Distribution of Charophyte Oospores in the Curonian Lagoon and their Relationship to Environmental Forcing

Abstract
1. Introduction
2. Study Area
3. Materials and Methods
3.1. Field Sampling
3.2. Data Analysis
3.3. Statistical Analysis
4. Results
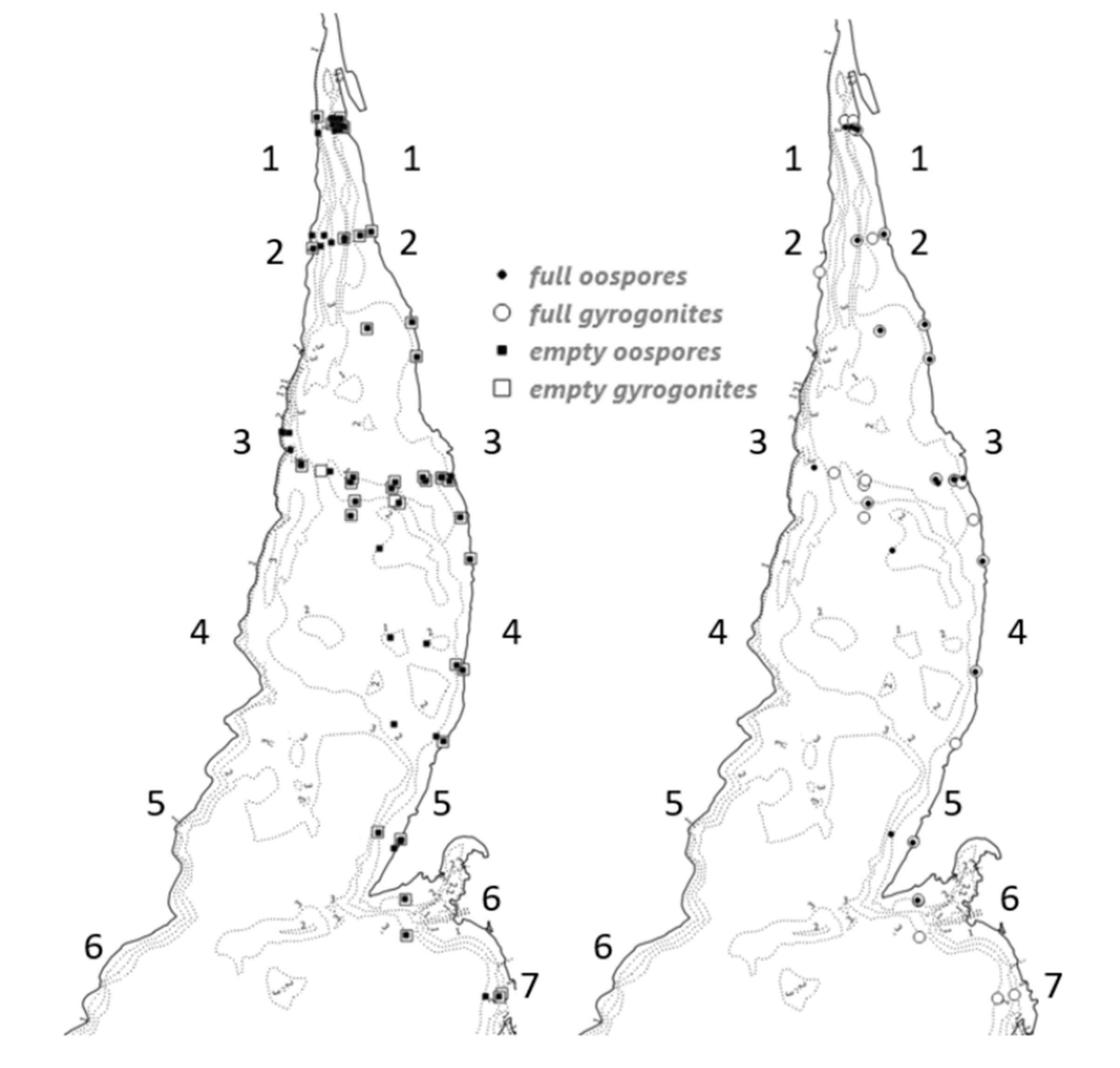
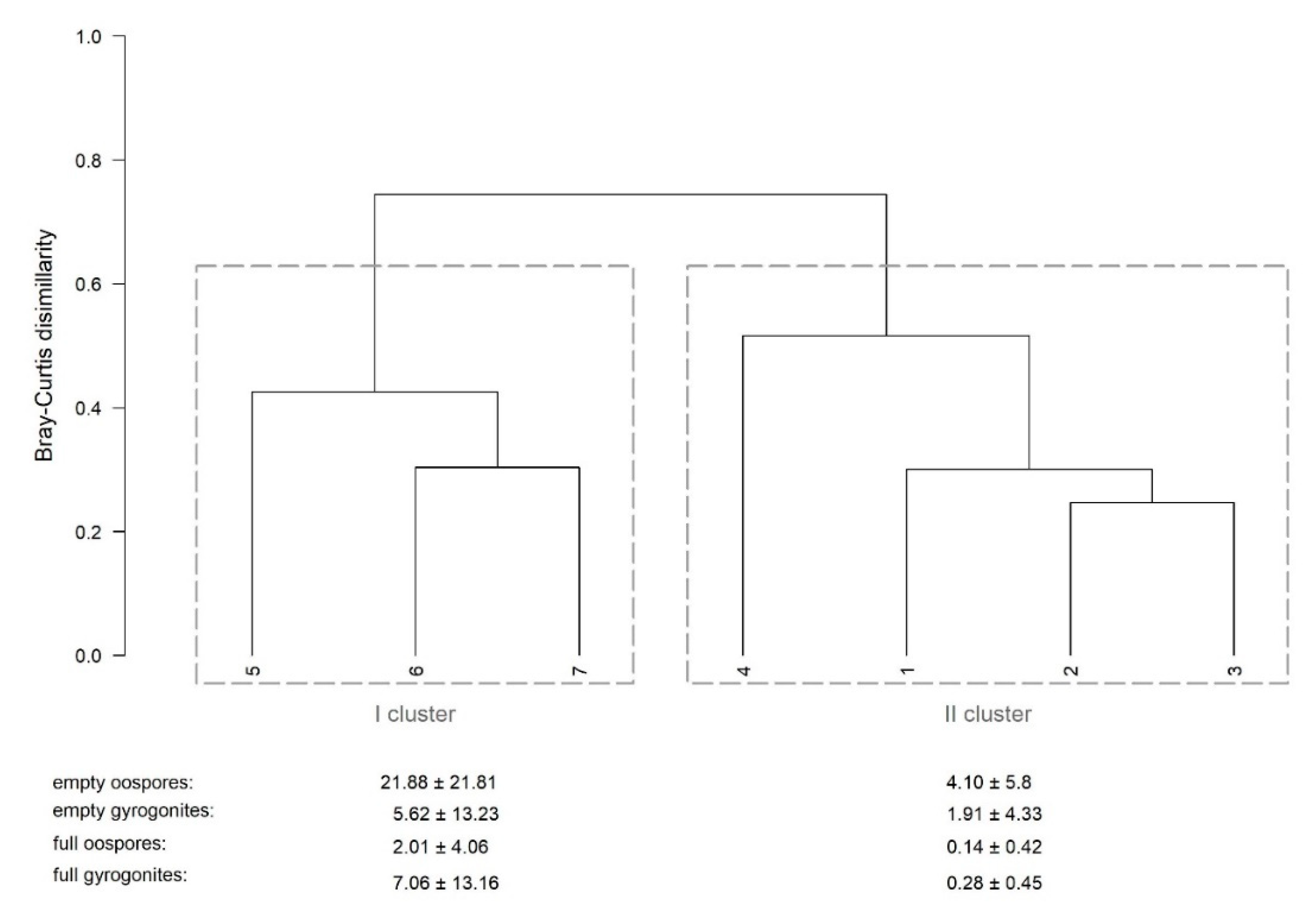
4.1. Structure and Distribution of Oospore Bank
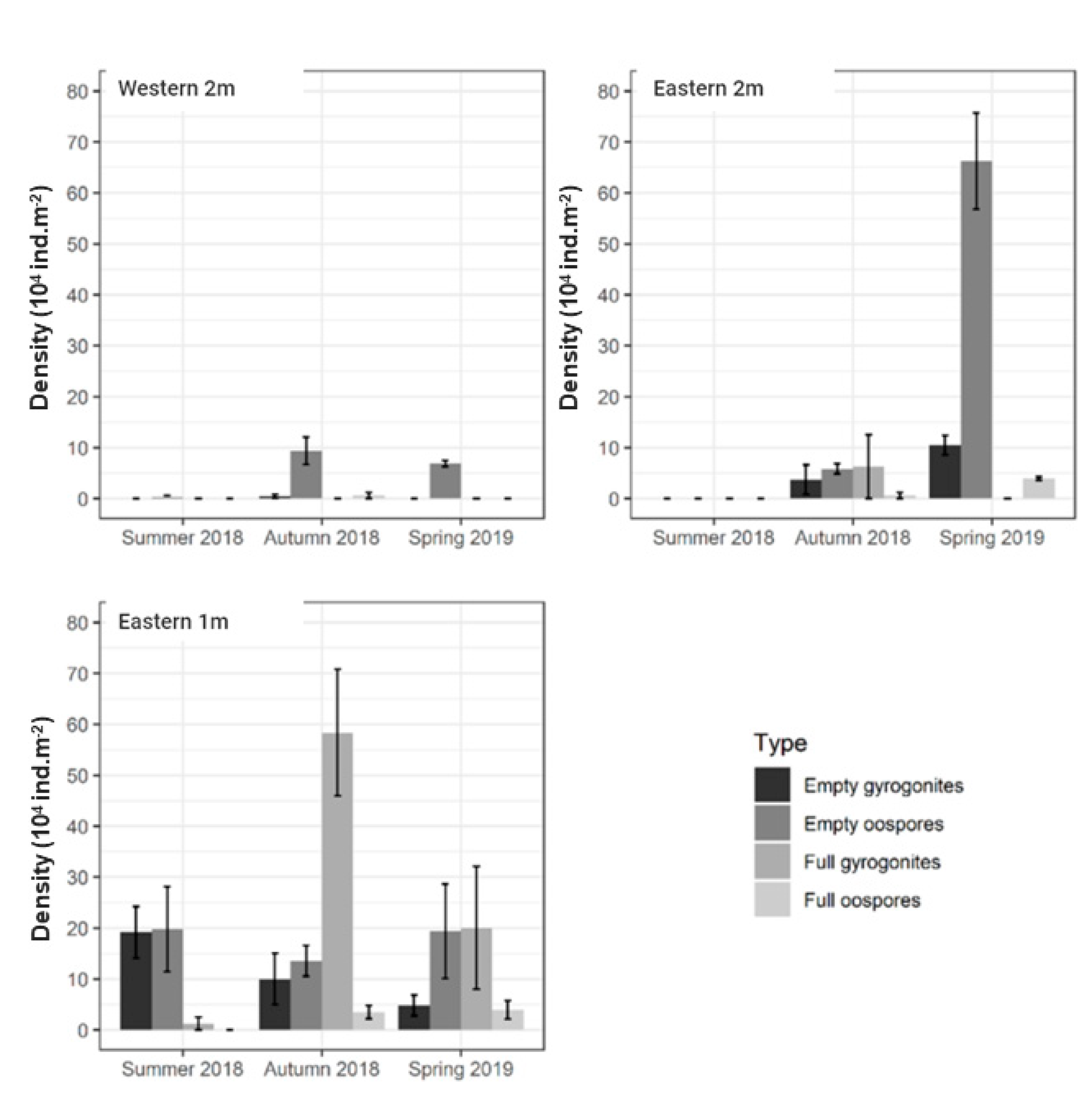
4.2. Seasonal Patterns
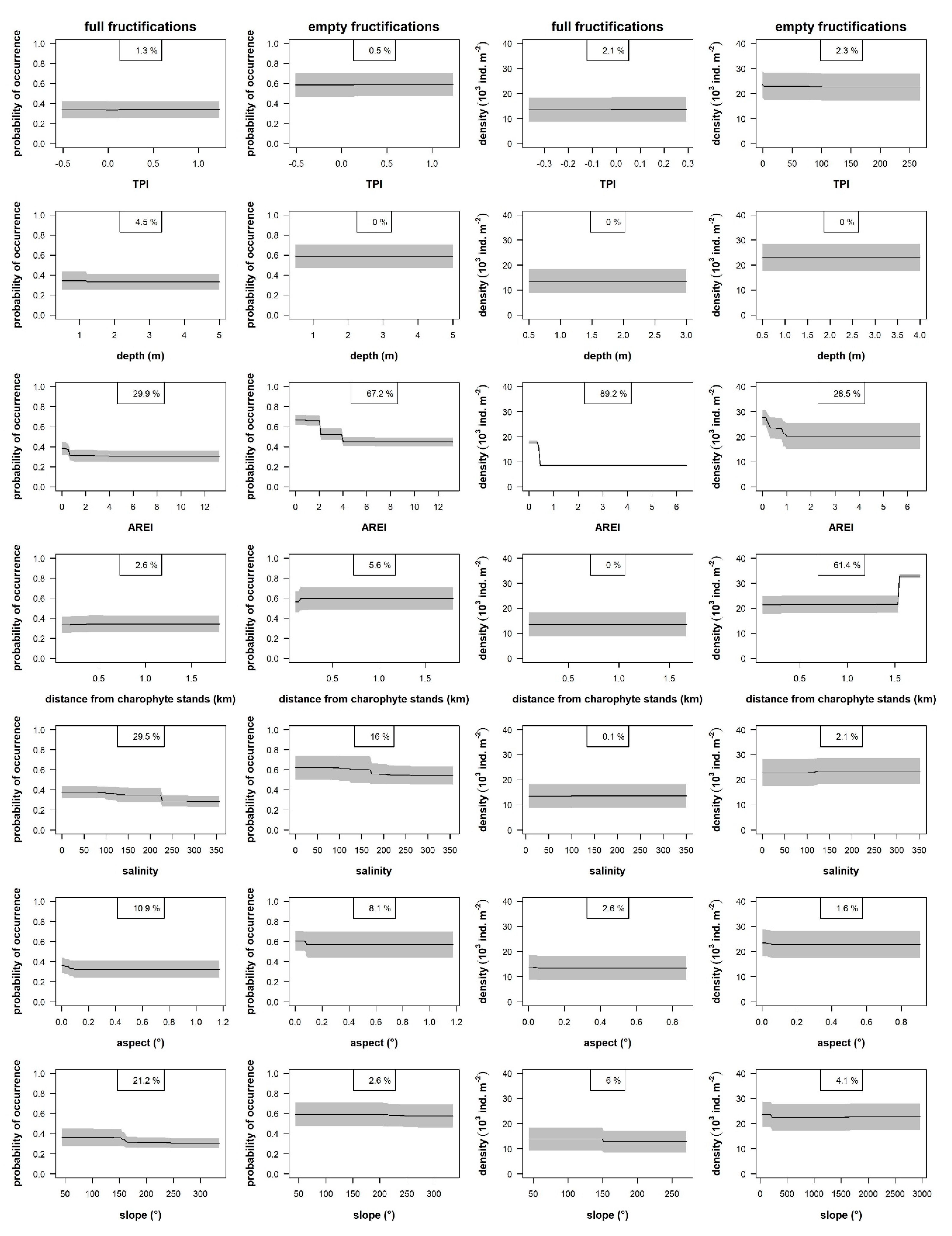
4.3. Environmental Factors Explaining Patterns in Oospore Bank
5. Discussion
5.1. Species Composition
5.2. Density and Distribution
5.3. Importance of Environmental Factors
5.3.1. Distance from Charophyte Stands
5.3.2. Hydrodynamics and Transport of Fructification
5.3.3. Salinity
5.3.4. Depth
5.3.5. Topography of the Seafloor (Proxy of Intensity of Water Movement)
5.3.6. Waterfowl
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonis, A.; Grillas, P. Deposition, germination and spatio-temporal patterns of charophyte propagule banks: A review. Aquat. Bot. 2002, 72, 235–248. [Google Scholar] [CrossRef]
- Stobbe, A.; Gregor, T.; Röpke, A. Long-lived banks of oospores in lake sediments from the Trans-Urals (Russia) indicated by germination in over 300 years old radiocarbondated sediments. Aquat. Bot. 2014, 119, 84–90. [Google Scholar] [CrossRef]
- Steinhardt, T.; Selig, U. Comparison of recent vegetation and diaspore banks along abiotic gradients in brackish coastal lagoons. Aquat. Bot. 2009, 91, 20–26. [Google Scholar] [CrossRef]
