
Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats

Abstract
1. Introduction
2. Materials and Methods
2.1. Site Characterization and Sampling
2.2. Genomics and Data Analysis
3. Results
3.1. Water Chemistry
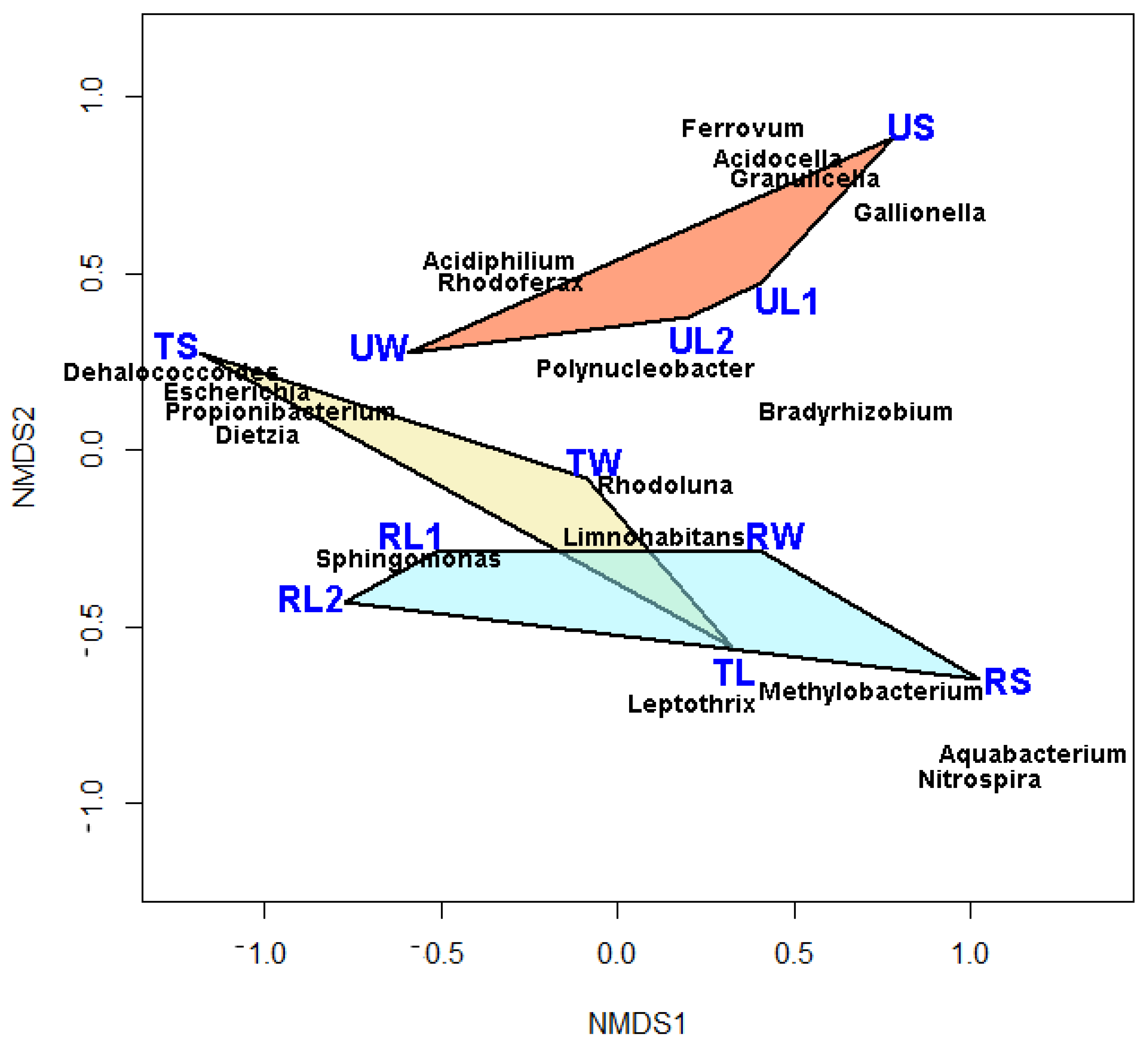
3.2. Microbial Community Composition
4. Discussion
4.1. Water Chemistry
4.2. Microbial Community Composition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rothschild, L.; Mancinelli, R. Life in extreme environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.; Banfield, J. Microbial communities in acid mine drainage. FEMS Microbiol. Ecol. 2003, 44, 139–152. [Google Scholar] [CrossRef]
- Aguinaga, O.E.; McMahon, A.; White, K.N.; Dean, A.P.; Pittman, J.K. Microbial community shifts in response to acid mine drainage pollution within a natural wetland ecosystem. Front. Microbiol. 2018, 9, 1445. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.V.; Mooney, H.A.; Cropper, A.; Capistrano, D.; Carpenter, S.R.; Chopra, K.; Dasgupta, P.; Dietz, T.; Duraiappah, A.K.; Hassan, R.; et al. Ecosystems and Human Well-Being-SYNTHESIS: A Report of the Millennium Ecosystem Assessment; Sarukhan, J., Whyte, A., Board, M., Eds.; Island Press: Washington, DC, USA, 2005; Volume 5. [Google Scholar]
- Younger, P.L.; Banwart, S.A.; Hedin, R.S. Mine water hydrology. In Mine Water; Springer: Dordrecht, The Netherlands, 2002; Volume 5, pp. 127–270. [Google Scholar] [CrossRef]
- Simate, G.S.; Ndlovu, S. Acid mine drainage: Challenges and opportunities. J. Environ. Chem. Eng. 2014, 2, 1785–1803. [Google Scholar] [CrossRef]
- Watzlaf, G.R.; Schroeder, K.T.; Kleinmann, R.L.; Kairies, C.L.; Nairn, R.W. The Passive Treatment of Coal Mine Drainage; National Energy Technology Laboratory: Pittsburgh, PA, USA, 2004; pp. 1–72. [Google Scholar]
- Hogsden, K.L.; Harding, J.S. Anthropogenic and natural sources of acidity and metals and their influence on the structure of stream food webs. Environ. Pollut. 2012, 162, 466–474. [Google Scholar] [CrossRef]
- Johnson, D.B.; Hallberg, K.B. Pitfalls of passive mine water treatment. Rev. Environ. Sci. Biotechnol. 2002, 1, 335–343. [Google Scholar] [CrossRef]
