
Meeting the Salinity Requirements of the Bivalve Mollusc Crassostrea gigas in the Depuration Process and Posterior Shelf-Life Period to Improve Food Safety and Product Quality


 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Area
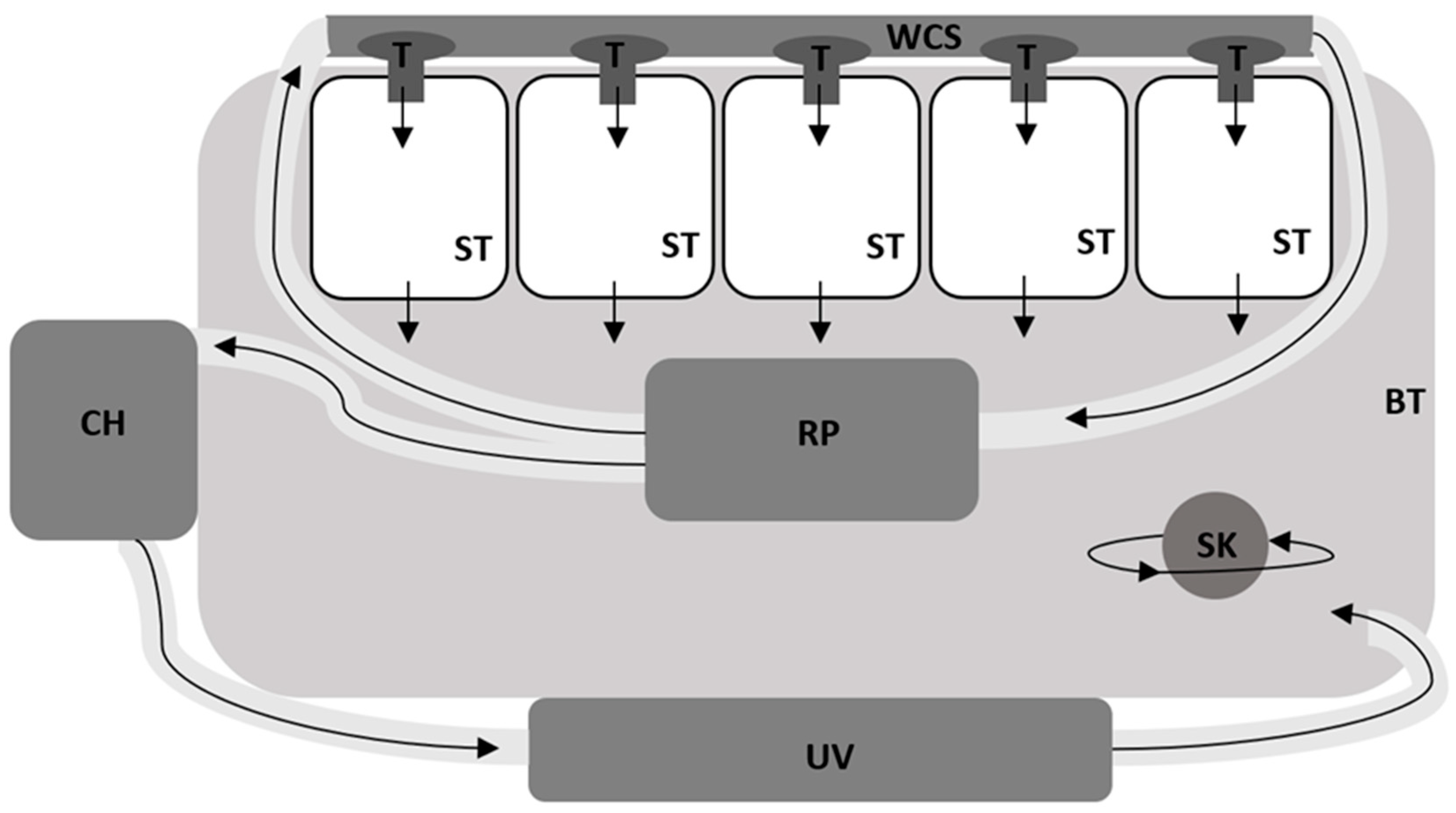
2.2. Experimental System
2.3. Depuration Process
2.4. Microbiological Analysis
2.5. Experimental Procedure
2.6. Statistical Analysis
3. Results
3.1. Microbiological Results
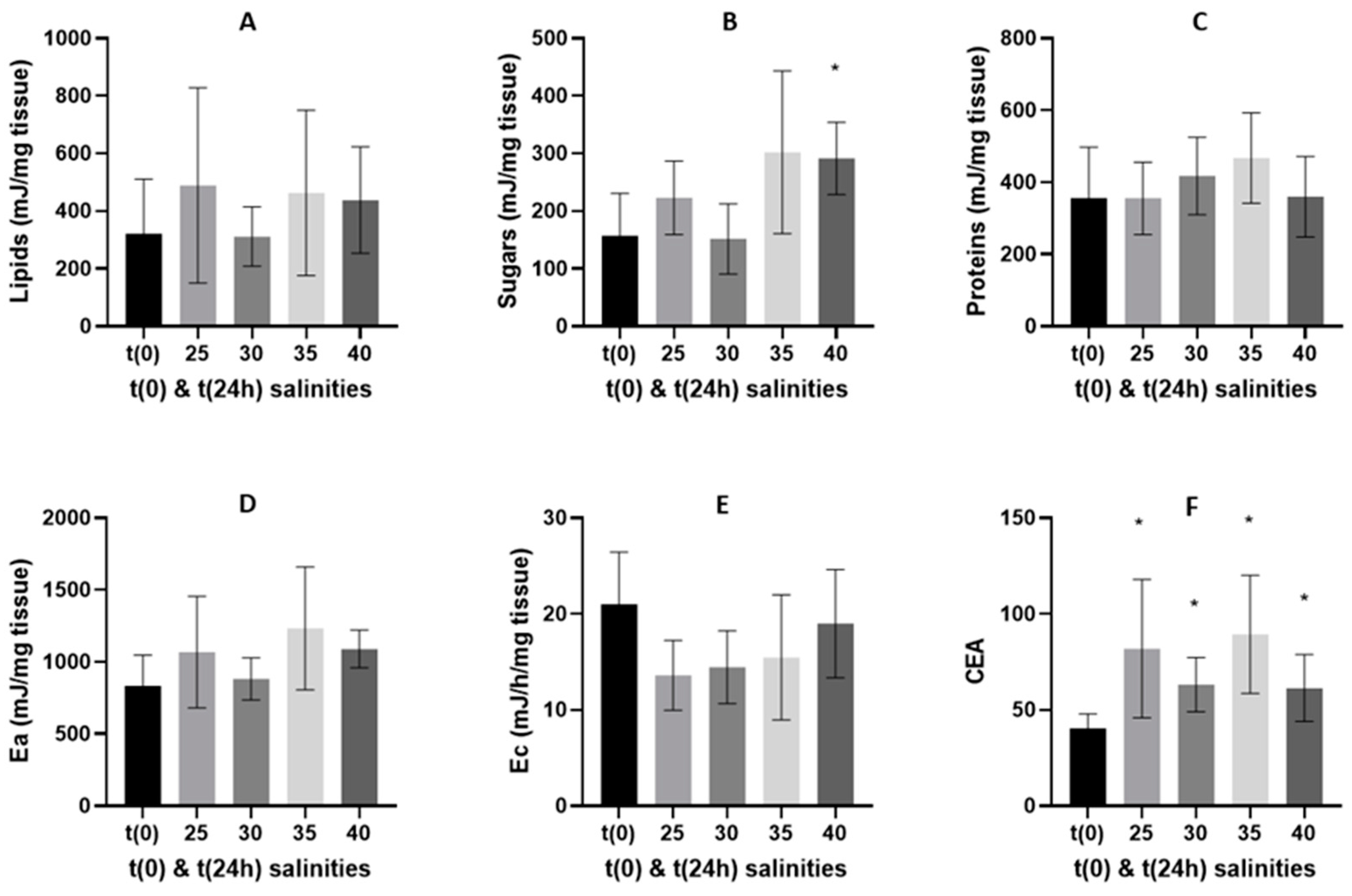
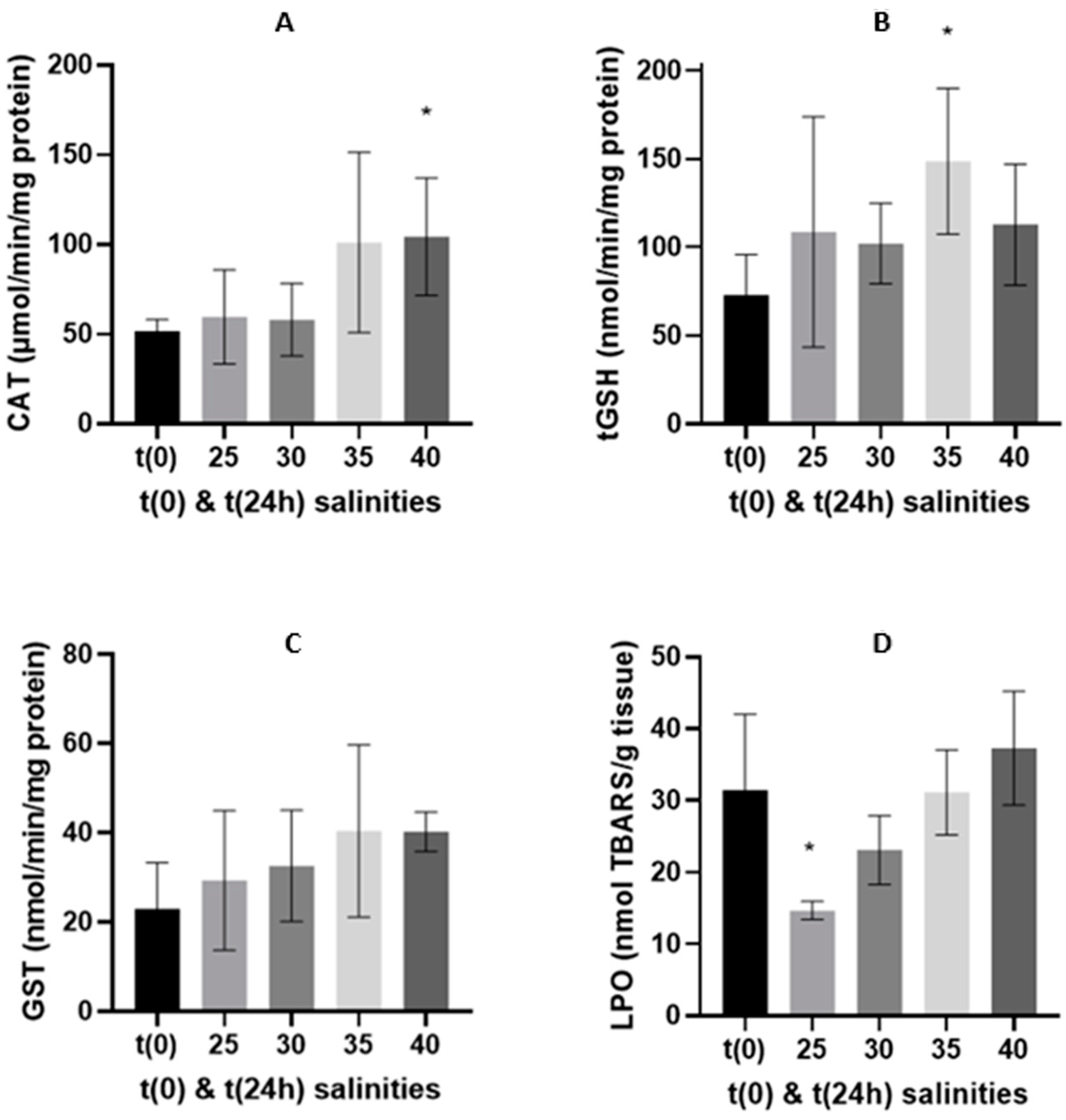
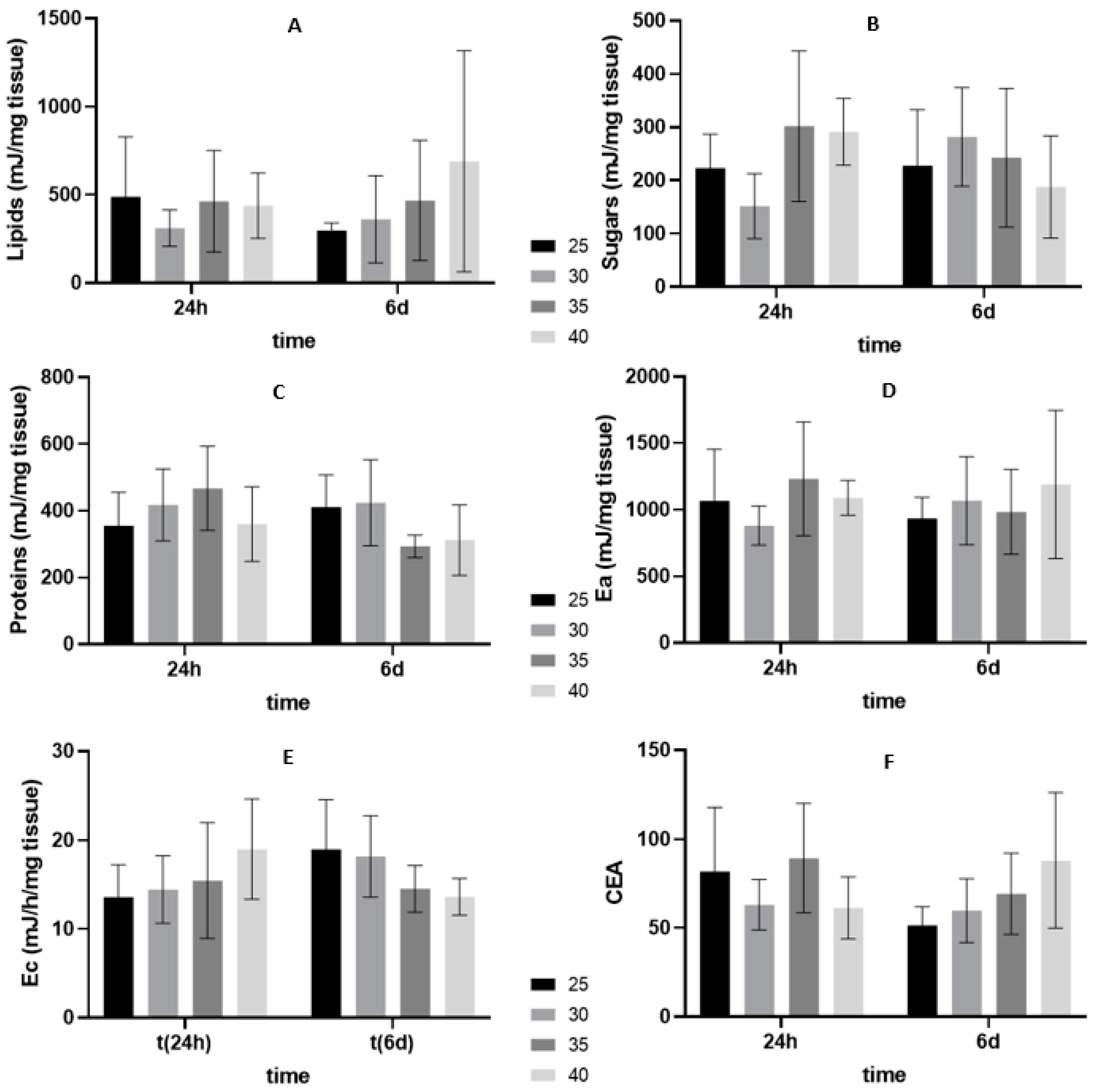
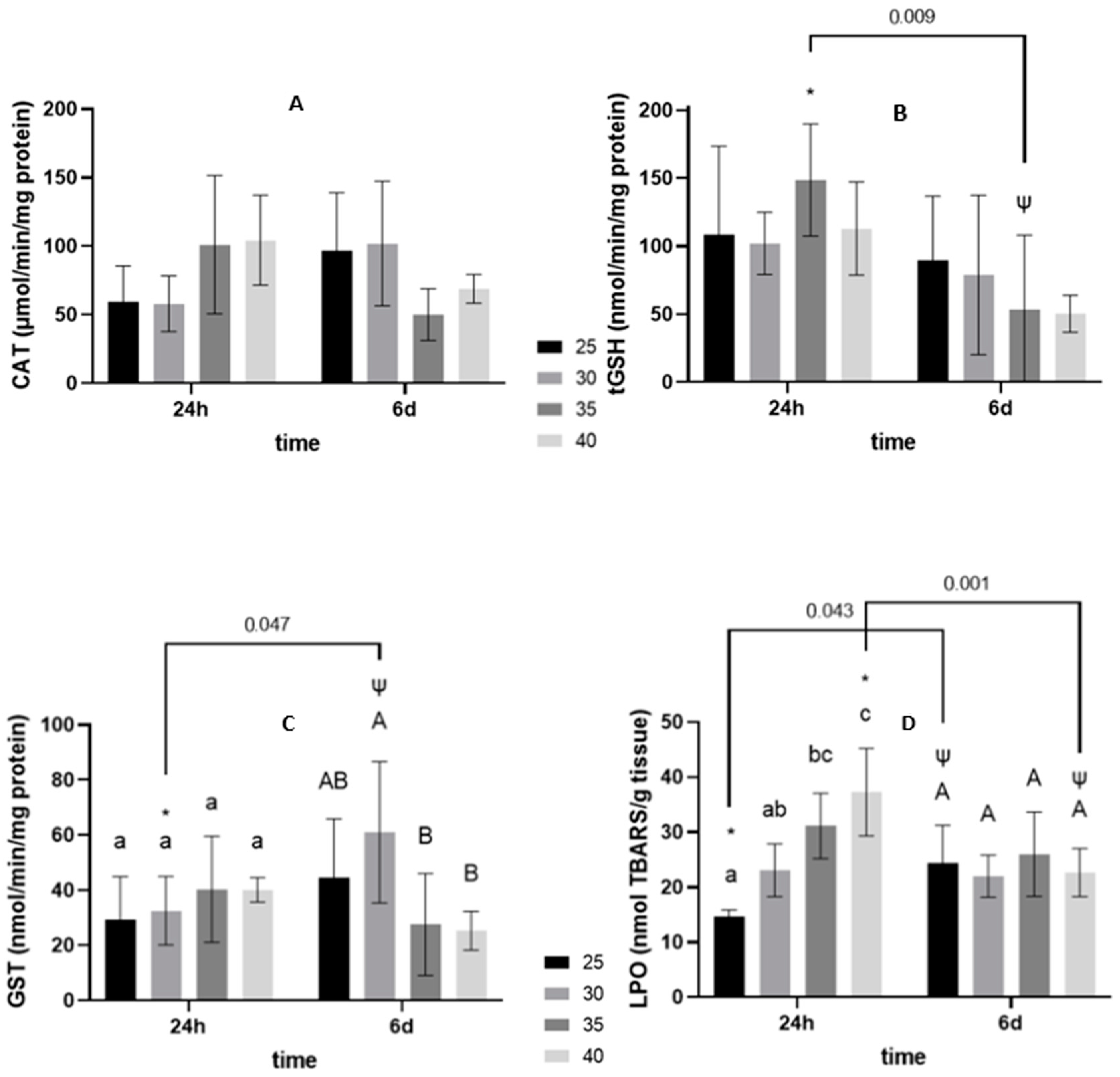
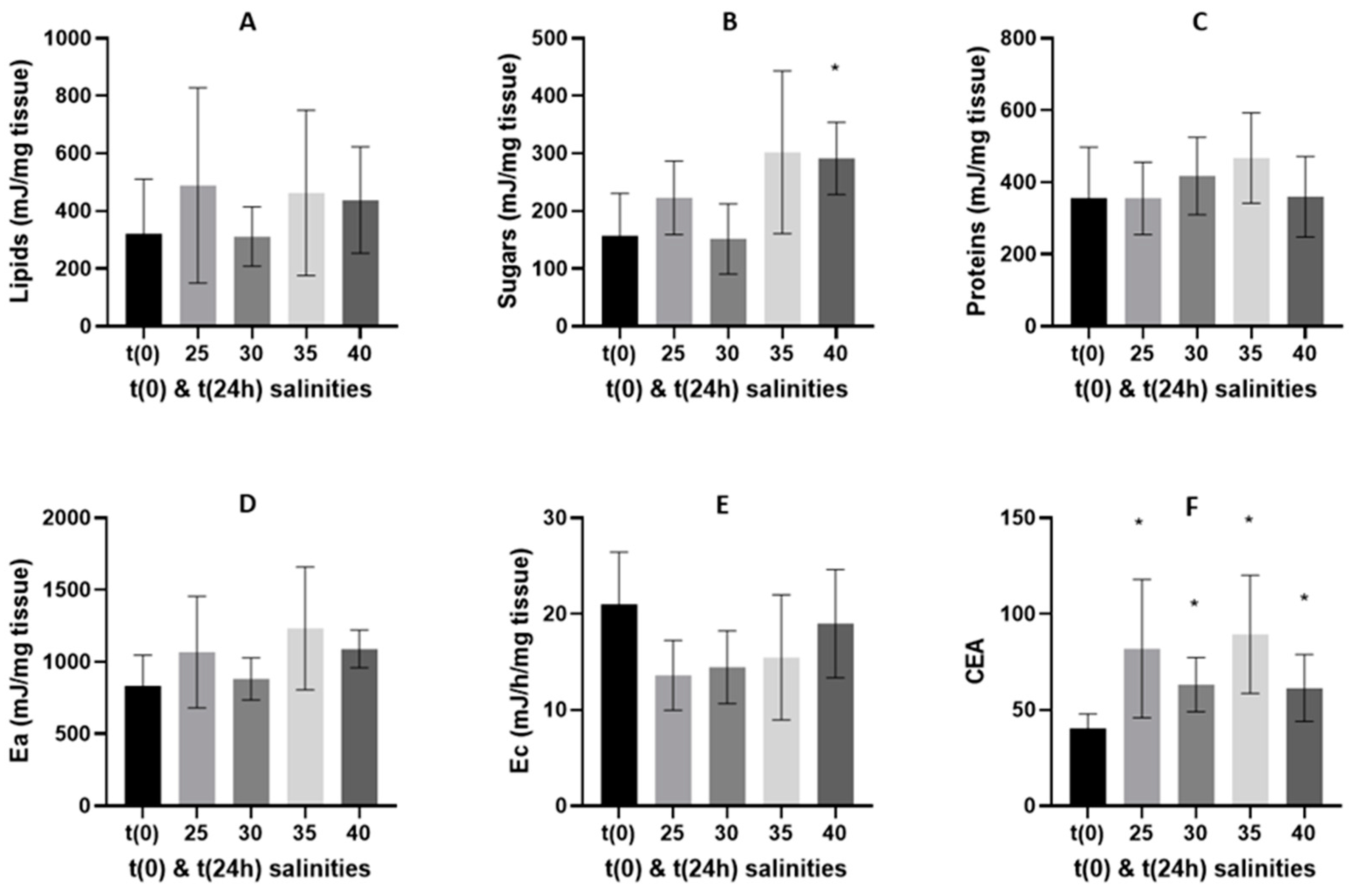
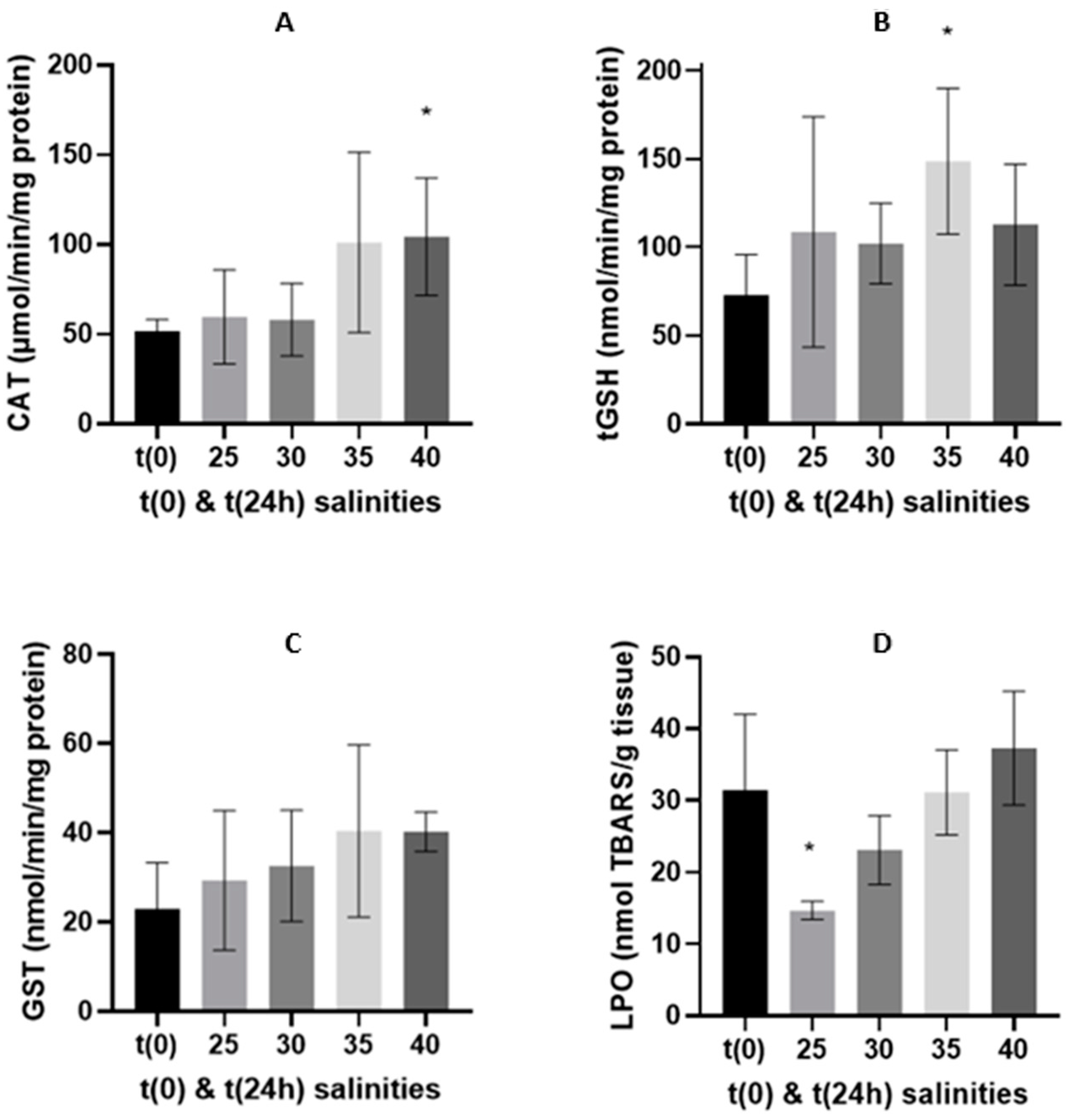