- Blindow, I.; Dahlke, S.; Dewart, A.; Flügge, S.; Hendreschke, M.; Kerkow, A.; Meyer, J. Long-term and interannual changes of submerged macrophytes and their associated diaspore reservoir in a shallow southern Baltic Sea bay: Influence of eutrophication and climate. Hydrobiologia 2016, 778, 121–136. [Google Scholar] [CrossRef]
- Nowak, P.; Steinhardt, T.; von Ammon, U.; Rohde, H.; Schoor, A.; Holzhausen, A.; Schaible, R.; Schubert, H. Diaspore bank analysis of Baltic coastal waters. Bot. Lett. 2018, 165, 159–173. [Google Scholar] [CrossRef]
- Blindow, I. Distribution of charophytes along the Swedish coast in relation to salinity and eutrophication. Int. Rev. Hydrobiol. 2000, 85, 707–717. [Google Scholar] [CrossRef]
- Brzeska, P.; Woźniczka, A.; Pełechaty, M.; Blindow, I. New records of Chara connivens P. Salzmann ex A. Braun1835—An extremely rare and protected species in Polish brackish waters. Acta Soc. Bot. Pol. 2015, 84, 143–146. [Google Scholar] [CrossRef][Green Version]
- Torn, K.; Kovtun-Kante, A.; Herkül, K.; Martin, G.; Mäemets, H. Distribution and predictive occurrence model of charophytes in Estonian waters. Aquat. Bot. 2015, 120, 142–149. [Google Scholar] [CrossRef]
- Kovtun, A.; Torn, K.; Martin, G.; Kullas, T.; Kotta, J.; Suursaar, Ü. Influence of abiotic environmental conditions on spatial distribution of charophytes in the coastal waters of West Estonian Archipelago, Baltic Sea. J. Coast. Res. 2011, 412–416. [Google Scholar] [CrossRef]
- Steinhardt, T.; Selig, U. Spatial distribution patterns and relationship between recent vegetation and diaspore bank of a brackish coastal lagoon on the southern Baltic Sea. Estuar. Coast. Shelf Sci. 2007, 74, 205–214. [Google Scholar] [CrossRef]
