Cometabolism of the Superphylum Patescibacteria with Anammox Bacteria in a Long-Term Freshwater Anammox Column Reactor

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Reactor Operation and Sampling
2.2. DNA Extraction and DNA Sequencing
2.3. Bioinformatics
3. Results and Discussion
3.1. Anammox Reactor Operation
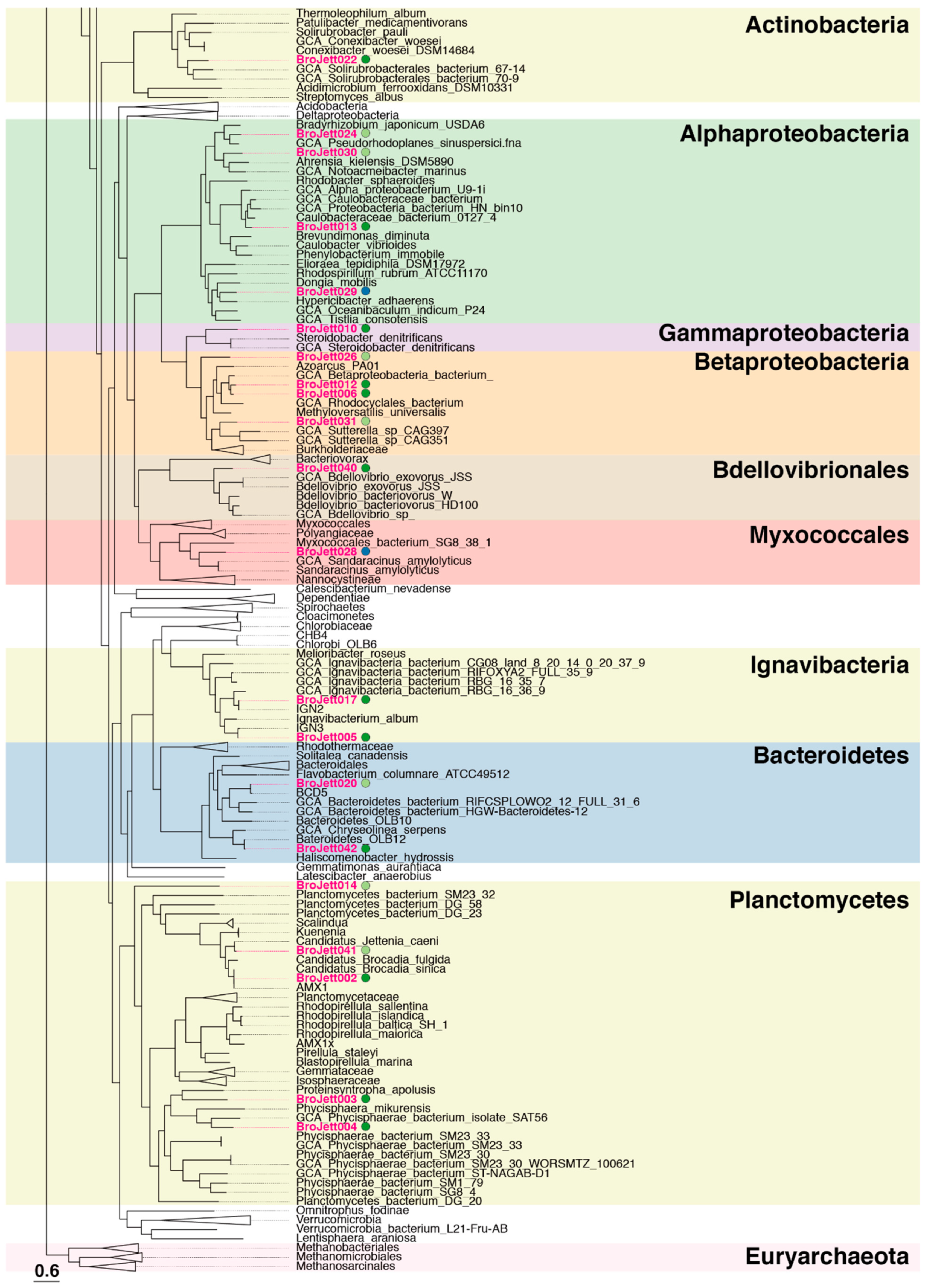
3.2. Genome Construction and Basic Information on Bins
3.3. Relative Abundance
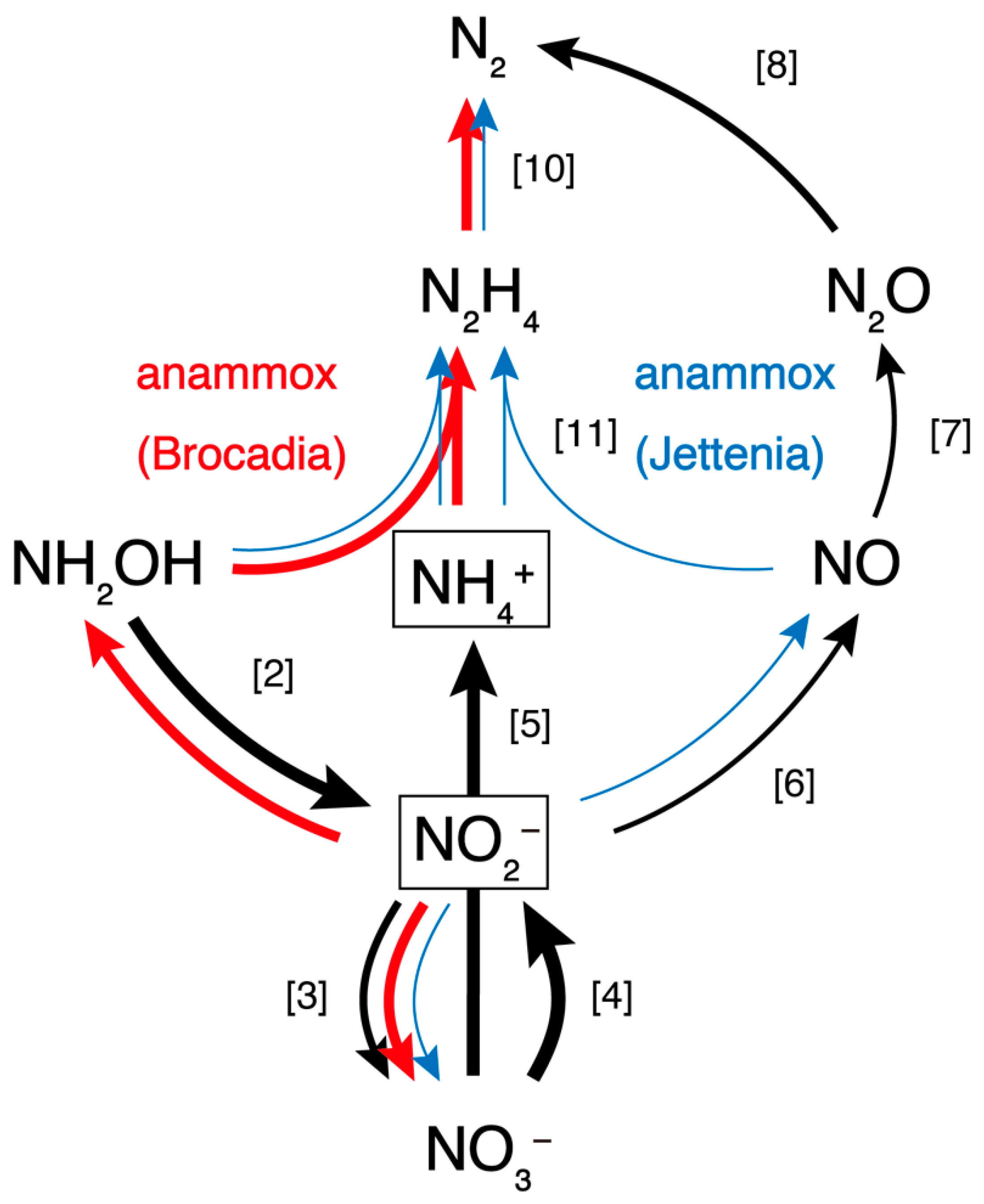
3.4. Nitrogen Cycle
3.5. Genomic Features of Patescibacteria Bins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Strous, M.; Fuerst, J.A.; Kramer, E.H.M.; Logemann, S.; Muyzer, G.; Van De Pas-Schoonen, K.T.; Webb, R.; Kuenen, J.G.; Jetten, M.S.M. Missing lithotroph identified as new planctomycete. Nature 1999, 400, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Khramenkov, S.V.; Kozlov, M.N.; Kevbrina, M.V.; Dorofeev, A.G.; Kazakova, E.A.; Grachev, V.A.; Kuznetsov, B.B.; Polyakov, D.Y.; Nikolaev, Y.A. A novel bacterium carrying out anaerobic ammonium oxidation in a reactor for biological treatment of the filtrate of wastewater fermented sludge. Microbiology 2013, 82, 628–636. [Google Scholar] [CrossRef]
- Oshiki, M.; Satoh, H.; Okabe, S. Ecology and physiology of anaerobic ammonium-oxidizing bacteria. Environ. Microbiol. 2016, 18, 2784–2796. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Oshiki, M.; Awata, T.; Isobe, K.; Kimura, Z.; Yoshikawa, H.; Hira, D.; Kindaichi, T.; Satoh, H.; Fujii, T.; et al. Physiological characterization of anaerobic ammonium oxidizing bacterium “Candidatus Jettenia caeni”. Environ. Microbiol. 2015, 17, 2172–2189. [Google Scholar] [CrossRef]
- Awata, T.; Oshiki, M.; Kindaichi, T.; Ozaki, N.; Ohashi, A.; Okabe, S. Physiological characterization of an anaerobic ammonium-oxidizing bacterium belonging to the “Candidatus Scalindua”’ group. Appl. Environ. Microbiol. 2013, 79, 4145–4148. [Google Scholar] [CrossRef]
- Oshiki, M.; Shimokawa, M.; Fujii, N.; Satoh, H.; Okabe, S. Physiological characteristics of the anaerobic ammonium-oxidizing bacterium “Candidatus Brocadia sinica”. Microbiology 2011, 157, 1706–1713. [Google Scholar] [CrossRef]
