, project administration, funding acquisition, writing—review and editing, Y.L. and X.G.; formal analysis, resources, J.L.; validation, supervision, Q.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Key R&D Program of China (No. 2020YFD0900201) and the Fundamental Research Funds for the Central Universities of China (No. 842212015).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data in this study are available from the corresponding author upon reasonable request.
Acknowledgments
We are grateful to Qiaohan Wang of Ocean University of China for experimental assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Komatsu, T.; Tatsukawa, K.; Filippi, J.B.; Sagawa, T.; Matsunaga, D.; Mikami, A.; Ishida, K.; Ajisaka, T.; Tanaka, K.; Aoki, M.; et al. Distribution of drifting seaweeds in eastern East China Sea. J. Mar. Syst. 2007, 67, 245–252. [Google Scholar] [CrossRef]
- Schell, J.M.; Goodwin, D.S.; Siuda, A.N.S. Recent Sargassum inundation events in the Caribbean: Shipboard observations reveal dominance of a previously rare form. Oceanography 2015, 28, 8–10. [Google Scholar] [CrossRef]
- Xing, Q.G.; Guo, R.H.; Wu, L.L.; An, D.Y.; Cong, M.; Qin, S.; Li, X.R. High-resolution satellite observations of a new hazard of golden tides caused by floating Sargassum in winter in the Yellow Sea. IEEE Geosci. Remote Sens. Lett. 2017, 14, 1815–1819. [Google Scholar] [CrossRef]
- García-Sánchez, M.; Graham, C.; Vera, E.; Escalante-Mancera, E.; Álvarez-Filip, L.; van Tussenbroek, B.I. Temporal changes in the composition and biomass of beached pelagic Sargassum species in the Mexican Caribbean. Aquat. Bot. 2020, 167, 103275. [Google Scholar] [CrossRef]
- Lapointe, B.E. A comparison of nutrient-limited productivity in Sargassum natans from neritic vs. oceanic waters of the western North Atlantic Ocean. Limnol. Oceanogr. 1995, 40, 625–633. [Google Scholar] [CrossRef]
- Huffard, C.L.; von Thun, S.; Sherman, A.D.; Sealey, K.; Smith, K.L. Pelagic Sargassum community change over a 40-year period: Temporal and spatial variability. Mar. Biol. 2014, 161, 2735–2751. [Google Scholar] [CrossRef]
- Hu, Z.M.; Uwai, S.; Yu, S.H.; Komatsu, T.; Ajisaka, T.; Duan, D.L. Phylogeographic heterogeneity of the brown macroalga Sargassum horneri (Fucaceae) in the northwestern Pacific in relation to late Pleistocene glaciation and tectonic configurations. Mol. Ecol. 2011, 20, 3894–3909. [Google Scholar] [CrossRef]