- Van den Berg, M.S.; Coops, H.; Simons, J. Propagule bank buildup of Chara aspera and its significance for colonization of a shallow lake. Hydrobiologia 2001, 462, 9–17. [Google Scholar] [CrossRef]
- Birk, S.; Bonne, W.; Borja, A.; Brucet, S.; Courrat, A.; Poikane, S.; Solimini, A.; Van De Bund, W.; Zampoukas, N.; Hering, D. Three hundred ways to assess Europe’s surface waters: An almost complete overview of biological methods to implement the Water Framework Directive. Ecol. Indic. 2012, 18, 31–41. [Google Scholar] [CrossRef]
- Sinkevičienė, Z.; Bučas, M.; Ilginė, R.; Vaičiūtė, D.; Kataržytė, M.; Petkuvienė, J. Charophytes in the estuarine Curonian Lagoon: Have the changes in diversity, abundance and distribution occurred since the late 1940s? Oceanol. Hydrobiol. Stud. 2017, 46, 186. [Google Scholar] [CrossRef]
- Bučas, M.; Sinkevičienė, Z.; Kataržytė, M.; Vaičiūtė, D.; Petkuvienė, J.; Stragauskaitė, V.; Ilginė, R. How much can the occurrence and coverage of charophytes in an estuarine lagoon (Curonian Lagoon) be explained by environmental factors? Estuar. Coast. Shelf Sci. 2019, 216, 128–138. [Google Scholar] [CrossRef]
- Christian, F.; Arturas, R.; Saulius, G.; Georg, U.; Lina, B. Hydraulic regime-based zonation scheme of the Curonian Lagoon. Hydrobiologia 2008, 611, 133–146. [Google Scholar] [CrossRef]
- Dailidienė, I.; Davulienė, L. Long-term mean salinity in the Curonian Lagoon in 1993–2005. Acta Zool Lit. 2007, 17, 172–181. [Google Scholar] [CrossRef]
