
What’s on the Menu for the Resident Brown Trout in a Rich Limestone Stream?

 and
and
Abstract
:1. Introduction
2. Materials and Methods
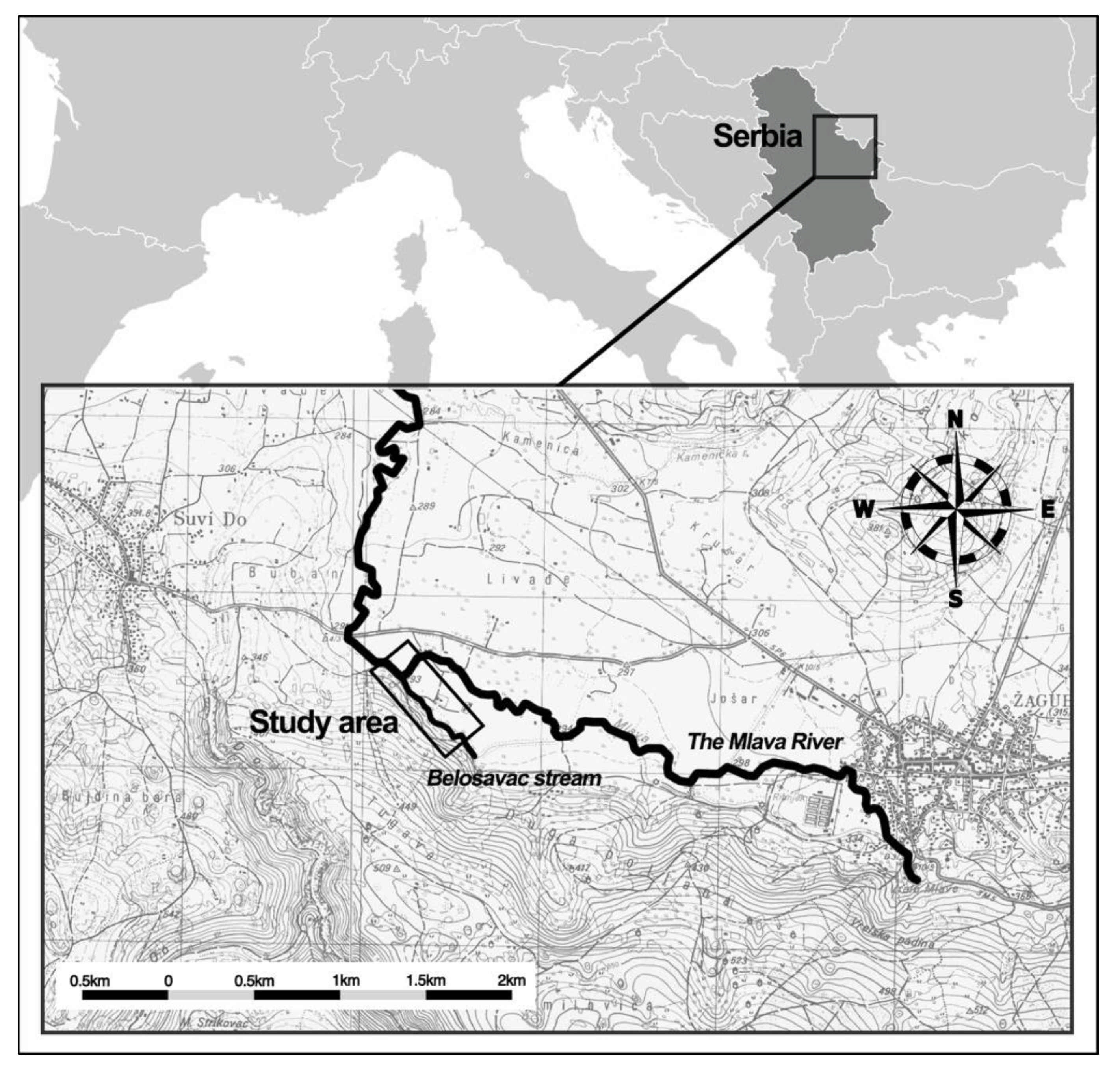
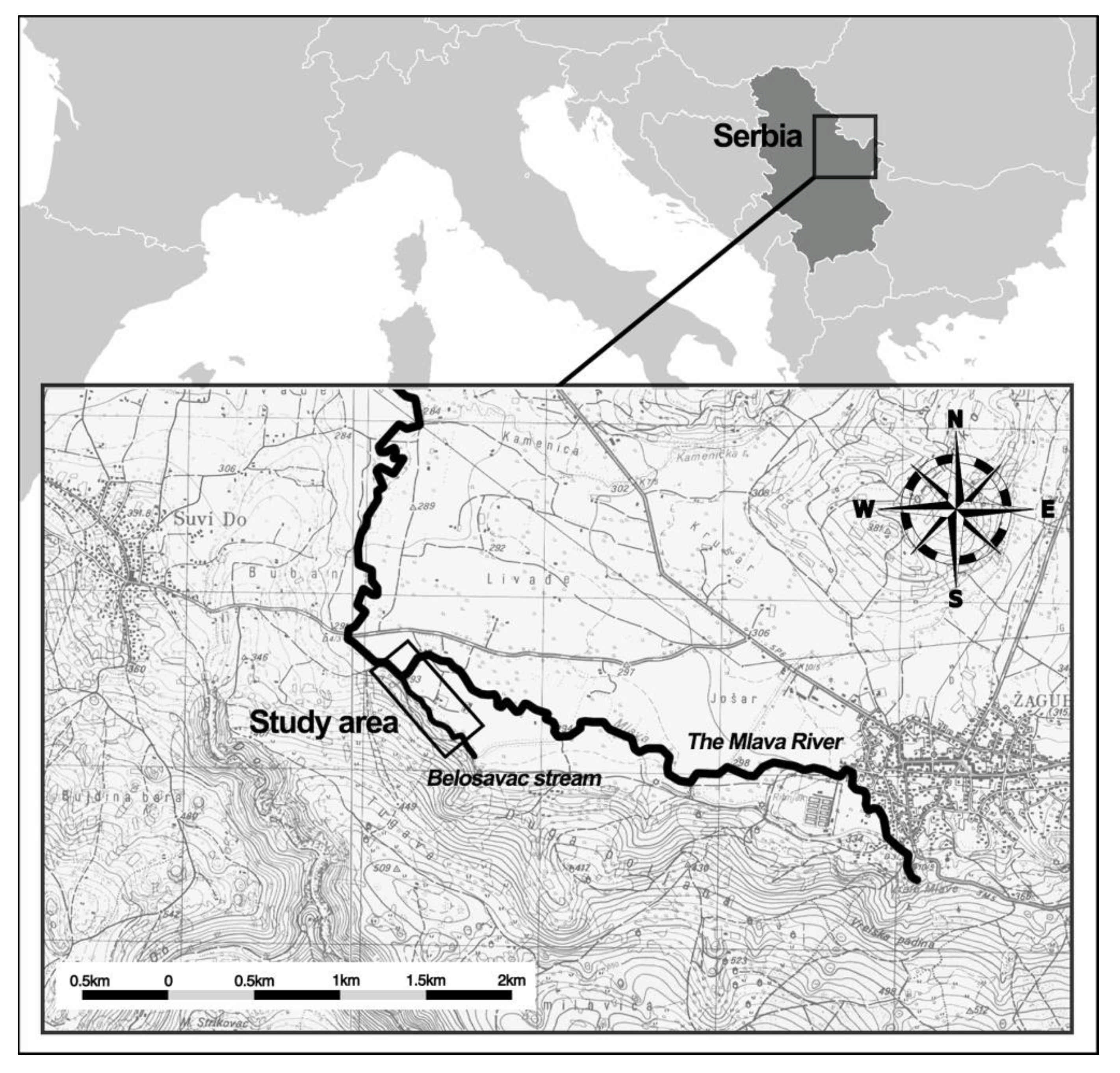
2.1. Study Design and Site/Area Description
2.2. Fish Measuring and Diet Sampling
2.3. Von Bertalanffy Growth Model, Condition Factor, and Length–Weight Regression
2.4. Macroinvertebrates from the Environment and Stomach Content
2.5. Prey Importance and Diet Diversity
2.6. Importance of Terrestrial Prey in a Food-Rich Stream
2.7. Diet Overlap among Age Classes
2.8. Predation Selectivity
2.9. Statistical Analysis
3. Results
3.1. Fish Body Measurements and Growth Parameters
3.2. Benthos, Drift, and Diet Samples
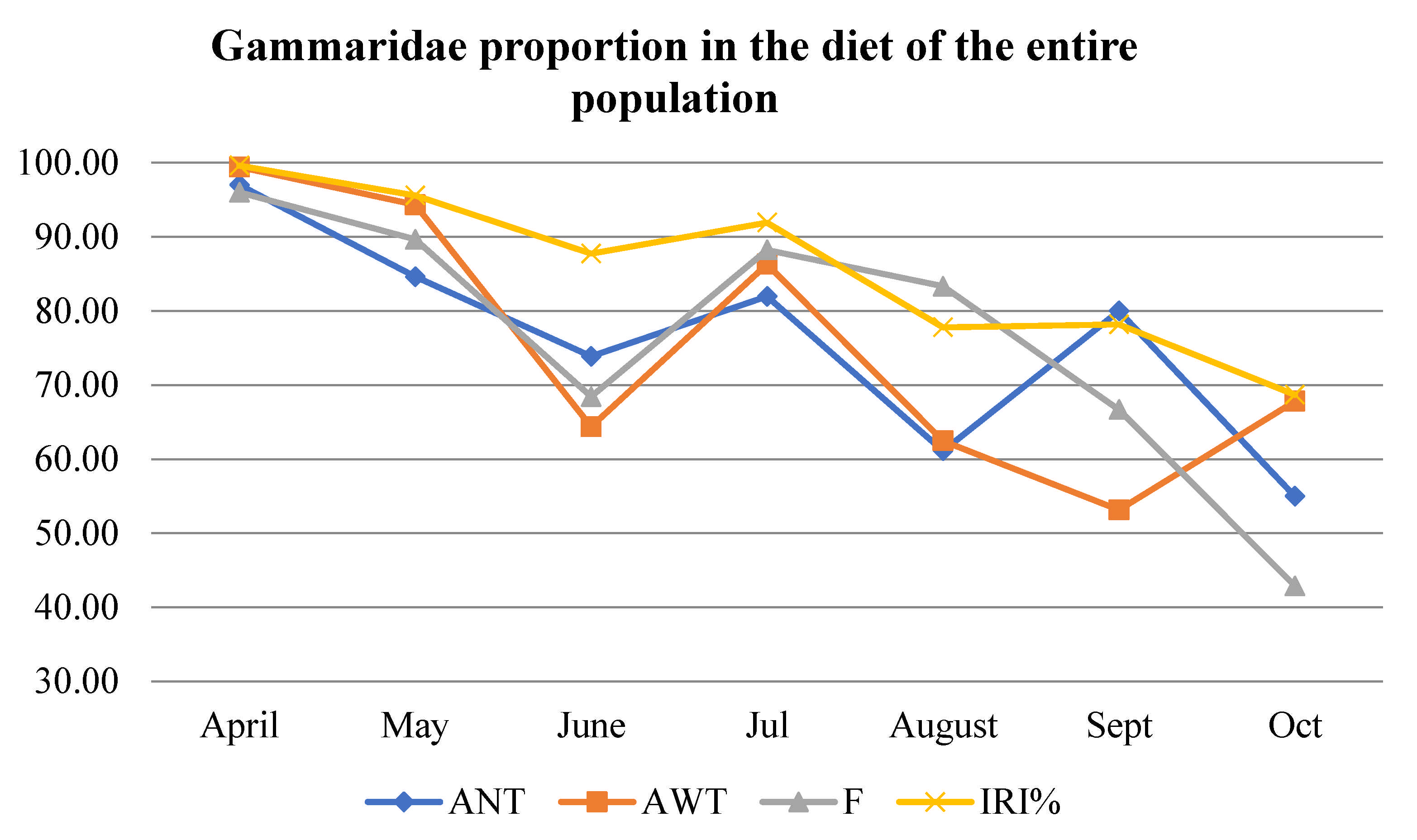
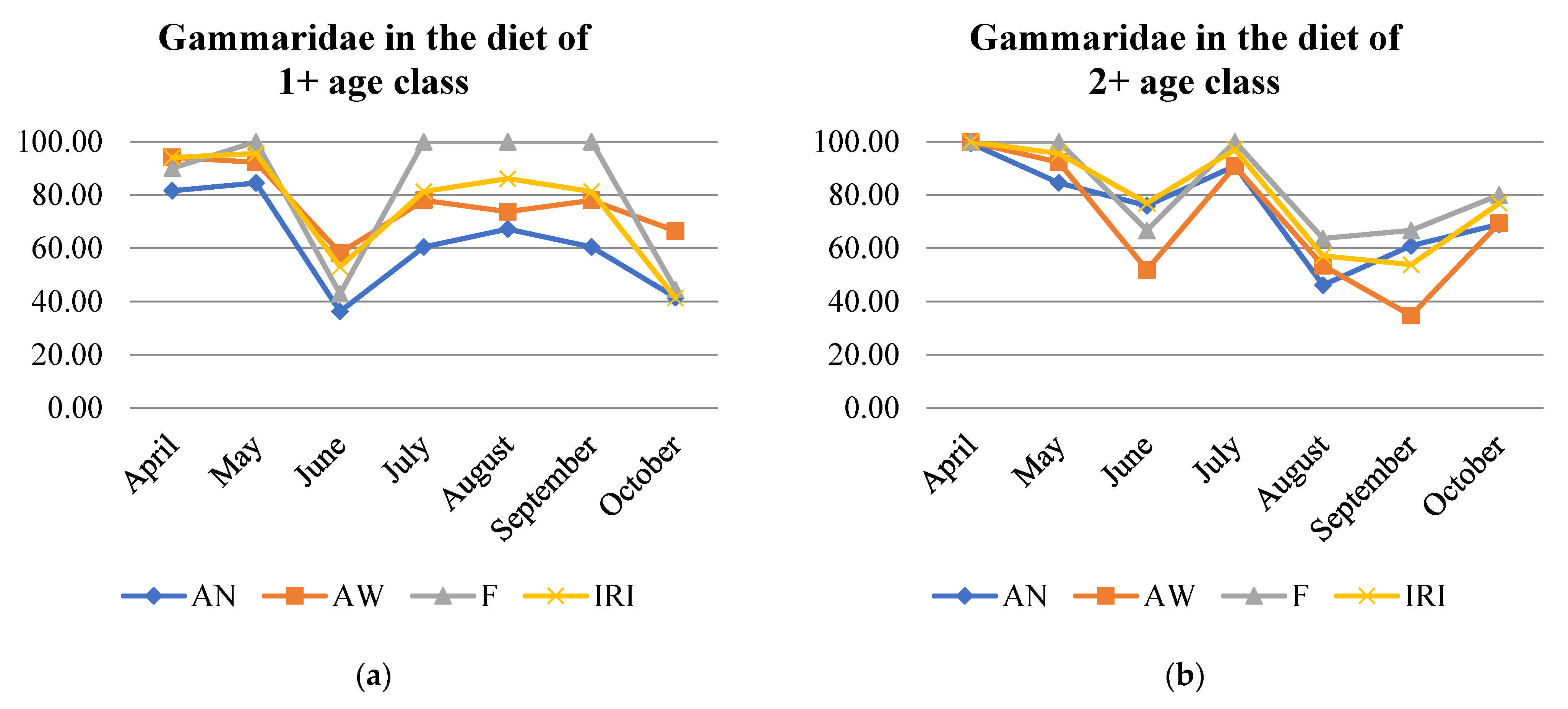
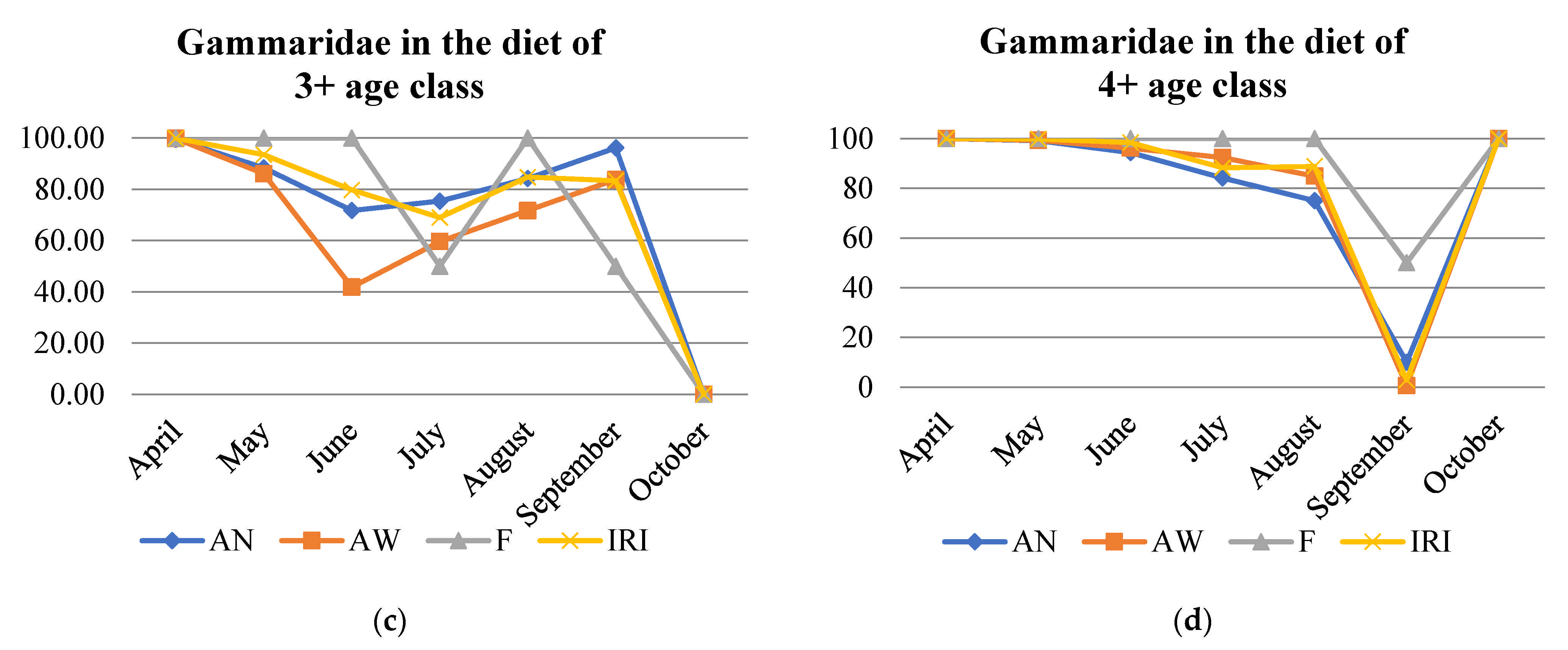
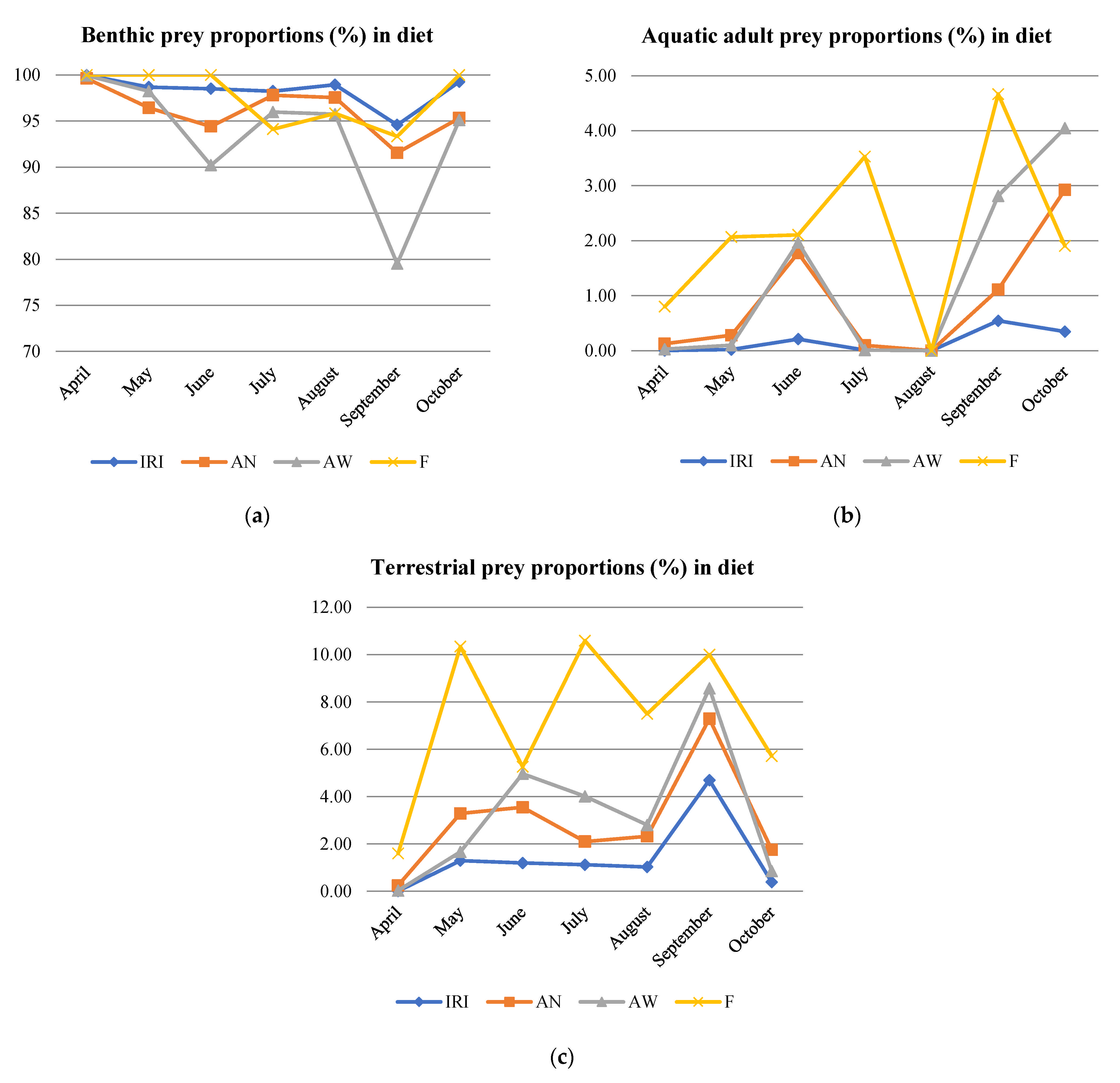
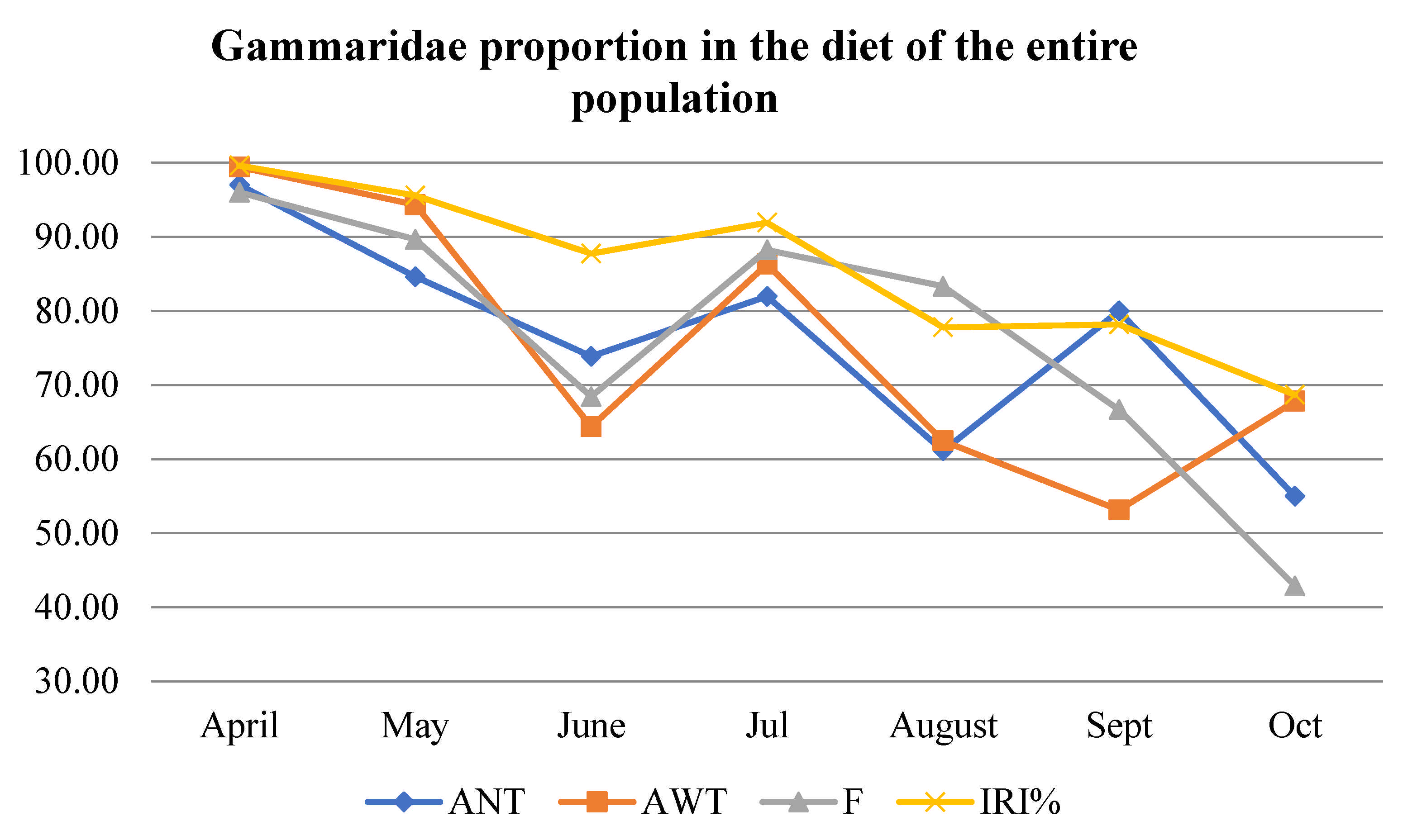
3.3. Diet Description According to Number (AN) and Weight (AW) Abundance, Frequency of Occurrence (F), and Index of Relative Importance (IRI)
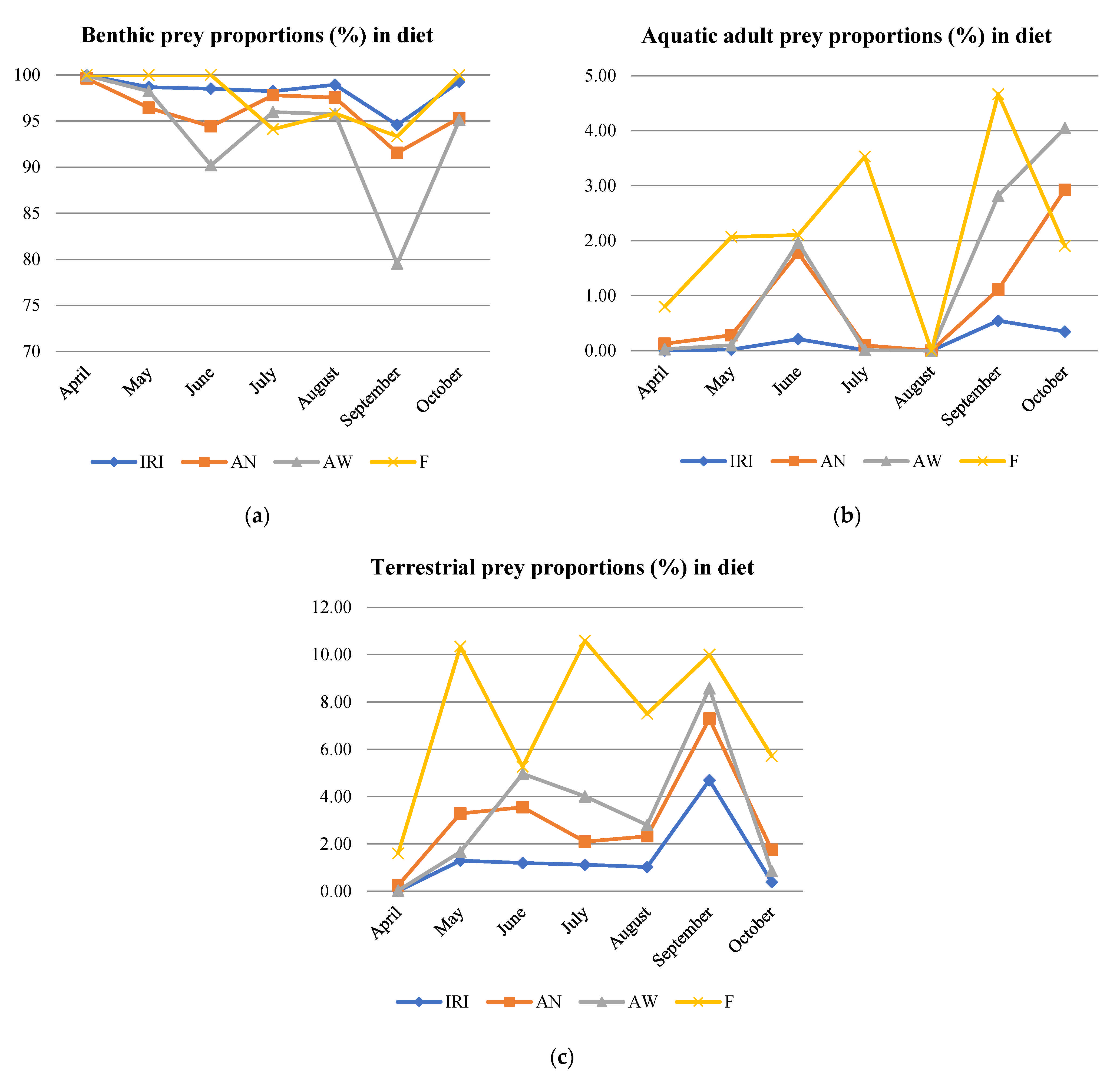
3.4. Terrestrial Prey
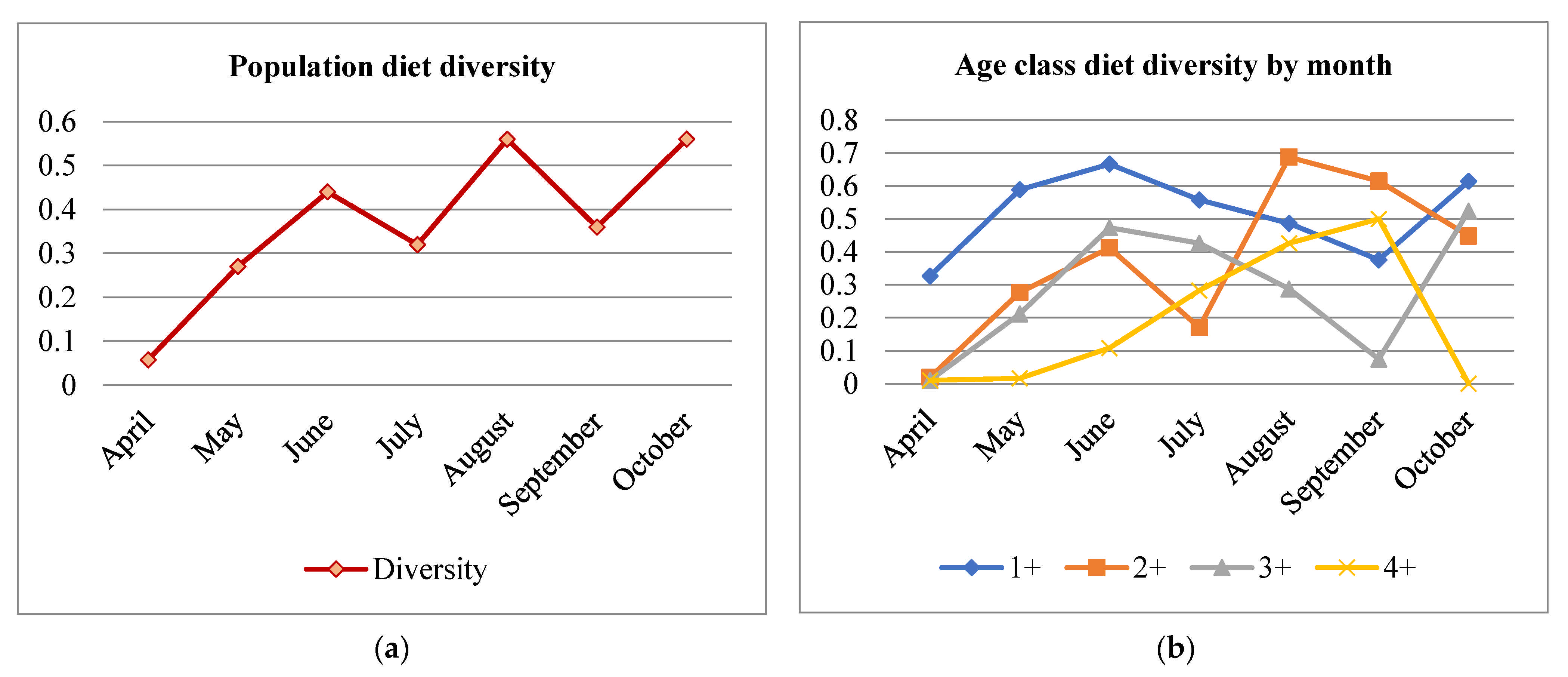
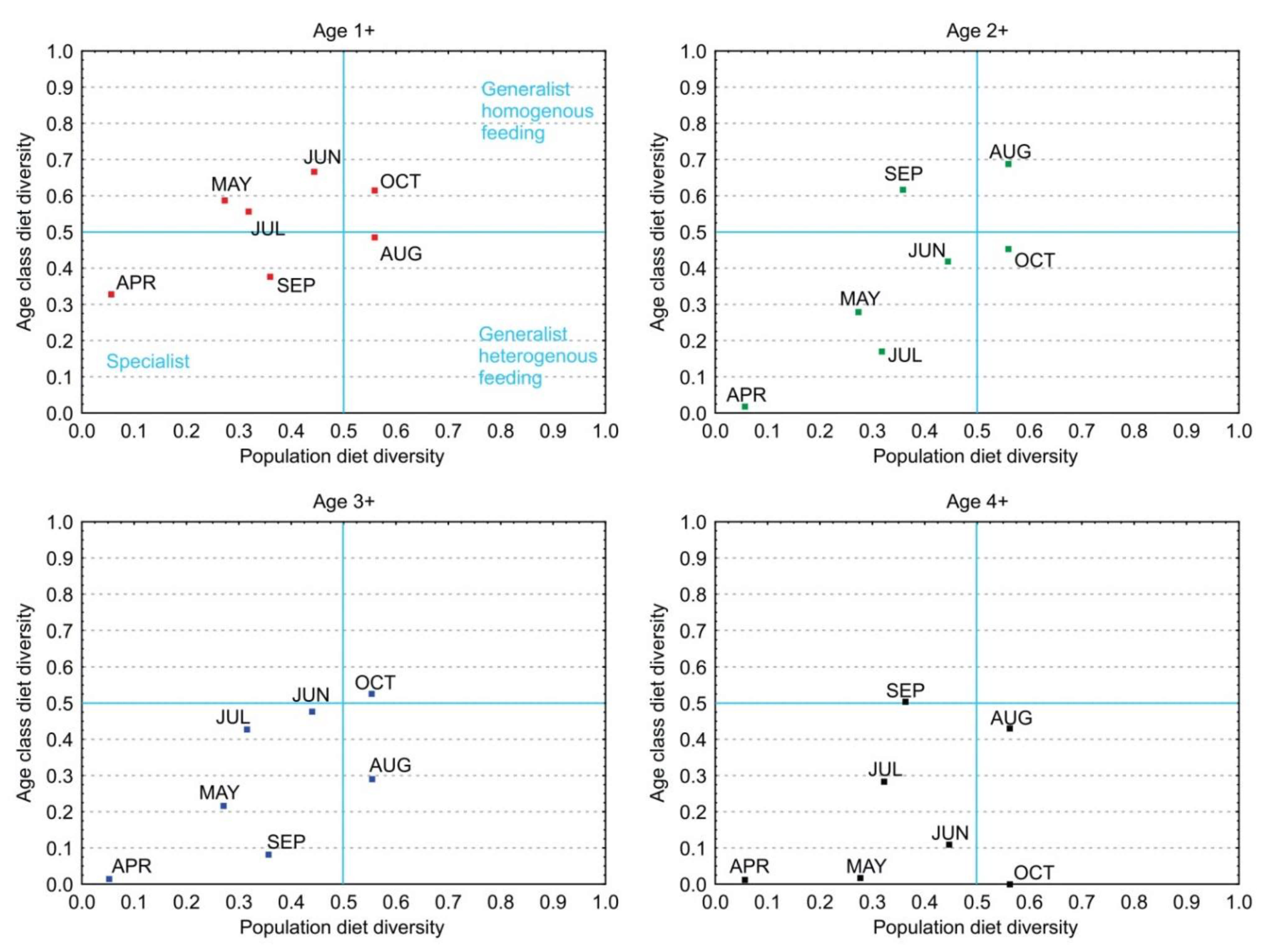
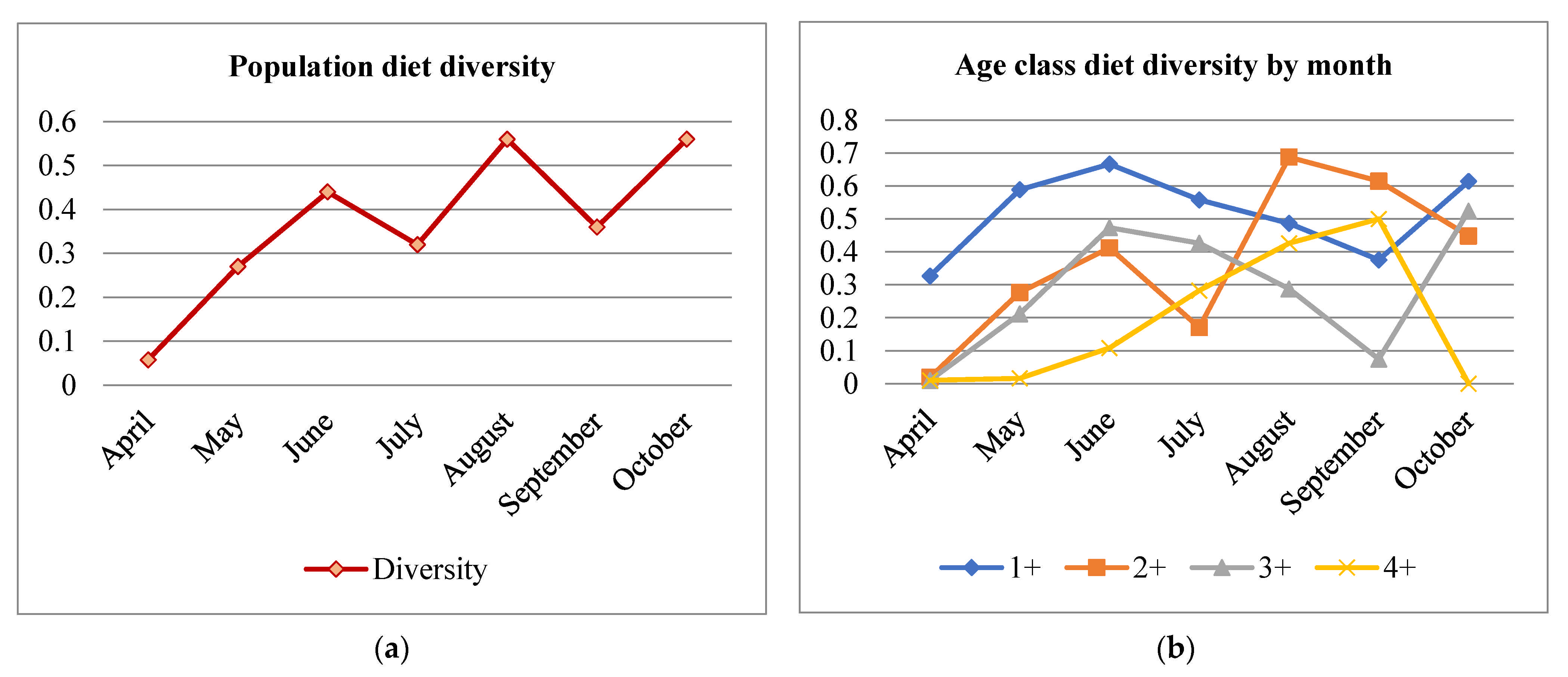
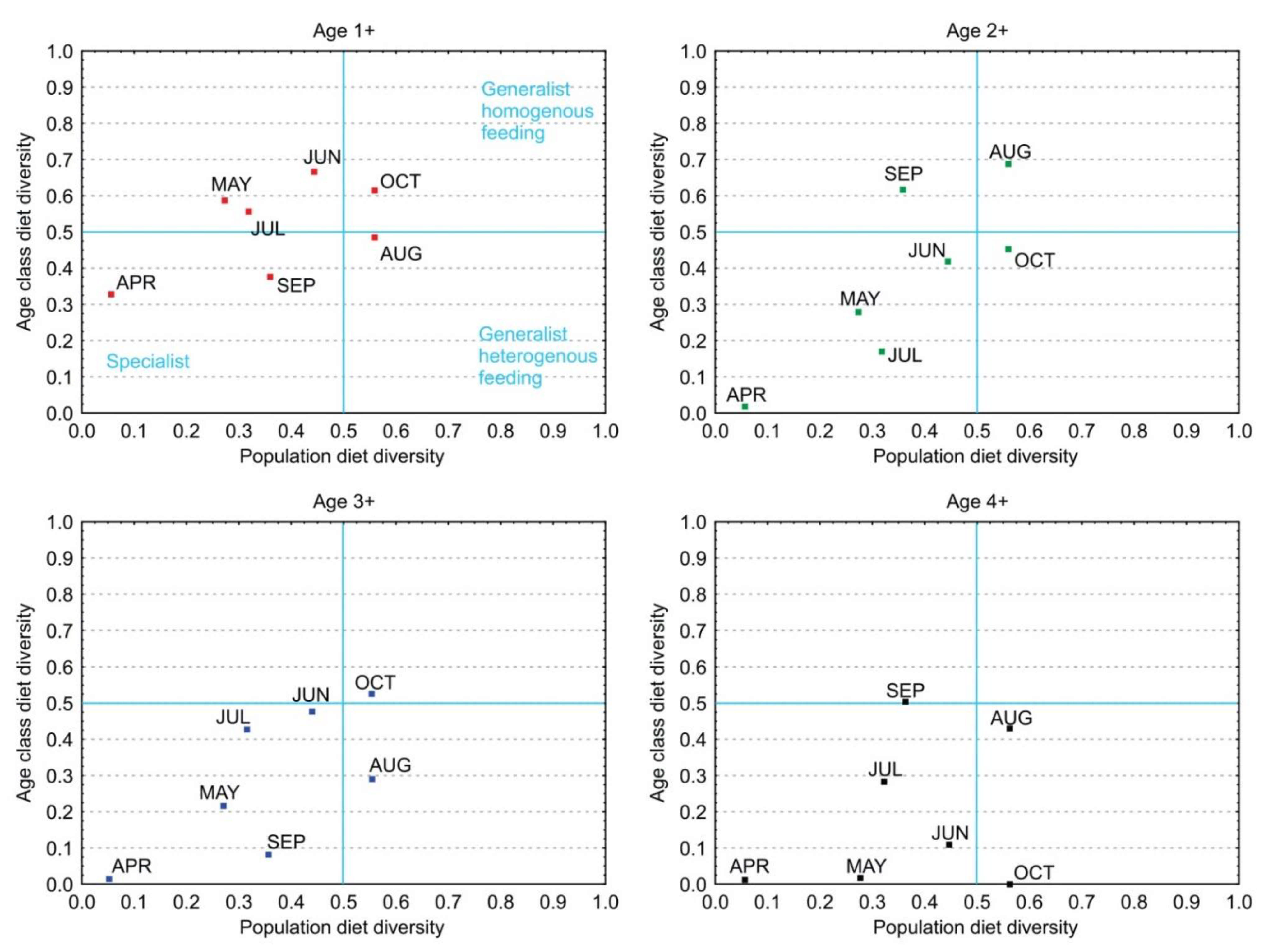