- Xu, M.; Sakamoto, S.; Komatsu, T. Attachment strength of the subtidal seaweed Sargassum horneri (Turner) C. Agardh varies among development stages and depths. J. Appl. Phycol. 2016, 28, 3679–3687. [Google Scholar] [CrossRef]
- Sun, J.Z.; Chen, W.D.; Zhuang, D.G.; Zheng, H.Y.; Li, L.; Pang, S.J. In situ ecological studies of the subtidal brown alga Sargasssum horneri at Nanji Island of China. S. China Fish. Sci. 2008, 4, 58–63, (In Chinese with English abstract). [Google Scholar]
- Qi, L.; Hu, C.M.; Wang, M.Q.; Shang, S.L.; Wilson, C. Floating algae blooms in the East China Sea. Geophys. Res. Lett. 2017, 44, 11501–11509. [Google Scholar] [CrossRef]
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Rivera, E.; Flores-Díaz, M.; Hawkins, A. A fish kill coincident with dense Sargassum accumulation in a tropical bay. Bull. Mar. Sci. 2015, 91, 455–456. [Google Scholar] [CrossRef]
- Solarin, B.B.; Bolaji, D.A.; Fakayode, O.S.; Akinnigbagbe, R.O. Impacts of an invasive seaweed Sargassum hystrix var. fluitans (Børgesen 1914) on the fisheries and other economic implications for the Nigerian coastal waters. IOSR J. Agric. Vet. Sci. 2014, 7, 1–6. [Google Scholar]
- Milledge, J.J.; Harvey, P.J. Golden tides: Problem or golden opportunity? The valorisation of Sargassum from beach inundations. J. Mar. Sci. Eng. 2016, 4, 60. [Google Scholar] [CrossRef]
- Wang, X.M.; Shan, T.F.; Pang, S.J.; Su, L. Assessment of the genetic relationship between the recently established benthic population and the adjacent floating populations of Sargassum horneri (Phaeophyceae) in Dalian of China by newly developed trinucleotide microsatellite markers. J. Appl. Phycol. 2019, 31, 3989–3996. [Google Scholar] [CrossRef]
- Dickie, I.A.; Schnitzer, S.A.; Reich, P.B.; Hobbie, S.E. Spatially disjunct effects of co-occurring competition and facilitation. Ecol. Lett. 2005, 8, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Narwani, A.; Bentlage, B.; Alexandrou, M.A.; Fritschie, K.J.; Delwiche, C.; Oakley, T.H.; Cardinale, B.J. Ecological interactions and coexistence are predicted by gene expression similarity in freshwater green algae. J. Ecol. 2017, 105, 580–591. [Google Scholar] [CrossRef]
- Cameron, H.; Coulson, T.; Marshall, D.J. Size and density mediate transitions between competition and facilitation. Ecol. Lett. 2019, 22, 1879–1888. [Google Scholar] [CrossRef]
- Choi, H.G.; Norton, T.A. Competition and facilitation between germlings of Ascophyllum nodosum and Fucus vesiculosus. Mar. Biol. 2005, 147, 525–532. [Google Scholar] [CrossRef]
