
Late Quaternary Landscape Dynamics at the La Spezia Gulf (NW Italy): A Multi-Proxy Approach Reveals Environmental Variability within a Rocky Embayment

 , ,
, ,
Abstract
1. Introduction
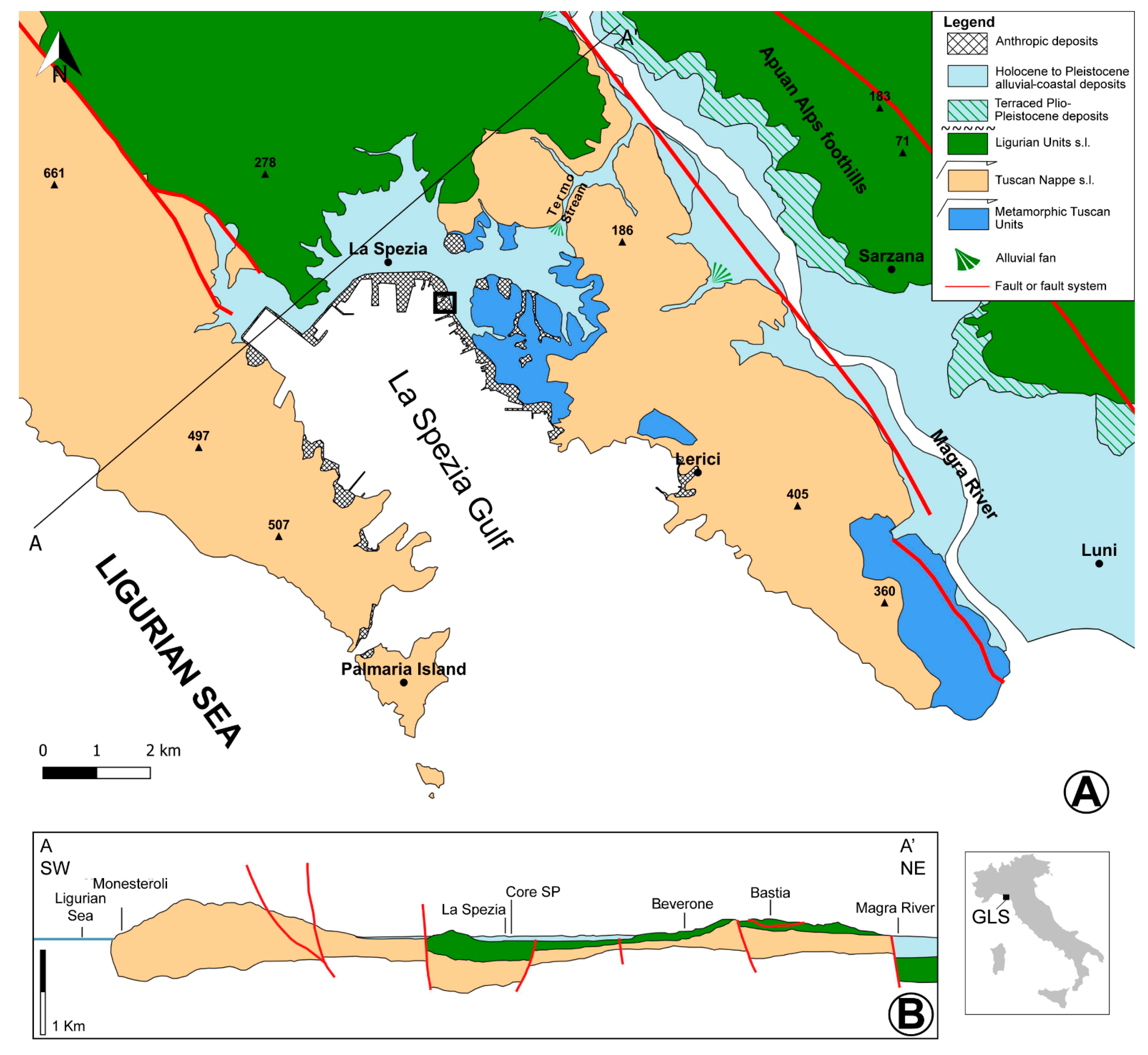
2. Study Area
2.1. Geology and Geomorphology
2.2. Climate and Vegetation
3. Materials and Methods
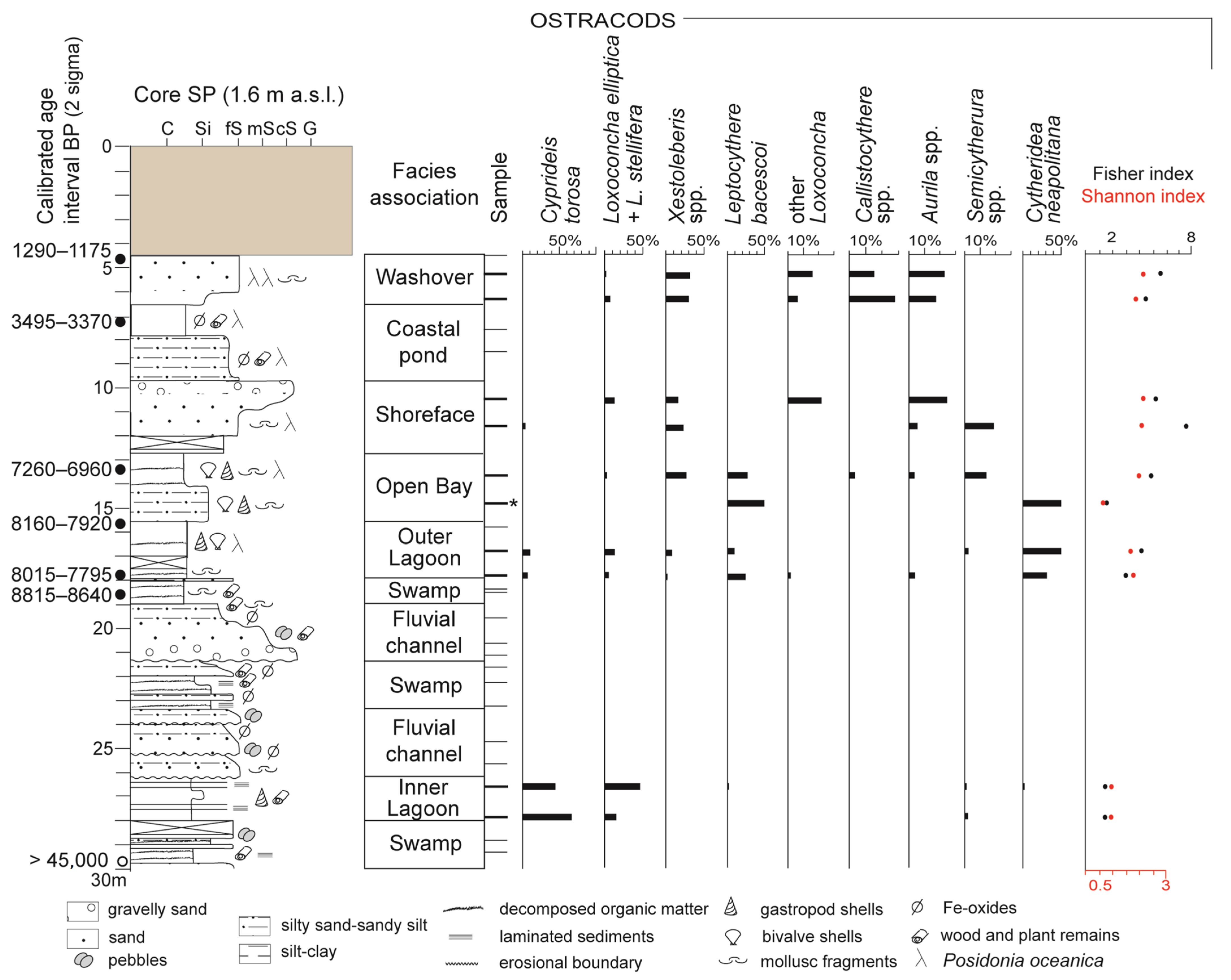
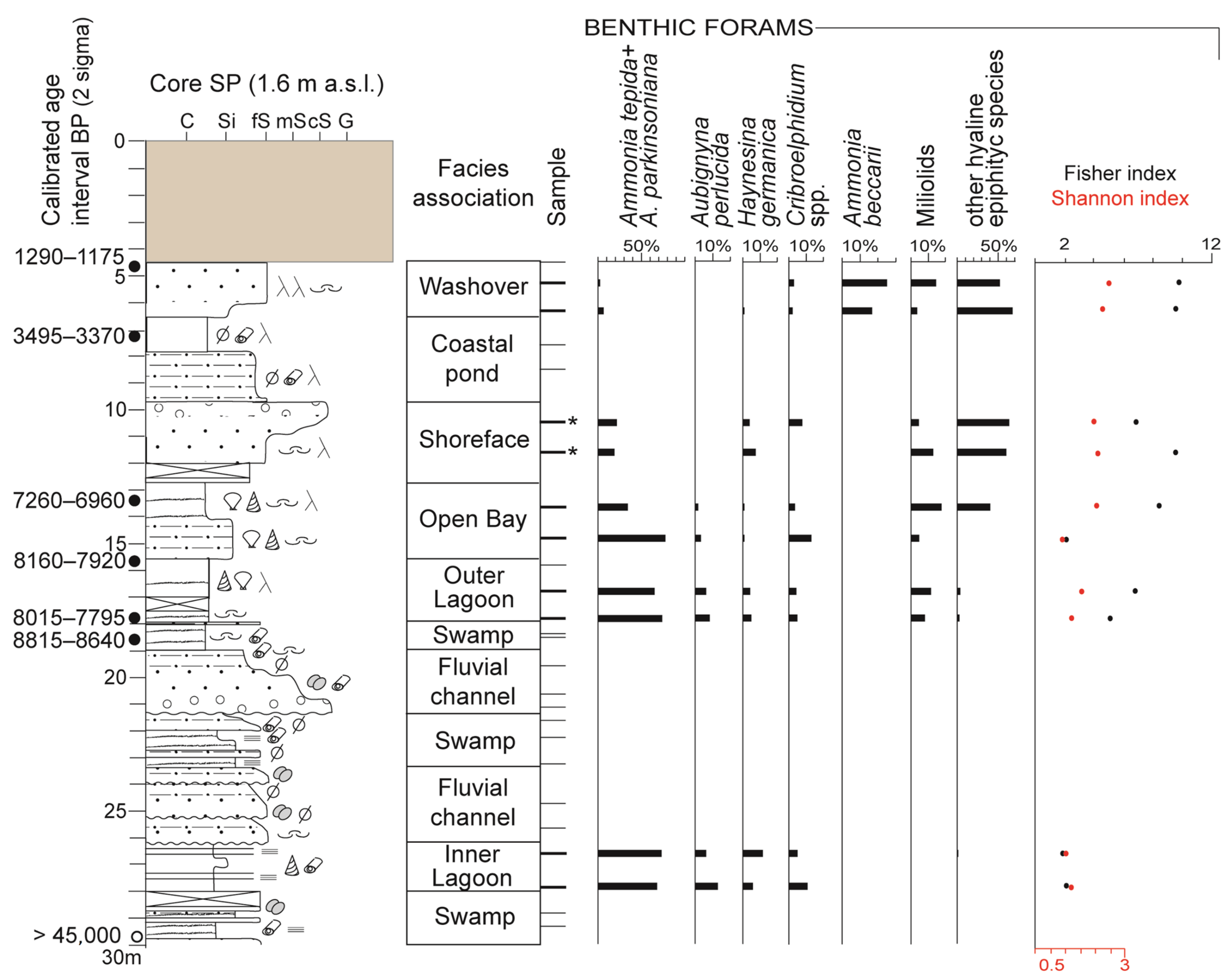
4. Results
4.1. Facies Associations
4.1.1. Fluvial-Channel Facies Association (~26–23.50 m; ~21.50–19 m)
Description
Interpretation
4.1.2. Swamp Facies Association (~30–28 m; ~23.50–21.50 m; 19–18 m)
Description
Interpretation
4.1.3. Coastal pond Facies Association (~9.80–6.50 m)
Description
Interpretation
4.1.4. Inner Lagoon Facies Association (~28–26 m)
Description
Interpretation
4.1.5. Outer Lagoon Facies Association (~18–15.50 m)
Description
Interpretation
4.1.6. Open bay Facies Association (~15.50–12.60 m)
Description
Interpretation
4.1.7. Shoreface Facies Association (~12.60–9.80 m)
Description
Interpretation
4.1.8. Washover Facies Association (~6.50–4.50 m)
Description
Interpretation
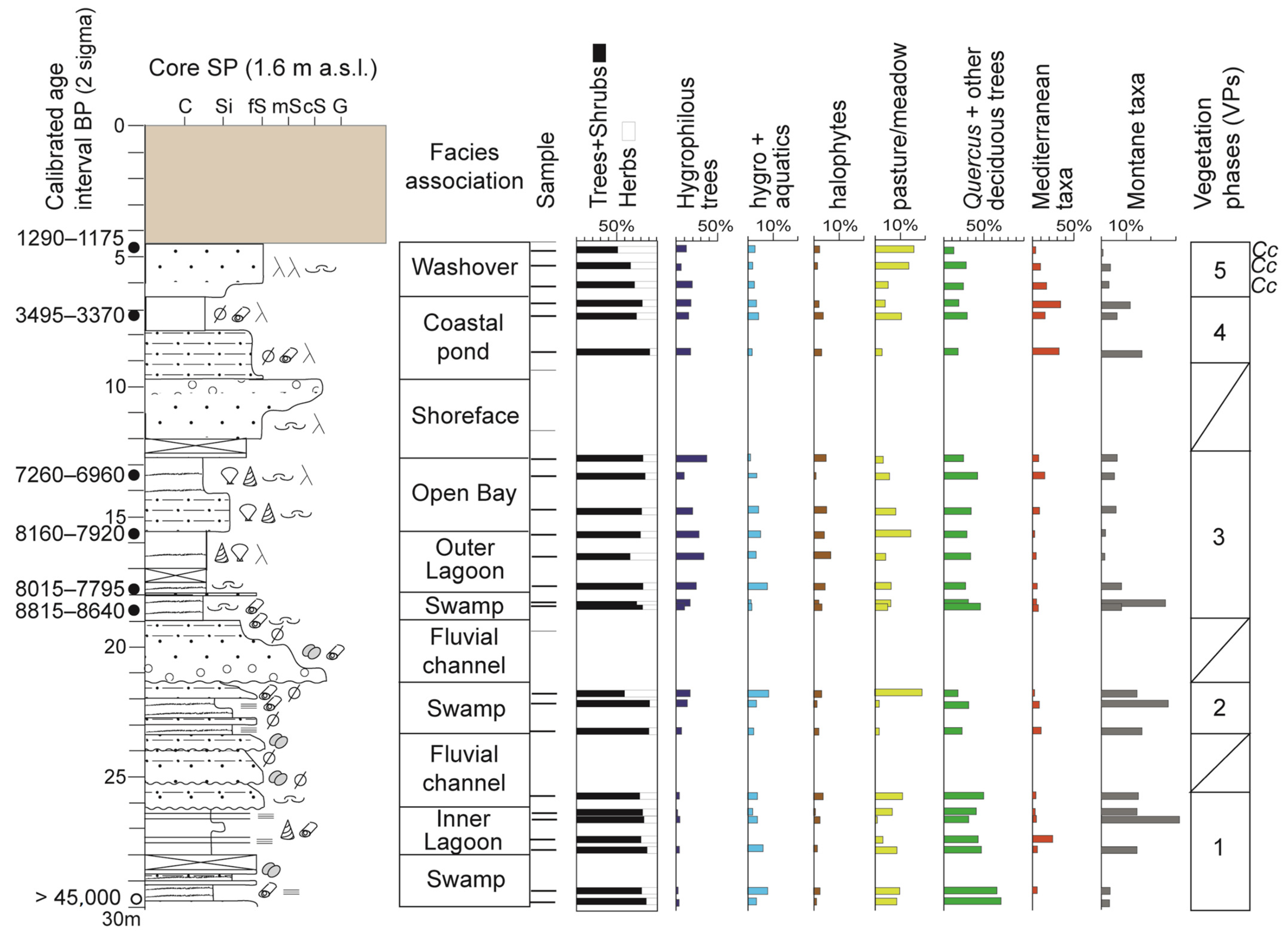
4.2. Vegetation Phases
4.2.1. VP 1
4.2.2. VP 2
4.2.3. VP 3
4.2.4. VP 4
4.2.5. VP 5
5. Discussion
5.1. Late Pleistocene Barrier-Lagoon System
5.2. Late Pleistocene Alluvial System
5.3. Early Holocene Barrier-Lagoon System
5.4. Mid-Late Holocene Coastal Bay System
6. Conclusions
- The interplay between Milankovitch-scale eustatic-climate oscillations and inherited basin morphologies shaped the GLS landscape through time, leading to the alternation of transitional-shallow marine and alluvial environments during interglacial and glacial periods, respectively;
- The GLS rocky embayment proved to be a very dynamic setting able to host a variety of transitional-shallow marine environments during interglacial periods. All sedimentary facies developed under highstand or transgressive RSL conditions showed a low degree of fluvial influence, consistent with the present-day hydrographic context. Even under high confined conditions (those of inner lagoons, coastal ponds, etc.) river fluxes played a negligible role in the environmental dynamics, as testified by the absence of a freshwater-hypohaline ostracod fauna and by the formation of gypsum crystals/rosettes. Only during the last glacial period was the gulf subject to strong sedimentary inputs from the hinterland, and an alluvial fan progradation occurred favoured by the proximity to steep promontories;