3.5. Diet Diversity
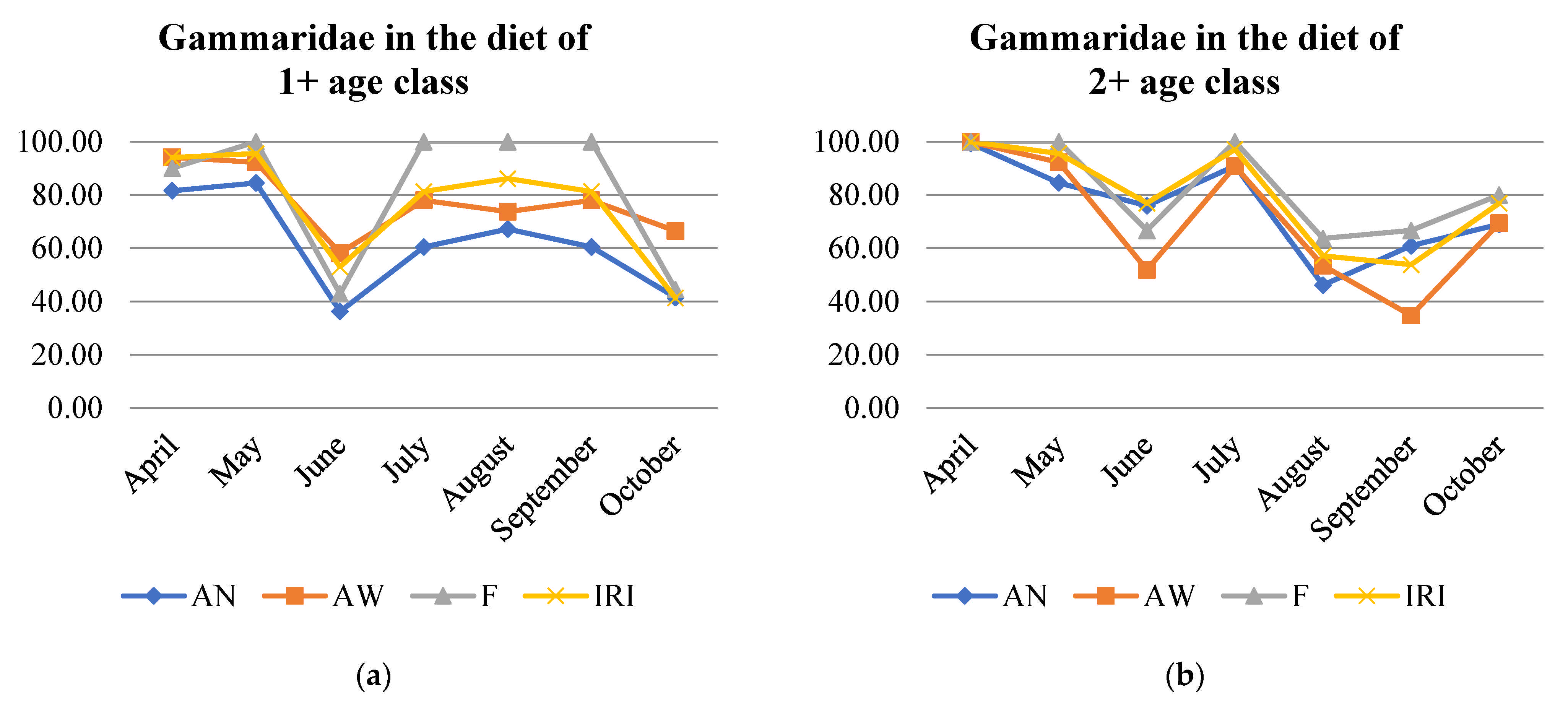
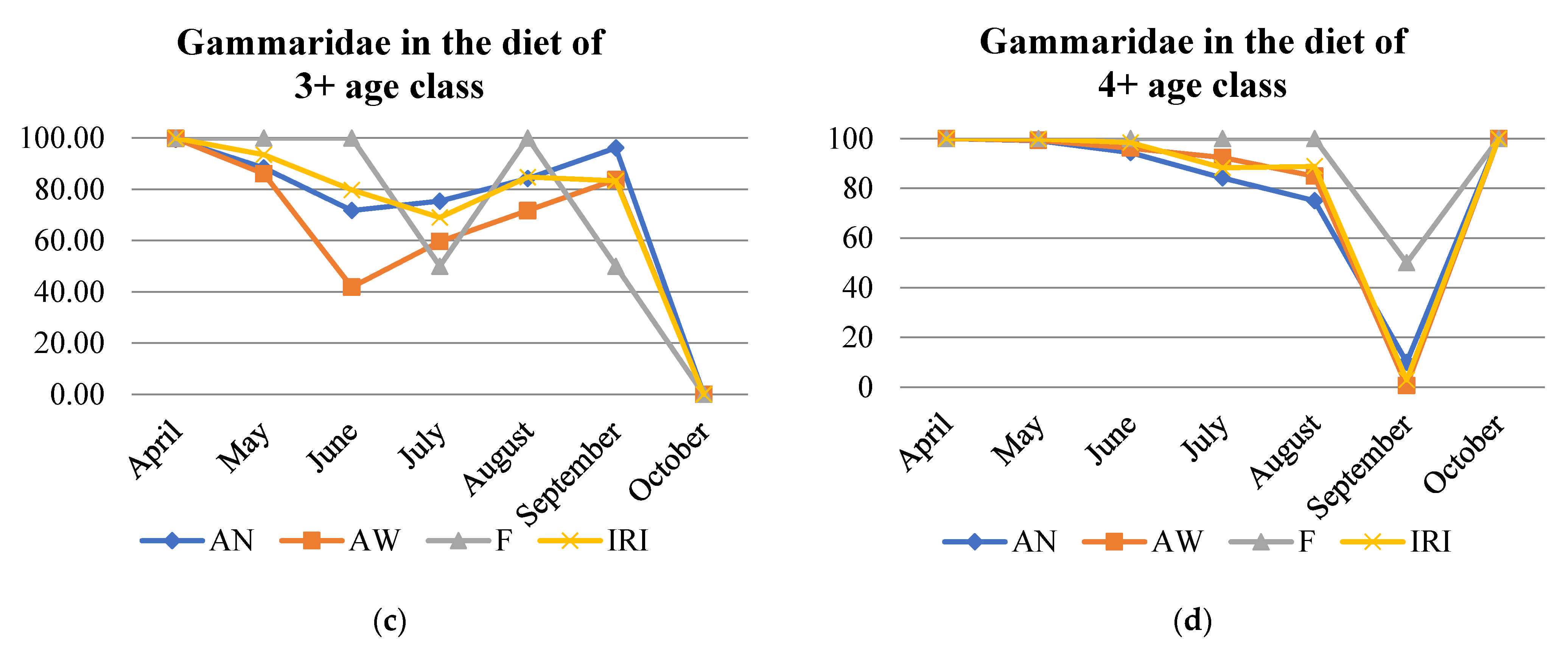
3.6. Diet Overlapping among Age Classes
3.7. Selectivity (Electivity and Tokeshi’s Graphical Model)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elliott, J.M. Quantitative Ecology and the Brown Trout; Oxford University Press: Oxford, UK, 1994; Volume 64, p. 301. ISBN 0198546785. [Google Scholar]
- Elliott, J.M. The food of trout (Salmo trutta) in a Dartmoor stream. J. Appl. Ecol. 1967, 4, 59–71. [Google Scholar] [CrossRef]
- Bridcut, E.E.; Giller, P.S. Diet variability and foraging strategies in brown trout (Salmo trutta): An analysis from subpopulations to individuals. Can. J. Fish. Aquat. Sci. 1995, 52, 2543–2552. [Google Scholar] [CrossRef]
- De Crespin De Billy, V.; Usseglio-Polatera, P. Traits of brown trout prey in relation to habitat characteristics and benthic invertebrate communities. J. Fish Biol. 2002, 60, 687–714. [Google Scholar] [CrossRef]
- De Crespin De Billy, V.; Dumont, B.; Lagarrigue, T.; Baran, P.; Statzner, B. Invertebrate accessibility and vulnerability in the analysis of brown trout (Salmo trutta L.) summer habitat suitability. River Res. Appl. 2002, 18, 533–553. [Google Scholar] [CrossRef]
