
Population Status and Ecology of the Steno-Endemic Fairy Shrimp Chirocephalus sibyllae Cottarelli and Mura, 1975 Inhabiting a Mountain Temporary Pond (Central Italy)

 ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Biology and Ecology of Chirocephalus sibyllae
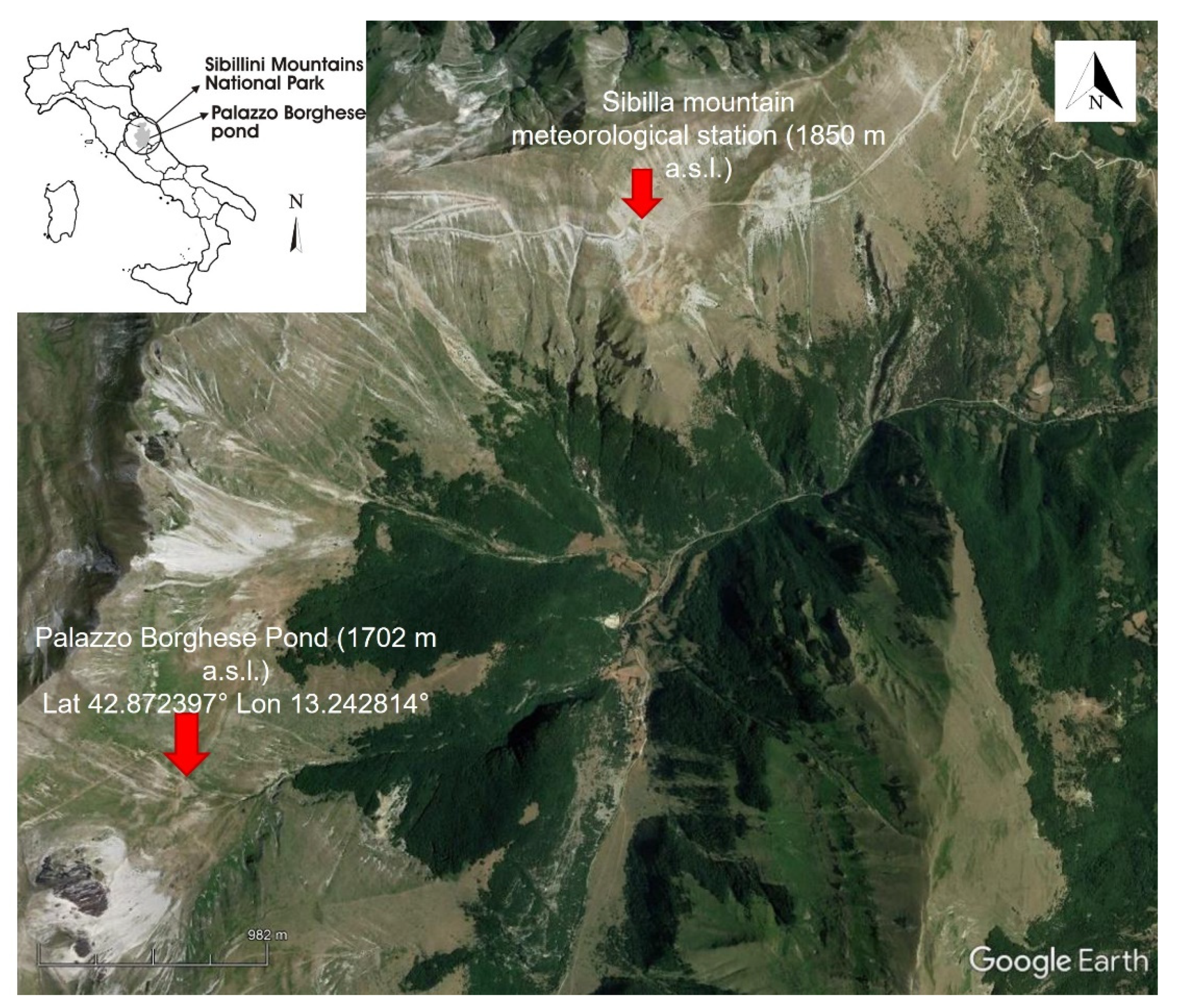
2.2. Study Area
2.3. Data Collection
- Environmental characterization
- 2.
- Zooplankton community composition and C. sibyllae population status
2.4. Laboratory Analysis
2.5. Statistical Analysis
3. Results
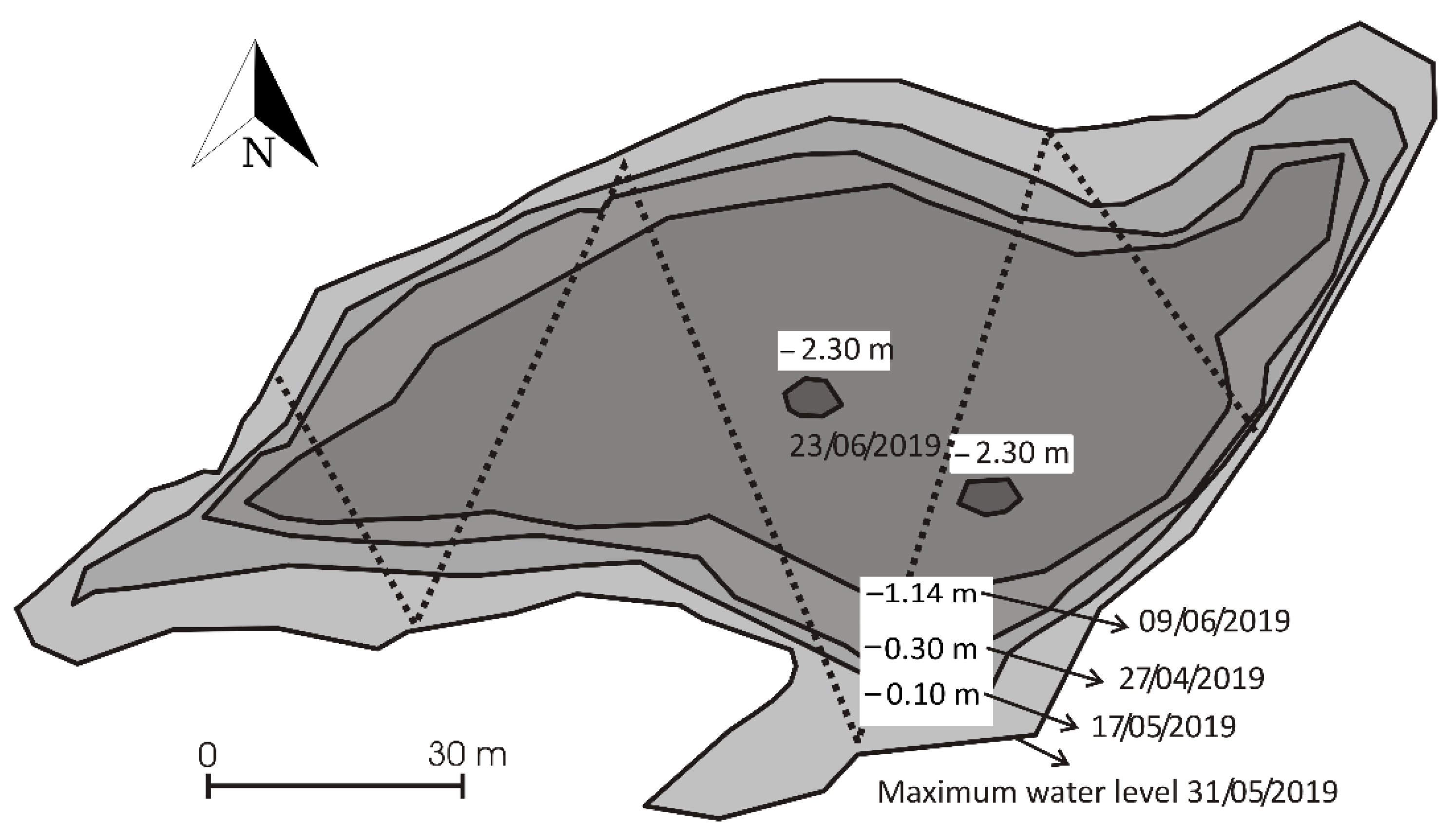
3.1. Environmental Characterization
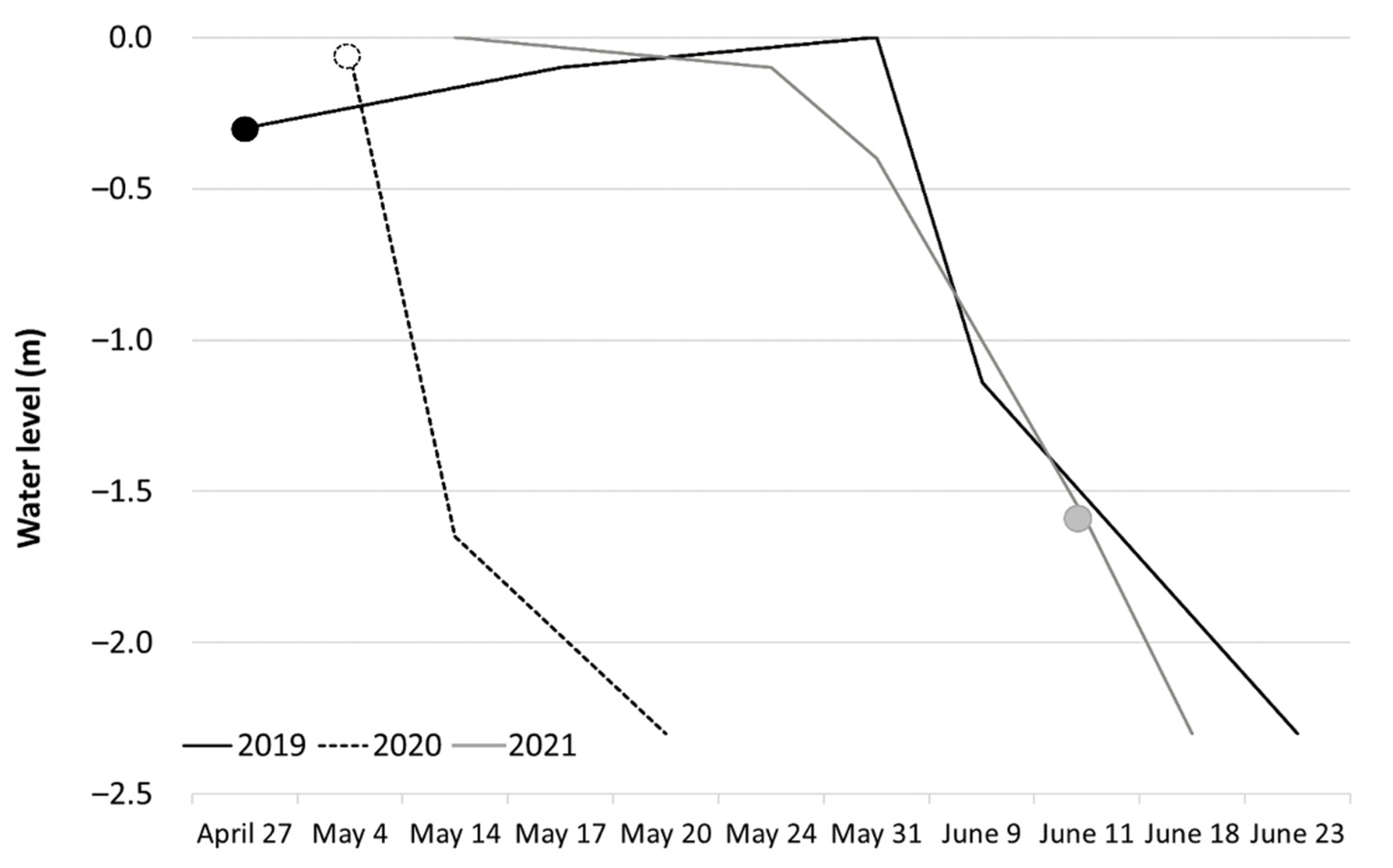
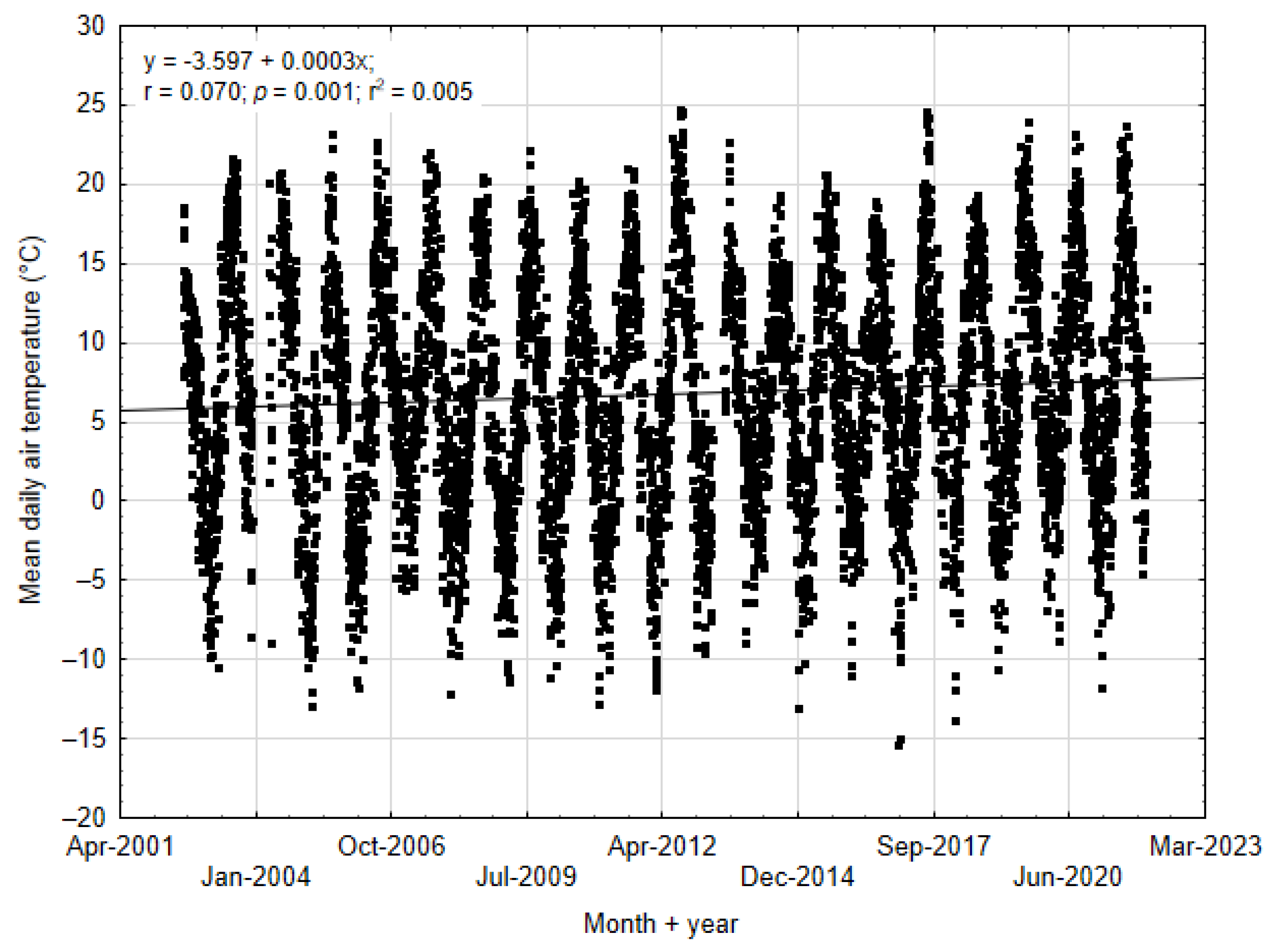