- Ziemkiewicz, P.F.; Skousen, J.G.; Simmons, J. Long-term performance of passive acid mine drainage treatment systems. Mine Water Environ. 2003, 22, 118–129. [Google Scholar] [CrossRef]
- Skousen, J.; Zipper, C.E.; Rose, A.; Ziemkiewicz, P.F.; Nairn, R.; McDonald, L.M.; Kleinmann, R.L. Review of passive systems for acid mine drainage treatment. Mine Water Environ. 2017, 36, 133–153. [Google Scholar] [CrossRef]
- Nelson, S.M.; Roline, R.A. Recovery of a stream macroinvertebrate community from mine drainage disturbance. Hydrobiologia 1996, 339, 73–84. [Google Scholar] [CrossRef]
- DeNicola, D.M.; Stapleton, M.G. Benthic diatoms as indicators of long-term changes in a watershed receiving passive treatment for acid mine drainage. Hydrobiologia 2014, 732, 29–48. [Google Scholar] [CrossRef]
- Williams, K.M.; Turner, A.M. Acid mine drainage and stream recovery: Effects of restoration on water quality, macroinvertebrates, and fish. Knowl. Manag. Aquat. Ecosyst. 2015, 416, 18. [Google Scholar] [CrossRef]
- Rawat, N.; Singh, G. The role of micro-organisms in the formation of acid mine drainage in the north-eastern coal field of India. Int. J. Mine Water 1982, 2, 29–36. [Google Scholar] [CrossRef]
- Ly, T.; Wright, J.R.; Weit, N.; McLimans, C.J.; Ulrich, N.; Tokarev, V.; Valkanas, M.M.; Trun, N.; Rummel, S.; Grant, C.J.; et al. Microbial communities associated with passive acidic abandoned coal mine remediation. Front. Microbiol. 2019, 10, 1955. [Google Scholar] [CrossRef] [PubMed]
- Allan, J.D.; Castillo, M.M.; Capps, K. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Niyogi, D.K.; Lewis, W.M., Jr.; McKnight, D.M. Litter breakdown in mountain streams affected by mine drainage: Biotic mediation of abiotic controls. Ecol. Appl. 2001, 11, 506–516. [Google Scholar] [CrossRef]
- Schlief, J. Leaf associated microbial activities in a stream affected by acid mine drainage. Int. Rev. Hydrobiol. 2004, 89, 467–475. [Google Scholar] [CrossRef]