- White, L.F.; Shurin, J.B. Density dependent effects of an exotic marine macroalga on native community diversity. J. Exp. Mar. Biol. Ecol. 2011, 405, 111–119. [Google Scholar] [CrossRef]
- Gavina, M.K.A.; Tahara, T.; Tainaka, K.; Ito, H.; Morita, S.; Ichinose, G.; Okabe, T.; Togashi, T.; Nagatani, T.; Yoshimura, J. Multi-species coexistence in Lotka–Volterra competitive systems with crowding effects. Sci. Rep. 2018, 8, 1198. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Lin, L.; Ma, Z.; Zhang, T.; Chen, W.; Zou, D. Carbon and nitrogen accumulation and interspecific competition in two algae species, Pyropia haitanensis and Ulva lactuca, under ocean acidification conditions. Aquac. Int. 2019, 27, 721–733. [Google Scholar] [CrossRef]
- Li, X.S.; Xu, J.T.; He, P.M. Comparative research on inorganic carbon acquisition by the macroalgae Ulva prolifera (Chlorophyta) and Pyropia yezoensis (Rhodophyta). J. Appl. Phycol. 2016, 28, 491–497. [Google Scholar] [CrossRef]
- Sun, J.Y.; Bao, M.L.; Xu, T.P.; Li, F.T.; Wu, H.L.; Li, X.S.; Xu, J.T. Elevated CO2 influences competition for growth, photosynthetic performance and biochemical composition in Neopyropia yezoensis and Ulva prolifera. Algal Res. 2021, 56, 102313. [Google Scholar] [CrossRef]
- Sukenik, A.; Eshkol, R.; Livne, A.; Hadas, O.; Rom, M.; Tchernov, D.; Vardi, A.; Kaplan, A. Inhibition of growth and photosynthesis of the dinoflagellate Peridinium gatunense by Microcystis sp. (Cyanobacteria): A novel allelopathic mechanism. Limnol. Oceanogr. 2002, 47, 1656–1663. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Z.M.; Song, X.X.; Tang, X.X.; Zhang, S.D. Effects of macroalgae Ulva pertusa (Chlorophyta) and Gracilaria lemaneiformis (Rhodophyta) on growth of four species of bloom-forming dinoflagellates. Aquat. Bot. 2007, 86, 139–147. [Google Scholar] [CrossRef]
- He, D.; Liu, J.; Hao, Q.; Ran, L.H.; Zhou, B.; Tang, X.X. Interspecific competition and allelopathic interaction between Karenia mikimotoi and Dunaliella salina in laboratory culture. Chin. J. Oceanol. Limnol. 2016, 34, 301–313. [Google Scholar] [CrossRef]
- Yuan, C.Y.; Zhao, Y.; Yang, S.; Cui, Q.M. Allelopathic effect of Sargassum fusiforme on growth of Karenia mikimotoi. Appl. Mech. Mater. 2014, 522–524, 721–724. [Google Scholar] [CrossRef]
- Patil, V.; Abate, R.; Yang, Y.F.; Zhang, J.W.; Lin, H.N.; Chen, C.P.; Liang, J.R.; Sun, L.; Li, X.S.; Gao, Y.H. Allelopathic effect of Pyropia haitanensis (Rhodophyta) on the bloom-forming Skeletonema costatum (Bacillariophyta). J. Appl. Phycol. 2020, 32, 1275–1286. [Google Scholar] [CrossRef]