- Pollen taxa/groups mainly reflecting the local pollen rain proved to be excellent palaeoenvironmental indicators, which strengthened and complemented the information derived from the meiofaunal and sedimentological records. More specifically, pollen assemblages reflect the degree of hygrophily/humidity in the coastal plain (hygrophilous trees and hygro + aquatic herbs), offering a complete picture of gulf palaeolandscapes. Moreover, the group of halophytes made it possible to update and refine the environmental information derived from the meiofauna in terms of salinity and degree of confinement;
- The relative abundances of mesophilous taxa (Quercetum taxa + other deciduous trees), Mediterranean taxa (mainly Q. ilex) and Montane taxa (mainly A. alba) helped to track the main changes in regional vegetation cover during the late Quaternary, and to provide vegetation-derived palaeoclimate inferences at a Milankovitch timescale. Apart from present interglacial (Holocene) vegetation phases (VPs 5–3), two older VPs were recognised and assigned to the last glacial (VP 2) and interglacial (VP 1) periods. The high tree cover (>50–60%) that characterised all the VPs and the particular composition of VP 2 (co-dominance of cool-temperate and meso-thermophilous trees) suggest that GLS acted as an important tree refuge, especially during glacial times, when favourable physiographic features (high promontories surrounding the gulf and proximity to the sea) are likely to have fostered mild temperatures and high moisture conditions;
- The Holocene RSL rise caused GLS inundation and the establishment at around 9000–8500 cal year BP of a low-confined lagoon environment, protected seaward by an inherited relief (sandy barrier) located at ~13–12 m water depths on the gulf bottom. Around 8000 cal year BP, the barrier was submerged under accelerating RSL rise, which led to the development of the modern coastal bay, only partially sheltered by the surrounding promontories. This phase, which also includes shoreline progradation under RSL highstand (last 7000 years), occurred during optimum-like vegetation conditions that showed an increase in mediterraneity during the last few millennia (VPs 4–5);
- The Middle-Late Holocene depositional and palaeoenvironmental dynamics, occurred within the GLS, reflect evolutionary patterns that move away from those already documented for several Western Mediterranean plains. The superposition of shoreface and coastal wetlands deposits onto open bay sediments tracks the progradation of the coastline into a rocky embayment subject to predominant marine processes;