- Strous, M.; Heijnen, J.J.; Kuenen, J.G.; Jetten, M.S.M. The sequencing batch reactor is a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms. Appl. Microbiol. Biotechnol. 1998, 50, 589–596. [Google Scholar] [CrossRef]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef]
- Albertsen, M.; Hugenholtz, P.; Skarshewski, A.; Nielsen, K.L.; Tyson, G.W.; Nielsen, P.H. Genome sequences of rare, uncultured bacteria obtained by differential coverage binning of multiple metagenomes. Nat. Biotechnol. 2013, 31, 533–538. [Google Scholar] [CrossRef]
- Brown, C.T.; Hug, L.A.; Thomas, B.C.; Sharon, I.; Castelle, C.J.; Singh, A.; Wilkins, M.J.; Wrighton, K.C.; Williams, K.H.; Banfield, J.F. Unusual biology across a group comprising more than 15% of domain Bacteria. Nature 2015, 523, 208–211. [Google Scholar] [CrossRef]
- Kindaichi, T.; Yamaoka, S.; Uehara, R.; Ozaki, N.; Ohashi, A.; Albertsen, M.; Nielsen, P.H.; Nielsen, J.L. Phylogenetic diversity and ecophysiology of Candidate phylum Saccharibacteria in activated sludge. FEMS Microbiol. Ecol. 2016, 92, fiw078. [Google Scholar] [CrossRef] [PubMed]
- Speth, D.R.; In’t Zandt, M.H.; Guerrezo-Cruz, S.; Dutilh, B.E.; Jettenm, M.S.M. Genome-based microbial ecology of anammox granules in a full-scale wastewater treatment system. Nat. Commun. 2016, 7, 11172. [Google Scholar] [CrossRef] [PubMed]
- Kindaichi, T.; Awata, T.; Suzuki, Y.; Tanabe, K.; Hatamoto, M.; Ozaki, N.; Ohashi, A. Enrichment using an up-flow column reactor and community structure of marine anammox bacteria from coastal sediment. Microbes Environ. 2011, 26, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Tsushima, I.; Ogasawara, Y.; Kindaichi, T.; Satoh, H.; Okabe, S. Development of high-rate anaerobic ammonium-oxidizing (anammox) biofilm reactors. Water Res. 2007, 41, 1623–1634. [Google Scholar] [CrossRef]
- Van de Graaf, A.A.; De Bruijn, P.; Robertson, L.A.; Jetten, M.S.M.; Kuenen, J.G. Autotrophic growth of anaerobic ammonium-oxidizing microorganisms in a fluidized bed reactor. Microbiology 1996, 142, 2187–2196. [Google Scholar] [CrossRef]
