
Ecological Influences of Water-Level Fluctuation on Food Web Network


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
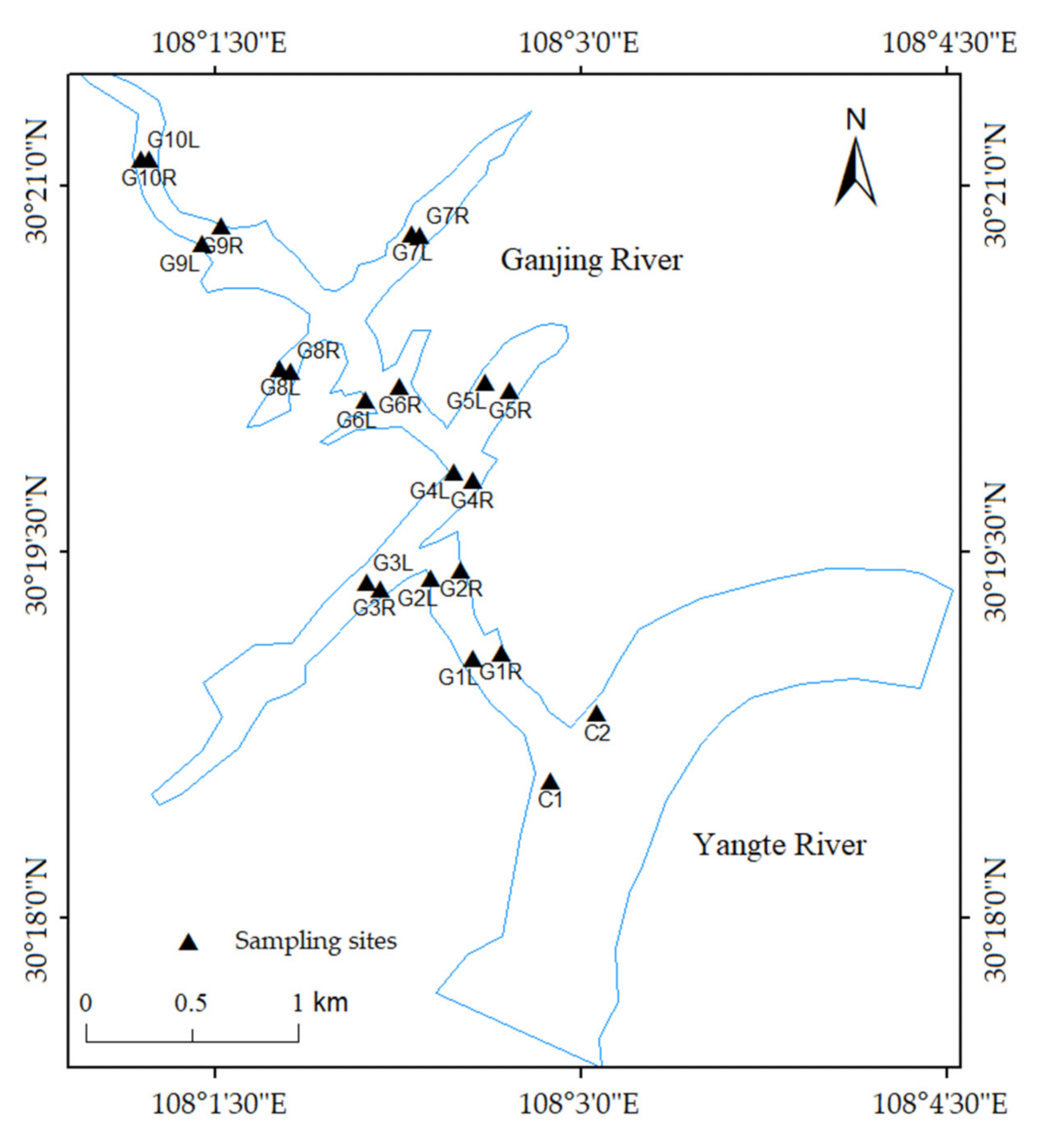
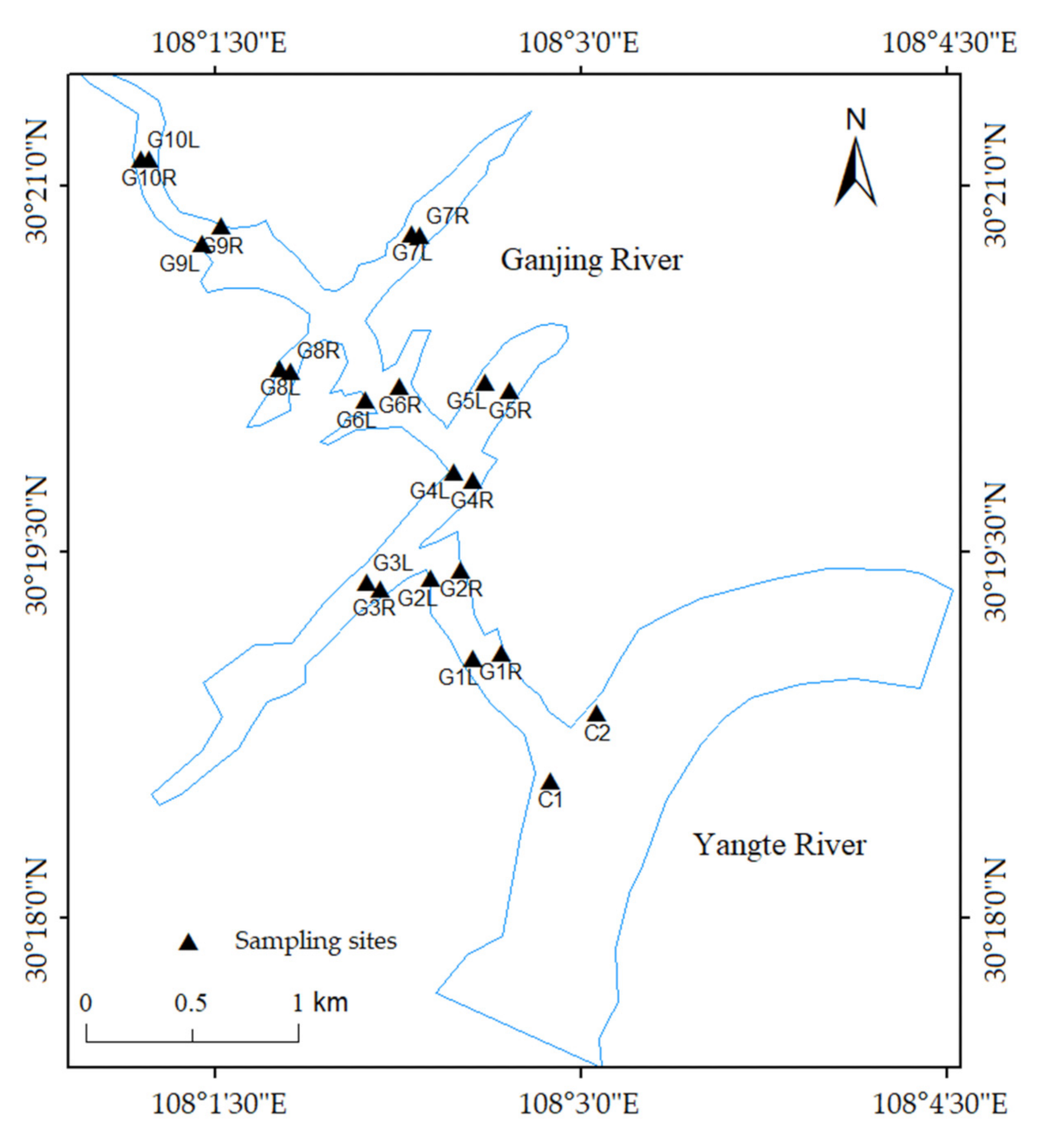
2.1. Study Area
2.2. Sample Collection
2.3. Analysis of Stable Isotopes
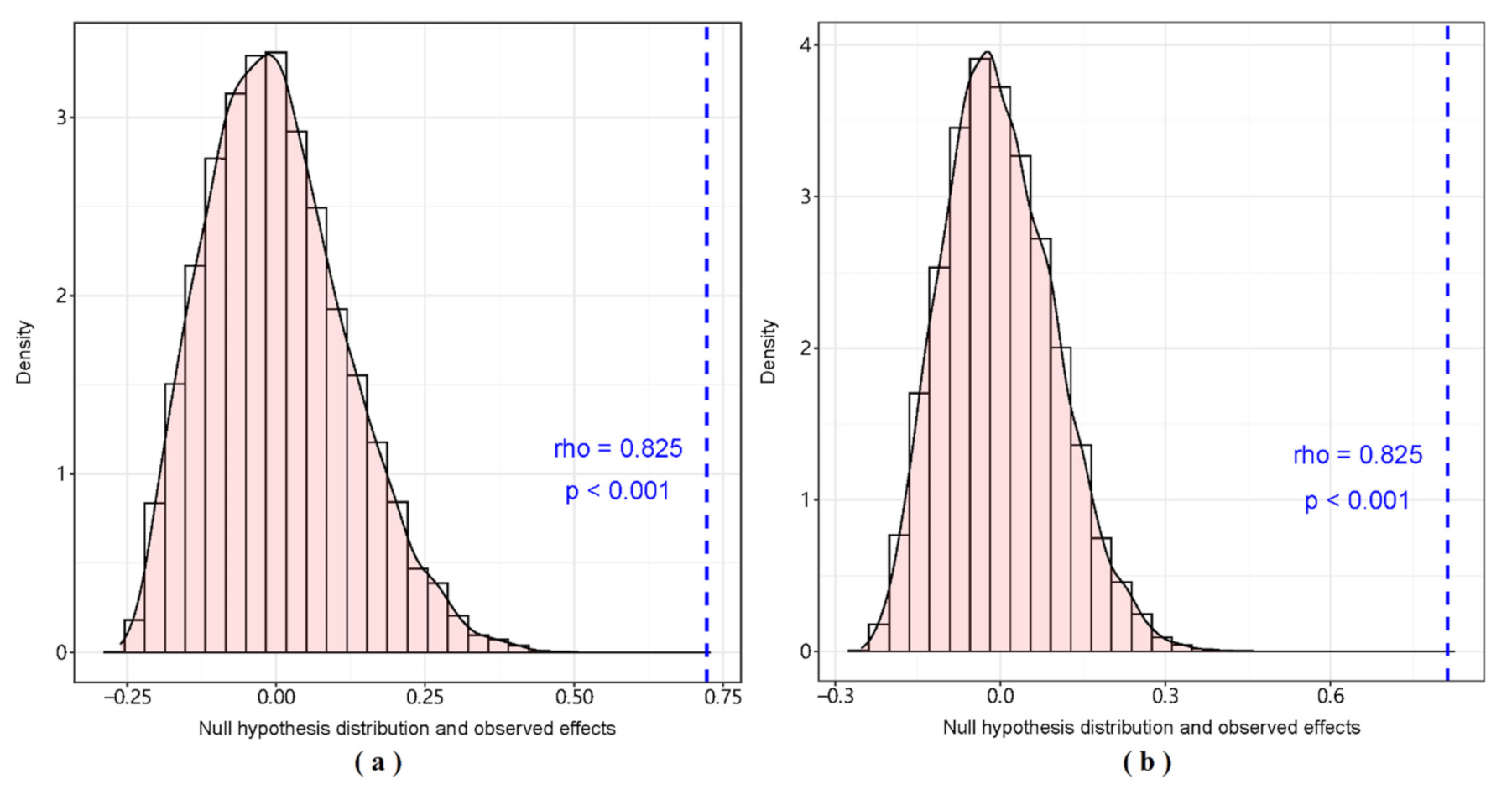
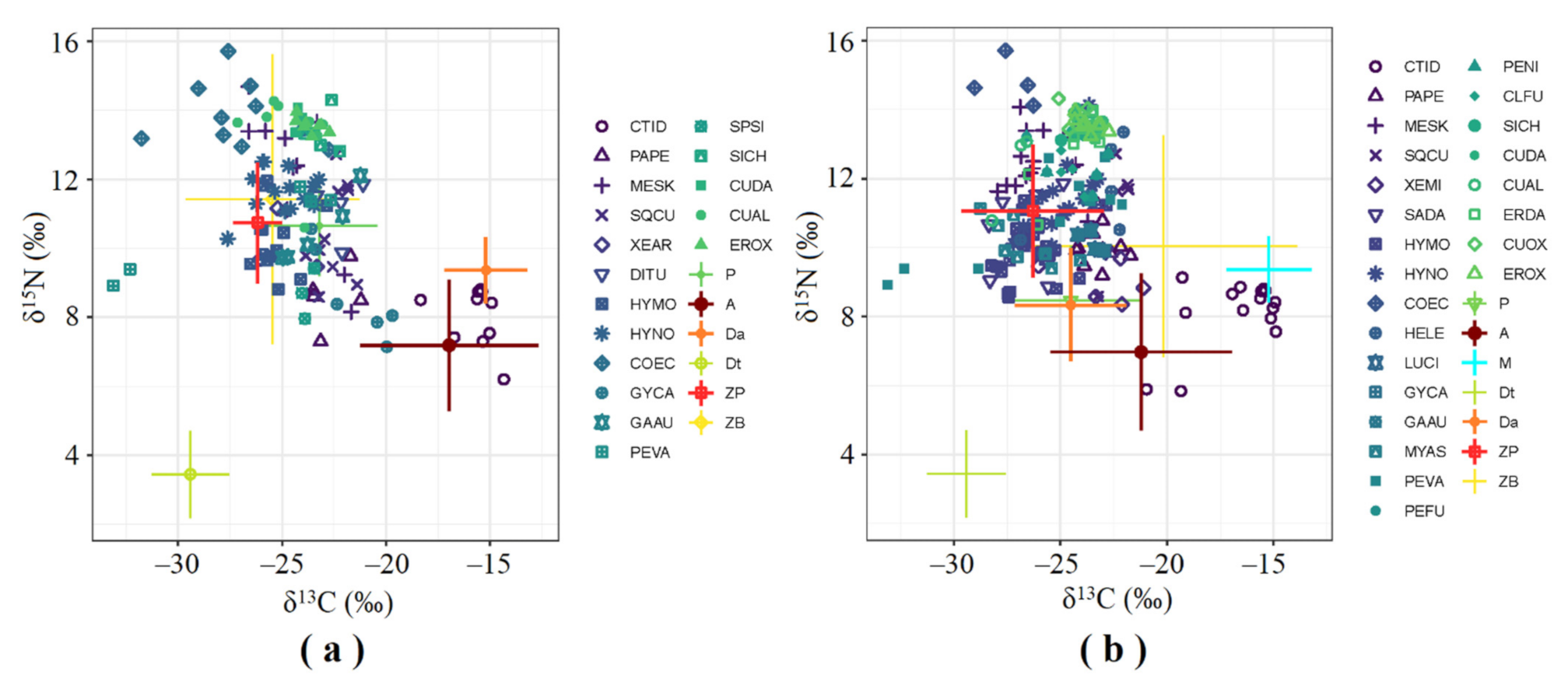
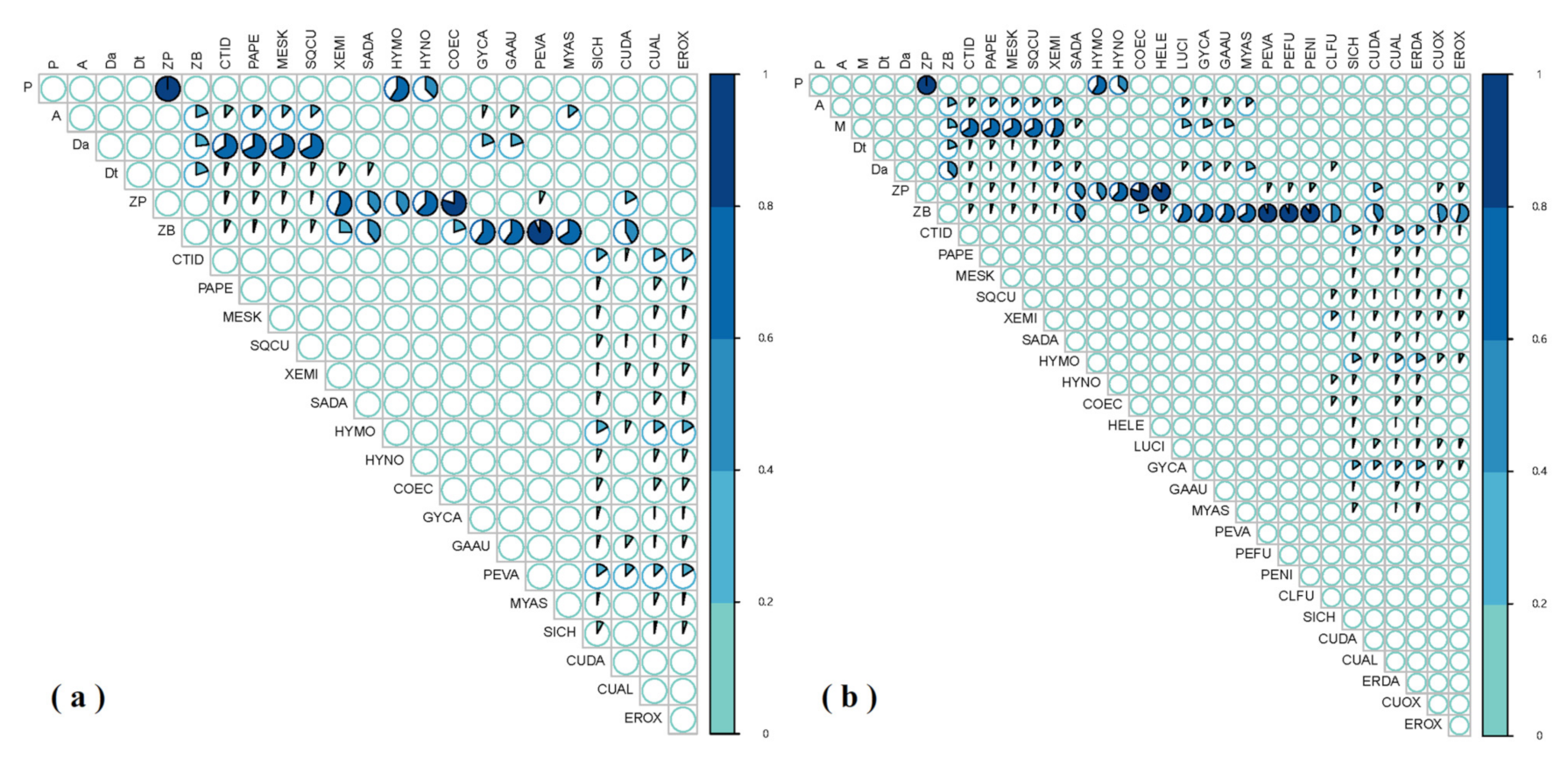