- Steentoft, M.; Irvine, L.M.; Farnham, W.F. Two terete species of Gracilaria and Gracilariopsis (Gracilariales, Rhodophyta) in Britain. Phycologia 1995, 34, 113–127. [Google Scholar] [CrossRef]
- Yoshida, T. Marine Algae of Japan; Uchida Rokakuho Publishing Company: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Iyer, R.; De Clerck, O.; Bolton, J.J.; Coyne, V.E. Morphological and taxonomic studies of Gracilaria and Gracilariopsis species (Gracilariales, Rhodophyta) from South Africa. S. Afr. J. Bot. 2004, 70, 521–539. [Google Scholar] [CrossRef]
- Qi, Z.H.; Liu, H.M.; Li, B.; Mao, Y.Z.; Jiang, Z.J.; Zhang, J.H.; Fang, J.G. Suitability of two seaweeds, Gracilaria lemaneiformis and Sargassum pallidum, as feed for the abalone Haliotis discus hannai Ino. Aquaculture 2010, 300, 189–193. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, Q.F.; Huan, Z.Y.; Zhong, M.Q.; Chen, W.Z.; Du, H. Identification and characterization of glutamine synthetase isozymes in Gracilaria lemaneiformis. Aquat. Bot. 2018, 146, 23–30. [Google Scholar] [CrossRef]
- Yuan, S.L.; Duan, Z.H.; Lu, Y.N.; Ma, X.L.; Wang, S. Optimization of decolorization process in agar production from Gracilaria lemaneiformis and evaluation of ntioxidant activities of the extract rich in natural pigments. 3 Biotech 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Fishery Bureau of Agriculture Ministry of China. Chinese Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2021; pp. 23–28. (In Chinese)
- Yang, Y.F.; Chai, Z.Y.; Wang, Q.; Chen, W.Z.; He, Z.L.; Jiang, S.J. Cultivation of seaweed Gracilaria in Chinese coastal waters and its contribution to environmental improvements. Algal Res. 2015, 9, 236–244. [Google Scholar] [CrossRef]
- Kim, J.K.; Yarish, C.; Hwang, E.K.; Park, M.; Kim, Y. Seaweed aquaculture: Cultivation technologies, challenges and its ecosystem services. Algae 2017, 32, 1–13. [Google Scholar] [CrossRef]
- Gao, X.; Kim, J.H.; Park, S.K.; Yu, O.H.; Kim, Y.S.; Choi, H.G. Diverse responses of sporophytic photochemical efficiency and gametophytic growth for two edible kelps, Saccharina japonica and Undaria pinnatifida, to ocean acidification and warming. Mar. Pollut. Bull. 2019, 142, 315–320. [Google Scholar] [CrossRef]