- Zemlys, P.; Ferrarin, C.; Umgiesser, G.; Gulbinskas, S.; Bellafiore, D. Investigation of saline water intrusions into the Curonian Lagoon (Lithuania) and two-layer flow in the Klaipėda Strait using finite element hydrodynamic model. Ocean Sci. 2013, 9. [Google Scholar] [CrossRef]
- Lesutiene, J.; Bukaveckas, P.A.; Gasiunaite, Z.R.; Pilkaityte, R.; Razinkovas-Baziukas, A. Tracing the isotopic signal of a cyanobacteria bloom through the food web of a Baltic Sea coastal lagoon. Estuar. Coast. Shelf Sci. 2014, 138, 47–56. [Google Scholar] [CrossRef]
- Bresciani, M.; Adamo, M.; De Carolis, G.; Matta, E.; Pasquariello, G.; Vaičiūtė, D.; Giardino, C. Monitoring blooms and surface accumulation of cyanobacteria in the Curonian Lagoon by combining MERIS and ASAR data. Remote Sens. Environ. 2014, 146, 124–135. [Google Scholar] [CrossRef]
- Vaičiūtė, D.; Bresciani, M.; Bartoli, M.; Giardino, C.; Bučas, M. Spatial and temporal distribution of coloured dissolved organic matter in a hypertrophic freshwater lagoon. J. Limnol. 2015, 74, 572–583. [Google Scholar] [CrossRef][Green Version]
- Trimonis, E.; Gulbinskas, S.; Kuzavinis, M. The Curonian Lagoon bottom sediments in the Lithuanian water area. Baltica 2003, 16, 13–20. [Google Scholar]
- Krause, W. Zur Bestimmungsmöglichkeit subfossiler Characeen-Oosporen an Beispielen aus Schweizer Seen. Vierteljahrsschr. Naturforsch. Ges. Zürich. 1986, 141, 295–313. [Google Scholar]
- Haas, J.N. First identification key for charophyte oospores from central Europe. Pflugers. Arch. 1994, 29, 227–235. [Google Scholar] [CrossRef]
