
Revealing the Coral Species Diversity in Xiamen Bay: Spatial Distribution of Genus Astrogorgia (Cnidaria, Alcyonacea, Plexauridae) and Newly Recorded Species


Abstract
:1. Introduction
2. Materials and Methods
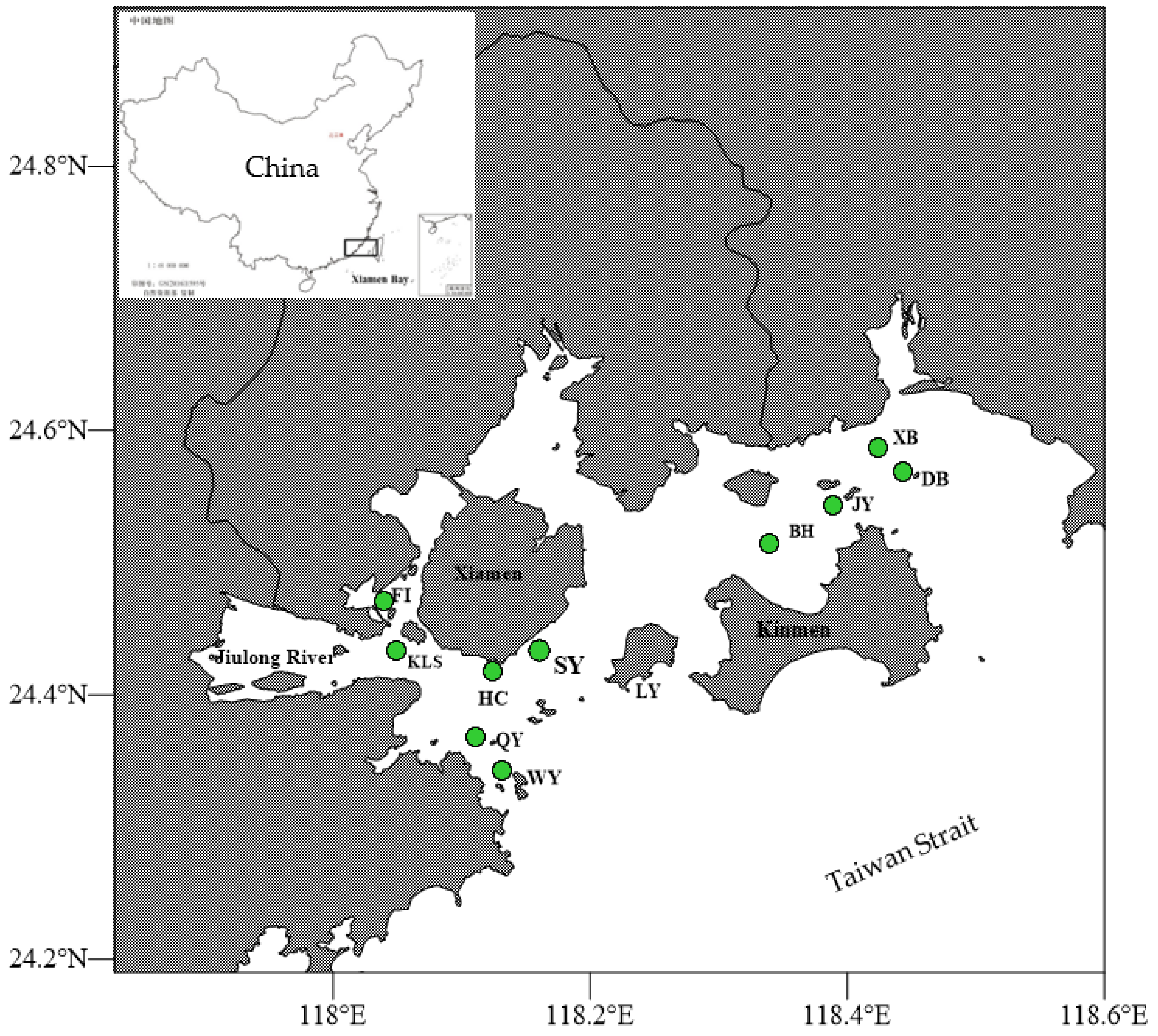
2.1. Sample Collection and Study Area
2.2. Sample Processing and Observation
2.3. Species Identification
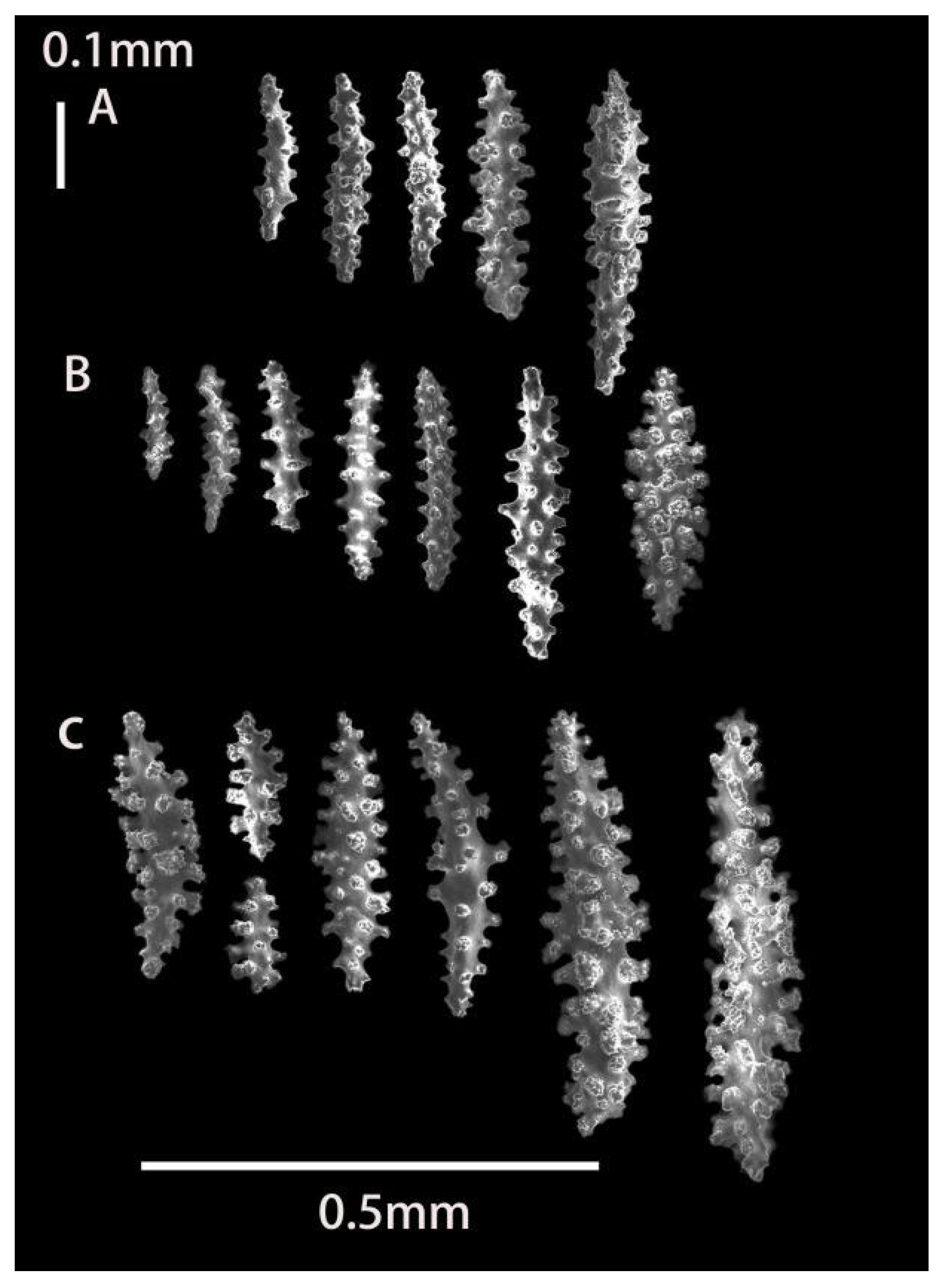
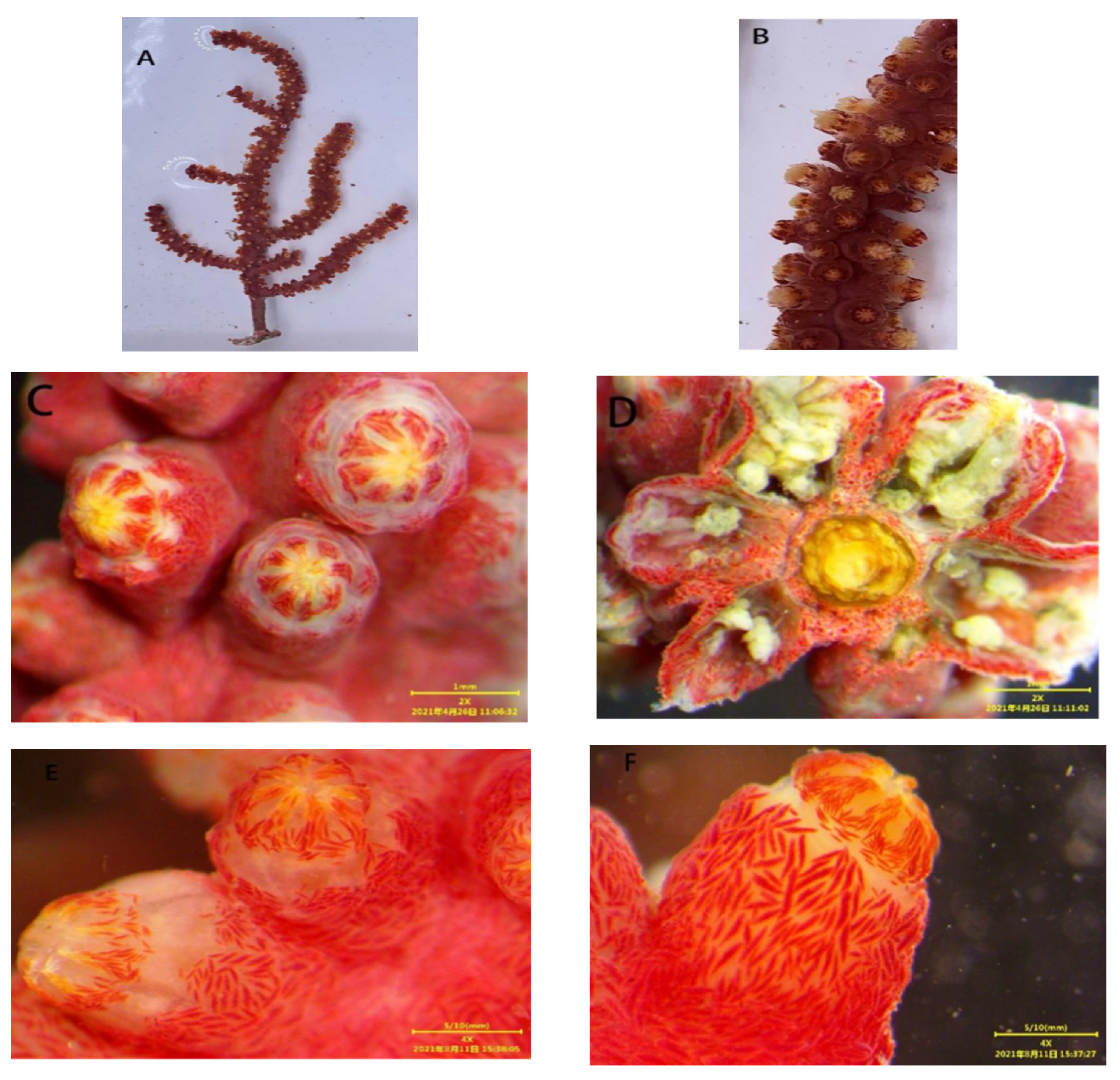
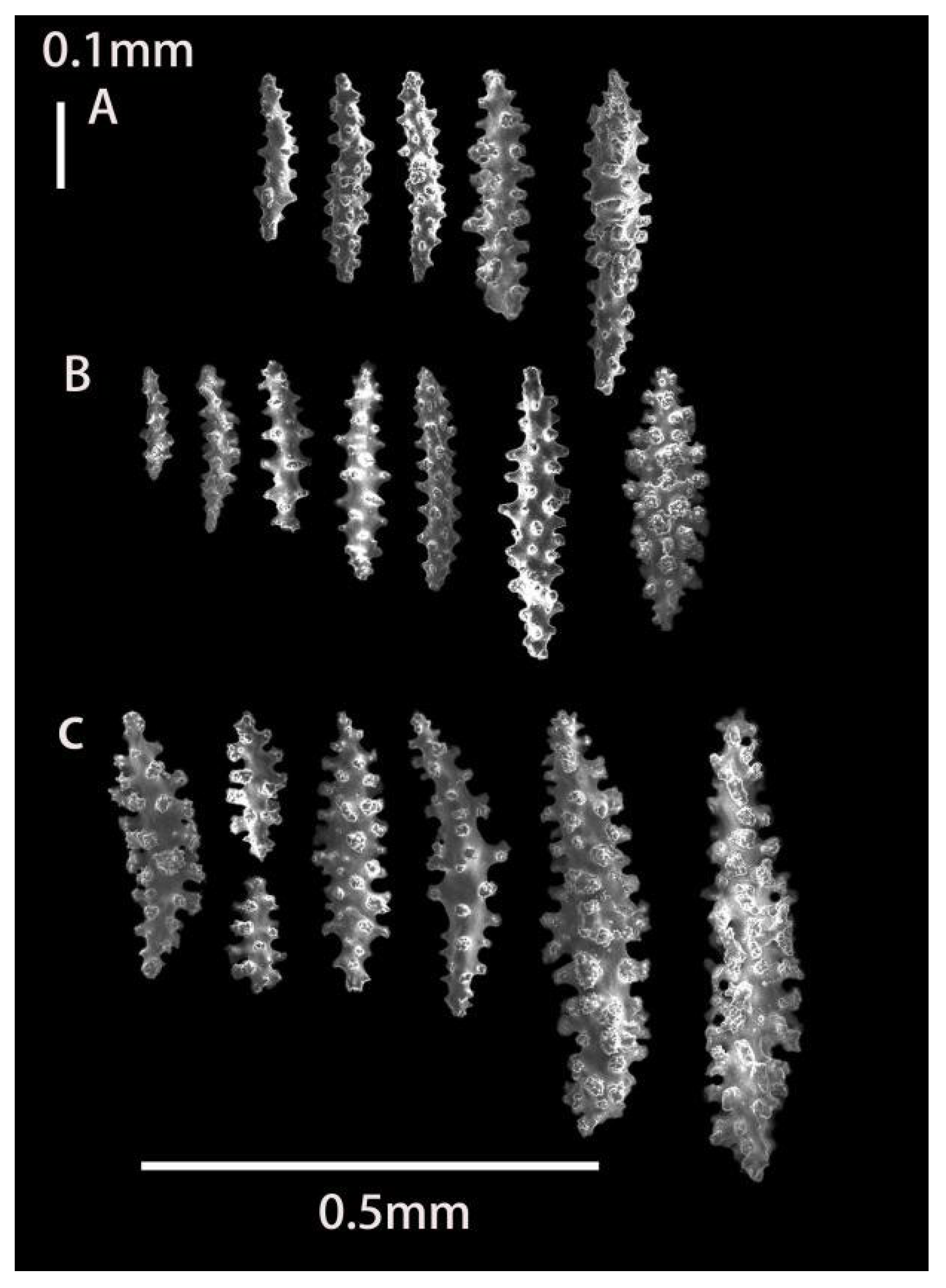
2.3.1. Morphology
2.3.2. DNA Extraction and PCR Amplification
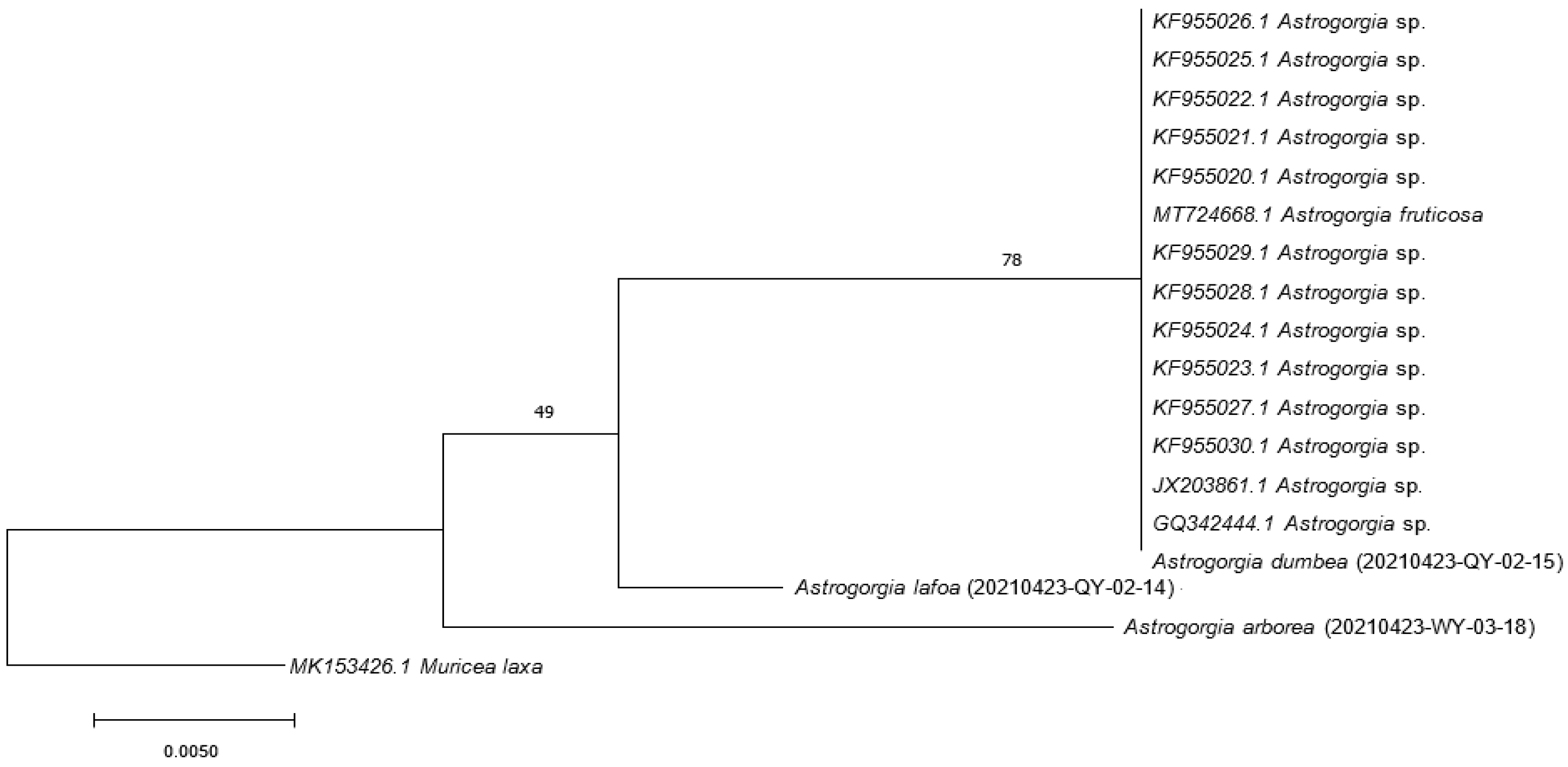
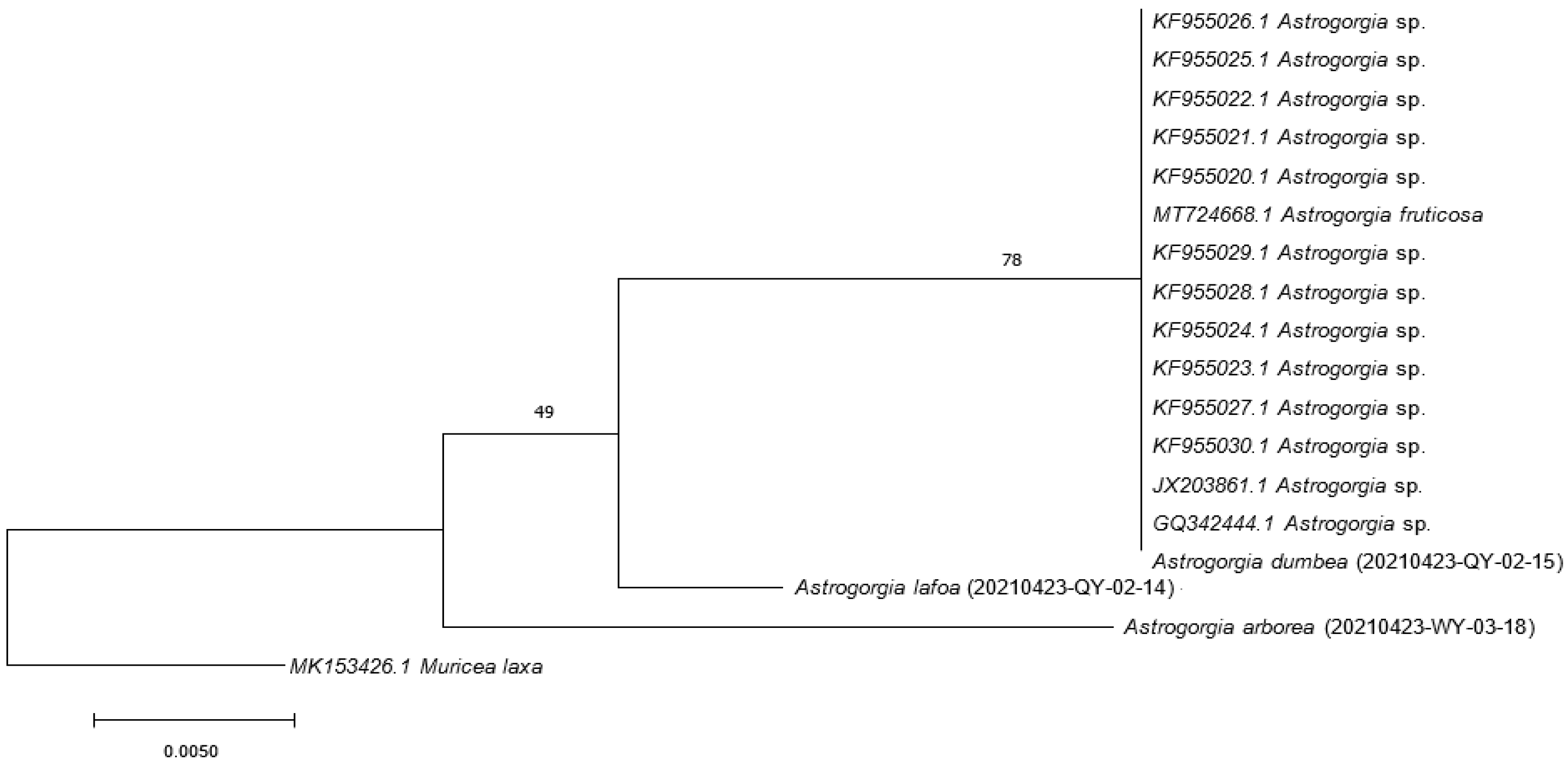