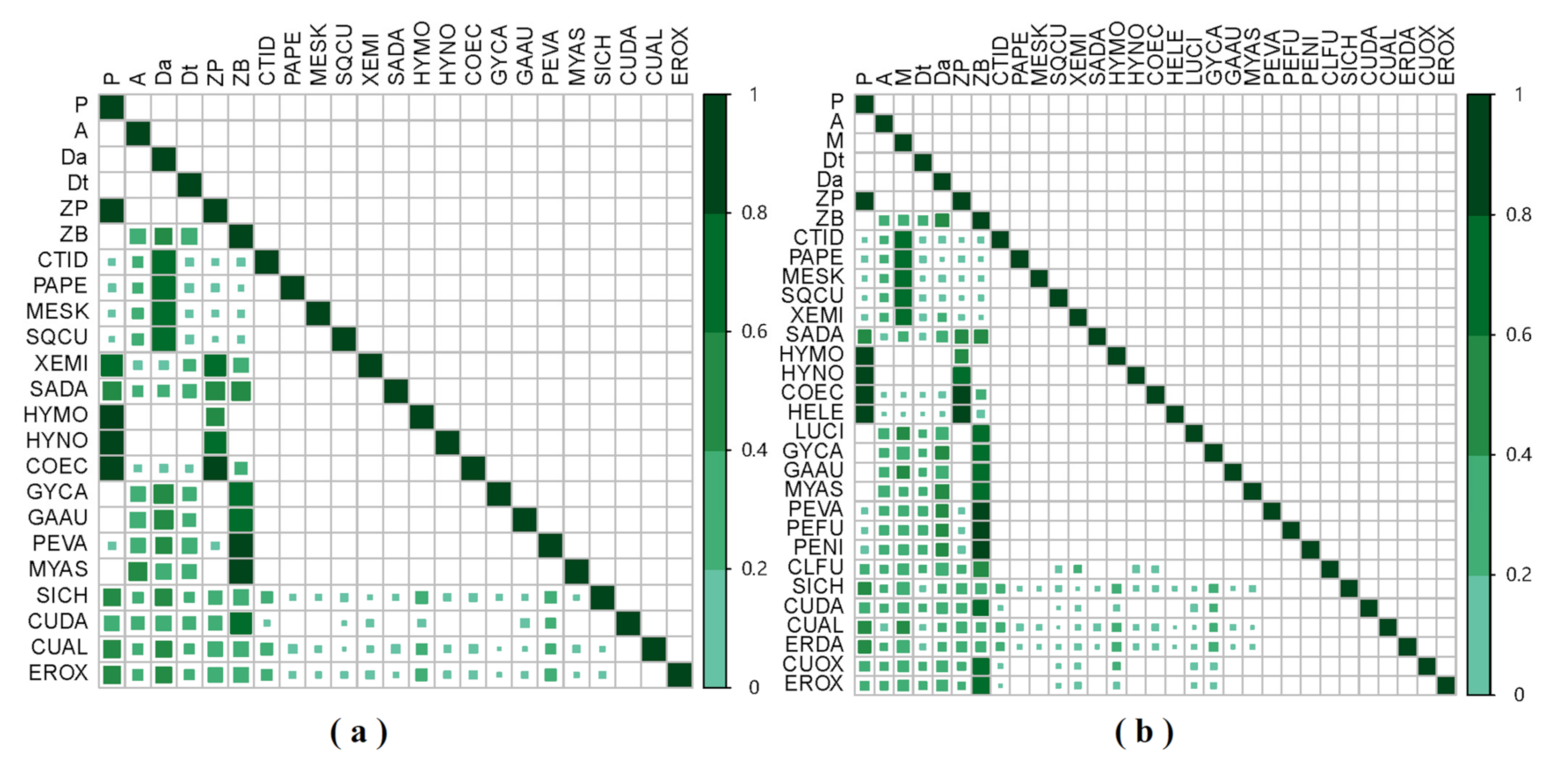
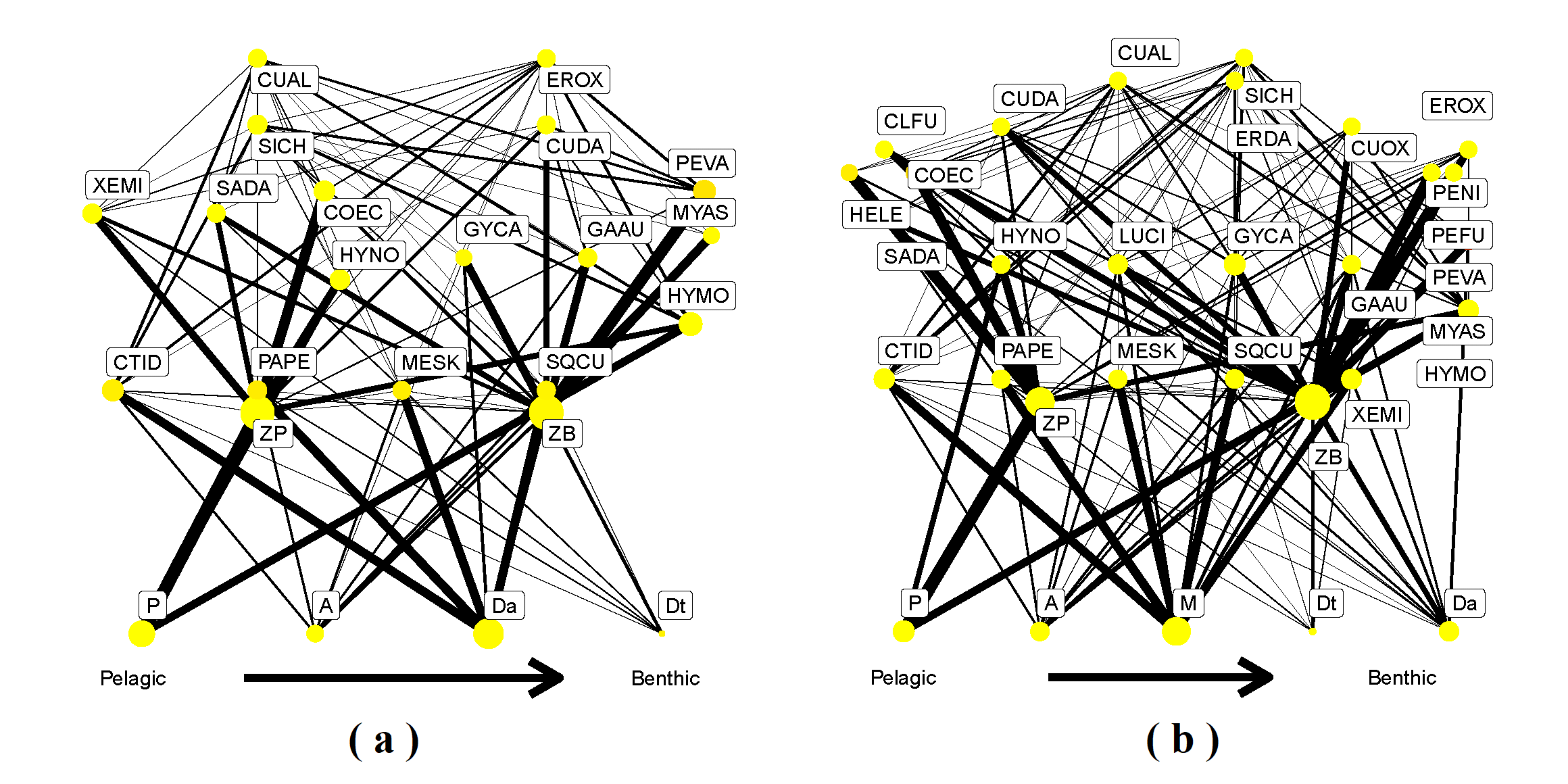
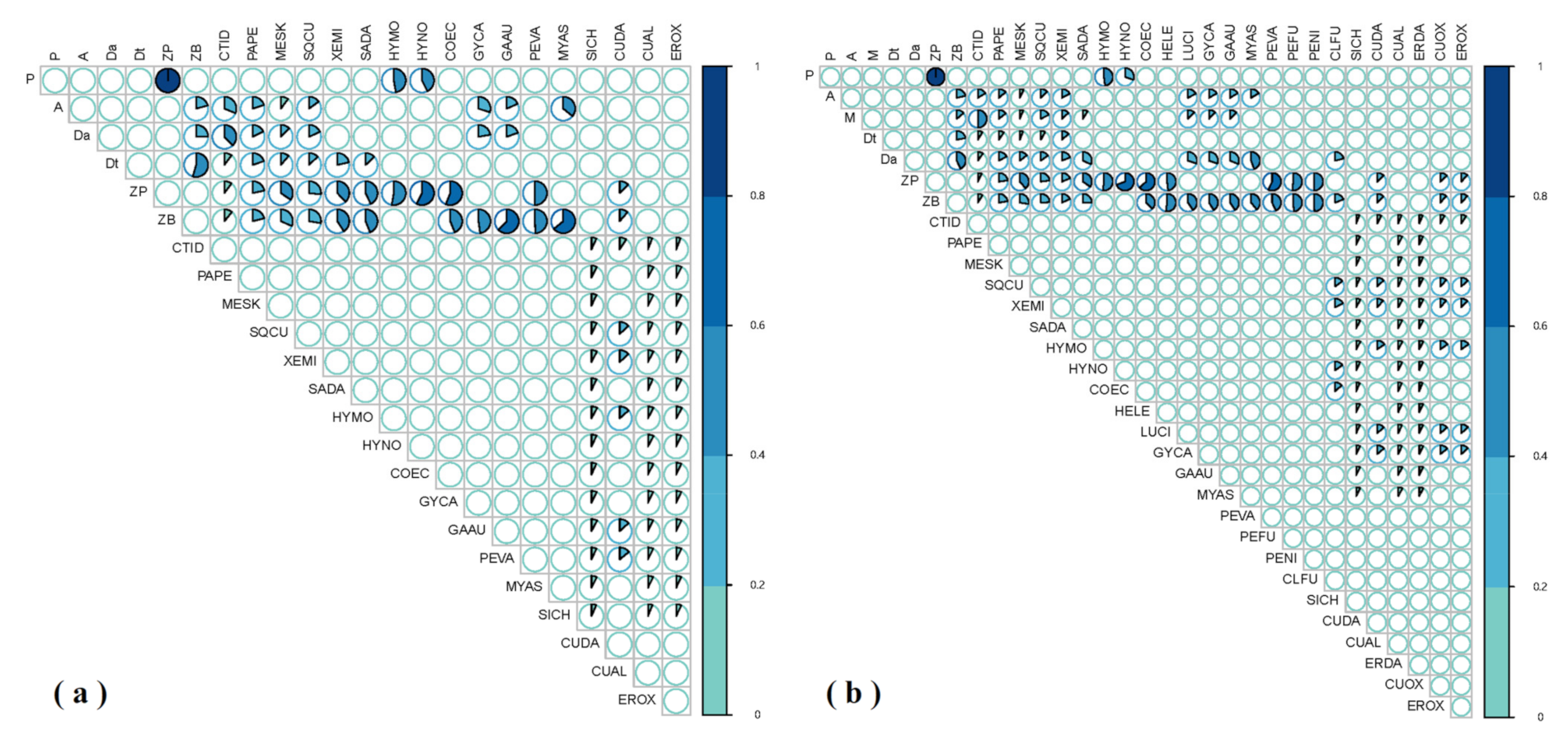
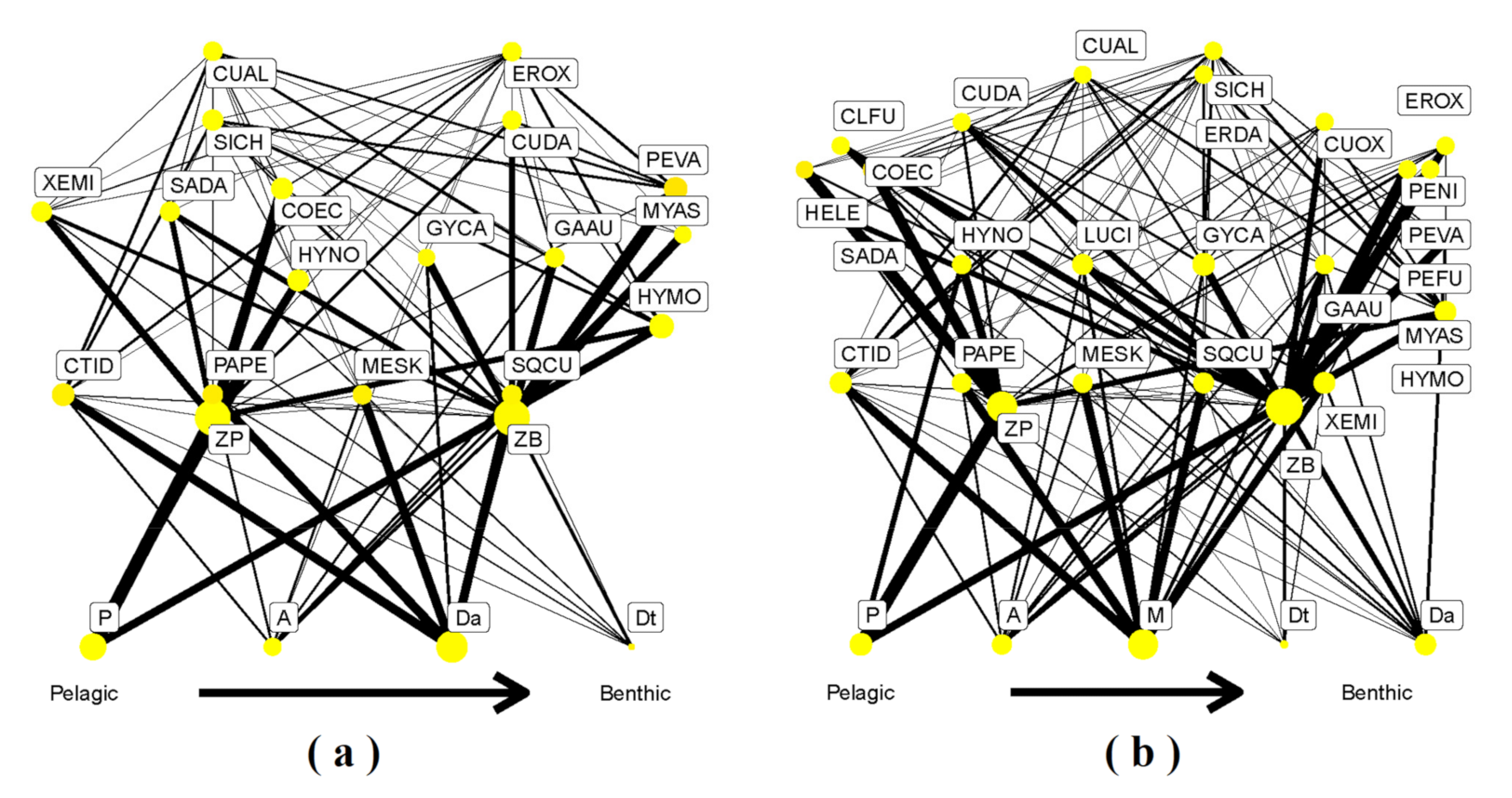
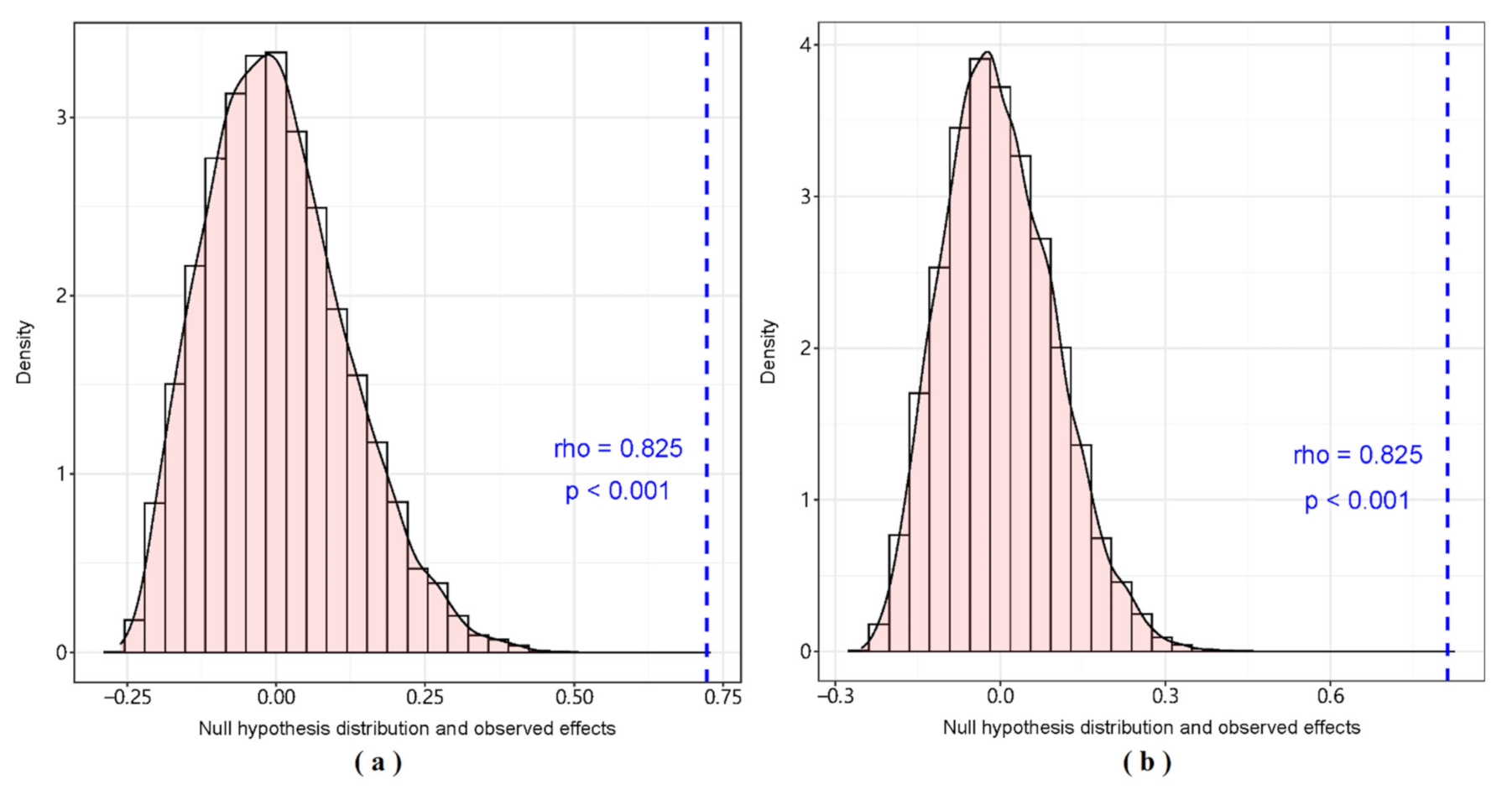
3. Results
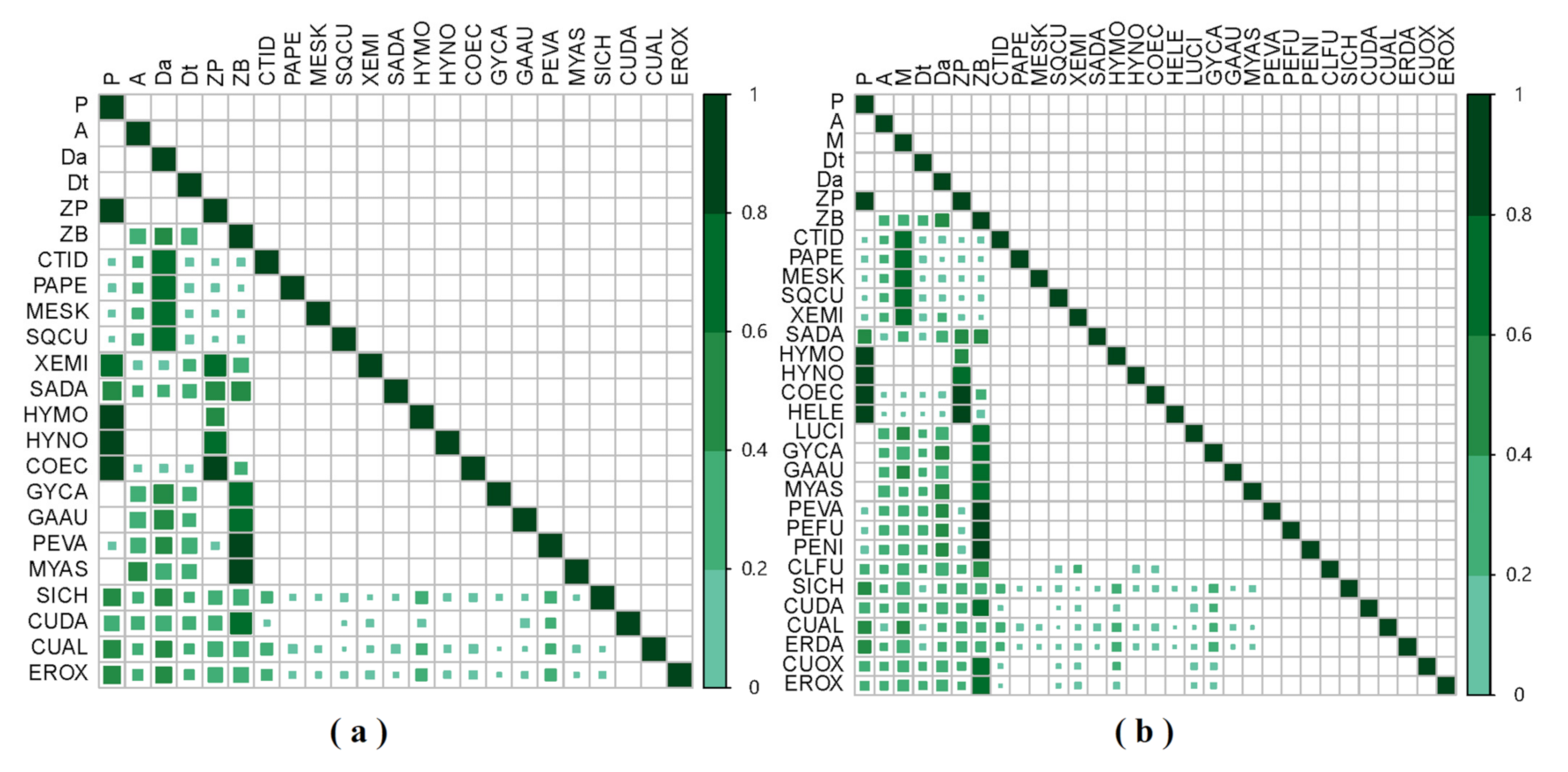
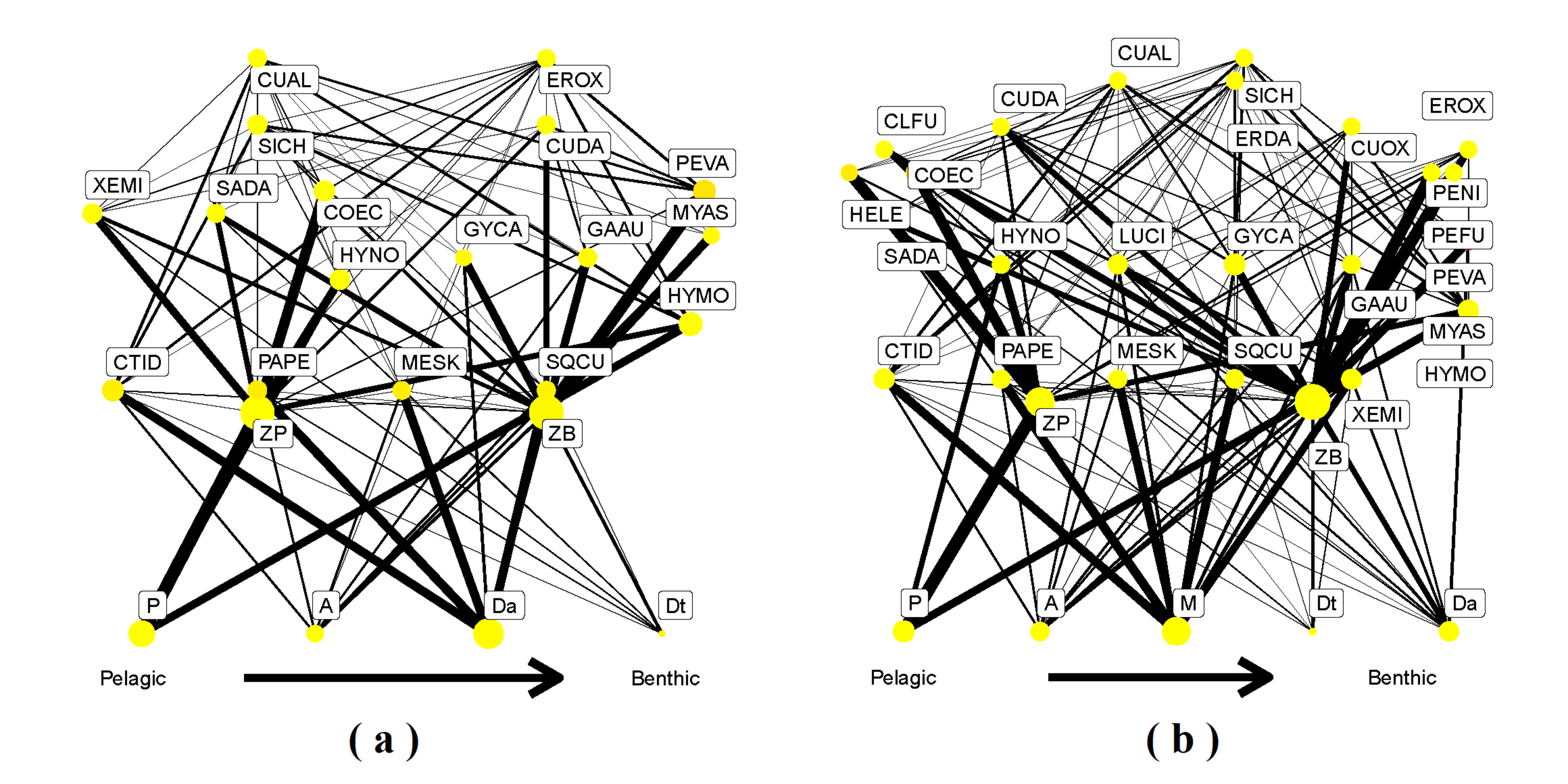
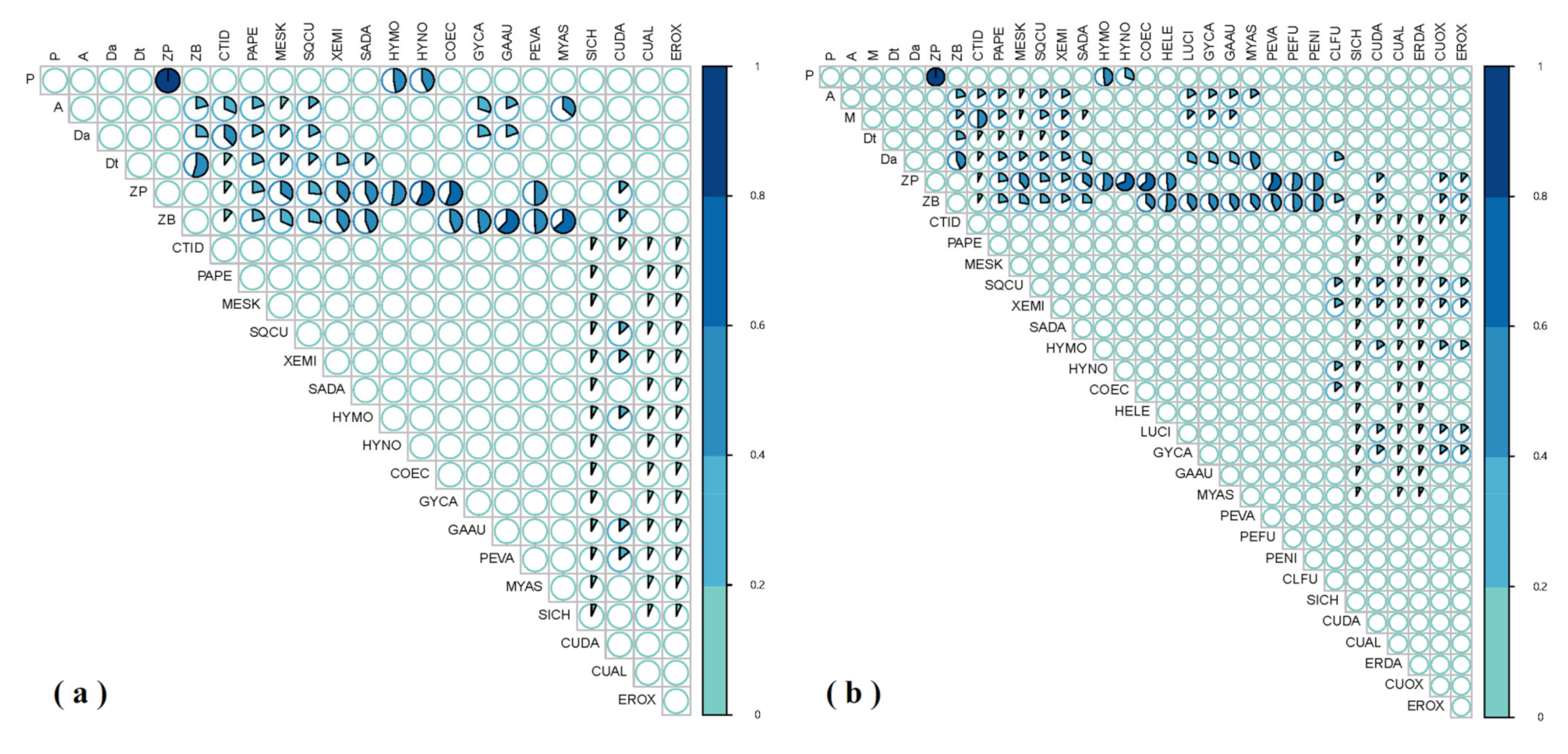
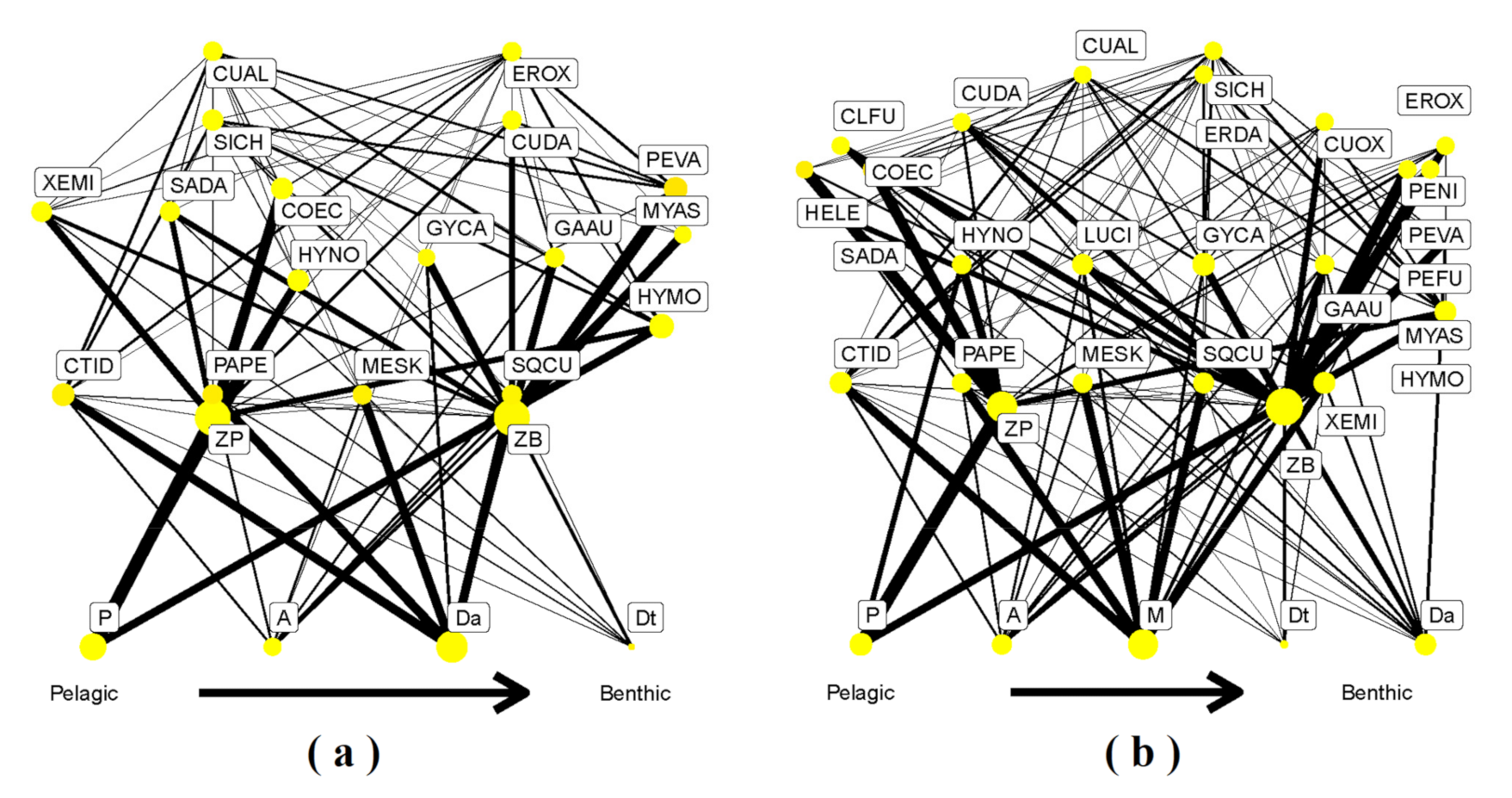
3.1. GCA Food Web
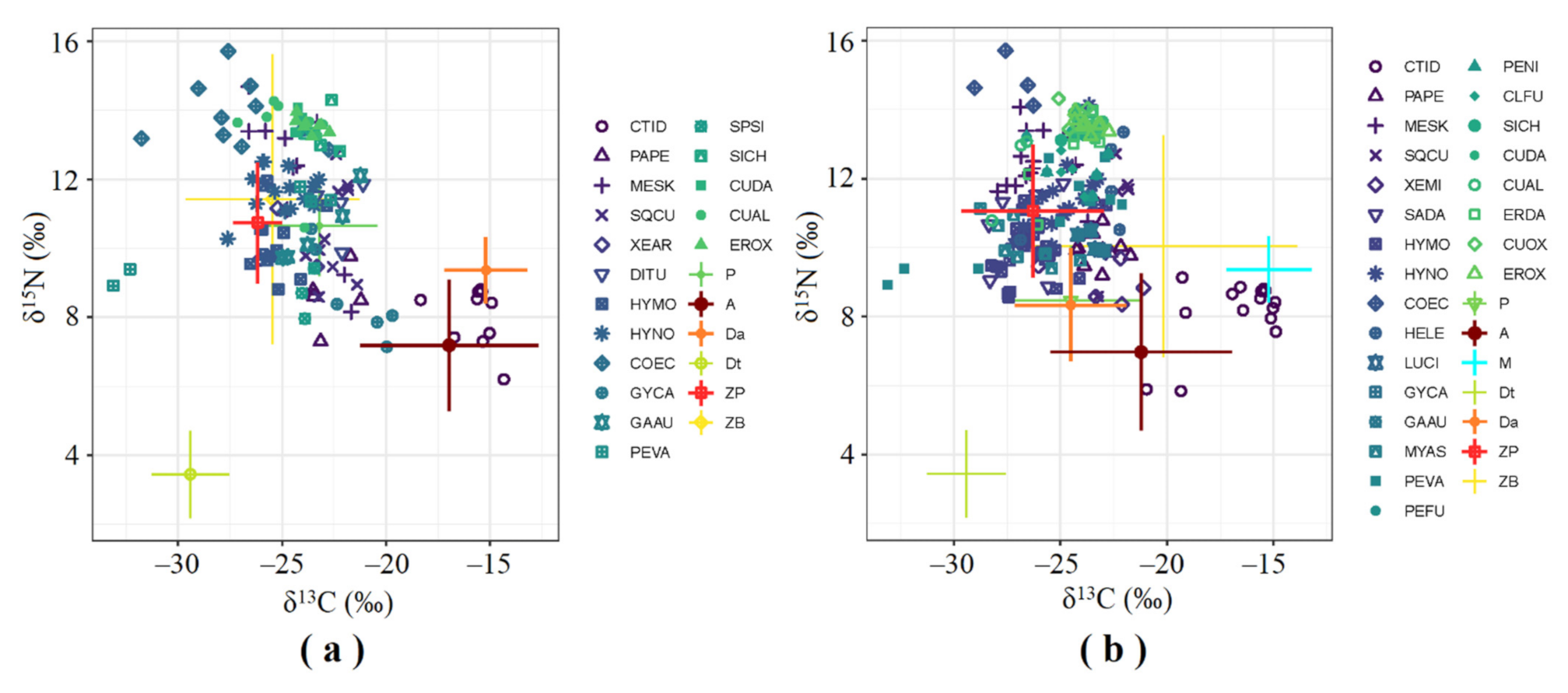