- Awata, T.; Goto, Y.; Kindaichi, T.; Ozaki, N.; Ohashi, A. Nitrogen removal using an anammox membrane bioreactor at low temperature. Water Sci. Technol. 2015, 72, 2148–2153. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Wu, Y.-W.; Simmons, B.A.; Singer, S.W. MaxBin 2.0: An automated binning algorithm to recover genomes from multiple metagenomic datasets. Bioinformatics 2016, 32, 605–607. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing of the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Graham, E.D.; Heidelberg, J.F.; Tully, B.J. Potential for primary productivity in a globally-distributed bacterial phototroph. ISME J. 2018, 12, 1861–1866. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Asnicar, F.; Thomas, A.M.; Beghini, F.; Mengoni, C.; Manara, S.; Manghi, P.; Zhu, Q.; Bolzan, M.; Cumbo, F.; May, U.; et al. Precise phylogenetic analysis of microbial isolates and genomes from metagenomes using PhyloPhlAn 3.0. Nat. Commun. 2020, 11, 2500. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Shaw, D.R.; Albertsen, M.; Saikaly, P.E. Comparative genome-centric analysis of freshwater and marine anammox cultures suggests functional redundancy in nitrogen removal processes. Front. Microbiol. 2020, 11, 1637. [Google Scholar] [CrossRef]
- Oshiki, M.; Ali, M.; Shinyako-Hata, K.; Satoh, H.; Okabe, S. Hydroxylamine-dependent anaerobic ammonium oxidation (anammox) by “Candidatus Brocadia sinica”. Environ. Microbiol. 2016, 18, 3133–3143. [Google Scholar] [CrossRef]
- Lau, K.W.K.; Ng, C.Y.M.; Ren, J.; Lau, S.C.L.; Qian, P.Y.; Wong, P.K.; Lau, T.C.; Wu, M. Owenweeksia Hongkongensis gen. nov., sp. nov., a novel marine bacterium of the phylum ‘Bacteroidetes’. Int. J. Syst. Evol. Microbiol. 2005, 55, 1051–1057. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, J.H.; Liu, Y.H.; Zhou, Y.G.; Liu, Z.P. Dongia mobilis gen. nov., sp. nov., a new member of the family Rhodospirillaceae isolated from a sequencing batch reactor for treatment of malachite green effluent. Int. J. Syst. Evol. Microbiol. 2010, 60, 2780–2785. [Google Scholar] [CrossRef]
- Lai, Q.; Yuan, J.; Wu, C.; Shao, Z. Oceanibaculum indicum gen. nov., sp. nov., isolated from deep seawater of the Indian Ocean. Int. J. Syst. Evol. Microbiol. 2009, 59, 1733–1737. [Google Scholar] [CrossRef]
- Kartal, B.; De Almeida, N.M.; Maalcke, W.J.; Op den Camp, H.J.M.; Jetten, M.S.M.; Keltjens, J.T. How to make a living from anaerobic ammonium oxidation. FEMS Microbiol. Rev. 2013, 37, 428–461. [Google Scholar] [CrossRef] [PubMed]