- Kara, C.; Alp, A. Feeding habits and diet composition of brown trout (Salmo trutta) in the upper streams of River Ceyhan and River Euphrates in Turkey. Turkish J. Vet. Anim. Sci. 2005, 29, 417–428. [Google Scholar]
- Alp, A.; Kara, C.; Büyükcapar, H.M. Age, growth and diet composition of the resident brown trout, Salmo trutta macrostigma Dumeril 1858, in Fırnız Stream of the River Ceyhan, Turkey. Turkish J. Vet. Anim. Sci. 2005, 29, 285–295. [Google Scholar]
- Montori, A.; De Figueroa, J.M.T.; Santos, X. The Diet of the Brown Trout Salmo trutta (L.) during the Reproductive Period: Size-Related and Sexual Effects. Int. Rev. Hydrobiol. 2006, 91, 438–450. [Google Scholar] [CrossRef]
- Frank, M.; Piccolo, J.J.; Baret, P.V. A review of ecological models for brown trout: Towards a new demogenetic model. Ecol. Freshw. Fish 2011, 20, 167–198. [Google Scholar] [CrossRef]
- Sánchez-Hernández, J.; Cobo, F. Summer differences in behavioral feeding habits and use of feeding habitat among brown trout (Pisces) age classes in a temperate area. Ital. J. Zool. 2012, 79, 468–478. [Google Scholar] [CrossRef] [Green Version]
- Giller, P.; Greenberg, L. The relationship between individual habitat use and diet in brown trout. Freshw. Biol. 2015, 60, 256–266. [Google Scholar] [CrossRef]