- Until the foundation of the old town of La Spezia in the early Middle Ages, coastal wetlands represented natural topographic lows in the coastal plain, where the hygrophyte plants flourished. The earliest evidence of human disturbance in the vegetation landscape dates back to pre-Roman times, about 3000 cal year BP, as highlighted by the stratigraphic records of cultivated plants. A dramatic increase of Castanea sativa is recorded along the hill slopes during the early medieval period.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carbognin, L.; Teatini, P.; Tomasin, A.; Tosi, L. Global Change and Relative Sea Level Rise at Venice: What Impact in Term of Flooding. Clim. Dyn. 2010, 35, 1055–1063. [Google Scholar] [CrossRef]
- Marsico, A.; Lisco, S.; Lo Presti, V.; Antonioli, F.; Amorosi, A.; Anzidei, M.; Deiana, G.; De Falco, G.; Fontana, A.; Fontolan, G.; et al. Flooding Scenario for Four Italian Coastal Plains Using Three Relative Sea Level Rise Models. J. Maps 2017, 13, 961–967. [Google Scholar] [CrossRef]
- Antonioli, F.; De Falco, G.; Lo Presti, V.; Moretti, L.; Scardino, G.; Anzidei, M.; Bonaldo, D.; Carniel, S.; Leoni, G.; Furlani, S.; et al. Relative Sea-Level Rise and Potential Submersion Risk for 2100 on 16 Coastal Plains of the Mediterranean Sea. Water 2020, 12, 2173. [Google Scholar] [CrossRef]
- Di Rita, F.; Simone, O.; Caldara, M.; Gehrels, W.R.; Magri, D. Holocene Environmental Changes in the Coastal Tavoliere Plain (Apulia, Southern Italy): A Multiproxy Approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 139–151. [Google Scholar] [CrossRef]
- Di Rita, F.; Molisso, F.; Sacchi, M. Late Holocene Environmental Dynamics, Vegetation History, Human Impact, and Climate Change in the Ancient Literna Palus (Lago Patria; Campania, Italy). Rev. Palaeobot. Palynol. 2018, 258, 48–61. [Google Scholar] [CrossRef]
- Longhitano, S.G.; Della Luna, R.; Milone, A.L.; Cilumbriello, A.; Spilotro, G. The 20,000-years-long sedimentary record of the Lesina coastal system (southern Italy): From alluvial, to tidal, to wave process regime change. Holocene 2015, 5, 18–21. [Google Scholar] [CrossRef]
- Bellotti, P.; Calderoni, G.; Dall’Aglio, P.L.; D’Amico, C.; Davoli, L.; Di Bella, L.; D’Orefice, M.; Esu, D.; Ferrari, K.; Bandini Mazzanti, M.; et al. Middle-to Late-Holocene Environmental Changes in the Garigliano Delta Plain (Central Italy): Which Landscape Witnessed the Development of the Minturnae Roman Colony? Holocene 2016, 26, 1457–1471. [Google Scholar] [CrossRef]