- Li, S.F.; Liu, Y.; Gong, Q.L.; Gao, X.; Li, J.Y. Physiological and ultrastructural responses of the brown seaweed Undaria pinnatifida to triphenyltin chloride (TPTCL) stress. Mar. Pollut. Bull. 2020, 153, 110978. [Google Scholar] [CrossRef]
- Liu, C.X.; Zou, D.H.; Yang, Y.F.; Chen, B.B.; Jiang, H. Temperature responses of pigment contents, chlorophyll fluorescence characteristics, and antioxidant defenses in Gracilariopsis lemaneiformis (Gracilariales, Rhodophyta) under different CO2 levels. J. Appl. Phycol. 2017, 29, 983–991. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, Y.Q.; Liu, X.J.; Zhong, M.Q.; Chen, W.Z.; Wang, F.; Du, H. Response of Gracilaria lemaneiformis to nitrogen deprivation. Algal Res. 2018, 34, 82–96. [Google Scholar] [CrossRef]
- Yang, Y.L.; Li, W.; Li, Y.H.; Xu, N.J. Photophysiological responses of the marine macroalga Gracilariopsis lemaneiformis to ocean acidification and warming. Mar. Environ. Res. 2020, 163, 105204. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.R.; Pang, T.; Liu, J.G. Changes in the temperature tolerance profile of Gracilariopsis lemaneiformis from the perspective of photosynthesis, respiration, and biochemical markers after many years of vegetative propagation. J. Appl. Phycol. 2022, 34, 1045–1058. [Google Scholar] [CrossRef]
- Tatewaki, M. Formation of a crustaceous sporophyte with unilocular sporangia in Scytosiphon lomentaria. Phycologia 1966, 6, 62–66. [Google Scholar] [CrossRef]
- Li, S.; Liu, D. Improvement of spectrophotometric determination of phosphorus in water by phospho–molybdenum blue. Environ. Prot. Chem. Ind. 2006, 26, 78–80, (In Chinese with English abstract). [Google Scholar]
- Sun, X.Y.; Hong, L.C.; Ye, H.M. Experiment determining nitrate nitrogen in water samples by on-line cadmium column reduction-flow injection method. Water Resour. Prot. 2010, 26, 75–77, (In Chinese with English abstract). [Google Scholar]
- Kochert, G. Protein determination by dye binding. In Handbook of Phycological Methods. Physiological and Biochemical Methods; Hellebust, J.A., Craigie, J.S., Eds.; Cambridge University Press: Cambridge, UK, 1978; pp. 91–93. [Google Scholar]
- Tang, S.Q.; Gong, M.G.; Chen, D.C. Study of methods for determination of carbohydrates in sea water 2. The anthrone method for total amount of particulate carbohydrate. J. Shandong Coll. Oceanol. 1985, 15, 47–54. (In Chinese) [Google Scholar]
- Hirsch, A.M.; Bauer, W.D.; Bird, D.M.; Cullimore, J.; Tyler, B.; Yoder, J.I. Molecular signals and receptors: Controlling rhizosphere interactions between plants and other organisms. Ecology 2003, 84, 858–868. [Google Scholar] [CrossRef]