- Baeseman, J.L.; Smith, R.L.; Silverstein, J. Denitrification potential in stream sediments impacted by acid mine drainage: Effects of pH, various electron donors, and iron. Microb. Ecol. 2006, 51, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Bott, T.L.; Jackson, J.K.; McTammany, M.E.; Newbold, J.D.; Rier, S.T.; Sweeney, B.W.; Battle, J.M. Abandoned coal mine drainage and its remediation: Impacts on stream ecosystem structure and function. Ecol. Appl. 2012, 22, 2144–2163. [Google Scholar] [CrossRef]
- Lear, G.; Niyogi, D.; Harding, J.; Dong, Y.; Lewis, G. Biofilm bacterial community structure in streams affected by acid mine drainage. Appl. Environ. Microbiol. 2009, 75, 3455. [Google Scholar] [CrossRef] [PubMed]
- Kuang, J.L.; Huang, L.N.; Chen, L.X.; Hua, Z.S.; Li, S.J.; Hu, M.; Li, J.T.; Shu, W.S. Contemporary environmental variation determines microbial diversity patterns in acid mine drainage. ISME J. 2013, 7, 1038–1050. [Google Scholar] [CrossRef]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar] [CrossRef] [PubMed]
- Findlay, S. Stream microbial ecology. J. N. Am. Benthol. Soc. 2010, 29, 170–181. [Google Scholar] [CrossRef]
- Webster, J.R.; Benfield, E.F. Vascular plant breakdown in freshwater ecosystems. Annu. Rev. Ecol. Evol. Syst. 1986, 17, 567–594. [Google Scholar] [CrossRef]
- Simmons, J.A.; Lawrence, E.R.; Jones, T.G. Treated and untreated acid mine drainage effects on stream periphyton biomass, leaf decomposition, and macroinvertebrate diversity. J. Freshwater Ecol. 2005, 20, 413–424. [Google Scholar] [CrossRef][Green Version]
- Johnson, K.S.; Thompson, P.C.; Gromen, L.; Bowman, J. Use of leaf litter breakdown and macroinvertebrates to evaluate gradient of recovery in an acid mine impacted stream remediated with an active alkaline doser. Environ. Monit. Assess. 2014, 186, 4111–4127. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.; Odum, W.E.; Mills, A. Leaf litter decomposition in a reservoir affected by acid mine drainage. Oikos 1983, 41, 165–172. [Google Scholar] [CrossRef]
- McNamara, C.J.; Leff, L.G. Bacterial community composition in biofilms on leaves in a northeastern Ohio stream. J. N. Am. Benthol. Soc. 2004, 23, 677–685. [Google Scholar] [CrossRef]
- Datashed. Available online: https://www.datashed.org/ (accessed on 3 November 2021).