- Poff, N.L.; Huryn, A.D. Multi-scale determinants of secondary production in Atlantic salmon (Salmo salar) streams. Can. J. Fish. Aquat. Sci. 1998, 55, 201–217. [Google Scholar] [CrossRef]
- Gerking, S.D. Feeding Ecology of Fish, 1st ed.; Elsevier: San Diego, CA, USA, 1994; ISBN 9781483288529. [Google Scholar]
- Keeley, E.R.; Grant, J.W.A. Prey size of salmonid fishes in streams, lakes, and oceans. Can. J. Fish. Aquat. Sci. 2001, 58, 1122–1132. [Google Scholar] [CrossRef]
- Keeley, E.R.; Grant, J. Allometry of diet selectivity in juvenile Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 1997, 54, 1894–1902. [Google Scholar] [CrossRef] [Green Version]
- Kahilainen, K.; Lehtonen, H. Brown trout (Salmo trutta L.) and Arctic charr (Salvelinus alpinus (L.)) as predators on three sympatric whitefish (Coregonus lavaretus (L.)) forms in the subarctic Lake Muddusjärvi. Ecol. Freshw. Fish 2002, 11, 158–167. [Google Scholar] [CrossRef]
- Jensen, H.; Bøhn, T.; Amundsen, P.A.; Aspholm, P.E. Feeding ecology of piscivorous brown trout (Salmo trutta L.) in a subarctic watercourse. Ann. Zool. Fenn. 2004, 41, 319–328. [Google Scholar]
- Vøllestad, L.; Andersen, R. Resource partitioning of various age groups of brown trout Salmo trutta in the littoral zone of Lake Selura, Norway. Arch. Hydrobiol. 1985, 105, 177–185. [Google Scholar]