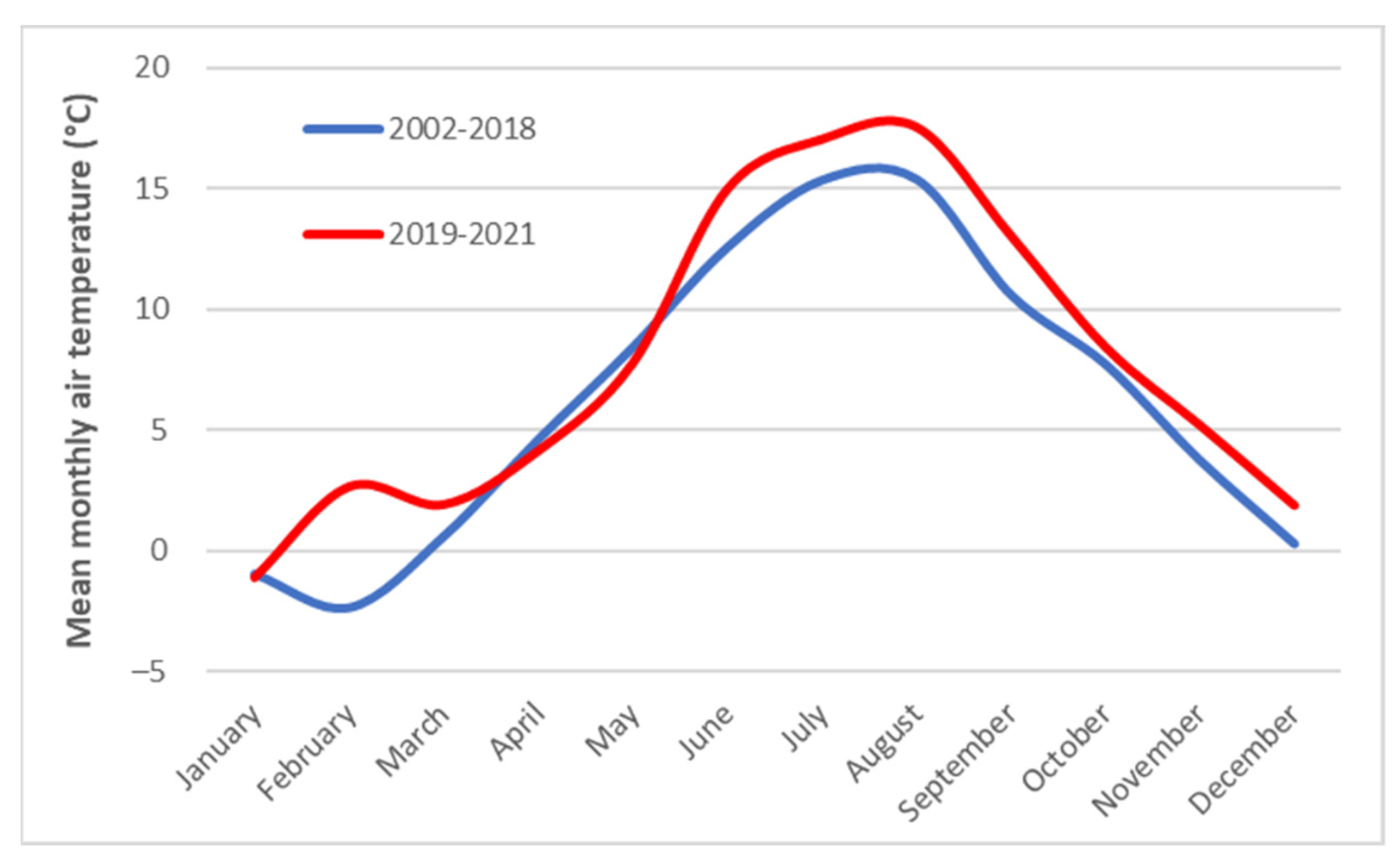
3.2. Climatic Conditions
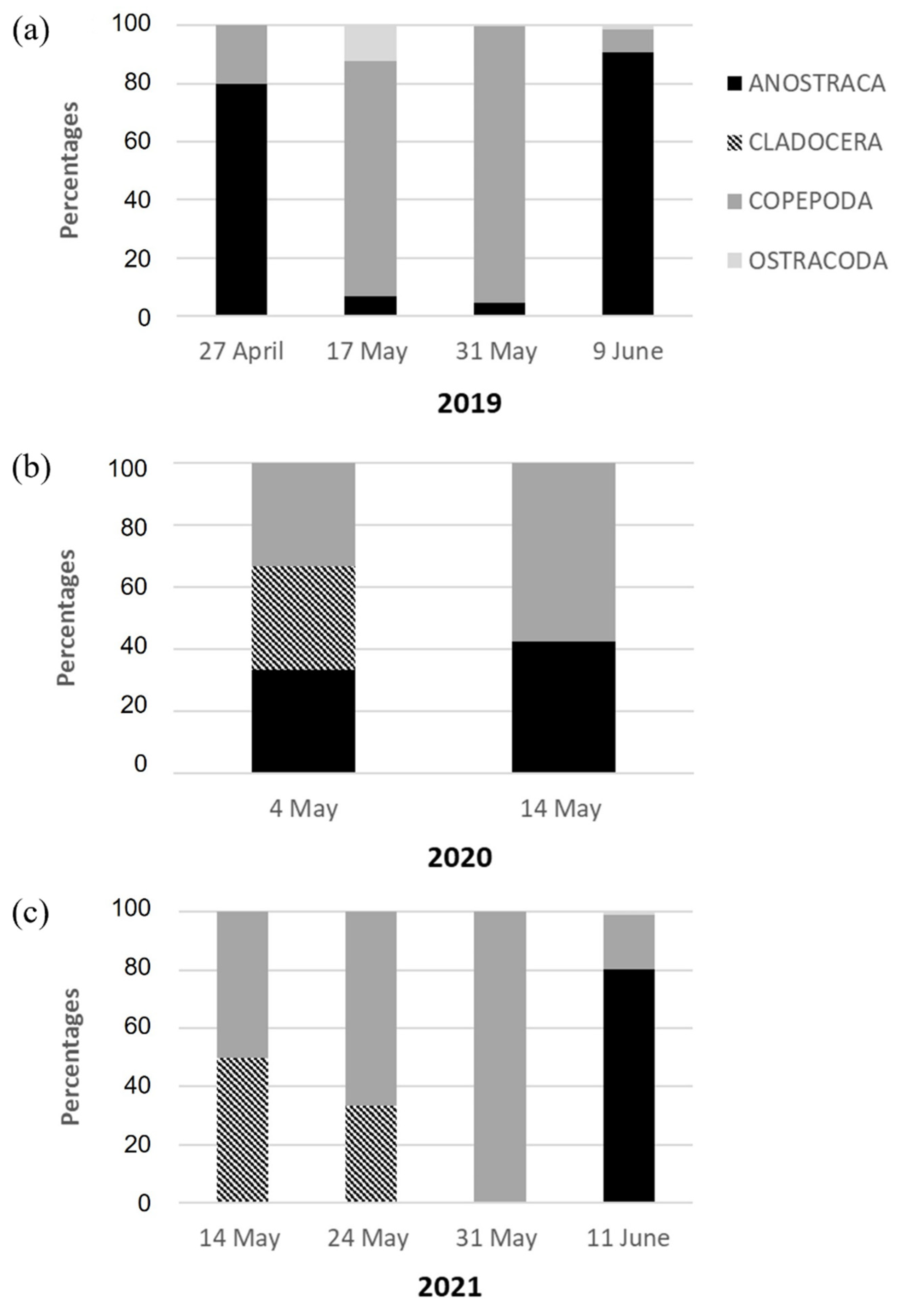
3.3. Composition of the Zooplankton Community
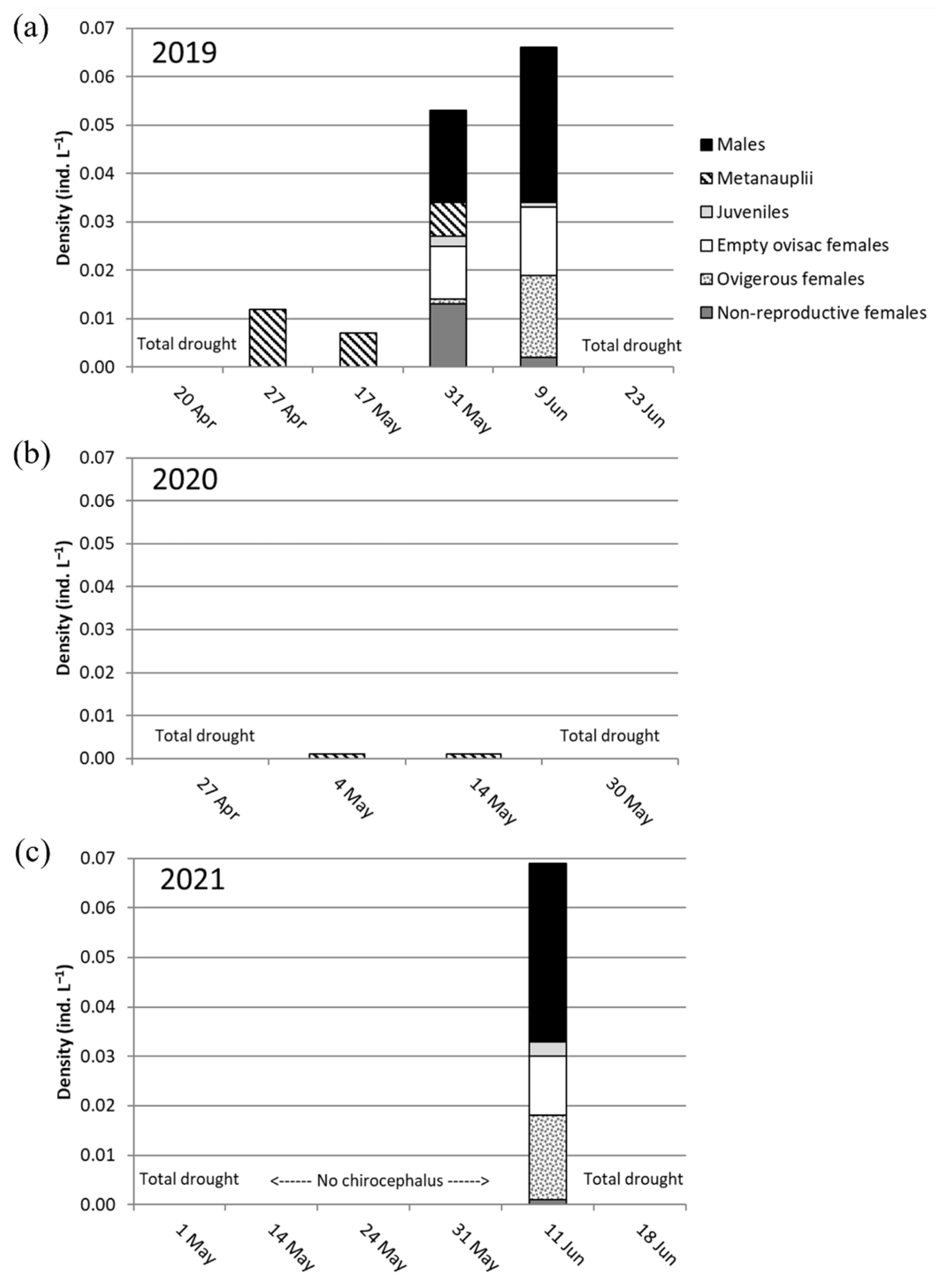
3.4. Chirocephalus sibyllae Population Status, and Life History Traits
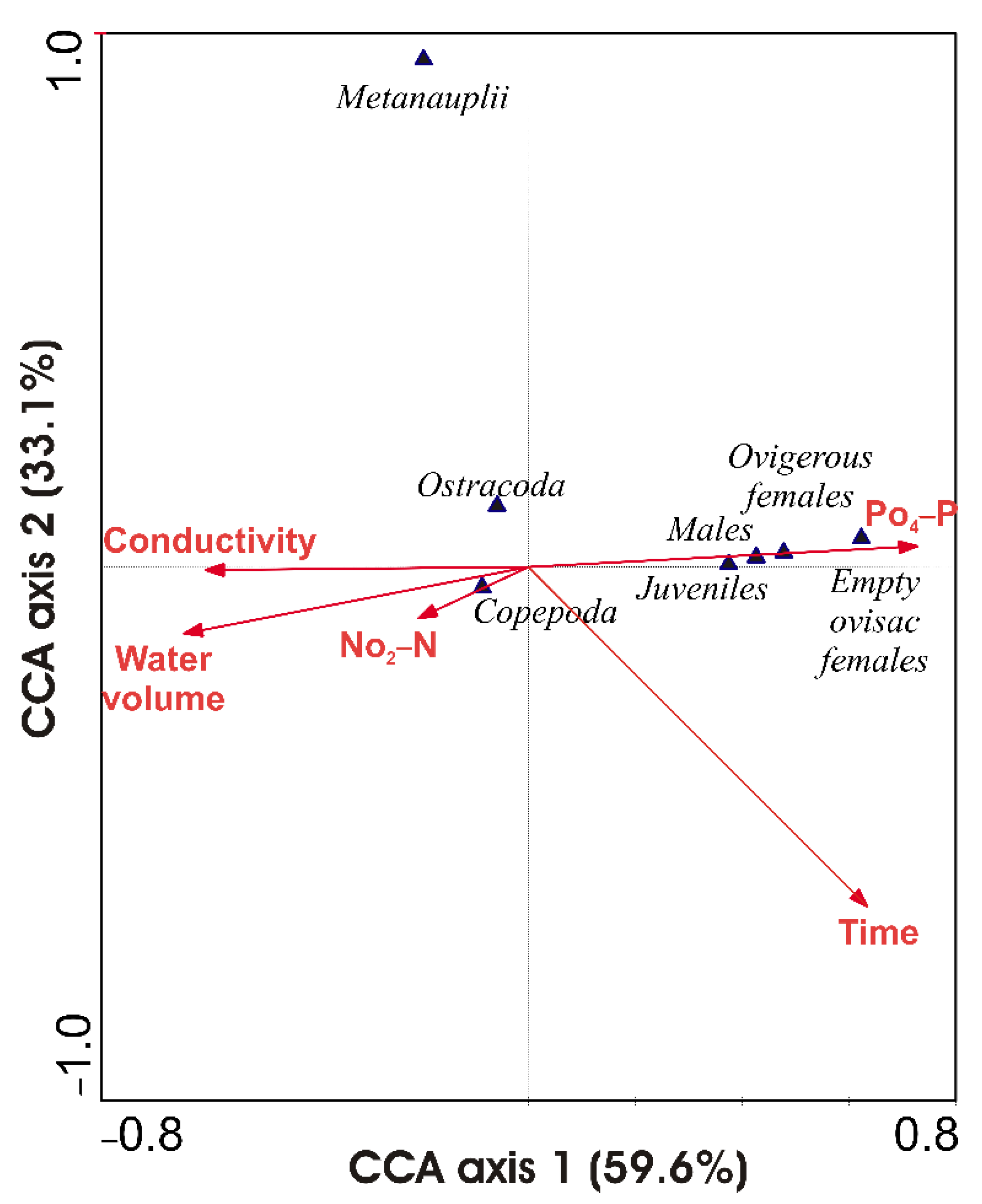
3.5. Biotic Responses to Environmental Drivers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adrian, R.; O’Reilly, C.M.; Zagarese, H.; Baines, S.B.; Hessen, D.O.; Keller, W.; Livingstone, D.M.; Sommaruga, R.; Straile, D.; Van Donk, E.; et al. Lakes as sentinels of climate change. Limnol. Oceanogr. 2009, 54, 2283–2297. [Google Scholar] [CrossRef]