- Lemos, L.N.; Medeiros, J.D.; Dini-Andreote, F.; Fernandes, G.R.; Varani, A.M.; Oliveira, G.; Pylro, V.S. Genomic signatures and co-occurrence patterns of the ultra-small Saccharimonadia (phylum CPR/Patescibacteria) suggest a symbiotic lifestyle. Mol. Ecol. 2019, 28, 4259–4271. [Google Scholar] [CrossRef] [PubMed]
- Beam, J.P.; Becraft, E.D.; Brown, J.M.; Schulz, F.; Jarett, J.K.; Bezuidt, O.; Poulton, N.J.; Clark, K.; Dunfield, P.F.; Ravin, N.V.; et al. Ancestral absence of electron transport chains in Patescibacteria and DPANN. Front. Microbiol. 2020, 11, 1848. [Google Scholar] [CrossRef] [PubMed]
- Chung, F.Z.; Tsujibo, H.; Bhattacharyya, U.; Sharief, F.S.; Li, S.S. Genomic organization of human lactate dehydrogenase-A gene. Biochem. J. 1985, 231, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Overkamp, K.M.; Kötter, P.; van der Hoek, R.; Schoondermark-Stolk, S.; Luttik, M.A.H.; van Dijken, J.P.; Pronk, J.T. Functional analysis of structural genes for NAD+-dependent formate dehydrogenase in Saccharomyces cerevisiae. Yeast 2002, 19, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Fukui, T.; Fujiwara, S.; Atomi, H.; Imanaka, T. Concerted action of diacetylchitobiose deacetylase and exo-β-D-glucosaminidase in a novel chitinolytic pathway in the hyperthermophilic archaeon Thermococcus kodakaraensis KOD1. J. Biol. Chem. 2004, 279, 30021–30027. [Google Scholar] [CrossRef]
- Tharanathan, R.N.; Kittur, F.S. Chitin–the undisputed biomolecule of great potential. Crit. Rev. Food Sci. Nutr. 2003, 43, 61–87. [Google Scholar] [CrossRef]
- Takeo, M.; Kimura, K.; Mayilraj, S.; Inoue, T.; Tada, S.; Miyamoto, K.; Kashiwa, M.; Ikemoto, K.; Baranwal, P.; Kato, D.; et al. Biosynthetic pathway and genes of chitin/chitosan-like bioflocculant in the genus Citrobacter. Polymers 2018, 10, 237. [Google Scholar] [CrossRef]
- Itoh, Y.; Rice, J.D.; Goller, C.; Pannuri, A.; Taylor, J.; Meisner, J.; Beveridge, T.J.; Preston, J.F.; Romeo, T. Roles of pgaABCD genes in synthesis, modification, and export of the Escherichia coli biofilm adhesin poly-beta-1,6-N-acetyl-D-glucosamine. J. Bacteriol. 2008, 190, 3670–3680. [Google Scholar] [CrossRef]