- Ejarque, A.; Julià, R.; Reed, J.M.; Mesquita-Joanes, F.; Barba, J.M.; Riera, S. Coastal Evolution in a Mediterranean Microtidal Zone: Mid to Late Holocene Natural Dynamics and Human Management of the Castelló Lagoon, NE Spain. PLoS ONE 2016, 11, e0155446. [Google Scholar] [CrossRef] [PubMed]
- Melis, R.T.; Di Rita, F.; French, C.; Marriner, N.; Montis, F.; Serreli, G.; Sulas, F.; Vacchi, M. 8000 years of Coastal Changes on a Western Mediterranean Island: A Multiproxy Approach from the Posada Plain of Sardinia. Mar. Geol. 2018, 403, 93–108. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, A.; Blázquez, A.M.; Guillem, J.; Usera, J. Maximum Flood Area during MIS 1 in the Almenara Marshland (Western Mediterranean): Benthic Foraminifera and Sedimentary Record. Holocene 2018, 28, 1452–1466. [Google Scholar] [CrossRef]
- Revelles, J.; Ghilardi, M.; Rossi, V.; Currás, A.; López-Bultó, O.; Brkojewitsch, G.; Vacchi, M. Coastal Landscape Evolution of Corsica Island (W. Mediterranean): Palaeoenvironments, Vegetation History and Human Impacts since the Early Neolithic Period. Quat. Sci. Rev. 2019, 225. [Google Scholar] [CrossRef]
- D’Orefice, M.; Bellotti, P.; Bertini, A.; Calderoni, G.; Neri, P.C.; Di Bella, L.; Fiorenza, D.; Foresi, L.M.; Louvari, M.A.; Rainone, L.; et al. Holocene Evolution of the Burano Paleo-Lagoon (Southern Tuscany, Italy). Water 2020, 12, 1007. [Google Scholar] [CrossRef]
- Amorosi, A.; Colalongo, M.L.; Fusco, F.; Pasini, G.; Fiorini, F. Glacio-Eustatic Control of Continental-Shallow Marine Cyclicity from Late Quaternary Deposits of the Southeastern Po Plain, Northern Italy. Quat. Res. 1999, 52, 1–13. [Google Scholar] [CrossRef]
- Aguzzi, M.; Amorosi, A.; Colalongo, M.L.; Lucchi, M.R.; Rossi, V.; Sarti, G.; Vaiani, S.C. Late Quaternary Climatic Evolution of the Arno Coastal Plain (Western Tuscany, Italy) from Subsurface Data. Sediment. Geol. 2007, 202, 211–229. [Google Scholar] [CrossRef]
- Cacciari, M.; Amorosi, A.; Campo, B.; Marchesini, M.; Rossi, V. Palynology of the Late Quaternary Succession of the Arno Plain (Northern Italy): New Insights on Palaeoenvironmental Trends and Climate Dynamics. Boll. Soc. Paleontol. Ital. 2019, 58, 201–221. [Google Scholar] [CrossRef]
- Campo, B.; Bruno, L.; Amorosi, A. Basin-Scale Stratigraphic Correlation of Late Pleistocene-Holocene (MIS 5e–MIS 1) Strata across the Rapidly Subsiding Po Basin (Northern Italy). Quat. Sci. Rev. 2020, 237. [Google Scholar] [CrossRef]