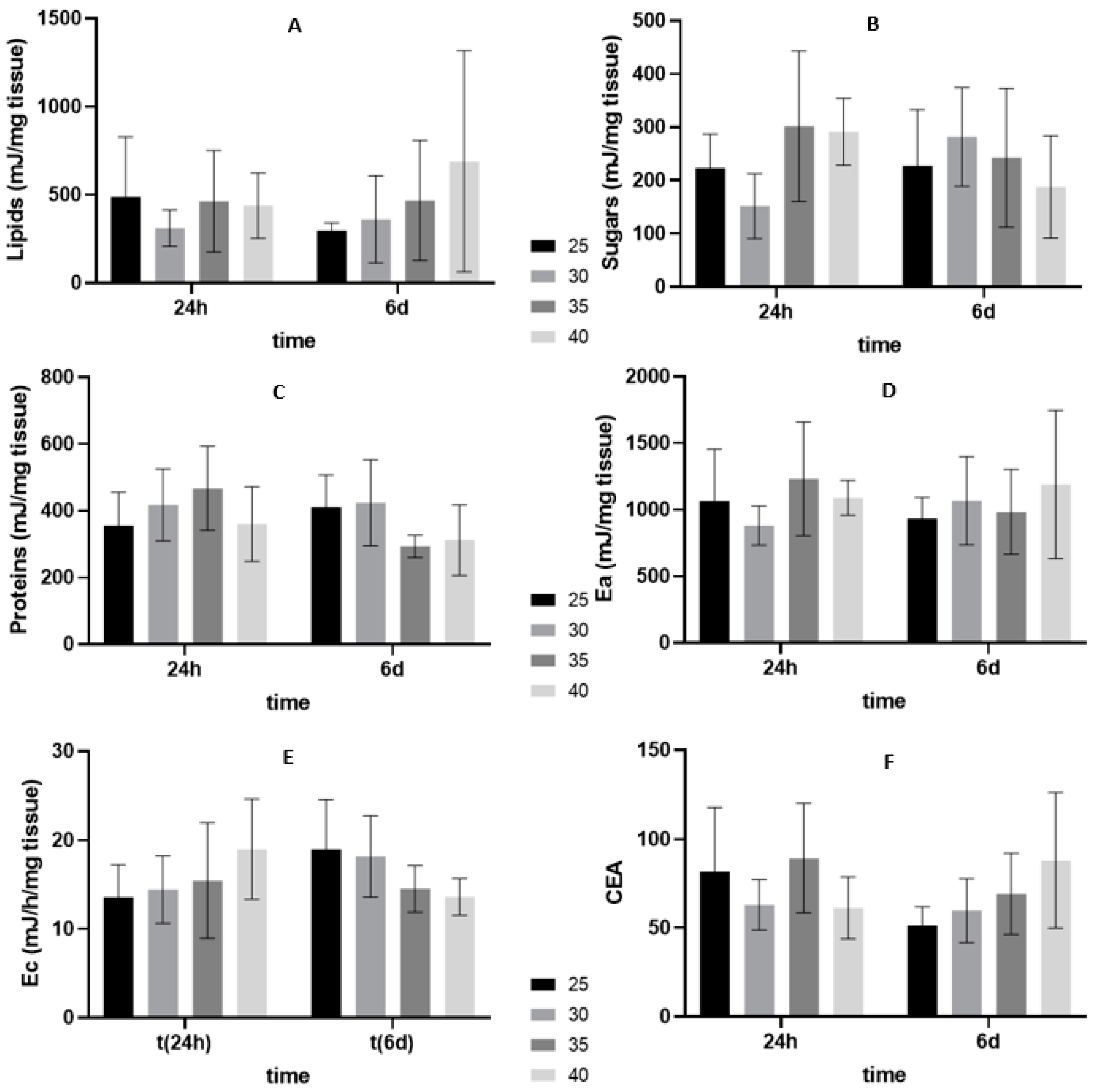
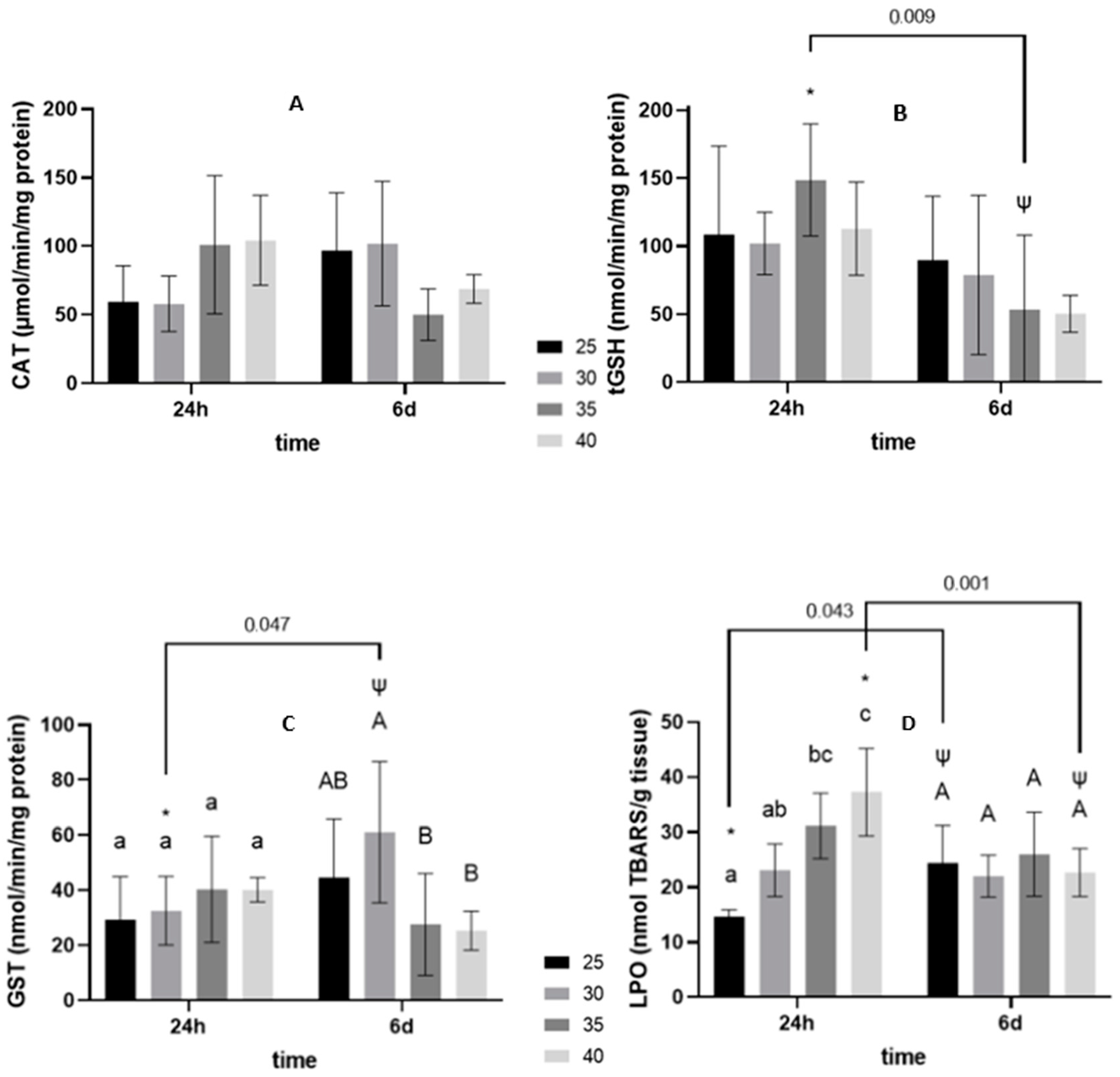
3.2. Oxidative Stress Related Biomarkers
3.2.1. 24-h Depuration Period
3.2.2. Six Days of Shelf-Life
4. Discussion
4.1. Microbiological Analysis
4.2. Energy Budget and Oxidative Stress-Related Biomarkers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. In Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Soares, K.M.P.; Gonçalves, A.A. Seafood quality and safety. Inst. Adolfo Lutz 2012, 71, 1–10. [Google Scholar]
- Pérez, A.C.A.; Avdalov, N.; Neiva, C.R.P.; Neto, M.J.L.; Lopes, R.G.; Tomita, R.Y. Sanitary-Hygienic Procedures for Industry and Seafood Inspectors: Recommendations; Insituto de Pesca: Santos, Brazil, 2007. [Google Scholar]