- Xu, D.; Gao, Z.Q.; Zhang, X.F.; Fan, X.; Wang, Y.T.; Li, D.M.; Wang, W.; Zhuang, Z.M.; Ye, N.H. Allelopathic interactions between the opportunistic species Ulva prolifera and the native macroalga Gracilaria lichvoides. PLoS ONE 2012, 7, e33648. [Google Scholar] [CrossRef]
- Bieberich, J.; Lauerer, M.; Drachsler, M.; Heinrichs, J.; Müller, S.; Feldhaar, H. Species- and developmental stage-specific effects of allelopathy and competition of invasive Impatiens glandulifera on cooccurring plants. PLoS ONE 2018, 13, e0205843. [Google Scholar] [CrossRef]
- Pan, Z.L.; Yu, Y.Y.; Chen, Y.L.; Yu, C.Z.; Xu, N.J.; Li, Y.H. Combined effects of biomass density and low-nighttime temperature on the competition for growth and physiological performance of Gracilariopsis lemaneiformis and Ulva prolifera. Algal Res. 2022, 62, 102638. [Google Scholar] [CrossRef]
- Bais, H.P.; Venkatachalam, L.; Biedrzycki, M.L. Stimulation or inhibition: Conflicting evidence for (±)-catechin’s role as a chemical facilitator and disease protecting agent. Plant Signal. Behav. 2010, 5, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Huang, G.H.; Fu, H.Y.; An, C.J.; Yao, Y.; Cheng, G.H.; Suo, M.Q. Allelopathy inhibitory effects of Hydrodictyon reticulatum on Chlorella pyrenoidosa under co-culture and liquor-cultured conditions. Water 2017, 9, 416. [Google Scholar] [CrossRef]
- Belz, R.G. Stimulation versus inhibition–bioactivity of parthenin, a phytochemical from Parthenium hysterophorus L. Dose-Response 2008, 6, 80–96. [Google Scholar] [CrossRef]
- Belz, R.G. Investigating a potential auxin–related mode of hermetic/inhibitory action of the phytotoxin parthenin. J. Chem. Ecol. 2016, 42, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Budzałek, G.; Śliwińska-Wilczewska, S.; Wiśniewska, K.; Wochna, A.; Bubak, I.; Latała, A.; Wiktor, J.M. Macroalgal defense against competitors and herbivores. Int. J. Mol. Sci. 2021, 22, 7865. [Google Scholar] [CrossRef]
- Xie, Z.H.; Xiao, H.; Cai, H.J.; Wang, R.J.; Tang, X.X. Influence of UV-B irradiation on the interspecific growth interaction between Heterosigma akashiwo and Prorocentrum donghaiense. Int. Rev. Hydrobiol. 2006, 91, 555–573. [Google Scholar] [CrossRef]
- Nagayama, K.; Shibata, T.; Fujimoto, K.; Honjo, T.; Nakamura, T. Algicidal effect of phlorotannins from the brown alga Ecklonia kurome on red tide microalgae. Aquaculture 2003, 218, 601–611. [Google Scholar] [CrossRef]