- Wańkowski, J.W.J.; Thorpe, J.E. The role of food particle size in the growth of juvenile Atlantic salmon (Salmo salar L.). J. Fish Biol. 1979, 14, 351–370. [Google Scholar] [CrossRef]
- Fochetti, R.; Argano, R.; Tierno de Figueroa, J.M. Feeding ecology of various age-classes of brown trout in River Nera, Central Italy. Belgian J. Zool. 2008, 138, 128–131. [Google Scholar]
- Fausch, K.D. Profitable stream positions for salmonids: Relating specific growth rate to net energy gain. Can. J. Zool. 1984, 62, 441–451. [Google Scholar] [CrossRef]
- Hunt, P.C.; Jones, J.W. The food of brown trout in Llyn Alaw, Anglesey, North Wales. J. Fish Biol. 1972, 4, 333–352. [Google Scholar] [CrossRef]
- Hynes, H.B.N. The Ecology of Running Waters; Liverpool University Press: Liverpool, UK, 1970. [Google Scholar]
- Chesson, J. Measuring preference in selective predation. Ecology 1978, 59, 211–215. [Google Scholar] [CrossRef]
- Fochetti, R.; Amici, I.; Argano, R. Seasonal changes and selectivity in the diet of brown trout in the River Nera (central Italy). J. Freshw. Ecol. 2003, 18, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Rosenbauer, T. The Orvis Guide to Reading Trout Streams; Lyons Press: New York, NY, USA, 1999; ISBN 1558219331. [Google Scholar]