- Carosi, A.; Padula, R.; Ghetti, L.; Lorenzoni, M. Endemic freshwater fish range shifts related to global climate changes: A long-term study provides some observational evidence for the Mediterranean area. Water 2019, 11, 2349. [Google Scholar] [CrossRef]
- Williamson, C.E.; Saros, J.E.; Vincent, W.F.; Smol, J.P. Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnol. Oceanogr. 2009, 54, 2273–2282. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Phil. Trans. R. Soc. B 2010, 365, 2093–2106. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Forconi, A.; Lorenzoni, M. Fish community of the river Tiber basin (Umbria-Italy): Temporal changes and possible threats to native biodiversity. Knowl. Manag. Aquat. Ecosyst. 2015, 416, 22. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; IPCC: Geneva, Switzerland, 2021; in press. [Google Scholar]
- Padula, R.; Carosi, A.; Rossetti, A.; Lorenzoni, M. The Diatomic Diversity of Two Mediterranean High-Elevation Lakes in the Sibillini Mountains National Park (Central Italy). Environments 2021, 8, 79. [Google Scholar] [CrossRef]
- Wissinger, S.A.; Oertli, B.; Rosset, V. Invertebrate Communities of Alpine Ponds. In Invertebrates in Freshwater Wetlands; Batzer, D., Boix, D., Eds.; Springer: Cham, Switzerland, 2016; pp. 55–103. [Google Scholar] [CrossRef]