3.2. SIA Food Web
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Naselli-Flores, L.; Barone, R. Importance of water-level fluctuation on population dynamics of cladocerans in a hypertrophic reservoir (Lake Arancio, south-west Sicily, Italy). Hydrobiologia 1997, 360, 223–232. [Google Scholar] [CrossRef]
- Wantzen, K.M.; Rothhaupt, K.-O.; Mörtl, M.; Cantonati, M.; G.-Tóth, L.; Fischer, P. Ecological effects of water-level fluctuations in lakes: An urgent issue. Hydrobiologia 2008, 613, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Wu, J.; Chen, J.; Wu, Q.; Lei, G. Freshwater fish biodiversity in the Yangtze River basin of China: Patterns, threats and conservation. Biodivers. Conserv. 2003, 12, 1649–1685. [Google Scholar] [CrossRef]
- Reidy, L.C.; Christer, N.; James, R.; Ng, R.Y. Implications of dam obstruction for global freshwater fish diversity. Bioscience 2012, 62, 539–548. [Google Scholar]
- Mao, Z.; Gu, X.; Cao, Y.; Luo, J.; Jeppesen, E. Pelagic energy flow supports the food web of a shallow lake following a dramatic regime shift driven by water level changes. Sci. Total Environ. 2020, 756, 143642. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, X.; Li, W.; Xu, J.; Chen, Y.; Fan, N. Potential influence