2.3.3. Cladogram Topology
3. Results
3.1. Systematic
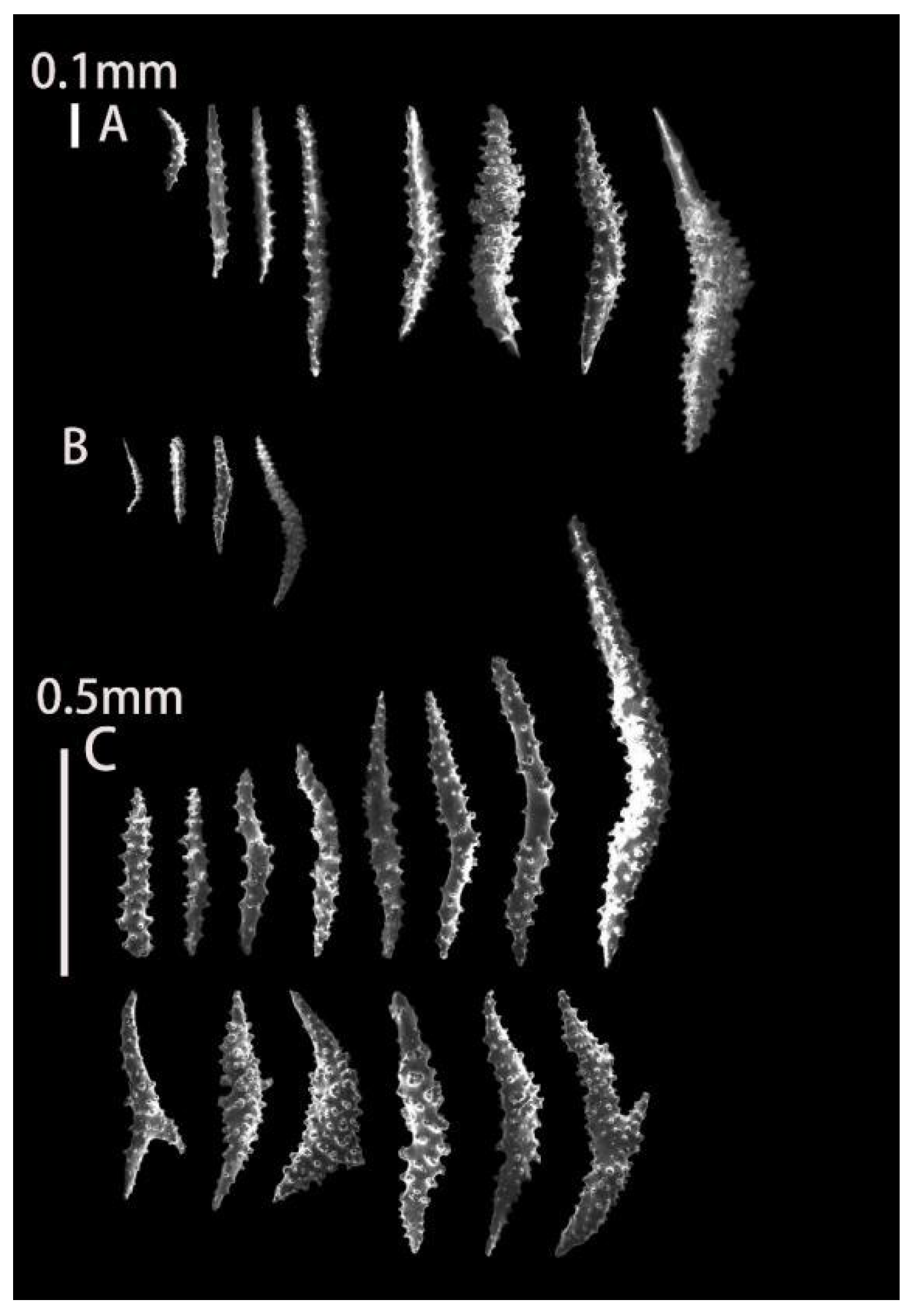
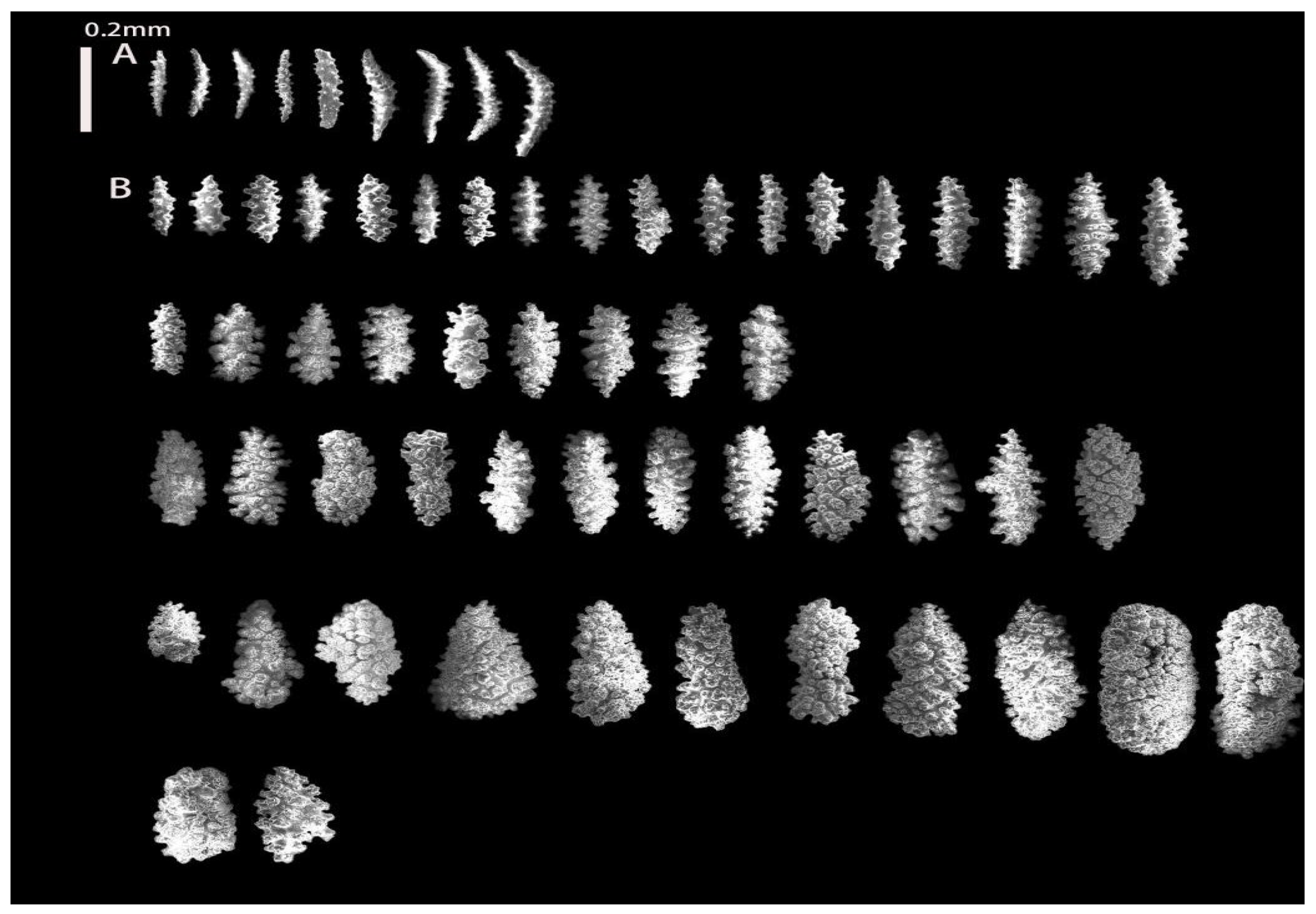
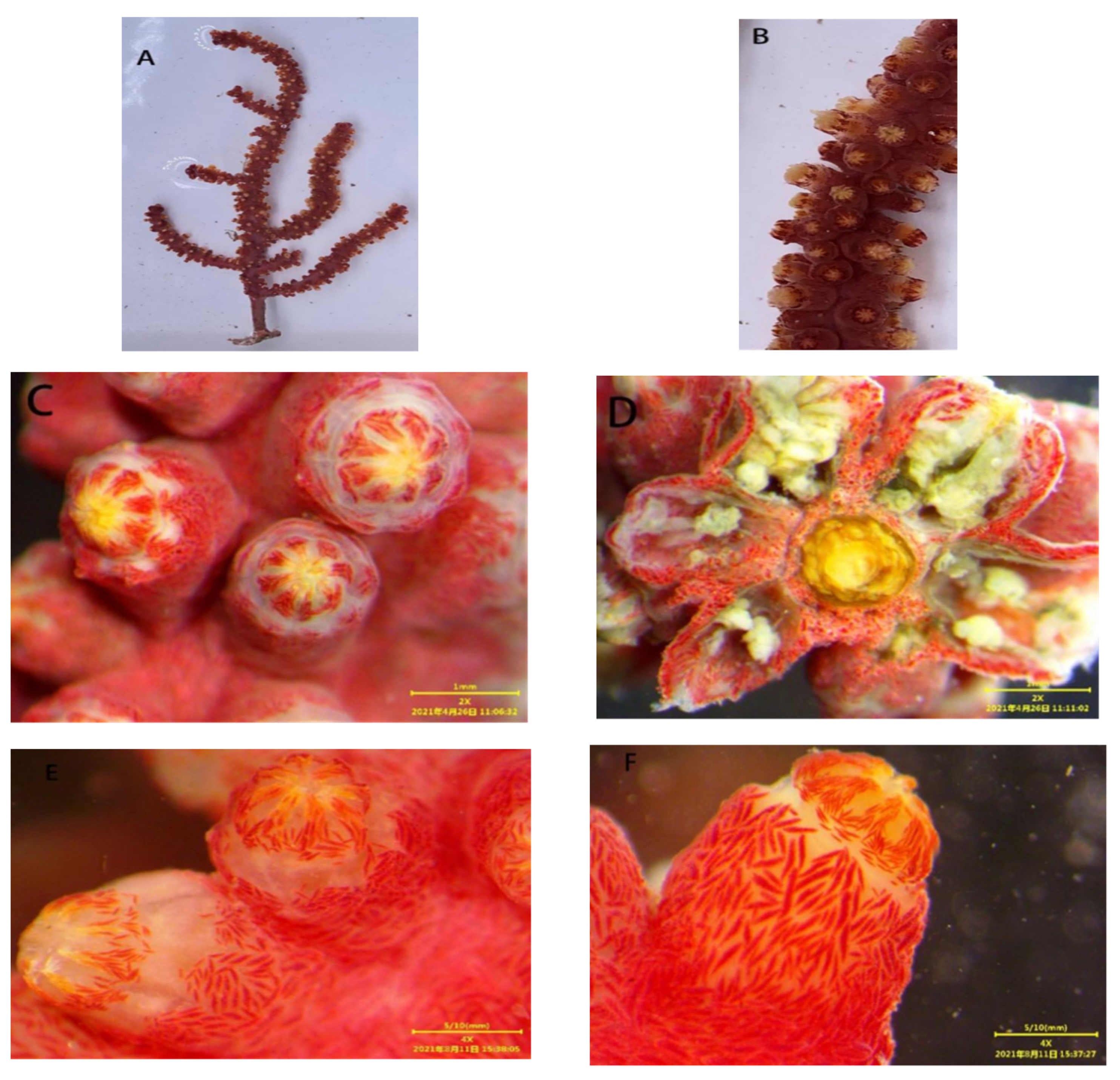
3.1.1. Astrogorgia lafoa Grasshoff, 1999
3.1.2. Astrogorgia arborea (Thomson & Simpson, 1909)
3.1.3. Astrogorgia dumbea Grasshoff, 1999
3.2. DNA Analysis
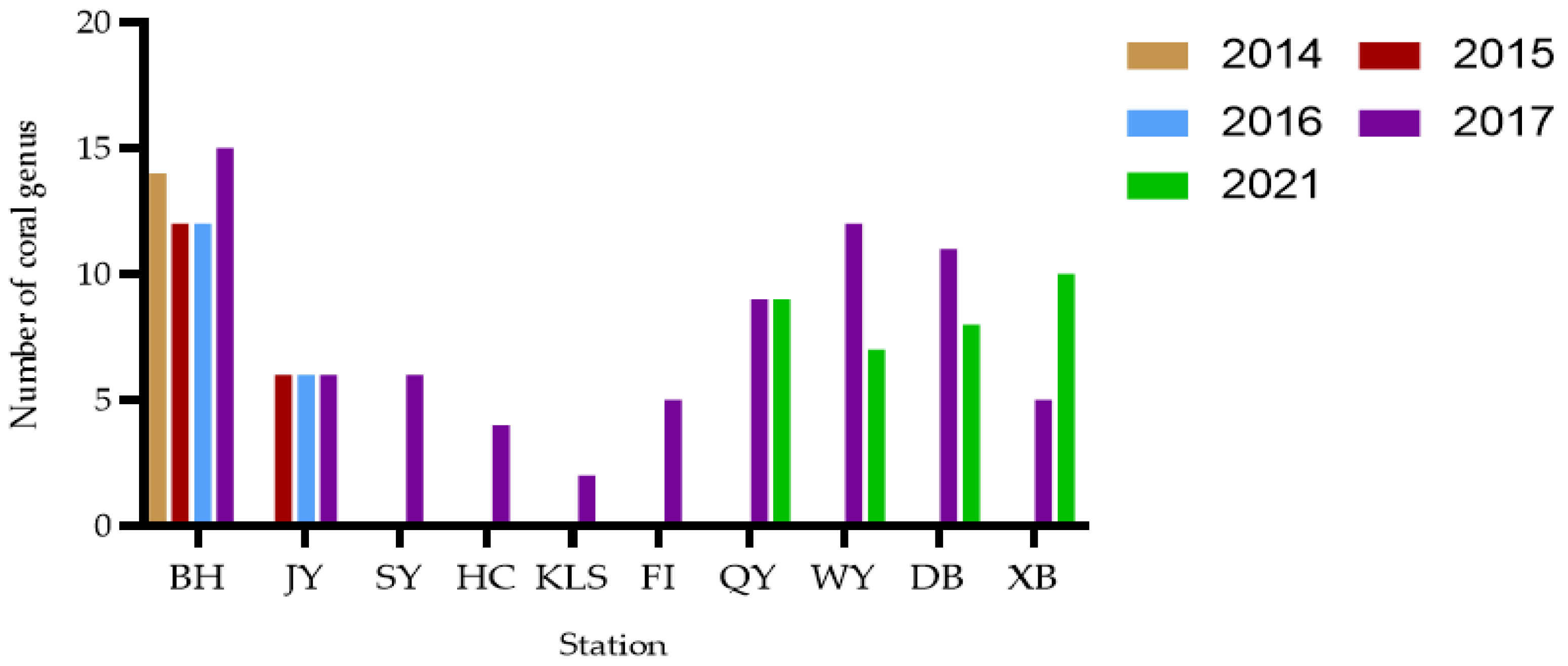
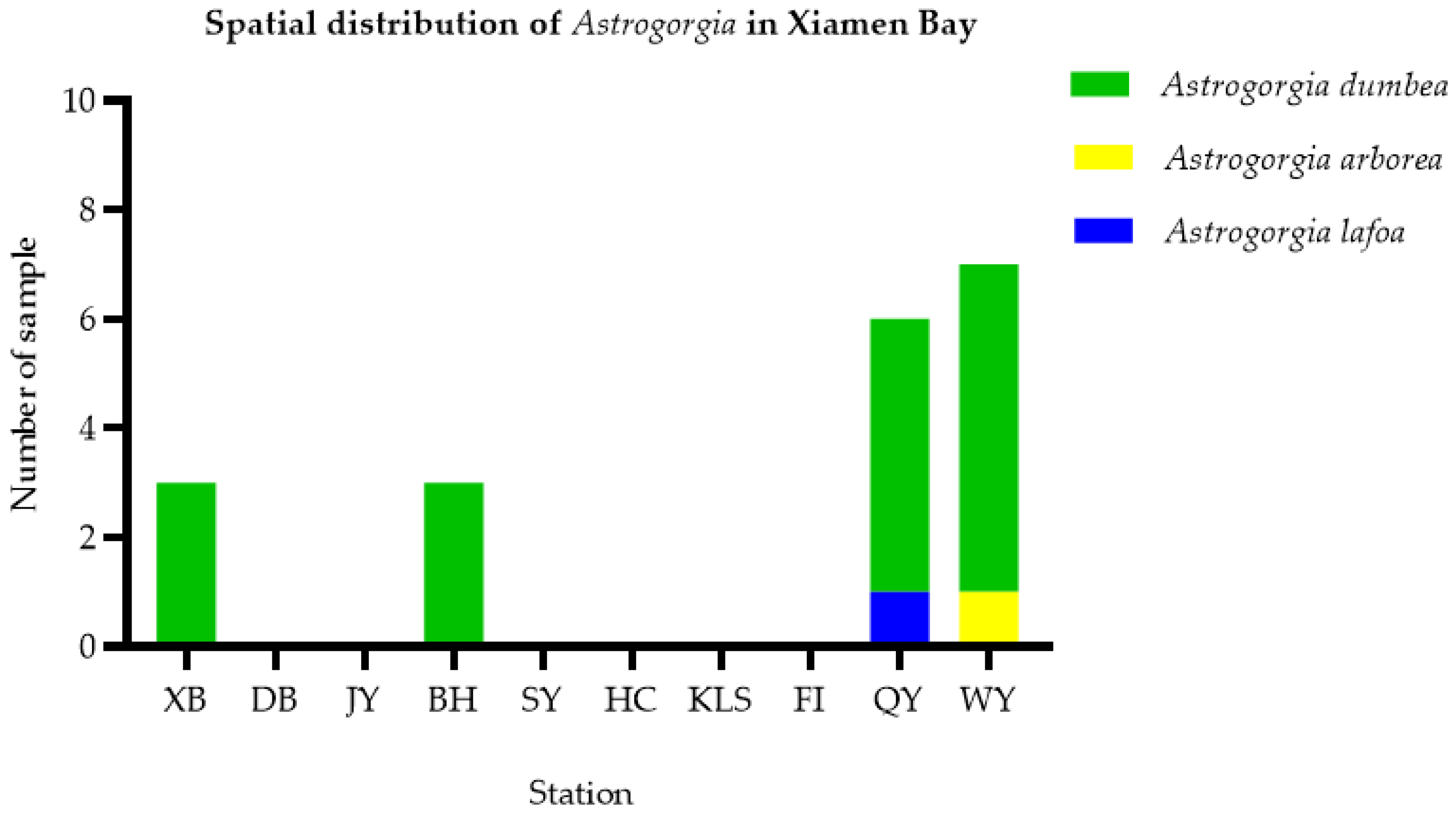
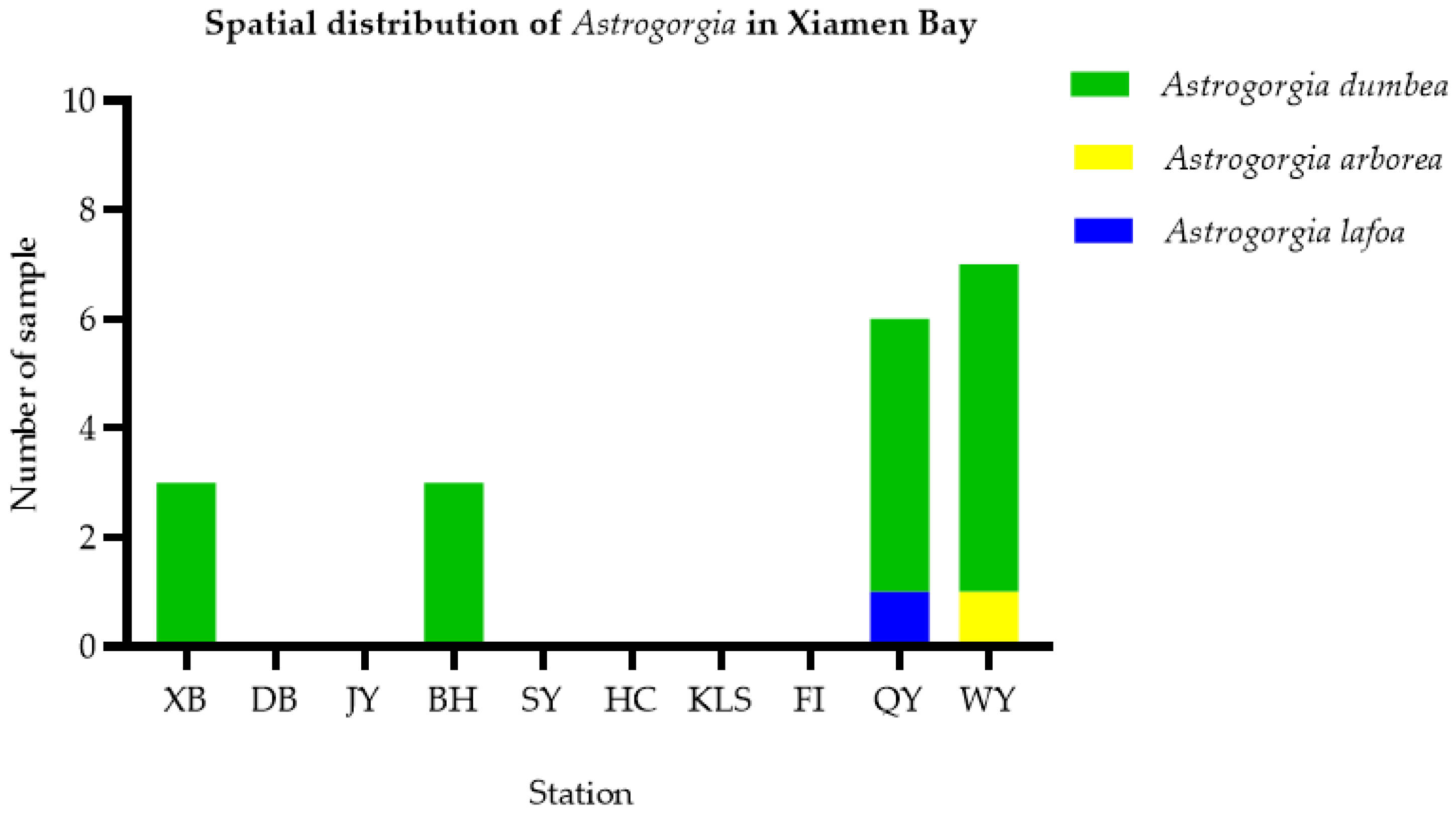
3.3. Geographical Distribution and Habit Characteristics of Astrogorgia
4. Discussion
4.1. Historical Evolution Process of Family-Level and Genus-Level Taxonomy of Some Species of Astrogorgia