- Vedder, F. Morphologie und Taxonomie rezenter und subfossiler Characeen-Oosporen aus der Ostsee. Rostock. Meeresbiolog. Beitr. 2004, 13, 43–54. [Google Scholar]
- Soulié-Märsche, I.; García, A. Gyrogonites and oospores, complementary viewpoints to improve the study of the charophytes (Charales). Aquat. Bot. 2015, 120, 7–17. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. v 2.18.12- Las Palmas. Open Source Geospatial Found. Proj. 2020. Available online: https://doi.org/http://www.qgis.org/ (accessed on 18 April 2020).
- Malhotra, A.; Fonseca, M.S. WEMo (Wave Exposure Model): Formulation, Procedures and Validation; NOAA Technical Memorandum NOS NCCOS 65: Beaufort, NC, USA, 2007. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 June 2020).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling; R Package Version 3.3-13; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://CRAN.R-project.org/package=raster (accessed on 20 June 2020).
- Bivand, R.; Keitt, T.; Rowlingson, B. rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library; R Package Version 1.5-17; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://CRAN.R-project.org/package=rgdal (accessed on 20 June 2020).
- Hijmans, R.J.; Williams, E.; Vennes, C. Geosphere: Spherical Trigonometry; R Package Version 1.3-11; R Package Version 1.5-17; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: https://cran.r-project.org/web/packages/geosphere (accessed on 20 June 2020).