- Kindaichi, T.; Ito, T.; Okabe, S. Ecophysiological interaction between nitrifying bacteria and heterotrophic bacteria in autotrophic nitrifying biofilms as determined by microautoradiography-fluorescence in situ Hybridization. Appl. Environ. Microbiol. 2004, 70, 1641–1650. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bin ID | Taxonomy | Complete-ness | Contamination | Bin Size (Mbp) | Number of Contigs | Relative Abundance (%) | ||
|---|---|---|---|---|---|---|---|---|
| Day 4989 | Day 5054 | Day 5073 | ||||||
| BroJett025 | Patescibacteria, Candidatus Pacebacteria | 97.67 * | 0 * | 1.17 | 1 | 0.2 | 0.3 | 0.2 |
| BroJett032 | Patescibacteria, Candidatus Pacebacteria | 100 * | 0 * | 1.18 | 5 | 0.1 | 0.2 | 0.1 |
| BroJett037 | Patescibacteria, Candidate division WS6 | 95.35 * | 0 * | 1.18 | 3 | 0.1 | 0.1 | 0.1 |
| BroJett019 | Patescibacteria, Candidate division WS6 | 95.35 * | 0* | 1.06 | 5 | 0.3 | 0.5 | 0.5 |
| BroJett008 | Patescibacteria | 97.67* | 2.33 * | 0.57 | 8 | 2.0 | 0.1 | 0.0 |
| BroJett034 | Patescibacteria, Berkelbacteria | 93.02 * | 0 * | 0.68 | 2 | 0.1 | 0.1 | 0.2 |
| BroJett039 | Chloroflexi | 23.2 | 0 | 1.71 | 48 | 0.1 | 0.0 | 0.1 |
| BroJett021 | Chloroflexi | 90.91 | 0.91 | 6.39 | 43 | 0.3 | 0.2 | 0.1 |
| BroJett001 | Chloroflexi | 93.64 | 3.09 | 3.71 | 153 | 23.8 | 8.1 | 7.1 |
| BroJett015 | Chloroflexi | 75.64 | 3.82 | 5.45 | 63 | 0.6 | 0.7 | 0.2 |
| BroJett018 | Chloroflexi | 91.82 | 0.91 | 6.27 | 45 | 0.4 | 0.4 | 0.1 |
| BroJett007 | Chloroflexi | 98.18 | 0 | 4.26 | 35 | 5.3 | 2.0 | 1.5 |
| BroJett038 | Chloroflexi | 77.27 | 0.91 | 4.12 | 49 | 0.1 | 0.1 | 0.1 |
| BroJett033 | Chloroflexi | 28.8 | 0 | 1.12 | 41 | 0.1 | 0.0 | 0.0 |
| BroJett011 | Chloroflexi | 84.85 | 3.64 | 9.27 | 84 | 1.1 | 0.4 | 0.2 |
| BroJett009 | Armatimonadetes | 91.76 | 0 | 2.69 | 23 | 1.4 | 0.4 | 0.4 |
| BroJett022 | Actinobacteria | 96.98 | 0 | 2.70 | 5 | 0.3 | 0.2 | 0.0 |
| BroJett024 | Alphaproteobacteria | 87.29 | 6.96 | 4.67 | 34 | 0.2 | 0.1 | 0.0 |
| BroJett030 | Alphaproteobacteria | 87.95 | 1.2 | 3.49 | 37 | 0.1 | 0.1 | 0.0 |
| BroJett013 | Alphaproteobacteria | 95.02 | 0.6 | 3.78 | 19 | 0.9 | 0.3 | 0.0 |
| BroJett029 | Alphaproteobacteria | 75.86 | 6.03 | 4.62 | 51 | 0.2 | 0.1 | 0.0 |
| BroJett010 | Gammaproteobacteria | 90.31 | 1.5 | 2.94 | 10 | 1.3 | 0.5 | 0.5 |
| BroJett026 | Betaproteobacteria | 87.56 | 2.68 | 4.49 | 28 | 0.2 | 0.1 | 0.0 |