- Huss, H.H.; Ababouch, L.; Gram, L. Assessment and Management of Seafood Safety and Quality; Food and Agriculture Organization Fisheries Technical Paper; Food and Agriculture Organization: Roma, Italy, 2003; Volume 444, p. 230. [Google Scholar]
- Ifft, J.; Roland-Holst, D.; Zilberman, D. Consumer valuation of safety-labeled free-range chicken: Results of a field experiment in Hanoi. Agric. Econ. 2012, 43, 607–620. [Google Scholar] [CrossRef]
- De Groote, H.; Narrod, C.; Kimenju, S.C.; Bett, C.; Scott, R.P.; Tiongco, M.M.; Gitonga, Z.M. Measuring rural consumers’ willingness to pay for quality labels using experimental auctions: The case of aflatoxin-free maize in Kenya. Agric. Econ. 2016, 47, 33–45. [Google Scholar] [CrossRef]
- Adak, G.K.; Meakins, S.M.; Yip, H.; Lopman, B.A.; O’Brien, S.J. Disease risks from foods, England and Wales, 1996–2000. Emerg. Infect. Dis. 2005, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, I.A.; Adak, G.K.; O’Brien, S.J.; Brett, M.M.; Bolton, F.J. General outbreaks of infectious intestinal disease associated with fish and shellfish, England and Wales, 1992–1999. Commun. Dis. Public Health 2001, 4, 117–123. [Google Scholar] [PubMed]
- Prins, T.C.; Smaal, A.C. The role of the blue mussel Mytilus edulis in the cycling of the nutrients in the Oosterscheidc estuary (The Netherlands). In The Oosterschelde Estuary (The Netherlands): A Case-Study of a Changing