4.2. Species Identification of Astrogorgia from Two Aspects of Morphology and Molecular Biology
4.3. Distribution and Habitat Analysis
4.4. Management Recommendations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Sixth Status of Corals of the World: 2020 Report. Available online: https://gcrmn.net/2020-report/ (accessed on 2 June 2022).
- Zhao, H.T.; Wang, L.R. The general characteristics of the coral reef ecosystem. Chin. J. Ecol. 2001, 20, 41–45. (In Chinese) [Google Scholar]
- Zhao, H.T.; Wang, L.R.; Yuan, J.Y. Sustainable development of the coral reefs in the South China Sea Islands. Trop. Geogr. 2016, 36, 55–65. (In Chinese) [Google Scholar] [CrossRef]
- Spalding, M.D.; Ravilious, C.; Green, E.P. World. Atlas of Coral Reefs; University of California Press: Berkeley, CA, USA, 2001. [Google Scholar]
- Zou, R.L. Fauna Sinica: Coelenterata. Anthozoa. Scleractinia. Hermatypic Coral; Science Press: Beijing, China, 2001. (In Chinese) [Google Scholar]
- Liu, R.Y. Catalogue of Marine Life in China; Science Press: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Huang, L.T.; Huang, H.; Jiang, L. A revised taxonomy for Chinese hermatypic corals. Biodiv. Sci. 2020, 28, 515–523. [Google Scholar] [CrossRef]