- Bivand, R.; Rundel, C. Rgeos: Interface to Geometry Engine—Open Source (GEOS); R Package Version 0.3-11; R Package Version 1.5-17; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://cran.r-project.org/web/packages/rgeos/i (accessed on 20 June 2020).
- Evans, J.S. SpatialEco; R Package Version 1.3-1; R Package Version 1.5-17; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://github.com/jeffreyevans/spatialEco (accessed on 20 June 2020).
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Ordination—First encounter. Analysing Ecological Data. 2007, 189–192. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.4-3; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://cran.r-project.org/web/packages/vegan (accessed on 20 June 2020).
- Peters, G. Userfriendlyscience: Quantitative Analysis made Accessible; R Package Version 0.7.0; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://cran.r-project.org/web/packages/userfriendlyscience (accessed on 20 June 2020).
- Friedman, J.H. Greedy function approximation: A gradient boosting machine. Ann. Stat. 2001, 1189–1232. [Google Scholar] [CrossRef]
- Torn, K.; Peterson, A.; Herkül, K.; Suursaar, U. Effects of climate change on the occurrence of charophytes and angiosperms in a brackish environment. Webbia. 2019, 74, 167–177. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- LeDell, E.; Gill, N.; Aiello, S.; Fu, A.; Candel, A.; Click, C.; Kraljevic, T.; Nykodym, T.; Aboyoun, P.; Kurka, M.; et al. h2o: R Interface for the ‘H2O’ Scalable Machine Learning Platform; R Package Version 3.30.1.3; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://cran.r-project.org/web/packages/h2o/h2o.pdf (accessed on 20 June 2020).