Ecosystem; Nienhuis, P.H., Smaal, A.C., Eds.; Springer: Dordrecht, The Netherlands, 1994; Volume 97. [Google Scholar] [CrossRef]
- Capelle, J.J. Production Efficiency of Mussel Bottom Culture. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2017. [Google Scholar]
- Anrooy, R.V.; Secretan, P.A.D.; Lou, Y.; Roberts, R.; Upare, M. Review of the Current State of World Aquaculture Insurance; Fisheries Technical Paper 493; FAO Fisheries Department: Rome, Italy, 2006; p. 107. [Google Scholar]
- Potasman, I.; Paz, A.; Odeh, M. Infectious outbreaks associated with bivalve shellfish consumption: A worldwide perspective. Clin. Infect. Dis. 2002, 35, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Hussenot, J.M.E. Floating Skimmers Improve Water Quality in Tanks, Ponds; Global Aquaculture Advocate: L’Houmeau, France, 2004. [Google Scholar]
- Matthiessen, G.C. Oyster Culture; Fishing News Books: London, UK, 2001; p. 163. [Google Scholar]
- Gosling, E. Bivalve Molluscs: Biology, Ecology and Culture; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Telesh, I.V.; Khlebovich, V.V. Principal processes within the estuarine salinity gradient: A review. Mar. Pollut. Bull. 2010, 61, 149–155. [Google Scholar] [CrossRef]
- Pauley, G.P.; Van der Raay, B.; Troutt, D. Species Profile, Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates (Pacific Northwest); US Fish and Wildlife Service: Corvallis, OR, USA, 1988.