- Accoroni, S.; Percopo, I.; Cerino, F.; Romagnoli, T.; Pichierri, S.; Perrone, C.; Totti, C. Allelopathic interactions between the HAB dinoflagellate Ostreopsis cf. ovata and macroalgae. Harmful Algae 2015, 49, 147–155. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.Y.; Ge, F.J.; Liu, Q.; Zhang, Y.Y.; Zhou, Q.H.; Xu, D.; Wu, Z.B. Interspecific competition for nutrients between submerged macrophytes (Vallisneria natans, Ceratophyllum demersum) and filamentous green algae (Cladophora oligoclona) in a co-culture system. Pol. J. Environ. Stud. 2019, 28, 1483–1494. [Google Scholar] [CrossRef]
- Zhang, S.D.; Yu, Z.M.; Song, X.X.; Song, F.; Wang, Y. Competition about nutrients between Gracilaria lemaneiformis and Prorocentrum donghaiense. Acta Ecol. Sin. 2005, 25, 2676–2680, (In Chinese with English abstract). [Google Scholar]
- Liu, Z.Y.; Sun, P.; Qin, S.; Li, J.J.; Zhuang, L.C.; Song, W.L.; Bi, Y.X.; Zhong, Z.H. Comparative analysis of growth and photosynthetic characteristics in benthic and floating Sargassum horneri. Chin. J. Ecol. 2021, 40, 76–83, (In Chinese with English abstract). [Google Scholar]
- Chen, B.B.; Zou, D.H.; Jiang, H. Elevated CO2 exacerbates competition for growth and photosynthesis between Gracilaria lemaneiformis and Ulva lactuca. Aquaculture 2015, 443, 49–55. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, B.; Tang, X. Effects of two species of macroalgae—Ulva pertusa and Gracilaria lemaneiformis—On growth of Heterosigma akashiwo (Raphidophyceae). J. Appl. Phycol. 2009, 21, 375–385. [Google Scholar] [CrossRef]
- Lu, H.M.; Xie, H.H.; Gong, Y.X.; Wang, Q.; Yang, Y.F. Secondary metabolites from the seaweed Gracilaria lemaneiformis and their allelopathic effects on Skeletonema costatum. Biochem. Syst. Ecol. 2011, 39, 397–400. [Google Scholar] [CrossRef]
Figure 1.
The RGRs of Gracilariopsis lemaneiformis (A) and Sargassum horneri (B) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.
Figure 1.
The RGRs of Gracilariopsis lemaneiformis (A) and Sargassum horneri (B) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.
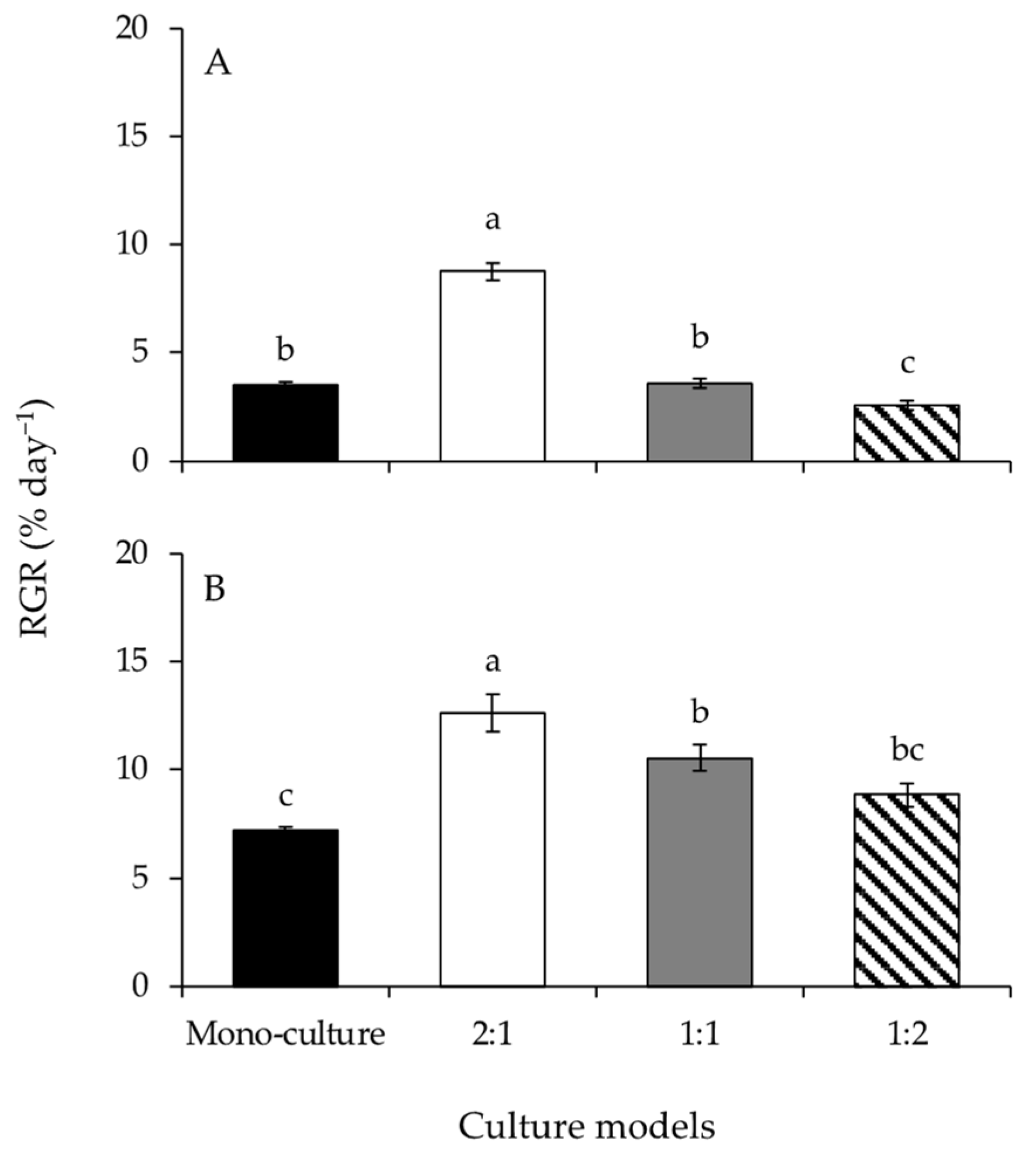
Figure 2.
The net photosynthetic rates of Gracilariopsis lemaneiformis (A) and Sargassum horneri (B) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.
Figure 2.
The net photosynthetic rates of Gracilariopsis lemaneiformis (A) and Sargassum horneri (B) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.
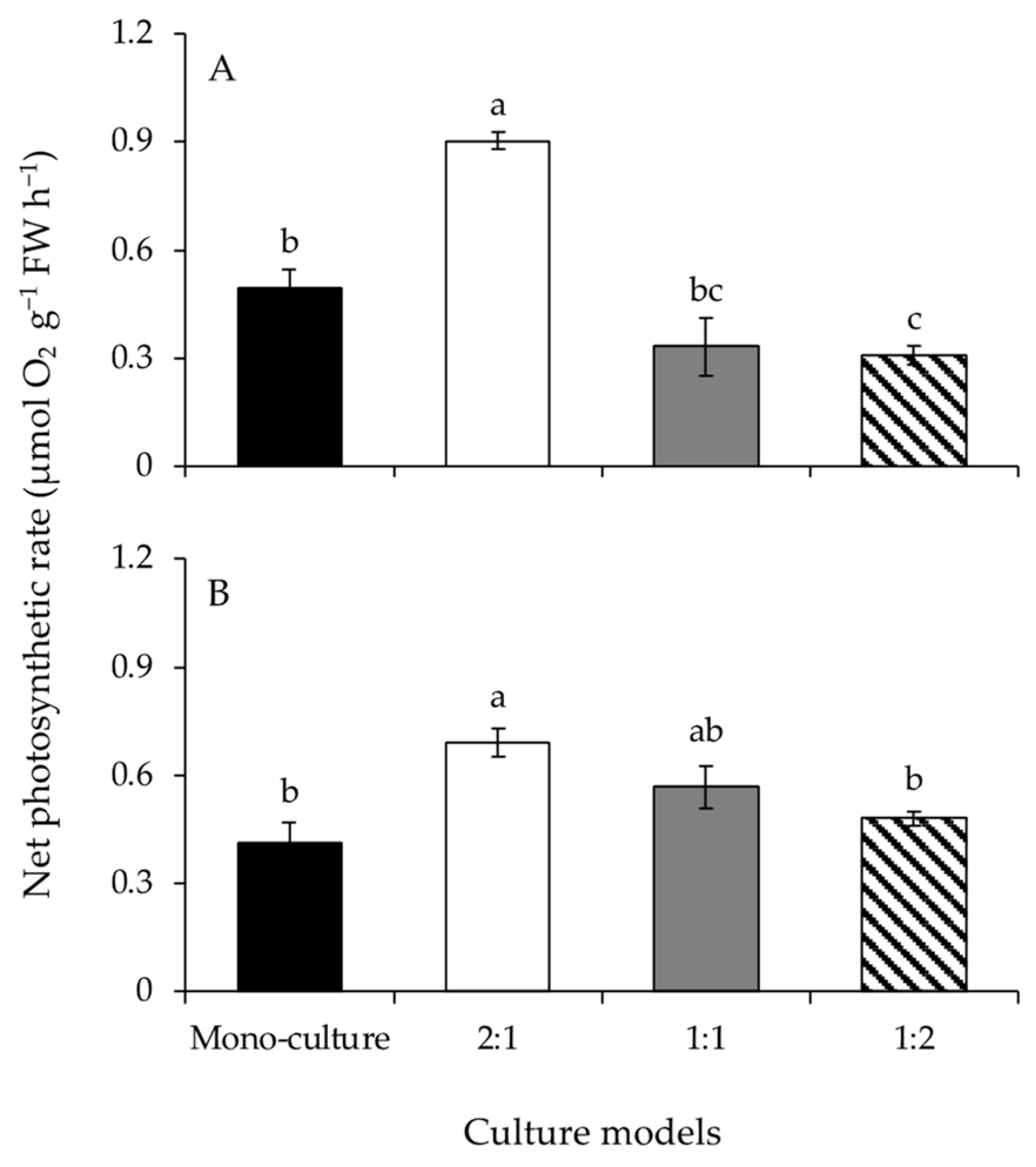
Figure 3.