- Dolez, L.; Salel, T.; Bruneton, H.; Colpo, G.; Devillers, B.; Lefèvre, D.; Muller, S.D.; Sanchez, C. Holocene Palaeoenvironments of the Bages-Sigean Lagoon (France). Geobios 2015, 48, 297–308. [Google Scholar] [CrossRef]
- Milli, S.; Girasoli, D.E.; Tentori, D.; Tortora, P. Sedimentology and Coastal Dynamics of Carbonate Pocket Beaches: The Ionian-Sea Apulia Coast between Torre Colimena and Porto Cesareo (Southern Italy). J. Mediterr. Earth Sci. 2017, 9, 29–66. [Google Scholar] [CrossRef]
- Aiello, G.; Amato, V.; Barra, D.; Caporaso, L.; Caruso, T.; Giaccio, B.; Parisi, R.; Rossi, A. Late Quaternary Benthic Foraminiferal and Ostracod Response to Palaeoenvironmental Changes in a Mediterranean Coastal Area, Port of Salerno, Tyrrhenian Sea. Reg. Stud. Mar. Sci. 2020, 40, 101498. [Google Scholar] [CrossRef]
- Furlani, S.; Pappalardo, M.; Gómez-Pujol, L.; Chelli, A. The rock coast of the Mediterranean and Black seas. In Rock Coast Geomophology; Kennedy, D.M., Stephenson, W.J., Naylor, L.A., Eds.; Geological Society, Memoirs: London, UK, 2014; pp. 89–123. [Google Scholar]
- Chelli, A.; Pappalardo, M.; Llopis, I.A.; Federici, P.R. The Relative Influence of Lithology and Weathering in Shaping Shore Platforms along the Coastline of the Gulf of La Spezia (NW Italy) as Revealed by Rock Strength. Geomorphology 2010, 118, 93–104. [Google Scholar] [CrossRef]
- Rovere, A.; Vacchi, M.; Firpo, M.; Carobene, L. Underwater Geomorphology of the Rocky Coastal Tracts between Finale Ligure and Vado Ligure (Western Liguria, NW Mediterranean Sea). Quat. Int. 2011, 232, 187–200. [Google Scholar] [CrossRef]
- Antonioli, F.; Lo Presti, V.; Rovere, A.; Ferranti, L.; Anzidei, M.; Furlani, S.; Mastronuzzi, G.; Orru, P.E.; Scicchitano, G.; Sannino, G.; et al. Tidal Notches in Mediterranean Sea: A Comprehensive Analysis. Quat. Sci. Rev. 2015, 119, 66–84. [Google Scholar] [CrossRef]
- Furlani, S. The Rocky Coasts of the Mediterranean Sea: Field Studies. In Proceedings of the Conférence Méditerranéenne Côtière et Maritime, Split, Croatia, 29 November–1 December 2017; pp. 1–6. [Google Scholar]
- Arobba, D.; Caramiello, R.; Firpo, M.; Mercalli, L.; Morandi, L.F.; Rossi, S. New Evidence on the Earliest Human Presence in the Urban Area of Genoa (Liguria, Italy): A Multi-Proxy Study of a Mid-Holocene Deposit at the Mouth of the Bisagno River. Holocene 2018, 28, 1918–1935. [Google Scholar] [CrossRef]
- Geoportale. Available online: http://srvcarto.regione.liguria.it/geoviewer2/pages/apps/geoportale/index.html (accessed on 27 December 2020).