- American Public Health Association. Standard Methods for the Evaluation of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1998; p. 766. [Google Scholar]
- DeNicola, D.M.; Lellock, A.J. Nutrient limitation of algal periphyton in streams along an acid mine drainage gradient. J. Phycol. 2015, 51, 739–749. [Google Scholar] [CrossRef] [PubMed]
- DeNicola, D.M.; Stapleton, M.G.; Onufrak, A.J. Examination of effects of acid and metals on benthic algae in streams using chemical diffusing substrata. Ecol. Indic. 2018, 88, 17–26. [Google Scholar] [CrossRef]
- Lind, O.T. Handbook of Common Methods in Limnology, 2nd ed.; Kendall/Hunt: Dubuque, IA, USA, 1985; p. 199. [Google Scholar]
- Bower, C.E.; Holm-Hansen, T. A salicylate-hypochloritemethod for determining ammonia in seawater. Can. J. Fish. Aquat. Sci. 1980, 37, 794–798. [Google Scholar] [CrossRef]
- Chen, W.P.; Kuo, T.T. A simple and rapid method for the preparation of gram-negative bacterial genomic DNA. Nucleic Acids Res. 1993, 21, 2260. [Google Scholar] [CrossRef]
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate Multiscale Spatial Analysis [R Package Version 0.0-9]. 2019. Available online: https://cran.r-project.org/web/packages/adespatial (accessed on 13 October 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.B.; Simpson, G.; Solymos, P.; et al. Vegan [R Package Version 2.5-6]. 2020. Available online: https://cran.r-project.org/package=vegan (accessed on 13 October 2021).
- Drerup, S.A.; Vis, M.L. Extracellular enzyme activity suggests phosphorus limitation of biofilm productivity in acid mine drainage remediated streams. Hydrobiologia 2017, 784, 225–236. [Google Scholar] [CrossRef]
- Kasalický, V.; Jezbera, J.; Hahn, M.W.; Šimek, K. The diversity of the Limnohabitans genus, an important group of freshwater bacterioplankton, by characterization of 35 isolated strains. PLoS ONE 2013, 8, e58209. [Google Scholar] [CrossRef]
- Hahn, M.W.; Schmidt, J.; Taipale, S.J.; Doolittle, W.F.; Koll, U. Rhodoluna lacicola gen. nov., sp. nov., a planktonic freshwater bacterium with stream-lined genome. IJSEM 2014, 64, 3254. [Google Scholar] [CrossRef] [PubMed]
- Kalmbach, S.; Manz, W.; Bendinger, B.; Szewzyk, U. In situ probing reveals Aquabacterium commune as a widespread and highly abundant bacterial species in drinking water biofilms. Water Res. 2000, 34, 575–581. [Google Scholar] [CrossRef]
- Koch, H.; Lücker, S.; Albertsen, M.; Kitzinger, K.; Herbold, C.; Spieck, E.; Nielsen, P.H.; Wagner, M.; Daims, H. Expanded metabolic versatility of ubiquitous nitrite-oxidizing bacteria from the genus Nitrospira. Proc. Natl. Acad. Sci. USA 2015, 112, 11371–11376. [Google Scholar] [CrossRef]
- Korajkic, A.; Wanjugi, P.; Brooks, L.; Cao, Y.; Harwood, V.J. Persistence and decay of fecal microbiota in aquatic habitats. Microbiol. Mol. Biol. Rev. 2019, 83, e00005-19. [Google Scholar] [CrossRef]
- Hayashi, S.; Furusaka, C. Enrichment of Propionibacterium in paddy soil by addition of various organic substances. Antonie Van Leeuwenhoek 1980, 46, 313–320. [Google Scholar] [CrossRef]