The NO3-N uptake rates of Gracilariopsis lemaneiformis (A) and Sargassum horneri (B) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.
Figure 3.
The NO3-N uptake rates of Gracilariopsis lemaneiformis (A) and Sargassum horneri (B) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.
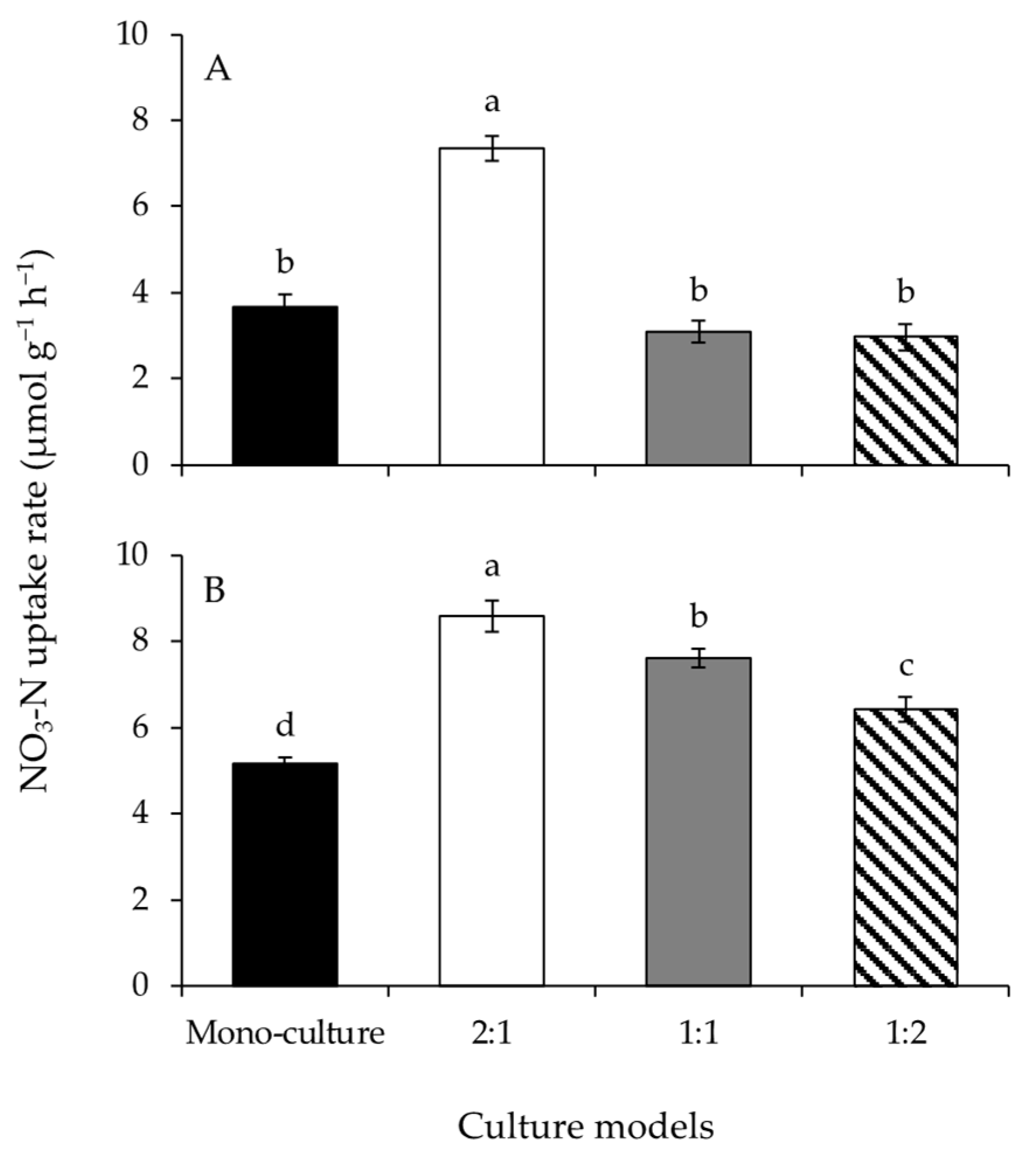
Figure 4.
The contents of Chl a, soluble protein, and soluble carbohydrate of Gracilariopsis lemaneiformis (A,C,E) and Sargassum horneri (B,D,F) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.
Figure 4.
The contents of Chl a, soluble protein, and soluble carbohydrate of Gracilariopsis lemaneiformis (A,C,E) and Sargassum horneri (B,D,F) cultured for twelve days at mono-culture and co-culture with biomass density ratios of 2:1, 1:1, and 1:2. Different lowercase letters represent significant differences (p < 0.05) among culture models.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
One-way ANOVA testing of the effects of biomass density ratio on RGR, net photosynthetic rate, NO3-N uptake rate, Chl a, soluble protein, and carbohydrate of Gracilariopsis lemaneiformis and Sargassum horneri.
Table 1.
One-way ANOVA testing of the effects of biomass density ratio on RGR, net photosynthetic rate, NO3-N uptake rate, Chl a, soluble protein, and carbohydrate of Gracilariopsis lemaneiformis and Sargassum horneri.
| Factors | Gracilariopsis lemaneiformis | Sargassum horneri | ||||
|---|---|---|---|---|---|---|
| df | F | p | df | F | p | |
| RGR | 3 | 112.721 | <0.001 | 3 | 16.595 | <0.001 |
| Net photosynthetic rate | 3 | 29.975 | <0.001 | 3 | 6.836 | <0.05 |
| NO3-N uptake rate | 3 | 52.763 | <0.001 | 3 | 32.004 | <0.001 |
| Chl a | 3 | 4.510 | <0.05 | 3 | 4.157 | <0.05 |
| Soluble protein | 3 | 6.398 | <0.05 | 3 | 11.253 | <0.01 |
| Soluble carbohydrate | 3 | 39.980 | <0.001 | 3 | 33.784 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Song, H.; Liu, Y.; Li, J.; Gong, Q.; Gao, X. Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions. Water 2022, 14, 2664. https://doi.org/10.3390/w14172664
AMA Style
Song H, Liu Y, Li J, Gong Q, Gao X. Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions. Water. 2022; 14(17):2664. https://doi.org/10.3390/w14172664
Chicago/Turabian StyleSong, Hanmo, Yan Liu, Jingyu Li, Qingli Gong, and Xu Gao. 2022. "Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions" Water 14, no. 17: 2664. https://doi.org/10.3390/w14172664
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.