- Gasparini, G.P.; Abbate, M.; Bordone, A.; Cerrati, G.; Galli, C.; Lazzoni, E.; Negri, A. Circulation and Biomass Distribution during Warm Season in the Gulf of La Spezia (North-Western Mediterranean). J. Mar. Syst. 2009, 78, 548–562. [Google Scholar] [CrossRef]
- Bellini, C.; Mariotti-Lippi, M.; Montanari, C. The Holocene Landscape History of the NW Italian Coasts. Holocene 2009, 19, 1161–1172. [Google Scholar] [CrossRef]
- Vacchi, M.; Marriner, N.; Morhange, C.; Spada, G.; Fontana, A.; Rovere, A. Multiproxy Assessment of Holocene Relative Sea-Level Changes in the Western Mediterranean: Sea-Level Variability and Improvements in the Definition of the Isostatic Signal. Earth Sci. Rev. 2016, 155, 172–197. [Google Scholar] [CrossRef]
- Breslau, L.R.; Edgerton, H.E. The Gulf of La Spezia, Italy: A case history of seismic-sedimentologic correlation. In The Mediterranean Sea: A Natural Sedimentation Laboratory; Stanlet, D.J., Ed.; Dowden, Hutchinson and Ross, Inc.: Stroudsburg, PA, USA, 1972; pp. 177–188. [Google Scholar]
- Mariani, M.; Prato, R. I Bacini Neogenici Costieri Del Margine Tirrenico: Approccio Sismico-Stratigrafico. Mem. Soc. Geol. Ital. 1988, 41, 519–531. [Google Scholar]
- Boccaletti, M.; Corti, G.; Martelli, L. Recent and Active Tectonics of the External Zone of the Northern Apennines (Italy). Int. J. Earth Sci. 2011, 100, 1331–1348. [Google Scholar] [CrossRef]
- Conti, P.; Cornamusini, G.; Carmignani, L. An Outline of the Geology of the Northern Apennines (Italy), with Geological Map at 1:250,000 Scale. Ital. J. Geosci. 2020, 139, 149–194. [Google Scholar] [CrossRef]
- Gandolfi, G.; Paganelli, L. Il Macigno Costiero fra La Spezia e Grosseto. G. Geol. 1992, 54, 163–179. [Google Scholar]
- Abbate, E.; Fanucci, F.; Benvenuti, M.; Bruni, P.; Cipriani, N.; Falorni, P.; Fazzuoli, M.; Morelli, D.; Pandeli, E.; Papini, M.; et al. Note Illustrative della Carta Geologica d’Italia alla Scala 1:50.000. Foglio 248 La Spezia; S.EL.CA. s.r.l.: Florence, Italy, 2005; 288p. [Google Scholar]
- Nisio, S.I. Sinkholes in Liguria. Mem. Descr. Cart. Geol. Ital. 2008, 85, 399–408. [Google Scholar]
- Climate Data. Dati Climatici Sulle Città del Mondo. Available online: https://it.climate-data.org/ (accessed on 27 December 2020).