- Yang, S.L.; Yang, L.; Zhao, D.B.; Ren, Y.S. Species composition and distribution of neritic Scleractinian and Gorgonian corals in coastal waters of Fujian. J. Appl. Oceanogr. 2015, 34, 209–218. (In Chinese) [Google Scholar]
- Huang, Z.G. Diversity of Species in Xiamen Bay, China; Ocean Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Ni, Z. Study on Investigation and Taxonomy of Coral resources in Baiha Reef; Jimei University: Xiamen, China, 2018. (In Chinese) [Google Scholar]
- Verrill, A.E. Critical remarks on the halcyonoid polyps in the Museum of Yale College, with descriptions of new genera. Ame. J. Sci. Arts 1868, 45, 411–415. [Google Scholar]
- Verrill, A.E. Synopsis of the polyps and corals of the North Pacific Exploring Expedition, 1853–1856, with descriptions of some additional species from the West Coast of North America. III Madreporaria. Salem Commun. Essex Inst. 1866, 5, 17–32. [Google Scholar]
- Alderslade, P.A.; Fabricius, K. Soft Corals and Sea Fans; Australian Institute of Marine Science: Crawley, Australia, 2008.
- Horton, T.; Kroh, A.; Ahyong, S.; Bailly, N.; Boury-Esnault, N.; Brandão, S.N.; Mees, J. World Register of Marine Species (WoRMS); WoRMS Editorial Board, 2018. Available online: https://www.gbif.org/publisher/d1f6b74b-1d53-44db-bed5-d08497095900 (accessed on 15 June 2022).