| BroJett012 | Betaproteobacteria | 91.11 | 0.52 | 3.22 | 101 | 1.0 | 2.6 | 5.5 |
| BroJett006 | Betaproteobacteria | 95.56 | 0.45 | 3.30 | 135 | 5.8 | 17.7 | 10.7 |
| BroJett031 | Betaproteobacteria | 88.27 | 0.62 | 4.03 | 13 | 0.1 | 0.1 | 0.0 |
| BroJett040 | Oligoflexia | 92.86 | 0 | 2.76 | 21 | 0.0 | 0.1 | 0.1 |
| BroJett028 | Deltaproteobacteria | 75.91 | 5.38 | 8.63 | 98 | 0.2 | 0.2 | 0.0 |
| BroJett017 | Ignavibacteriae | 94.97 | 0 | 3.60 | 22 | 0.5 | 0.1 | 0.1 |
| BroJett005 | Ignavibacteriae | 96.65 | 0.56 | 3.85 | 120 | 9.9 | 3.7 | 3.2 |
| BroJett020 | Bacteroidetes | 89.25 | 0 | 2.70 | 21 | 0.3 | 0.2 | 0.5 |
| BroJett042 | Bacteroidetes | 93.99 | 0 | 3.74 | 27 | 0.0 | 0.0 | 0.2 |
| BroJett014 | Planctomycetes | 81.82 | 4.55 | 4.02 | 32 | 0.9 | 1.0 | 0.6 |
| BroJett041 | Planctomycetes (Jettenia) | 84.62 | 1.1 | 3.02 | 23 | 0.0 | 0.2 | 0.1 |
| BroJett002 | Planctomycetes (Brocadia) | 97.8 | 1.65 | 4.10 | 23 | 14.6 | 39.0 | 58.5 |
| BroJett003 | Planctomycetes | 95.01 | 2.94 | 3.92 | 437 | 13.1 | 18.0 | 7.6 |
| BroJett004 | Planctomycetes | 97.66 | 0 | 3.22 | 20 | 13.0 | 1.8 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosokawa, S.; Kuroda, K.; Narihiro, T.; Aoi, Y.; Ozaki, N.; Ohashi, A.; Kindaichi, T. Cometabolism of the Superphylum Patescibacteria with Anammox Bacteria in a Long-Term Freshwater Anammox Column Reactor. Water 2021, 13, 208. https://doi.org/10.3390/w13020208
Hosokawa S, Kuroda K, Narihiro T, Aoi Y, Ozaki N, Ohashi A, Kindaichi T. Cometabolism of the Superphylum Patescibacteria with Anammox Bacteria in a Long-Term Freshwater Anammox Column Reactor. Water. 2021; 13(2):208. https://doi.org/10.3390/w13020208
Chicago/Turabian StyleHosokawa, Suguru, Kyohei Kuroda, Takashi Narihiro, Yoshiteru Aoi, Noriatsu Ozaki, Akiyoshi Ohashi, and Tomonori Kindaichi. 2021. "Cometabolism of the Superphylum Patescibacteria with Anammox Bacteria in a Long-Term Freshwater Anammox Column Reactor" Water 13, no. 2: 208. https://doi.org/10.3390/w13020208
APA StyleHosokawa, S., Kuroda, K., Narihiro, T., Aoi, Y., Ozaki, N., Ohashi, A., & Kindaichi, T. (2021). Cometabolism of the Superphylum Patescibacteria with Anammox Bacteria in a Long-Term Freshwater Anammox Column Reactor. Water, 13(2), 208. https://doi.org/10.3390/w13020208