- Fabisch, M.; Freyer, G.; Johnson, C.A.; Büchel, G.; Akob, D.M.; Neu, T.R.; Küsel, K. Dominance of ‘Gallionella capsiferriformans’ and heavy metal association with Gallionella-like stalks in metal-rich pH 6 mine water discharge. Geobiology 2016, 14, 68–90. [Google Scholar] [CrossRef]
- Percak-Dennett, E.M. Microbial Iron Redox Cycling in Terrestrial Environments. Ph.D. Thesis, The University of Wisconsin-Madison, Madison, WI, USA, 2014. [Google Scholar]
- Giddings, L.A.; Chlipala, G.; Kunstman, K.; Green, S.; Morillo, K.; Bhave, K.; Peterson, H.; Driscoll, H.; Maienschein-Cline, M. Characterization of an acid rock drainage microbiome and transcriptome at the Ely Copper Mine Superfund site. PLoS ONE 2020, 15, e0237599. [Google Scholar] [CrossRef]
- Ormeño-Orrillo, E.; Martínez-Romero, E. A genomotaxonomy view of the Bradyrhizobium genus. Front. Microbiol. 2019, 10, 1334. [Google Scholar] [CrossRef]
- Tláskal, V.; Voříšková, J.; Baldrian, P. Bacterial succession on decomposing leaf litter exhibits a specific occurrence pattern of cellulolytic taxa and potential decomposers of fungal mycelia. FEMS Microbiol. Ecol. 2016, 92, fiw177. [Google Scholar] [CrossRef]
- Gołębiewski, M.; Tarasek, A.; Sikora, M.; Deja-Sikora, E.; Tretyn, A.; Niklińska, M. Rapid microbial community changes during initial stages of pine litter decomposition. Microb. Ecol. 2019, 77, 56–75. [Google Scholar] [CrossRef]
- Mcnamara, C.J.; Leff, L.G. Response of biofilm bacteria to dissolved organic matter from decomposing maple leaves. Microb. Ecol. 2004, 48, 324–330. [Google Scholar] [CrossRef]
- Newman, M.M.; Liles, M.R.; Feminella, J.W. Litter breakdown and microbial succession on two submerged leaf species in a small forested stream. PLoS ONE 2015, 10, e0130801. [Google Scholar] [CrossRef]
- Edberg, F.; Andersson, A.F.; Holmström, S.J. Bacterial community composition in the water column of a lake formed by a former uranium open pit mine. Microb. Ecol. 2012, 64, 870–880. [Google Scholar] [CrossRef]
- Ettamimi, S.; Carlier, J.D.; Cox, C.J.; Elamine, Y.; Hammani, K.; Ghazal, H.; Costa, M.C. A meta-taxonomic investigation of the prokaryotic diversity of water bodies impacted by acid mine drainage from the São Domingos mine in southern Portugal. Extremophiles 2019, 23, 821–834. [Google Scholar] [CrossRef]
- Ferreira, V.; Koricheva, J.; Duarte, S.; Niyogi, D.K.; Guérold, F. Effects of anthropogenic heavy metal contamination on litter decomposition in streams–A meta-analysis. Environ. Pollut. 2016, 210, 261–270. [Google Scholar] [CrossRef]
- Li, X.; Kapoor, V.; Impellitteri, C.; Chandrran, K.; Santo Domingo, J. Measuring nitrification inhibition by metals in wastewater treatment systems: Current state of science and fundamental research needs. Crit. Rev. Environ. Sci. Technol. 2016, 46, 249–289. [Google Scholar] [CrossRef]


{kind=link}
| Parameter | Reference (R) | Treated (T) | Untreated (U) |
|---|---|---|---|
| Discharge (m3/s) | 0.17 | 0.07 | 0.01 |
| pH | 7.2 | 6.6 | 3.1 |
| SO4 (mg/L) | 39 | 90 | 770 |