- Lu, Y.-N.; Cui, P.; Tian, X.-Q.; Lou, L.-G.; Fan, C.-Q. Unusual Cytotoxic Steroidal Saponins from the Gorgonian Astrogorgia dumbea. Planta Med. 2016, 82, 882–887. [Google Scholar] [CrossRef] [Green Version]
- Li, X.B.; Huang, H.; Lian, J.S.; Yang, J.H.; Huang, L.M. Diversity and spatial distribution of gorgonian coral in Dongshan-coastal waters in Fujian, China. J. Oceanogr. Taiwan Strait 2011, 30, 92–96. (In Chinese) [Google Scholar]
- Dai, C.F. Taxonomic morphologies and taxonomic systems of Octocorallia. In A Complete Map of Corals in Taiwan: Eight Corals, 1st ed.; Li, J.H., Hu, J.Y., Eds.; Owl: Taiwan, China, 2022; pp. 1–408. (In Chinese) [Google Scholar]
- Wang, L.H.; Sheu, J.H.; Kao, S.Y.; Su, J.H.; Chen, Y.H.; Chen, Y.H.; Sung, P.J. Natural product chemistry of gorgonian corals of the family Plexauridae distributed in the Indo-Pacific Ocea. Mar. Drugs 2012, 10, 2415–2434. [Google Scholar] [CrossRef] [Green Version]
- Berrue, F.; Kerr, R.G. Diterpenes from gorgonian corals. Nat. Prod. Rep. 2009, 26, 681–710. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Yu, S.; van Ofwegen, L.; Totzke, F.; Proksch, P.; Lin, W. 9,10-Secosteroids, protein kinase inhibitors from the Chinese gorgonian Astrogorgia sp. Bioorg. Med. Chem. 2011, 19, 6873–6880. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, Y.W. A pharmaceutical perspective on gorgonian: Chemistry and bioactivity. Chin. J. Nat. Med. 2003, 1, 69–75. [Google Scholar]
- Kandou, F.E.F.; Maabuat, P.V.; Katili, D.Y. Seleksi Aktivitas Antibakteri dari Beberapa Gorgonia dari Perairan Manado Ter-hadap Bakteri Staphylococcus aureus, Escherichia coli, dan Edwarsiella tarda. J. MIPA 2019, 8, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Sibero, M.T.; Zilda, D.S.; Haryanti, D.; Igarashi, Y. Astrogorgia sp. from Saparua, Maluku: Phytochemical Content, Antimicrobial, Antioxidant, and Cytotoxicity Properties. IOP Conf. Series. Earth Environ. Sci. 2021, 750, 12062. [Google Scholar] [CrossRef]
- Hay, M.E.; Kappel, Q.E.; Fenical, W. Synergisms in Plant Defenses against Herbivores: Interactions of Chemistry, Calcification, and Plant Quality. Ecology 1994, 75, 1714–1726. [Google Scholar] [CrossRef]
- Paul, V.J.; Hay, M.E. Seaweed susceptibility to herbivory: Chemical and morphological correlates. Mar. Ecol. Prog. Ser. 1986, 33, 255–264. [Google Scholar] [CrossRef]
- Pennings, S.C.; Puglisi, M.P.; Pitlik, T.J.; Himaya, A.C.; Paul, V.J. Effects of secondary metabolites and CaCO3 on feeding by surgeonfishes and parrotfishes: Within-plant comparisons. Mar. Ecol. Prog. Ser. 1996, 134, 49–58. [Google Scholar] [CrossRef]
- Puglisi, M.P.; Paul, V.J.; Biggs, J.; Slattery, M. Co-occurrence of chemical and structural defenses in the gorgonian corals of Guam. Mar. Ecol. Prog. Ser. 2002, 239, 105–114. [Google Scholar] [CrossRef]
- Wirshing, H.H.; Messing, C.G.; Douady, C.J.; Reed, J.; Stanhope, M.J.; Shivji, M.S. Molecular evidence for multiple lineages in the gorgonian family Plexauridae (Anthozoa: Octocorallia). Mar. Biol. 2005, 147, 497–508. [Google Scholar] [CrossRef]
- Kim, M.-S.; Song, J.-I. Four New Records of Holaxonia (Anthozoa: Octocorallia: Alcyonacea) from Korea. Anim. Syst. Evol. Divers. 2014, 30, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Breedy, O.; Guzman, H.M. A revision of the genus Muricea Lamouroux, 1821 (Anthozoa, Octocorallia) in the eastern Pacific. Part I: Eumuricea Verrill, 1869 Revisited. ZooKeys 2015, 537, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, A.K.; van Ofwegen, L.P. Species of Elasmogorgia and Euplexaura (Cnidaria, Octocorallia) from Japan with a discussion about the genus Filigella. ZooKeys 2016, 589, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohr, K.E.; Ripple, K.; Patterson, J.T. Differential disturbance effects and phenotypic plasticity among out planted corals at patch and fore reef sites. J. Nat. Conserv. 2020, 55, 125827. [Google Scholar] [CrossRef]
- Bayer, F.M. Octocorallia. In Treatise on Invertebrate Paleontology, Part, F., Coelenterata; Moore, R.C., Ed.; University of Kansas Press: Lawrence, KS, USA, 1956; pp. 166–189. [Google Scholar]
- Deichmann, E. The Alcyonaria of the Western Part of the Atlantic Ocean; Printed for the Museum: Cambridge, MA, USA, 1936. [Google Scholar] [CrossRef]
- Kükenthal, W. Gorgonaria; Das Tierreich 47; Overview of the genus Briareum in the Indo-Pacific; W. de Gruyter & Co.: Berlin, Germany; Leipzig, Germany, 1924; 478p. [Google Scholar]
- Brockman, S.A.; McFadden, C.S. The Mitochondrial Genome of Paraminabea aldersladei (Cnidaria: Anthozoa: Octocorallia) Supports Intramolecular Recombination as the Primary Mechanism of Gene Rearrangement in Octocoral Mitochondrial Genomes. Genome Biol. Evol. 2012, 4, 994–1006. [Google Scholar] [CrossRef] [Green Version]
- McFadden, C.S.; France, S.C.; Sánchez, J.A.; Alderslade, P. A molecular phylogenetic analysis of the Octocorallia (Cnidaria: Anthozoa) based on mitochondrial protein-coding sequences. Mol. Phylogenetics Evol. 2006, 41, 513–527. [Google Scholar] [CrossRef]
- Wang, P.Z.; Yang, C.J.; Wang, H.S.; Li, W.D. Preliminary identification of Aldersladum sp in the South China Sea. J. Hainan Trop. Ocean. Univ. 2018, 25, 18–22. (In Chinese) [Google Scholar]
- Xu, Y. Taxonomy and Phylogeny of the Golden Coral Family Chrysogorgiidae (Anthozoa: Alcyonacea) from Seamounts in the Western Pacific; University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Sciences): Beijing, China, 2019. (In Chinese) [Google Scholar]
- Li, X.B.; Huang, H.; Zhang, J.B.; Dong, Z.J.; Huang, L.M. Phylogenetic Relationships with the Gorgonian (Cndaria: Anthozoa: Octocorallia) Based on 18S rDNA Sequences. Mar. Sci. Bull. 2006, 25, 10–19. (In Chinese) [Google Scholar]
- Grasshoff, M. The Shallow Water Gorgonians of New Caledonia and Adjacent Islands: (Coelenterata: Octocorallia); Sencken-bergische Naturforschende Gesellschaft: Frankfurt, Germany, 1999. [Google Scholar]
- Thomson, J.A.; Simpson, J.J. An account of The Alcyonarians Collected by the Royal Indian Marine Survey Ship ’Investigator’ in the Indian Ocean. II. The Alcyonarians of the Littoral Area; Trustees of the Indian Museum: Calcutta, India, 1909. [Google Scholar]
- Bayer, F.M. Key to the genera of Octocorallia exclusive of Pennatulacea (Coelenterata: Anthozoa), with diagnosis of new taxa. Proc. Biol. Soc. Wash. 1981, 94, 902–947. [Google Scholar]
- Stephen, D.C.; Herman, H.W. A phylogenetic analysis of the Primnoidae (Anthozoa: Octocorallia: Calcaxonia) with analyses of character evolution and a key to the genera and subgenera. BMC Evol. Biol. 2018, 18, 66. [Google Scholar]
- Hermanlimianto, M.; van Ofwegen, L. A new species of Astrogorgia (Coelenterata: Octocorallia: Plexauridae) from Bali. Zool. Meded. 2006, 80, 103–108. [Google Scholar]
- Ofwegen, L.V.; Hoeksema, B.W. Astrogorgia bayeri, a new gorgonian octocoral species from South Sulawesi (Coelenterata: Octocorallia: Plexauridae). Bull. Biol. Soc. Wash. 2001, 10, 66–70. [Google Scholar]
- Thomson, J.A.; Dean, L.M. The Alcyonacea of the Siboga-Expedition: With an addendum to the Gorgonacea; Siboga Expedition; E. J. Brill Publishing: Leiden, The Netherlands, 1931; pp. 1899–1900. [Google Scholar]
- Namin, K.S.; Van Ofwegen, L.P. Some shallow water octocorals (Coelenterata: Anthozoa) of the Persian Gulf. Zootaxa 2009, 2058, 1–52. [Google Scholar] [CrossRef]
- Grasshoff, M. The Gorgonians of the Sinai Coast and the Strait of Gubal, Red Sea (Coelenterata, Octocorallia); Courier Forschungsinstitut Senckenberg: Stuttgart, Germany, 2000. [Google Scholar]
- Thomson, J.A.; Henderson, W.D.; Simpson, J.J. An Account of the Alcyonarians Collected by the Royal Indian Marine Survey Ship Investigator in the Indian Ocean; Order of the Trustees of the Indian Museum: Calcutta, India, 1906. [Google Scholar]
- Verrill, A.E.; Edmondson, C.H. Hawaiian Shallow Water Anthozoa; The Museum: Honolulu, HI, USA, 1928. [Google Scholar] [CrossRef]
- Kükenthal, W.; Gorzawsky, H. Diagnosen neuer japanischer Gorgoniden (Reise Doflein 1904–05). Zool. Anzeiger 1908, 32, 621–631. [Google Scholar]
- Haywood, P.J.; Ryland, J.S. The Marine Fauna of the British Isles and the North West Europe. Vol 1. Introduction and Protozoans to Arthropods; Clarendon Press: Oxford, UK, 1990. [Google Scholar]
- Lamouroux, J.V.F. Exposition Méthodique des Genres de l’ordre des Polypiers; VVe Agasse: Paris, France, 1821. [Google Scholar]
- Hickson, S.J. The Gorgonacea of Panama Bay, Together with a Description of One Species from the Galapagos Islands and One from Trinidad; Dansk Naturhistorisk Forening: København, Denmark, 1928. [Google Scholar]
- Quattrini, A.M.; Wu, T.; Soong, K.; Jeng, M.S.; Benayahu, Y.; McFadden, C.S. A next generation approach to species delimi-tation reveals the role of hybridization in a cryptic species complex of corals. BMC Evol. Biol. 2019, 19, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Stoeckle, M.Y.; Zemlak, T.S.; Francis, C.M. Identification of Birds through DNA Barcodes. PLoS Biol. 2004, 2, e312. [Google Scholar] [CrossRef] [Green Version]
- Baco, A.R.; Cairns, S.D. Comparing Molecular Variation to Morphological Species Designations in the Deep-Sea Coral Narella Reveals New Insights into Seamount Coral Ranges. PLoS ONE 2012, 7, e45555. [Google Scholar] [CrossRef] [Green Version]
- Herrera, S.; Shank, T.M. RAD sequencing enables unprecedented phylogenetic resolution and objective species delimitation in recalcitrant divergent taxa. Mol. Phylogenetics Evol. 2016, 100, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Meier, R.; Todd, P.A.; Chou, L.M. More evidence for pervasive paraphyly in Scleractinia corals: Systematic study of Southeast Asian Faviidae (Cnidaria; Scleractinia) based on molecular and morphological data. Mol. Phylogenetics Evol. 2009, 50, 102–116. [Google Scholar] [CrossRef]
- Arrigoni, R.; Stefani, F.; Pichon, M.; Galli, P.; Benzoni, F. Molecular phylogeny of the Robust clade (Faviidae, Mussidae, Merulinidae, and Pectiniidae): An Indian Ocean perspective. Mol. Phylogenetics Evol. 2012, 65, 183–193. [Google Scholar] [CrossRef]
- Li, X.N.; Liu, L.; Liu, C.W. Phylogenetic relationship in 8 species of Scleractinia coral based on COI sequences. Prog. Fish. Sci. 2012, 33, 83–88. (In Chinese) [Google Scholar]
- Dai, C.F.; Qin, Q.X. Ecological Map of Bafang Coral in Dongsha, 1st ed.; Cai, N.S., Wu, D.Y., Eds.; Marine National Park Administration: Kaohsiung, Taiwan, 2017; pp. 10–11. (In Chinese)
- El-Naggar, H.A. Human Impacts on Coral Reef Ecosystem. In Natural Resources Management and Biological Sciences; Rhodes, E.R., Naser, H., Eds.; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Laura, E.D.; Stephanie, S.H.; Daniel, J.T. Conservation and management of ornamental coral reef wildlife: Successes, short-comings, and future directions. Biol. Conserv. 2014, 169, 225–237. [Google Scholar]
- National List of Nature Reserves-Nature Reserves in China. Available online: http://www.zrbhq.com.cn/index.php?m=content&c=index&a=lists&catid=81 (accessed on 15 June 2022).