- Schubert, H.; Blindow, I. Charophyte of the Baltic Sea; The Baltic Marine Biologists Publication, No. 19; A.R.G. Gantner Verlag Kommanditgesellschaft: Ruggell, Liechtenstein, 2003; pp. 27–36. [Google Scholar]
- Santamaría, L. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment. Acta oecol. 2002, 23, 137–154. [Google Scholar] [CrossRef]
- Doege, A.K.; van de Weyer, K.; Becker, R.; Schubert, H. Bioindikation mit Characeen [Bioindication with Characeae]. In Armleuchteralgen. Die Characeen Deutschlands [Stoneworts. Characeae of Germany]; Chara, A.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 8, pp. 97–138. [Google Scholar]
- Steinhardt, T.; Selig, U. Influence of salinity and sediment resuspension on macrophyte germination in coastal lakes. J. Limnol. 2011, 70, 11–20. [Google Scholar] [CrossRef]
- Casanova, M.T.; Brock, M.A. Life histories of charophytes from permanent and temporary wetlands in eastern Australia. Aust. J. Bot. 1999, 47, 383–397. [Google Scholar] [CrossRef]
- De Winton, M.D.; Casanova, M.T.; Clayton, J.S. Charophyte germination and establishment under low irradiance. Aquat. Bot. 2004, 79, 175–187. [Google Scholar] [CrossRef]
- Sanjuan, J.; Vicente, A.; Flor-Arnau, N.; Monleón, T.; Cambra, J.; Martín-Closas, C. Effects of light and temperature on Chara vulgaris (Charophyceae) gyrogonite productivity and polymorphism–palaeoenvironmental implications. Phycologia 2017, 56, 204–212. [Google Scholar] [CrossRef]
- Elliott, M.; Quintino, V. The estuarine quality paradox, environmental homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Pollut. Bull. 2007, 54, 640–645. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol.System. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Bertrin, V.; Boutry, S.; Alard, D.; Haury, J.; Jan, G.; Moreira, S.; Ribaudo, C. Prediction of macrophyte distribution: The role of natural versus anthropogenic physical disturbances. Appl. Veg. Sci. 2018, 21, 395–410. [Google Scholar] [CrossRef]
- Van Zuidam, B.G.; Peeters, E.T. Wave forces limit the establishment of submerged macrophytes in large shallow lakes. Limnol. Oceanogr. 2015, 60, 1536–1549. [Google Scholar] [CrossRef]
- Ruiz-Montoya, L.; Lowe, R.J.; Kendrick, G.A. Contemporary connectivity is sustained by wind-and current-driven seed dispersal among seagrass meadows. Mov. Ecol. 2015, 3, 9. [Google Scholar] [CrossRef]
- Schubert, H.; Feuerpfeil, P.; Marquardt, R.; Telesh, I.; Skarlato, S. Macroalgal diversity along the Baltic Sea salinity gradient challenges Remane’s species-minimum concept. Mar. Pollut. Bull. 2011, 62, 1948–1956. [Google Scholar] [CrossRef]
- Bonis, A.; Grillas, P.; van Wijck, C.; Lepart, J. The effect of salinity on the reproduction of coastal submerged macrophytes in experimental communities. J. Veg. Sci. 1993, 4, 461–468. [Google Scholar] [CrossRef]
- Van Leeuwen, C.H.; Van der Velde, G.; van Groenendael, J.M.; Klaassen, M. Gut travellers: Internal dispersal of aquatic organisms by waterfowl. J. Biogeogr. 2012, 39, 2031–2040. [Google Scholar] [CrossRef]