| Conductivity (µs/cm) | 136 | 223 | 1727 |
| Acidity (mg CaCO3/L) | 0 | 10 | 149 |
| Soluble Fe (mg/L) | 0.02–0.10 $ | 0.31–0.52 @ | 6.63–11.72 @ |
| Soluble Mn (mg/L) | 0.02–0.09 $ | 1.07–1.12 @ | 25.52–45.89 @ |
| Soluble Al (mg/L) | 0.01–0.12 $ | 0.05–0.05 @ | 5.73–13.89 @ |
| Metal Precipitate | No | Yes | Yes |
| Total P (µg/L) | 3–5 * | <2–17 # | <2–8 # |
| NO3 + NO2-N (mg/L) | 0.21–0.73 * | 0.2–0.4 # | 0.6–0.9 # |
| NH4-N (mg/L) | 0.006–0.012 * | 0.06–0.08 # | 0.25–0.58 # |
| Taxon | U-W | U-L | U-S | T-W | T-L | T-S | R-W | R-L | R-S |
|---|---|---|---|---|---|---|---|---|---|
| Sphingomonas | 0.49 | 0.06 | 0.00 | 0.09 | 0.39 | 0.00 | 0.02 | 0.83 | 0.01 |
| Escherichia | 0.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.22 | 0.00 | 0.02 | 0.00 |
| Polynucleobacter | 0.07 | 0.32 | 0.00 | 0.06 | 0.00 | 0.00 | 0.10 | 0.01 | 0.00 |
| Propionibacterium | 0.06 | 0.01 | 0.00 | 0.02 | 0.01 | 0.39 | 0.00 | 0.04 | 0.00 |
| Acidiphilium | 0.03 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Acidocella | 0.02 | 0.20 | 0.16 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ferrovum | 0.02 | 0.01 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Dehalococcoides | 0.00 | 0.00 | 0.13 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 |
| Rhodoluna | 0.00 | 0.00 | 0.02 | 0.30 | 0.01 | 0.00 | 0.15 | 0.01 | 0.00 |
| Granulicella | 0.00 | 0.15 | 0.02 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Dietzia | 0.04 | 0.00 | 0.00 | 0.00 | 0.00 | 0.14 | 0.00 | 0.01 | 0.00 |
| Gallionella | 0.00 | 0.07 | 0.27 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.02 |
| Limnohabitans | 0.00 | 0.01 | 0.00 | 0.37 | 0.02 | 0.00 | 0.39 | 0.02 | 0.00 |
| Bradyrhizobium | 0.00 | 0.01 | 0.18 | 0.02 | 0.04 | 0.02 | 0.02 | 0.00 | 0.10 |
| Methylobacterium | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.01 |
| Rhodoferax | 0.00 | 0.00 | 0.02 | 0.01 | 0.00 | 0.03 | 0.00 | 0.00 | 0.00 |
| Leptothrix | 0.00 | 0.00 | 0.00 | 0.01 | 0.02 | 0.00 | 0.01 | 0.01 | 0.06 |
| Nitrospira | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 | 0.20 |
| Aquabacterium | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.26 |
| Richness | 19 | 30 | 11 | 30 | 27 | 11 | 26 | 22 | 21 |
| Shannon Diversity | 2.02 | 2.14 | 1.99 | 1.93 | 2.51 | 1.75 | 2.14 | 0.86 | 2.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neff, A.N.; DeNicola, D.M.; Maltman, C. Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats. Water 2021, 13, 3300. https://doi.org/10.3390/w13223300
Neff AN, DeNicola DM, Maltman C. Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats. Water. 2021; 13(22):3300. https://doi.org/10.3390/w13223300
Chicago/Turabian StyleNeff, Alexis N., Dean M. DeNicola, and Chris Maltman. 2021. "Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats" Water 13, no. 22: 3300. https://doi.org/10.3390/w13223300
APA StyleNeff, A. N., DeNicola, D. M., & Maltman, C. (2021). Passive Treatment for Acid Mine Drainage Partially Restores Microbial Community Structure in Different Stream Habitats. Water, 13(22), 3300. https://doi.org/10.3390/w13223300