- Carregosa, V.; Velez, C.; Soares, A.M.V.M.; Figueira, E.; Freitas, R. Physiological and biochemical responses of three Veneridae clams exposed to salinity changes. Comp. Biochem. Physiol. Part B Biochem. Mol. Bio. 2014, 177–178, 1–9. [Google Scholar] [CrossRef]
- Dickinson, G.H.; Ivanina, A.V.; Matoo, O.B.; Pörtner, H.O.; Lannig, G.; Bock, C.; Beniash, E.; Sokolova, I.M. Interactive effects of salinity and elevated CO2 levels on juvenile eastern oysters, Crassostrea virginica. J. Exp. Biol. 2012, 215, 29–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilles, R. Osmoregulation in Three Molluscs: Acanthochitona discrepans (Brown), Glycymeris glycymeris (L.) and Mytilus edulis (L.). Biol. Bull. 1972, 142, 25–35. [Google Scholar] [CrossRef]
- Neufeld, D.S.; Wright, S.H. Response of cell volume in Mytilus gill to acute salinity change. J. Exp. Biol 1996, 199, 473–484. [Google Scholar]
- Shumway, S.E. Effect of fluctuating salinity on the tissue water content of eight species of bivalve molluscs. J. Comp. Physiol 1977, 116, 269–285. [Google Scholar] [CrossRef]
- Shumway, S.E. Effect of salinity fluctuation on the osmotic pressure and Na+, Ca2+ and Mg2+ ion concentrations in the hemolymph of bivalve molluscs. Mar. Biol. 1977, 41, 153–177. [Google Scholar] [CrossRef]
- Shumway, S.E.; Gabbott, P.A.; Youngson, A. Effect of fluctuating salinity on concentrations of free amino-acids ninhydrin substances or muscles of 8 species of bivalve mollusks. J. Exp. Mar. Biol. Ecol. 1977, 29, 131–150. [Google Scholar] [CrossRef]
- Rivera-Ingraham, G.A.; Lignot, J.-H. Osmoregulation, bioenergetics and oxidative stress in coastal marine invertebrates: Raising the questions for future research. J. Exp. Biol. 2017, 220, 1749–1760. [Google Scholar] [CrossRef] [Green Version]
- Rada, J.P.A.; Duarte, A.C.; Pato, P.; Cachada, A.; Carreira, R.S. Sewage contamination of sediments from two Portuguese Atlantic coastal systems, revealed by fecal sterols. Mar. Pollut. Bull. 2016, 103, 319–324. [Google Scholar] [CrossRef]
- Donovan, T.J.; Gallacher, S.; Anrews, N.J.; Greenwood, M.H.; Graham, J.; Russell, J.E.; Roberts, D.; Lee, R. Modification of the standard method used in the United Kingdom for counting Escherichia coli in live bivalve molluscs. Commun. Dis. Public Health 1998, 1, 188–196. [Google Scholar] [PubMed]
- Rodrigues, A.C.M.; Gravato, C.; Quintaneiro, C.; Golovko, O.; Zlabek, V.; Barata, C.; Soares, A.M.V.M.; Pestana, J.L.T. Life history and biochemical effects of chlorantraniliprole on Chironomus riparius. Sci. Total Environ. 2015, 508, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gnaiger, E. Calculation of Energetic and Biochemical Equivalents of Respiratory Oxygen Consumption. In Polarographic Oxygen Sensors; Gnaiger, E., Forstner, H., Eds.; Aquatic and Physiological Applications; Springer: Berlin/Heidelberg, Germany, 1983; pp. 337–345. [Google Scholar] [CrossRef]
- De Coen, W.; Janssen, C.R. The use of biomarkers in Daphnia magna toxicity testing. IV.Cellular Energy Allocation: A new methodology to assess the energy budget of toxicant-stressed Daphnia populations. J. Aquat. Ecosyst. Stress Recovery 1997, 6, 43–55. [Google Scholar] [CrossRef]
- Verslycke, T.; Ghekiere, A.; Janssen, C.R. Seasonal and spatial patterns in cellular energy allocation in the estuarine mysid Neomysis integer (Crustacea: Mysidacea) of the Scheldt estuary (The Netherlands). J. Exp. Mar. Biol. Ecol. 2004, 306, 245–267. [Google Scholar] [CrossRef]
- Clairborne, A. Catalase activity. In CRC Handbook of Methods in Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Habig, W.H.H.; Pabst, M.J.J.; Jacoby, W.B.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Baker, M.; Cerniglia, G.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal. Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidised glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Rodrigues, A.C.M.; Gravato, C.; Quintaneiro, C.; Bordalo, M.D.; Golovko, O.; Žlábek, V.; Barata, C.; Soares, A.M.V.M.; Pestana, J.L.T. Exposure to chlorantraniliprole affects the energy metabolism of the caddisfly Sericostoma vittatum. Environ. Toxicol. Chem. 2017, 36, 1584–1591. [Google Scholar] [CrossRef]
- Bird, R.P.; Draper, H.H. Comparative studies on different methods of malonaldehyde determination. Methods Enzymol. 1984, 105, 299–305. [Google Scholar] [PubMed]
- Lees, D.; Younger, A.; Dore, B. Depuration and relaying. In Safe Management of Shellfish and Harvest Waters; IWA Publishing: Lowestoft, UK, 2010; pp. 145–181. [Google Scholar]
- Cusson, M.; Tremblay, R.; Daigle, G.; Roussy, M. Modeling the depuration potential of blue mussels (Mytilus spp.) in response to thermal shock. Aquaculture 2005, 250, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.; Cunha, A.; Castilho, F.; Romalde, J.L.; Pereira, M.J. Microbial contaminants and purification of bivalve shellfish: Crucial aspects in monitoring and future perspectives: A mini-review. Food. Contr. 2011, 22, 805–816. [Google Scholar] [CrossRef]
- Love, D.C.; Lovelace, G.L.; Sobsey, M.D. Removal of Escherichia coli, Enterococcus fecalis, coliphages MS2, poliovirus and hepatitis A virus from oysters (Crassostrea virginica) and hardshell clam (Mercinaria mercinaria) by depuration. Int. J. Food Microbiol 2010, 143, 211–217. [Google Scholar] [CrossRef]
- Ostroumov, S.A. Some aspects of water filtering activity of filter-feeders. Hydrobiologia 2005, 542, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Barile, N.B.; Scopa, M.; Nerone, E.; Mascilongo, G. Study of the efficacy of a closed cycle depuration system on bivalve molluscs. Vet. Ital. 2009, 45, 555–566. [Google Scholar]
- Martini, F.; Ober, W.C. Fundamentals of Anatomy and Physiology, 5th ed.; Prentice-Hall: Hoboken, NJ, USA, 2001. [Google Scholar]