- Barberis, G.; Peccenini, S.; Vagge, I. Formazioni Arbustive Mediterranee in Liguria. Fitosociologia 1997, 32, 61–71. [Google Scholar]
- Vagge, I. La Diffusione Del Bioclima Mediterraneo in Liguria (Italia Nord Occidentale). Fitosociologia 2014, 36, 95–109. [Google Scholar]
- Debenay, J.P.; Guillou, J.J. Ecological Transitions Indicated by Foraminiferal Assemblages in Paralic Environments. Estuaries 2002, 25, 1107–1120. [Google Scholar] [CrossRef]
- Frenzel, P.; Boomer, I. The Use of Ostracods from Marginal Marine, Brackish Waters as Bioindicators of Modern and Quaternary Environmental Change. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 225, 68–92. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Horne, D.J.; Curry, B.B.; Mesquita-Joanes, F. Mutual climatic range methods for Quaternary ostracods. In Ostracoda as Proxies for Quaternary Climate Change. Developments in Quaternary Science; Horne, D.J., Holmes, J.A., Rodriguez-Lazaro, J., Viehberg, F.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 65–84. [Google Scholar]
- Milli, S.; D’Ambrogi, C.; Bellotti, P.; Calderoni, G.; Carboni, M.G.; Celant, A.; Di Bella, L.; Di Rita, F.; Frezza, V.; Magri, D.; et al. The Transition from Wave-Dominated Estuary to Wave-Dominated Delta: The Late Quaternary Stratigraphic Architecture of Tiber River Deltaic Succession (Italy). Sediment. Geol. 2013, 284–285, 159–180. [Google Scholar] [CrossRef]
- Bini, M.; Brückner, H.; Chelli, A.; Pappalardo, M.; Da Prato, S.; Gervasini, L. Palaeogeographies of the Magra Valley Coastal Plain to Constrain the Location of the Roman Harbour of Luna (NW Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 337–338, 37–51. [Google Scholar] [CrossRef]
- Barbieri, G.; Vaiani, S.C. Benthic Foraminifera or Ostracoda? Comparing the Accuracy of Palaeoenvironmental Indicators from a Pleistocene Lagoon of the Romagna Coastal Plain (Italy). J. Micropalaeontol. 2018, 37, 203–230. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T. Paleontological Data Analysis; Wiley-Blackwell: New York, NY, USA, 2005. [Google Scholar]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The Relation between the Number of Species and the Number of Individuals in a Random Sample of an Animal Population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar] [CrossRef]
- Shannon, C.A. Mathematical Theory of Communication. Bell Syst. Technol. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Lowe, J.J.; Accorsi, C.A.; Bandini Mazzanti, M.; Bishop, V.; der Kaars, S.; Forlani, L.; Mercuri, A.M.; Rivalenti, C.; Torri, P.; Watson, C. Pollen Stratigraphy of Sediment Sequences from Carter Lakes Albano and Nemi (near Rome) and from the Central Adriatic, Spanning the Interval from Oxygen Isotope Stage 2 to the Present Day. Mem. Ist. Ital. Idrobiol. 1996, 55, 71–98. [Google Scholar]
- Pignatti, S. I Piani Di Vegetazione in Italia. G. Bot. Ital. 1979, 113, 411–428. [Google Scholar] [CrossRef]
- Stuiver, M.; Reimer, P.J.; Reimer, R.W. CALIB 8.2. Available online: http://calib.org (accessed on 25 December 2020).
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 Cal KBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Amorosi, A.; Rossi, V.; Scarponi, D.; Vaiani, S.C.; Ghosh, A. Biosedimentary Record of Postglacial Coastal Dynamics: High-Resolution Sequence Stratigraphy from the Northern Tuscan Coast (Italy). Boreas 2014, 43, 939–954. [Google Scholar] [CrossRef]
- Cacciari, M.; Amorosi, A.; Marchesini, M.; Kaniewski, D.; Bruno, L.; Campo, B.; Rossi, V. Linking Holocene Vegetation Dynamics, Palaeoclimate Variability and Depositional Patterns in Coastal Successions: Insights from the Po Delta Plain of Northern Italy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 538, 109468. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Mazzanti, M.B.; Torri, P.; Vigliotti, L.; Bosi, G.; Florenzano, A.; Olmi, L.; N’siala, I.M. A Marine/Terrestrial Integration for Mid-Late Holocene Vegetation History and the Development of the Cultural Landscape in the Po Valley as a Result of Human Impact and Climate Change. Veg. Hist. Archaeobot. 2012, 21, 353–372. [Google Scholar] [CrossRef]
- Accorsi, C.A.; Bandini Mazzanti, M.; Forlani, L.; Mercuri, A.M.; Trevisan Grandi, G. An Overview of Holocene Forest Pollen Flora/ Vegetation of the Emilia Romagna Region—Northern Italy. Arch. Geobot. 1999, 5, 3–37. [Google Scholar]
- Federici, P.R.; Pappalardo, M. Evidence of Marine Isotope Stage 5.5 Highstand in Liguria (Italy) and Its Tectonic Significance. Quat. Int. 2006, 145–146, 68–77. [Google Scholar] [CrossRef]
- Ferranti, L.; Antonioli, F.; Mauz, B.; Amorosi, A.; Dai Pra, G.; Mastronuzzi, G.; Monaco, C.; Orrù, P.; Pappalardo, M.; Radtke, U.; et al. Markers of the Last Interglacial Sea-Level High Stand along the Coast of Italy: Tectonic Implications. Quat. Int. 2006, 145–146, 30–54. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.M.; Carraro, G.; Conedera, M.; et al. The Past Ecology of Abies Alba Provides New Perspectives on Future Responses of Silver Fir Forests to Global Warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef]