- Pinceel, T.; Vanschoenwinkel, B.; Hawinkel, W.; Tuytens, K.; Brendonck, L. Aridity promotes bet hedging via delayed hatching: A case study with two temporary pond crustaceans along a latitudinal gradient. Oecologia 2017, 184, 161–170. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Lorenzoni, M.; Barocco, R.; Carosi, A.; Giannetto, D.; Pompei, L. The fish fauna of the Apennine streams related to changes in the regime of wet depositions. Biol. Ambient. 2014, 28, 67–73. [Google Scholar]
- Belk, D. Global status and trends in ephemeral pool invertebrate conservation: Implications for Californian fairy shrimp. In Ecology, Conservation, and Management of Vernal Pool Ecosystems—Proceedings from a 1996 Conference; Witham, C.W., Bauder, E.T., Belk, D., Ferren, W.R., Jr., Ornduff, R., Eds.; California Native Plant Society: Sacramento, CA, USA, 1998; pp. 147–150. [Google Scholar]
- Carosi, A.; Barelli, M.G.; Ambrosi, A.; Rossetti, A.; Padula, R.; Bifulco, C.; Morandi, F.; Lorenzoni, M. Conservation status of Chirocephalus marchesonii Ruffo & Vesentini, 1957 in the Pilato Lake (Sibillini Mountains National Park, Central Italy). Fundam. Appl. Limnol. 2021, 194, 171–185. [Google Scholar]
- Rogers, D.C. A conceptual model for Anostracan biogeography. J. Crustacean Biol. 2015, 35, 686–699. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3. Available online: https://www.iucnredlist.org (accessed on 6 March 2022).