- Freitas, R.; Salamanca, L.; Velez, C.; Wrona, F.J.; Soares, A.M.V.M.; Figueira, E. Multiple stressors in estuarine waters: Effects of arsenic and salinity on Ruditapes philippinarum. Sci. Total Environ. 2016, 541, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, A.J.S.; Hilbish, T.J. The costs of cell volume regulation: Protein metabolism during hyperosmotic adjustment. J. Mar. Biol. Assoc. 1992, 72, 569–578. [Google Scholar] [CrossRef]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with water stress: Evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M. Energy-Limited Tolerance to Stress as a Conceptual Framework to Integrate the Effects of Multiple Stressors. Integr. Comp. Biol. 2013, 53, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Huey, R.B.; Berrigan, D. Temperature, demography, and ectotherm fitness. Am. Nat. 2001, 158, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef]
- Velez, C.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Native and introduced clams biochemical responses to salinity and pH changes. Sci. Total Environ. 2016, 566–567, 260–268. [Google Scholar] [CrossRef]
- Moreira, A.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Salinity influences the biochemical response of Crassostrea angulata to Arsenic. Environ. Pollut. 2016, 214, 756–766. [Google Scholar] [CrossRef]
- Gosling, E. Bivalve Mollusc; Wiley-Blackwell: Oxford, UK, 2003. [Google Scholar]
- Kim, W.S.; Huh, H.T.; Huh, S.-H.; Lee, T.W. Effects of salinity on endogenous rhythm of the Manila clam, Ruditapes philippinarum (Bivalvia: Veneridae). Mar. Biol. 2001, 138, 157–162. [Google Scholar] [CrossRef]
- Erk, M.; Muyssen, B.T.A.; Ghekiere, A.; Janssen, C.R. Metallothionein and cellular energy allocation in the estuarine mysid shrimp Neomysis integer exposed to cadmium at different salinities. J. Exp. Mar. Biol. Ecol. 2008, 357, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Amiard-Triquet, C.; Amiard, J.C.; Rainbow, P.S. Ecological Biomarkers: Indicators of Ecotoxicological Effects; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Batley, G.; Simpson, S. Sediment Quality Assessment: A Practical Guide; Csiro Publishing: Clayton, Australia, 2016. [Google Scholar]
- Dent, L.; Lutterschmidt, W.I. Comparative thermal physiology of two sympatric sunfishes (Centrarchidae: Perciformes) with a discussion of microhabitat utilisation. J. Ther. Biol. 2003, 28, 67–74. [Google Scholar] [CrossRef]
- Lam, K.; Tsui, T.; Nakano, K.; Randall, D.J. Physiological Adaptations of Fishes to Tropical Intertidal Environments. Fish Physiol. 2005, 21, 501–581. [Google Scholar]
- Newton, J.R.; Smith-Keune, C.; Jerry, D.R. Thermal tolerance varies in tropical and sub-tropical populations of barramundi (Lates calcarifer) consistent with local adaptation. Aquaculture 2010, 308, S128–S132. [Google Scholar] [CrossRef]
- Khessiba, A.; Roméo, M.; Aïssa, P. Effects of some environmental parameters on catalase activity measured in the mussel (Mytilus galloprovincialis) exposed to lindane. Environ. Pollut. 2005, 133, 275–281. [Google Scholar] [CrossRef]
- Cunha, I.; García, L.M.; Guilhermino, L. Sea-urchin (Paracentrotus lividus) glutathione S-transferases and cholinesterase activities as biomarkers of environmental contamination. J. Environ. Monit. 2005, 7, 288–294. [Google Scholar] [CrossRef]
- Hartl, M.G.J.; Kilemade, M.; Sheehan, D.; Mothersill, C.; O’Halloran, J.; O’Brien, N.M.; van Pelt, F.N.A.M. Hepatic biomarkers of sediment-associated pollution in juvenile turbot, Scophthalmus maximus L. Mar. Environ. Res. 2007, 64, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Hiraku, Y.; Yamashita, N.; Nishiguchi, M.; Kawanishi, S. Catechol estrogens induce oxidative DNA damage and estradiol enhances cell proliferation. Int. J. Cancer 2001, 92, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Zanette, J.; de Almeida, E.A.; da Silva, A.Z.; Guzenski, J.; Ferreira, J.F.; Di Mascio, P.; Marques, M.R.F.; Bainy, A.C.D. Salinity influences glutathione S-transferase activity and lipid peroxidation responses in the Crassostrea gigas oyster exposed to diesel oil. Sci. Total Environ. 2011, 409, 1976–1983. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Times | T 0 h | T 24 h | ||||||||
| Experimental salinities | - | - | 25 | 25 | 30 | 30 | 35 | 35 | 40 | 40 |
| E. coli (MPN/100 g) | 490 | 230 | 20 | <18 | 20 | 20 | <18 | 20 | <18 | <18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvestre, J.A.; Pires, S.F.S.; Pereira, V.; Colaço, M.; Costa, A.P.L.; Soares, A.M.V.M.; Matias, D.; Bettencourt, F.; Fernández-Boo, S.; Rocha, R.J.M.; et al. Meeting the Salinity Requirements of the Bivalve Mollusc Crassostrea gigas in the Depuration Process and Posterior Shelf-Life Period to Improve Food Safety and Product Quality. Water 2021, 13, 1126. https://doi.org/10.3390/w13081126
Silvestre JA, Pires SFS, Pereira V, Colaço M, Costa APL, Soares AMVM, Matias D, Bettencourt F, Fernández-Boo S, Rocha RJM, et al. Meeting the Salinity Requirements of the Bivalve Mollusc Crassostrea gigas in the Depuration Process and Posterior Shelf-Life Period to Improve Food Safety and Product Quality. Water. 2021; 13(8):1126. https://doi.org/10.3390/w13081126
Chicago/Turabian StyleSilvestre, João A., Sílvia F. S. Pires, Vitória Pereira, Miguel Colaço, Ana P. L. Costa, Amadeu M. V. M. Soares, Domitília Matias, Filipa Bettencourt, Sergio Fernández-Boo, Rui J. M. Rocha, and et al. 2021. "Meeting the Salinity Requirements of the Bivalve Mollusc Crassostrea gigas in the Depuration Process and Posterior Shelf-Life Period to Improve Food Safety and Product Quality" Water 13, no. 8: 1126. https://doi.org/10.3390/w13081126