- Morkūnė, R.; Petkuvienė, J.; Bružas, M.; Morkūnas, J.; Bartoli, M. Monthly Abundance Patterns and the Potential Role of Waterbirds as Phosphorus Sources to a Hypertrophic Baltic Lagoon. Water 2020, 12, 1392. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth, m | Empty Oospores, % | Empty Gyrogonites, % | Full Oospores, % | Full Gyrogonites, % |
|---|---|---|---|---|
| 0.5–1 | 72.5 | 10.7 | 6.9 | 9.8 |
| 1.5–2 | 75.0 | 16.1 | 0.5 | 8.3 |
| 2–2.5 | 87.4 | 6.1 | 1.6 | 4.9 |
| >2.5 | 96.4 | 3.6 | 0.0 | 0.0 |
| Sites with Presence of Fructifications in the Study Area | |||||
|---|---|---|---|---|---|
| Variable, unit | Minimum | Maximum | Mean | Median | Standard Deviation |
| Depth, m | 0.5 | 4.0 | 1.7 | 1.5 | 0.8 |
| REI | 44 | 271 | 150 | 140 | 67 |
| Distance from the charophyte stands, km | 0.01 | 6.5 | 1.2 | 0.6 | 1.5 |
| Salinity | 0.1 | 1.7 | 0.7 | 0.5 | 0.6 |
| Bottom slope,° | 0.003 | 0.907 | 0.127 | 0.051 | 0.214 |
| Aspect,° | 0.4 | 351.8 | 134.8 | 100.2 | 102.3 |
| TPI | −0.339 | 0.626 | 0.205 | 0.118 | 0.334 |
| Sites without Fructifications in the Study Area | |||||
| Depth, m | 0.5 | 5.0 | 1.8 | 1.5 | 1.0 |
| REI | 71 | 335 | 205 | 218 | 82 |
| Distance from the charophyte stands, km | 0.4 | 13.3 | 5.2 | 4.8 | 3.9 |
| Salinity | 0.1 | 1.8 | 0.3 | 0.1 | 0.4 |
| Bottom slope,° | 0.003 | 1.171 | 0.135 | 0.107 | 0.179 |
| Aspect,° | 1.7 | 357.2 | 203.4 | 225.8 | 109.9 |
| TPI | −0.511 | 1.231 | 0.128 | 0.063 | 0.305 |
| First Cluster | |||||
| Depth, m | 0.5 | 4.0 | 1.7 | 1.5 | 0.7 |
| REI | 44 | 285 | 144 | 137 | 80 |
| Distance from the charophyte stands, km | 0.01 | 6.9 | 1.2 | 0.6 | 1.4 |
| Salinity | 0.1 | 1.8 | 0.8 | 0.5 | 0.6 |
| Bottom slope,° | 0.003 | 0.171 | 0.137 | 0.036 | 0.252 |
| Aspect,° | 0.4 | 357.2 | 159.3 | 119.1 | 113.1 |
| TPI | −0.339 | 0.626 | 0.178 | 0.082 | 0.349 |
| Second Cluster | |||||
| Depth, m | 0.5 | 3.0 | 1.8 | 2.0 | 0.8 |
| REI | 91 | 335 | 202 | 199 | 76 |
| Distance from the charophyte stands, km | 2.1 | 13.3 | 6.1 | 6.3 | 3.6 |
| Salinity | 0.1 | 0.2 | 0.2 | 0.2 | 0.051 |
| Bottom slope,° | 0.009 | 0.308 | 0.119 | 0.102 | 0.084 |
| Aspect,° | 1.7 | 356.6 | 160.7 | 118.3 | 108.7 |
| TPI | −0.512 | 0.626 | 0.124 | 0.132 | 0.244 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stragauskaitė, V.; Bučas, M.; Martin, G. Distribution of Charophyte Oospores in the Curonian Lagoon and their Relationship to Environmental Forcing. Water 2021, 13, 117. https://doi.org/10.3390/w13020117
Stragauskaitė V, Bučas M, Martin G. Distribution of Charophyte Oospores in the Curonian Lagoon and their Relationship to Environmental Forcing. Water. 2021; 13(2):117. https://doi.org/10.3390/w13020117
Chicago/Turabian StyleStragauskaitė, Vaiva, Martynas Bučas, and Georg Martin. 2021. "Distribution of Charophyte Oospores in the Curonian Lagoon and their Relationship to Environmental Forcing" Water 13, no. 2: 117. https://doi.org/10.3390/w13020117
APA StyleStragauskaitė, V., Bučas, M., & Martin, G. (2021). Distribution of Charophyte Oospores in the Curonian Lagoon and their Relationship to Environmental Forcing. Water, 13(2), 117. https://doi.org/10.3390/w13020117