- Federici, P.R. La tettonica recente dell’Appennino: I—Il bacino villafranchiano di Sarzana e il suo significato nel quadro dei movimenti distensivi a nord-ovest delle Alpi Apuane. Boll. Soc. Geol. Ital. Ital. J. Geosci. 1973, 92, 287–301. [Google Scholar]
- Bond, G.; Showers, W.; Cheseby, M.; Lotti, R.; Almasi, P.; DeMenocal, P.; Priore, P.; Cullen, H.; Hajdas, I.; Bonani, G. A Pervasive Millennial-Scale Cycle in North Atlantic Holocene and Glacial Climates. Science 1997, 278, 1257–1266. [Google Scholar] [CrossRef]
- Zanchetta, G.; Isola, I.; Piccini, L.; Dini, A. The Corchia Cave (Alpi Apuane): A 2 Ma Long Temporal Window on the Earth Climate. Geol. Field Trips 2011, 3, 1–55. [Google Scholar] [CrossRef]
- Zhornyak, L.V.; Zanchetta, G.; Drysdale, R.N.; Hellstrom, J.C.; Isola, I.; Regattieri, E.; Piccini, L.; Baneschi, I.; Couchoud, I. Stratigraphic Evidence for a “Pluvial Phase” between ca 8200–7100 Ka from Renella Cave (Central Italy). Quat. Sci. Rev. 2011, 30, 409–417. [Google Scholar] [CrossRef]
- Regattieri, E.; Zanchetta, G.; Drysdale, R.N.; Isola, I.; Hellstrom, L.D. Lateglacial to Holocene Trace Element Record (Ba, Mg, Sr) from Corchia Cave (Apuan Alps, Central Italy): Paleoenvironmental Implications. J. Quat. Sci. 2014, 29, 381–392. [Google Scholar] [CrossRef]
- Carboni, M.G.; Bergamin, L.; Di Bella, L.; Esu, D.; Cerone, E.P.; Antonioli, F.; Verrubbi, V. Palaeoenvironmental Reconstruction of Late Quaternary Foraminifera and Molluscs from the ENEA Borehole (Versilian Plain, Tuscany, Italy). Quat. Res. 2010, 74, 265–276. [Google Scholar] [CrossRef]
- Bellotti, P.; Caputo, C.; Davoli, L.; Evangelista, S.; Garzanti, E.; Pugliese, F.; Valeri, P. Morpho-Sedimentary Characteristics and Holocene Evolution of the Emergent Part of the Ombrone River Delta (Southern Tuscany). Geomorphology 2004, 61, 71–90. [Google Scholar] [CrossRef]
- Di Rita, F.; Celant, A.; Milli, S.; Magri, D. Lateglacial-Early Holocene Vegetation History of the Tiber Delta (Rome, Italy) under the Influence of Climate Change and Sea Level Rise. Rev. Palaeobot. Palynol. 2015, 218, 204–216. [Google Scholar] [CrossRef]
- Amorosi, A.; Bruno, L.; Campo, B.; Morelli, A.; Rossi, V.; Scarponi, D.; Hong, W.; Bohacs, K.M.; Drexler, T.M. Global Sea-Level Control on Local Parasequence Architecture from the Holocene Record of the Po Plain, Italy. Mar. Pet. Geol. 2017, 87, 99–111. [Google Scholar] [CrossRef]
- Amorosi, A.; Bracone, V.; Campo, B.; D’Amico, C.; Rossi, V.; Rosskopf, C.M. A Late Quaternary Multiple Paleovalley System from the Adriatic Coastal Plain (Biferno River, Southern Italy). Geomorphology 2016, 254, 146–159. [Google Scholar] [CrossRef]
- Treccani. La Spezia. Available online: https://www.treccani.it/enciclopedia/la-spezia (accessed on 28 December 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Core Depth (m) | Material | Conventional Age (Year BP) | Calibrated 2σ Age (cal Year BP) | Calibration Dataset | Dating Laboratory; Sample Laboratory Number |
|---|---|---|---|---|---|
| 4.70 | Plant remains | 1268 ± 45 | 1290–1175 | IntCal20 | CEDAD Laboratory (Lecce, Italy); LTL16734A |
| 7.30 | Organic sediments | 3230 ± 36 | 3495–3370 | IntCal20 | Innova Laboratory (Caserta, Italy); DSH9969_SO |
| 13.40 | Organic sediments | 6509 ± 45 | 7260–6960 | Mixed Marine NoHem | CEDAD Laboratory (Lecce, Italy); LTL16735A |
| 15.55 | Organic sediments | 7457 ± 37 | 8160–7920 | Mixed Marine NoHem | Innova Laboratory (Caserta, Italy); DSH10001_SO |
| 17.70 | Organic sediments | 7369 ± 39 | 8015–7795 | Mixed Marine NoHem | Innova Laboratory (Caserta, Italy); DSH9967_SO |
| 18.70 | Organic sediments | 7941 ± 37 | 8815–8640 | IntCal20 | Innova Laboratory (Caserta, Italy); DSH9970_SO |
| 29.80 | Organic sediments | >45,000 | - | - | CEDAD Laboratory (Lecce, Italy); LTL16736A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, V.; Amorosi, A.; Marchesini, M.; Marvelli, S.; Cocchianella, A.; Lorenzini, L.; Trigona, S.L.; Valle, G.; Bini, M. Late Quaternary Landscape Dynamics at the La Spezia Gulf (NW Italy): A Multi-Proxy Approach Reveals Environmental Variability within a Rocky Embayment. Water 2021, 13, 427. https://doi.org/10.3390/w13040427
Rossi V, Amorosi A, Marchesini M, Marvelli S, Cocchianella A, Lorenzini L, Trigona SL, Valle G, Bini M. Late Quaternary Landscape Dynamics at the La Spezia Gulf (NW Italy): A Multi-Proxy Approach Reveals Environmental Variability within a Rocky Embayment. Water. 2021; 13(4):427. https://doi.org/10.3390/w13040427
Chicago/Turabian StyleRossi, Veronica, Alessandro Amorosi, Marco Marchesini, Silvia Marvelli, Andrea Cocchianella, Luca Lorenzini, Simon Luca Trigona, Gianfranco Valle, and Monica Bini. 2021. "Late Quaternary Landscape Dynamics at the La Spezia Gulf (NW Italy): A Multi-Proxy Approach Reveals Environmental Variability within a Rocky Embayment" Water 13, no. 4: 427. https://doi.org/10.3390/w13040427
APA StyleRossi, V., Amorosi, A., Marchesini, M., Marvelli, S., Cocchianella, A., Lorenzini, L., Trigona, S. L., Valle, G., & Bini, M. (2021). Late Quaternary Landscape Dynamics at the La Spezia Gulf (NW Italy): A Multi-Proxy Approach Reveals Environmental Variability within a Rocky Embayment. Water, 13(4), 427. https://doi.org/10.3390/w13040427