- Swisher, D.; Richards, C. Selective Trout; Lyons Press: New York, NY, USA, 2000; ISBN 1-58574-038-1. [Google Scholar]
- AQEM Consortium. Manual for the Application of the AQEM Method. A Comprehensive Method to Assess European Streams Using Benthic Macroinvertebrates, Developed for the Purpose of the Water Framework Directive, Version 1.0; AQEM Consortium: Duisburg, Germany, 2002. [Google Scholar]
- Čanak Atlagić, J.; Marić, A.; Đuknić, J.; Andjus, S.; Marinković, N.; Paunović, M.; Simonović, P. The efficiency of syringe stomach flushing in diet sampling of salmonids. Acta Ichthyol. Piscat. 2019, 49, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Pauly, D.; Munro, J. A simple method for comparing the growth of fishes and invertebrates. Fishbyte 1983, 1, 5–6. [Google Scholar]
- Von Bertalanffy, L. Quantitative laws in metabolism and growth. Q. Rev. Biol. 1957, 32, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Ricker, W.E. Computation and Interpretation of Biological Statistics of Fish Populations. Bull. Fish. Res. Board Can. 1975, 191, 1–382. [Google Scholar] [CrossRef]
- Pinkas, L.; Oliphant, M.; Iverson, L.K. Food Habits of Albacore, Bluefin Tuna and Bonito in California Waters; Department of Fish and Game, State of California: Sacramento, CA, USA, 1971; Volume 152.