- Huang, H.; Lian, J.S.; Li, Z.X.; Chen, Z.Q. Fujian Dongshan Coral Nature Reserve and Its Biodiversity; China Ocean Press: Beijing, China, 2009. (In Chinese) [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Station | Longitude | Latitude |
|---|---|---|---|
| 2014 | Baiha Reef (BH) | 118°22′06″–118°22′17″ | 24°31′39″–24°31′55″ |
| 2015 | Jiaoyu Island (JY) | 118°24′15″–118°24′40″ | 24°32′42″–24°33′22″ |
| Baiha Reef (BH) | 118°22′12″–118°22′17″ | 24°31′53″–24°31′55″ | |
| 2016 | Jiaoyu Island (JY) | 118°24′15″ | 24°33′40″ |
| Baiha Reef (BH) | 118°22′10″–118°22′17′ | 24°31′45″–24°31′51″ | |
| 2017 | Shangyu Island (SY) | 118°11′19″–118°11′25″ | 24°27′12″–24°27′13″ |
| Huangcuo (HC) | 118°07′50″ | 24°25′25″ | |
| Kulangsu Island (KLS) | 118°03′14″–118°03′41″ | 24°26′23″–24°26′35″ | |
| Fire Island (FI) | 118°03′54″ | 24°29′35″ | |
| Qingyu Island (QY) | 118°05′35″–118° 07′28″ | 24° 21′45″–24°21′55″ | |
| Wuyu Island (WY) | 118°03′54″–118° 08′35″ | 24° 20′23″–24° 29′35″ | |
| Dabai Island (DB) | 118°26′58″–118°27′40″ | 24°33′46″–24°34′9″ | |
| Xiaobai Island (XB) | 118°27′47″ | 24°33′21″ | |
| Jiaoyu Island (JY) | 118°24′14″ | 24°32′41″ | |
| Baiha Reef (BH) | 118°22′07″–118°22′17″ | 24°31′38″–24°31′55″ | |
| 2021 | Qingyu Island (QY) | 118°07′22″–118°07′50″ | 24°21′46″–24°22′11″ |
| Wuyu Island (WY) | 118°08′29″–118°08′57″ | 24°20′30″–24°20′51″ | |
| Dabai Island (DB) | 118°45′01″–118°46′06″ | 24°56′32″–24°57′51″ | |
| Xiaobai Island (XB) | 118°42′59″–118°43′39″ | 24°58′26″–24°58′96″ |
| Number | Species | GenBank Accession Number COI |
|---|---|---|
| 1 | Astrogorgia sp. | KF955026.1 |
| 2 | Astrogorgia sp. | KF955025.1 |
| 3 | Astrogorgia sp. | KF955022.1 |
| 4 | Astrogorgia sp. | KF955021.1 |
| 5 | Astrogorgia sp. | KF955020.1 |
| 6 | Astrogorgia fruticosa | MT724668.1 |
| 7 | Astrogorgia sp. | KF955029.1 |
| 8 | Astrogorgia sp. | KF955028.1 |
| 9 | Astrogorgia sp. | KF955024.1 |
| 10 | Astrogorgia sp. | KF955023.1 |
| 11 | Astrogorgia sp. | KF955027.1 |
| 12 | Astrogorgia sp. | KF955030.1 |
| 13 | Astrogorgia sp. | JX203861.1 |
| 14 | Astrogorgia sp. | GQ342444.1 |
| 15 | Astrogorgia lafoa * | ON748930 |
| 16 | Astrogorgia dumbea * | ON748931 |
| 17 | Astrogorgia arborea * | ON748932 |
| 18 | Muricea laxa | MK153426.1 |
| Number | Species | Distribution Area (Reference) |
|---|---|---|
| 1 | A. arborea | Indian Ocean [43] |
| 2 | A. balinensis | Bali [46] |
| 3 | A. bayeri | South Sulawesi [47] |
| 4 | A. begata | New Caledonia, the lagoon at 25–35 m [42] |
| 5 | A. canala | New Caledonia, 10–30 m [42] |
| 6 | A. dumbea | New Caledonia Lagoon 15–45 m [42]; Dongshan Bay, China [16] |
| 7 | A. filigella | Japan [48] |
| 8 | A. fruticosa | Some shallow waters in the Persian Gulf [49] |
| 9 | A. jiska | Sinai Coast and the Strait of Gubar in the Red Sea [50] |
| 10 | A. lafoa | New Caledonia, Cape St. Vincent near 70 m [42] |
| 11 | A. lea | Sinai coast and the Red Sea Gulbal Strait [50] |
| 12 | A. mengalia | New Caledonia, Eastern Reef 35–45 m [42] |
| 13 | A. milka | Sinai coast and the Red Sea Gulbal Strait [50] |
| 14 | A. ramosa | Andaman Sea, Indian Ocean 83–494 m deep [43]; |
| 15 | A. rubra | Deep sea [51] |
| 16 | A. sara | Sinai Coast and the Strait of Gubar in the Red Sea [50] |
| 17 | A. sinensis | Hong Kong, China [12]; Taiwan, China [17] |
| 18 | A. splendens | Indian Ocean [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.-Y.; Wang, Y.-P.; Yang, J.; Shih, Y.-J.; Chu, T.-J. Revealing the Coral Species Diversity in Xiamen Bay: Spatial Distribution of Genus Astrogorgia (Cnidaria, Alcyonacea, Plexauridae) and Newly Recorded Species. Water 2022, 14, 2417. https://doi.org/10.3390/w14152417
Liu J-Y, Wang Y-P, Yang J, Shih Y-J, Chu T-J. Revealing the Coral Species Diversity in Xiamen Bay: Spatial Distribution of Genus Astrogorgia (Cnidaria, Alcyonacea, Plexauridae) and Newly Recorded Species. Water. 2022; 14(15):2417. https://doi.org/10.3390/w14152417
Chicago/Turabian StyleLiu, Jia-Ying, Yun-Pei Wang, Jing Yang, Yi-Jia Shih, and Ta-Jen Chu. 2022. "Revealing the Coral Species Diversity in Xiamen Bay: Spatial Distribution of Genus Astrogorgia (Cnidaria, Alcyonacea, Plexauridae) and Newly Recorded Species" Water 14, no. 15: 2417. https://doi.org/10.3390/w14152417