- Cottarelli, V.; Mura, G. Una nuova specie di Anostraco (Crustacea, Branchiopoda) dell’Italia Peninsulare: Chirocephalus Sibyllae N.Sp. Ital. J. Zool. 1975, 42, 187–196. [Google Scholar] [CrossRef]
- Belk, D.; Brtek, J. Checklist of the Anostraca. Hydrobiologia 1995, 298, 315–353. [Google Scholar] [CrossRef]
- Ruffo, S.; Vesentini, G. Una nuova specie di Fillopode Anostraco dei Monti Sibillini. Mem. Mus. Civ. Di Stor. Nat. Di Verona 1957, 6, 1–8. [Google Scholar]
- Ketmaier, V.; Zarattini, P.; De Matthaeis, E.; Cobolli, M.; Mura, G. Intra- and inter-specific relationships in the six Italian species of the fairy shrimp genus Chirocephalus: Combining allozyme and mtDNA data. J. Zool. Syst. Evol. Res. 2003, 41, 276–285. [Google Scholar] [CrossRef]
- Incagnone, G.; Marrone, F.; Barone, R.; Robba, L.; Naselli- Flores, L. How do freshwater organisms cross the “dry ocean”? A review on passive dispersal and colonization processes with a special focus on temporary ponds. Hydrobiologia 2015, 750, 103–123. [Google Scholar] [CrossRef]
- Mura, G.; Calzecchi-Onesti, B. Osservazioni sul ciclo biologico in natura di Chirocephalus sibyllae. Riv. Idrobiol. 1983, 22, 2–3. [Google Scholar]
- Nourisson, M. Influence d’un thermoperiodisme journalier sur le développement des oeufs non assechés de Chirocephalus stagnalis Shaw (Crustacé Phillopode). C.R. Acad. Sci. 1961, 253, 1870–1872. [Google Scholar]
- Hall, R.E. The development of eggs of Chirocephalus diaphanus Prevost at low temperature. Hydrobiologia 1959, 13, 156–159. [Google Scholar] [CrossRef]
- Cottarelli, V.; Mura, G. Guide per il riconoscimento delle specie animali delle acque interne italiane. In Anostraci, Notostraci, Concostraci (Crustacea: Anostraca, Notostraca, Conchostraca); Collana CNR AQ/1/94; Stamperia Valdonega: Verona, Italy, 1983; pp. 36–38. [Google Scholar]
- Belk, D. Evolution of egg size strategies in fairy shrimps. Southwest. Nat. 1977, 22, 99–105. [Google Scholar] [CrossRef]
- Agenzia per la Protezione dell’Ambiente e per i servizi Tecnici, Consiglio Nazionale delle Ricerche, & Istituto di Ricerca sulle Acque (APAT). Analytical Methods for Water. Manuals and Guidelines; IGER srl: Rome, Italy, 2003; p. 1153. Available online: http://www.isprambiente.gov.it/it/pubblicazioni/manuali-e-linee-guida/metodi-analitici-per-le-acque (accessed on 6 March 2022).
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D.; American Public Health Association (APHA); American Water Works Association (AWWA); Water Environment Federation (WEF). Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005; p. 2671. [Google Scholar]
- Ghetti, P.F.; Mc Kenzie, K. Guide per il riconoscimento delle specie animali delle acque interne italiane. In Ostracodi (Crustacea, Ostracoda); Collana CNR AQ/1/108; Stamperia Valdonega: Verona, Italy, 1981. [Google Scholar]
- Stella, E. Guide per il Riconoscimento delle Specie Animali delle Acque Interne Italiane. Calanoidi (Crustacea, Copepoda, Calanoide); Collana CNR AQ/1/140; Stamperia Valdonega: Verona, Italy, 1982. [Google Scholar]
- Abramoff, M.D.; Magalhães, P.J.; Ram, S.J. Processing with Image. J. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Brendonck, L.; Rogers, D.C.; Olesen, J.; Weeks, S.; Hoeh, W.R. Global diversity of large branchiopods (Crustacea: Branchiopoda) in freshwater. Hydrobiologia 2008, 595, 167–176. [Google Scholar] [CrossRef]
- Horváth, Z.; Vad, C.F.; Vörös, L.; Boros, E. Distribution and conservation status of fairy shrimps (Crustacea: Anostraca) in the astatic soda pans of the Carpathian basin: The role of local and spatial factors. J. Limnol. 2013, 72, 103–116. [Google Scholar] [CrossRef]
- Lukić, D.; Waterkeyn, A.; Rabet, N.; Mioduchowska, M.; Geudens, B.; Vanschoenwinkel, B.; Pinceel, T. High genetic variation and phylogeographic relations among Palearctic fairy shrimp populations reflect persistence in multiple southern refugia during Pleistocene ice ages and postglacial colonization. Freshw. Biol. 2019, 64, 1896–1907. [Google Scholar] [CrossRef]
- Cohen, D. Optimizing reproduction in a randomly varying environment. J. Theor. Biol. 1966, 12, 119–129. [Google Scholar] [CrossRef]
- Brendonck, L. Diapause, quiescence, hatching requirements: What we can learn from large freshwater branchiopods (Crustacea: Branchiopoda: Anostraca, Notostraca, Conchostraca). Hydrobiologia 1996, 320, 85–97. [Google Scholar] [CrossRef]
- Maffei, C.; Vagaggini, D.; Zarattini, P.; Mura, G. The dormancy problem for Crustacea Anostraca: A rigorous model connecting hatching strategies and environmental conditions. Ecol. Model. 2005, 185, 469–481. [Google Scholar] [CrossRef]
- Mura, G. Structure and Functioning of the “Egg Bank” of a Fairy Shrimp in a Temporary Pool: Chirocephalus ruffoi from Pollino National Park (Southern Italy) as a Case Study. Internat. Rev. Hydrobiol. 2004, 89, 35–50. [Google Scholar] [CrossRef]