- Pita, C.; Gamito, S.; Erzini, K. Feeding habits of the gilthead seabream (Sparus aurata) from the Ria Formosa (southern Portugal) as compared to the back seabream (Spondyliosoma cantharus) and the annular seabream (Diplodus annularis). J. Appl. Ichthyol. 2002, 18, 81–86. [Google Scholar] [CrossRef]
- Pianka, E.R. The Structure of Lizard Communities. Annu. Rev. Ecol. Syst. 1973, 4, 53–74. [Google Scholar] [CrossRef] [Green Version]
- Vanderploeg, H.A.; Scavia, D. Two Electivity Indices for Feeding with Special Reference to Zooplankton Grazing. J. Fish. Res. Board Can. 1979, 36, 362–365. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Scavia, D. Calculation and use of selectivity coefficients of feeding: Zooplankton grazing. Ecol. Modell. 1979, 7, 135–149. [Google Scholar] [CrossRef]
- Tokeshi, M. Graphical analysis of predator feeding strategy and prey importance. Freshw. Forum 1991, 1, 179–183. [Google Scholar]
- Rader, R.B. A functional classification of the drift: Traits that influence invertebrate availability to salmonids. Can. J. Fish. Aquat. Sci. 1997, 54, 1211–1234. [Google Scholar] [CrossRef]
- Liao, H.; Pierce, C.L.; Larscheid, J.G. Empirical assessment of indices of prey importance in the diets of predacious fish. Trans. Am. Fish. Soc. 2001, 130, 583–591. [Google Scholar] [CrossRef]
- Roussel, J.; Bardonnet, A. Ontogeny of diel pattern of stream margin habitat use by emerging brown trout, Salmo trutta, in experimental channels: Influence of food and predator presence. Environ. Biol. Fishes 1999, 56, 253–262. [Google Scholar] [CrossRef]
- Ayllón, D.; Almodóvar, A.; Nicola, G.G.; Elvira, B. Ontogenetic and spatial variations in brown trout habitat selection. Ecol. Freshw. Fish 2010, 19, 420–432. [Google Scholar] [CrossRef]
- Nakano, S.; Kawaguchi, Y.; Taniguchi, Y.; Miyasaka, H.; Shibata, Y.; Urabe, H.; Kuhara, N. Selective foraging on terrestrial invertebrates by rainbow trout in a forested headwater stream in northern Japan. Ecol. Res. 1999, 14, 351–360. [Google Scholar] [CrossRef]
- Sánchez, J.; Cobo, F.; González, M.A. Biología y la alimentación del salvelino, “Salvelinus fontinalis” (Mitchill, 1814), en cinco lagunas glaciares de la Sierra de Gredos (Ávila, España). Nov. Acta Científica Compostel. 2007, 16, 129–144. [Google Scholar]
- Utz, R.M.; Hartman, K.J. Identification of critical prey items to Appalachian brook trout (Salvelinus fontinalis) with emphasis on terrestrial organisms. Hydrobiologia 2007, 575, 259–270. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Nakano, S. Contribution of terrestrial invertebrates to the annual resource budget for salmonids in forest and grassland reaches of a headwater stream. Freshw. Biol. 2001, 46, 303–316. [Google Scholar] [CrossRef]
- Almany, G.R.; Peacock, L.F.; Syms, C.; McCormick, M.I.; Jones, G.P. Predators target rare prey in coral reef fish assemblages. Oecologia 2007, 152, 751–761. [Google Scholar] [CrossRef]
- Sweka, J.A.; Hartman, K.J. Contribution of Terrestrial Invertebrates to Yearly Brook Trout Prey Consumption and Growth. Trans. Am. Fish. Soc. 2008, 137, 224–235. [Google Scholar] [CrossRef]
- Rosenbauer, T. Orvis Guide to Prospecting for Trout, New and Revised: How to Catch Fish When There’s No Hatch to Match; Lyons Press: New York, NY, USA, 2008; ISBN 1599211475. [Google Scholar]
- Simonović, P.; Marić, A.; Jurlina, D.Š.; Kanjuh, T.; Nikolić, V. Determination of resident brown trout Salmo trutta features by their habitat characteristics in streams of Serbia. Biologia 2019, 75, 103–114. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Class | April | May | June | July | August | September | October | Mean CF for Each Age Class |
|---|---|---|---|---|---|---|---|---|
| 0+ | - | - | - | - | 1.67 | 1.58 | 1.53 | 1.59 |
| 1+ | 1.49 | 1.82 | 1.65 | 1.81 | 1.71 | 1.64 | 1.45 | 1.66 |
| 2+ | 1.51 | 1.72 | 1.98 | 1.65 | 1.66 | 1.63 | 1.52 | 1.67 |
| 3+ | 1.67 | 1.74 | 1.85 | 1.79 | 1.62 | 1.84 | 1.56 | 1.73 |
| 4+ | 1.74 | 1.94 | 1.74 | 1.86 | 1.66 | 1.66 | 1.47 | 1.73 |
| Mean CF for each month | 1.60 | 1.81 | 1.81 | 1.78 | 1.67 | 1.67 | 1.51 |
| Prey Type | Included Taxa | |
|---|---|---|
| Benthic prey | ||
| 1 | Oligochaeta | Oligochaeta, Eiseniella tetraedra and others |
| 2 | Gammaridae | Gammarus sp. |
| 3 | Platyhelminthes and Hirudinea | Dugesia lugubris, Schmidt, 1861., Crenobia alpina, Dana, 1766, Erpobdella vilnensis, Liskiewicz, 1925 |
| 4 | Mollusca | Pisidium sp., Ancylus sp. |
| 5 | Hydracarina | Hydracarina |
| 6 | Chironomidae and Simuliidae | Chironomidae and Simuliidae (larvae and pupae) |
| 7 | Limoniidae and Pediciidae | Eloeophila sp., Dicranota sp., Dicranoptycha sp. (larvae) |
| 8 | Tipulidae and Tabanidae | Tabanus sp., Tipula sp. (larvae) |
| 9 | Ephemeroptera larvae | Baetis sp., Heptagenia sp., Ephemera danica, Müller, 1764, Ephemerella sp. |
| 10 | Plecoptera larvae | Nemoura sp., Rhabdiopteryx sp. |
| 11 | Trichoptera larvae with cases | Beraeidae, Glossosoma sp., Potamophylax |
| 12 | Trichoptera larvae caseless | Lype sp., Plectrocnemia sp., Rhyacophila sp. |
| 13 | Aquatic Coleoptera and Heteroptera larvae | Limnius sp., Elodes sp., Elmis aenea, Dytiscidae (larvae) |
| Flying adults of aquatic prey (aerial adults) | ||
| 14 | Ephemeroptera adults | Baetis sp. |
| 15 | Trichoptera adults | Glossosoma sp., Lype reducta, Potamophylax luctuosus, P. latipennis, Silo nigricornis, Psychomyiidae |
| 16 | Aquatic Diptera adults | Chironomidae, Simuliidae, Dolichopodidae |
| 17 | Plecoptera adults | Plecoptera |
| 18 | Aquatic Coleoptera and Heteroptera adults | Limnius sp., Hydraena gracilis, Esolus angustatus, Elmis aenea, Pomatinus substriatus, Mesoveliidae |
| Terrestrial prey (adults and larvae) | ||
| 19 | Acari, Araneae, and Opiliones | Acari, Araneae, and Opiliones |
| 20 | Collembola | Collembola |
| 21 | Gastropoda | Gastropoda |
| 22 | Diptera | Muscidae and others |
| 23 | Coleoptera and Staphylinidae | Chrysomelidae, Cantharidae, Curculionidae, Coccinellidae, Scarabaeidae and others, Staphylinidae |
| 24 | Hymenoptera | Hoplocampa flava, Vespoidea, and others |
| 25 | Hymenoptera Formicidae | Formicidae |
| 26 | Heteroptera | Cicadomorpha and others |
| 27 | Dermaptera, Mecoptera, Psocoptera and Neuroptera. | Forficula auricularia, Linnaeus, 1758, Panorpa sp., Osmylus sp. |
| 28 | Lepidoptera larvae | Lepidoptera larvae |
| 29 | Orthoptera | Ensifera |
| Fish | ||
| 30 | Fish | Cottus gobio, Phoxinus phoxinus |
| AN | APR | MAY | JUN | JUL | AUG | SEP | OCT |
|---|---|---|---|---|---|---|---|
| Gammaridae | 97.01 | 84.62 | 73.86 | 81.95 | 61.11 | 79.98 | 54.97 |
| Ephemeroptera larvae | 1.37 | 10.07 | 8.12 | 10.60 | 4.26 | 2.27 | |
| Plecoptera larvae | 0.75 | ||||||
| Chironomidae and Simuliidae | 0.50 | 0.84 | 3.06 | 23.39 | 4.33 | 37.04 | |
| Trichoptera adult | 0.12 | 2.14 | |||||
| Heteroptera | 2.66 | 0.67 | |||||
| Coleoptera and Staphylinidae | 0.35 | ||||||
| Trichoptera with cases | 5.84 | 1.15 | 4.91 | 3.22 | 1.17 | ||
| Platyhelminthes and Hirudinea | 3.81 | ||||||
| Acari, Araneae, and Opiliones | 1.27 | ||||||
| Aq. Coleoptera and Heteroptera larvae | 1.42 | ||||||
| Formicidae | 3.28 | ||||||
| Dermaptera, Mecoptera, Psocoptera, and Neuroptera | 0.97 | ||||||
| AW | APR | MAY | JUN | JUL | AUG | SEP | OCT |
| Gammaridae | 99.43 | 94.32 | 64.36 | 86.31 | 62.46 | 53.12 | 67.79 |
| Plecoptera larvae | 0.29 | ||||||
| Ephemeroptera larvae | 0.18 | 1.66 | 5.18 | ||||
| Chironomidae and Simuliidae | 0.05 | 1.03 | 5.81 | 11.57 | |||
| Trichoptera adult | 0.03 | 2.42 | 3.91 | ||||
| Trichoptera with cases | 1.24 | 16.61 | 2.39 | 21.79 | 21.90 | 12.54 | |
| Heteroptera | 0.81 | 1.62 | |||||
| Trichoptera caseless | 0.52 | 2.56 | |||||
| Platyhelminthes and Hirudinea | 4.01 | ||||||
| Phoxinus phoxinus | 2.86 | ||||||
| Cottus gobio | 9.10 | ||||||
| Oligochaeta | 1.81 | ||||||
| Muscidae | 1.61 | ||||||
| Formicidae | 2.54 | ||||||
| Tipulidae and Tabanidae | 1.86 | ||||||
| F | APR | MAY | JUN | JUL | AUG | SEP | OCT |
| Gammaridae | 96 | 89.66 | 68.42 | 88.24 | 83.33 | 66.67 | 42.86 |
| Ephemeroptera larvae | 28 | 48.28 | 21.05 | 58.82 | 33.33 | 40.00 | |
| Plecoptera larvae | 20 | ||||||
| Chironomidae and Simuliidae | 8 | 24.14 | 29.41 | 45.83 | 63.33 | 66.67 | |
| Trichoptera adult | 4 | 9.52 | |||||
| Heteroptera | 31.03 | ||||||
| Trichoptera with cases | 31.58 | 41.18 | 45.83 | 63.33 | 9.52 | ||
| Trichoptera caseless | 13.79 | ||||||
| Platyhelminthes and Hirudinea | 26.32 | ||||||
| Acari, Araneae, and Opiliones | 26.32 | 40.00 | |||||
| Formicidae | 29.41 | ||||||
| Aq. Coleoptera and Heteroptera larvae | 20.83 | ||||||
| Dermaptera, Mecoptera, Psocoptera and Neuroptera | 14.29 | ||||||
| IRI | APR | MAY | JUN | JUL | AUG | SEP | OCT |
| Gammarus | 99.63 | 95.59 | 87.75 | 91.93 | 77.78 | 77.68 | 60.03 |
| Ephemeroptera larvae | 0.23 | 3.38 | 1.90 | 5.75 | 1.47 | 1.18 | |
| Plecoptera larvae | 0.11 | ||||||
| Chironomidae and Simuliidae | 0.02 | 0.14 | 0.74 | 10.11 | 2.80 | 36.98 | |
| Trichoptera adult | 0.003 | 0.66 | |||||
| Heteroptera | 0.64 | ||||||
| Trichoptera with cases | 0.09 | 6.58 | 0.90 | 9.25 | 13.93 | 1.49 | |
| Trichoptera caseless | 0.45 | ||||||
| Platyhelminthes and Hirudinea | 1.91 | ||||||
| Acari, Araneae, and Opiliones | 0.45 | ||||||
| Formicidae | 0.21 | 1.53 | |||||
| Tipulidae and Tabanidae | 0.24 |
| 1+ and 2+ | 2+ and 3+ | 3+ and 4+ | 4+ and 1+ | 4+ and 2+ | 1+ and 3+ | |
|---|---|---|---|---|---|---|
| April | 0.99 | 1.00 | 1.00 | 0.99 | 1.00 | 0.99 |
| May | 0.84 | 0.99 | 0.99 | 0.78 | 0.99 | 0.78 |
| June | 0.64 | 0.99 | 0.99 | 0.62 | 0.99 | 0.62 |
| July | 0.93 | 0.99 | 0.99 | 0.94 | 1.00 | 0.93 |
| August | 0.95 | 0.84 | 0.98 | 0.98 | 0.90 | 0.95 |
| September | 0.93 | 0.98 | 0.16 | 0.16 | 0.21 | 0.91 |
| October | 0.89 | 0.36 | 0.00 | 0.67 | 0.01 | 0.72 |
| Gammaridae | APR | MAY | JUN | JUL | AUG | SEP | OCT | |
|---|---|---|---|---|---|---|---|---|
| 1+ | MZB | 0 | - | - | - | - | - | - |
| Drift | - | - | - | - | - | 0 | - | |
| 2+ | MZB | + | - | - | - | - | - | - |
| Drift | + | - | - | 0 | - | - | - | |
| 3+ | MZB | + | - | - | - | - | + | - |
| Drift | + | + | - | - | - | 0 | - | |
| 4+ | MZB | + | + | - | + | - | + | + |
| Drift | + | + | + | + | - | - | + | |
| Chironomidae and Simuliidae | APR | MAY | JUN | JUL | AUG | SEP | OCT | |
| 1+ | MZB | + | 0 | - | - | - | 0 | + |
| Drift | - | - | - | + | + | + | + | |
| 2+ | MZB | - | - | 0 | - | - | 0 | + |
| Drift | - | - | 0 | - | + | + | + | |
| 3+ | MZB | - | - | - | - | - | 0 | 0 |
| Drift | - | - | - | - | + | + | + | |
| 4+ | MZB | - | - | - | - | - | - | - |
| Drift | - | - | - | 0 | 0 | - | - | |
| Ephemeroptera larvae | APR | MAY | JUN | JUL | AUG | SEP | OCT | |
| 1+ | MZB | + | + | + | + | + | + | + |
| Drift | - | + | + | + | 0 | + | + | |
| 2+ | MZB | 0 | + | + | + | + | + | + |
| Drift | 0 | 0 | 0 | 0 | 0 | + | + | |
| 3+ | MZB | 0 | 0 | / | + | + | + | / |
| Drift | 0 | + | - | 0 | - | + | / | |
| 4+ | MZB | - | 0 | - | + | - | / | / |
| Drift | - | 0 | - | + | - | - | / | |
| Trichoptera with cases | APR | MAY | JUN | JUL | AUG | SEP | OCT | |
| 1+ | MZB | - | / | + | 0 | + | + | 0 |
| Drift | - | - | + | + | 0 | 0 | - | |
| 2+ | MZB | - | + | + | - | + | + | + |
| Drift | - | 0 | + | 0 | + | + | - | |
| 3+ | MZB | - | / | + | + | + | + | - |
| Drift | - | - | + | + | + | + | - | |
| 4+ | MZB | - | / | + | - | - | + | - |
| Drift | - | - | + | - | - | + | - | |
| Trichoptera caseless | APR | MAY | JUN | JUL | AUG | SEP | OCT | |
| 1+ | MZB | - | 0 | + | + | + | + | + |
| Drift | / | + | + | + | + | 0 | - | |
| 2+ | MZB | - | + | - | + | + | + | / |
| Drift | / | 0 | - | + | + | 0 | - | |
| 3+ | MZB | - | - | + | / | - | - | / |
| Drift | / | - | + | - | - | 0 | - | |
| 4+ | MZB | - | + | - | / | - | + | / |
| Drift | / | + | - | - | - | + | - | |
| Acari, Araneae, and Opiliones | APR | MAY | JUN | JUL | AUG | SEP | OCT | |
| 1+ | MZB | / | / | / | / | / | / | / |
| Drift | / | / | + | + | - | 0 | / | |
| 2+ | MZB | / | / | / | / | / | / | / |
| Drift | + | + | + | - | - | + | + | |
| 3+ | MZB | / | / | / | / | / | / | / |
| Drift | / | / | + | + | - | - | / | |
| 4+ | MZB | / | / | / | / | / | / | / |
| Drift | / | / | + | + | - | - | / | |
| Coleoptera and Staphylinidae | APR | MAY | JUN | JUL | AUG | SEP | OCT | |
| 1+ | MZB | / | / | / | / | / | / | / |
| Drift | / | + | - | + | / | + | + | |
| 2+ | MZB | / | / | / | / | / | / | / |
| Drift | / | + | / | / | + | + | / | |
| 3+ | MZB | / | / | / | / | / | / | / |
| Drift | / | / | 0 | / | / | / | / | |
| 4+ | MZB | / | / | / | / | / | / | / |
| Drift | / | / | - | / | / | / | / | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čanak Atlagić, J.; Marić, A.; Tubić, B.; Andjus, S.; Đuknić, J.; Marković, V.; Paunović, M.; Simonović, P. What’s on the Menu for the Resident Brown Trout in a Rich Limestone Stream? Water 2021, 13, 2492. https://doi.org/10.3390/w13182492
Čanak Atlagić J, Marić A, Tubić B, Andjus S, Đuknić J, Marković V, Paunović M, Simonović P. What’s on the Menu for the Resident Brown Trout in a Rich Limestone Stream? Water. 2021; 13(18):2492. https://doi.org/10.3390/w13182492
Chicago/Turabian StyleČanak Atlagić, Jelena, Ana Marić, Bojana Tubić, Stefan Andjus, Jelena Đuknić, Vanja Marković, Momir Paunović, and Predrag Simonović. 2021. "What’s on the Menu for the Resident Brown Trout in a Rich Limestone Stream?" Water 13, no. 18: 2492. https://doi.org/10.3390/w13182492