- De Stasio, B.T., Jr. The seed bank of a freshwater crustacean: Copepodology for the plant ecologist. Ecology 1989, 70, 1377–1389. [Google Scholar] [CrossRef]
- Stearns, S.C. Life-history tactics: A review of the ideas. Q. Rev. Biol. 1976, 51, 3–47. [Google Scholar] [CrossRef] [PubMed]
- Philippi, T.; Seger, J. Hedging one’s evolutionary bets, revisited. Trends Ecol. Evol. 1989, 4, 41–44. [Google Scholar] [CrossRef]
- Nix, M.H.; Jenkins, D.G. Life history comparisons of Daphnia obtusa from temporary ponds, cultured with a low-quality food. Aquat. Ecol. 2000, 34, 19–27. [Google Scholar] [CrossRef]
- Mura, G. Preliminary testing of Anostraca from Italy for use in freshwater fish culture. Hydrobiologia 1992, 241, 185–194. [Google Scholar] [CrossRef]
- Søndergaard, M.; Kristensen, P.; Jeppesen, E. Phosphorus release from resuspended sediment in the shallow and wind exposed Lake Arresø, Denmark. Hydrobiologia 1992, 228, 91–99. [Google Scholar] [CrossRef]
- Declerck, S.; De Bie, T.; Ercken, D.; Hampel, H.; Schrijvers, S.; Van Wichelen, J.; Gillard, V.; Mandiki, R.; Losson, B.; Bauwens, D.; et al. Ecological characteristics of small farmland ponds: Associations with land use practices at multiple spatial scales. Biol. Conserv. 2006, 131, 523–532. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | 2020 | 2021 | |||
|---|---|---|---|---|---|
| n = 4 | n = 2 | n = 4 | |||
| Range | Mean ± SD | Range | Range | Mean ± SD | |
| Air temperature (°C) | 5.0–22.5 | 10.4 ± 8.2 | 12.5–16.2 | 9.3–18.5 | 13.0 ± 4.3 |
| Ammonia (mg L−1 NH4+) | 0.06–0.10 | 0.07 ± 0.02 | 0.12–0.17 | 0.03–0.07 | 0.04 ± 0.01 |
| BOD5 (mg L−1) | 0.0–1.8 | 0.8 ± 0.76 | 1.7–1.8 | 0.4–3.0 | 1.6 ± 1.15 |
| Chlorides (mg L−1) | 4.0–23.0 | 14.0 ± 7.8 | 11.0–12.0 | 14.0–34.0 | 26.3 ± 8.7 |
| COD (mg L−1) | 8.0–32.4 | 16.0 ± 11.2 | 8.0–10.7 | 10.7–23.1 | 15.3 ± 5.5 |
| Conductivity (µS cm−1) | 193–211 | 201 ± 7.8 | 180–238 | 105–161 | 143 ± 26.3 |
| Dissolved oxygen (mg L−1) | 7.2–9.8 | 8.3 ± 1.1 | 7.2–8.8 | 6.8–11.4 | 9.2 ± 1.9 |
| Nitrate (mg L−1 NO3-N) | 0.30–1.10 | 0.58 ± 0.38 | 0.3–0.7 | 0.4–0.8 | 0.50 ± 0.20 |
| Nitrite (mg L−1 NO2-N) | 0.05–0.07 | 0.06 ± 0.01 | 0.07–0.09 | 0.06–0.09 | 0.08 ± 0.01 |
| pH (units) | 7.4–8.6 | 8.0 ± 0.53 | 7.4–7.8 | 7.5–8.3 | 8.0 ± 0.38 |
| Orthophosphates (mg L−1 PO4-P) | 0.07–0.22 | 0.12 ± 0.07 | 0.04–0.21 | 0.03–0.12 | 0.06 ± 0.04 |
| Sulphates (mg L−1) | 0.00–0.00 | 0.00 ± 0.00 | 0.00–5.00 | 0.00–0.00 | 0.00 ± 0.00 |
| Water temperature (°C) | 4.9–19.8 | 10.1 ± 6.7 | 8.8–18.0 | 13.6–6.6 | 13.6 ± 6.6 |
| Correlation with axis | ||||
|---|---|---|---|---|
| AX1 | p | AX2 | p | |
| Conductivity (µS cm−1) | −0.341 | 0.335 | 0.070 | 0.847 |
| Nitrites (mg L−1 NO2-N) | −0.365 | 0.299 | 0.271 | 0.449 |
| Orthophosphates (mg L−1 PO4-P) | 0.621 | 0.049 | −0.324 | 0.361 |
| Water volume (m3) | −0.413 | 0.236 | 0.030 | 0.398 |
| Time (days) | 0.777 | 0.008 | −0.790 | 0.007 |
| Non-reproductive females (ind L−1) | 0.074 | 0.839 | −0.281 | 0.431 |
| Ovigerous females (ind L−1) | 0.964 | 0.000 | −0.284 | 0.426 |
| Empty ovisac females (ind L−1) | 0.838 | 0.002 | −0.385 | 0.273 |
| Juveniles (ind L−1) | 0.670 | 0.034 | −0.357 | 0.311 |
| Metanauplii (ind L−1) | −0.356 | 0.313 | 0.368 | 0.295 |
| Males (ind L−1) | 0.904 | 0.000 | −0.365 | 0.300 |
| Copepoda (ind L−1) | −0.081 | 0.824 | −0.269 | 0.452 |
| Ostracoda (ind L−1) | −0.059 | 0.872 | −0.238 | 0.509 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carosi, A.; Barelli, M.G.; Ambrosi, A.; Rossetti, A.; Morandi, F.; Lorenzoni, F.; Tagliaferri, G.; Lorenzoni, M. Population Status and Ecology of the Steno-Endemic Fairy Shrimp Chirocephalus sibyllae Cottarelli and Mura, 1975 Inhabiting a Mountain Temporary Pond (Central Italy). Water 2022, 14, 1750. https://doi.org/10.3390/w14111750
Carosi A, Barelli MG, Ambrosi A, Rossetti A, Morandi F, Lorenzoni F, Tagliaferri G, Lorenzoni M. Population Status and Ecology of the Steno-Endemic Fairy Shrimp Chirocephalus sibyllae Cottarelli and Mura, 1975 Inhabiting a Mountain Temporary Pond (Central Italy). Water. 2022; 14(11):1750. https://doi.org/10.3390/w14111750
Chicago/Turabian StyleCarosi, Antonella, Maria Gaetana Barelli, Alessandro Ambrosi, Alessandro Rossetti, Federico Morandi, Francesca Lorenzoni, Giovanni Tagliaferri, and Massimo Lorenzoni. 2022. "Population Status and Ecology of the Steno-Endemic Fairy Shrimp Chirocephalus sibyllae Cottarelli and Mura, 1975 Inhabiting a Mountain Temporary Pond (Central Italy)" Water 14, no. 11: 1750. https://doi.org/10.3390/w14111750
APA StyleCarosi, A., Barelli, M. G., Ambrosi, A., Rossetti, A., Morandi, F., Lorenzoni, F., Tagliaferri, G., & Lorenzoni, M. (2022). Population Status and Ecology of the Steno-Endemic Fairy Shrimp Chirocephalus sibyllae Cottarelli and Mura, 1975 Inhabiting a Mountain Temporary Pond (Central Italy). Water, 14(11), 1750. https://doi.org/10.3390/w14111750