of water level changes on energy flows in a lake food web. Chin. Sci. Bull. 2011, 56, 2794–2802. [Google Scholar] [CrossRef] [Green Version]
- Atwood, T.B.; Wiegner, T.N.; MacKenzie, R.A. Effects of hydrological forcing on the structure of a tropical estuarine food web. Oikos 2012, 121, 277–289. [Google Scholar] [CrossRef]
- Liu, J.K. Lakes of the middle and lower basins of the Chang Jiang (China). In Lakes and Reservoirs; Elsevier: Amsterdam, The Netherlands, 1984. [Google Scholar]
- Wantzen, K.M.; Machado, F.D.A.; Voss, M.; Boriss, H.; Junk, W.J. Seasonal isotopic shifts in fish of the Pantanal wetland, Brazil. Aquat. Sci. 2002, 64, 239–251. [Google Scholar] [CrossRef]
- Post, D.M. Using Stable Isotopes to Estimate Trophic Position: Models, Methods, and Assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Xu, J.; Wen, Z.; Ke, Z.; Zhang, M.; Ping, X. Contrasting energy pathways at the community level as a consequence of regime shifts. Oecologia 2014, 175, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Compson, Z.G.; Monk, W.A.; Hayden, B.; Bush, A.; O’Malley, Z.; Hajibabaei, M.; Porter, T.M.; Wright, M.T.; Baker, C.J.; Manir, M.S.A.; et al. Network-Based Biomonitoring: Exploring Freshwater Food Webs With Stable Isotope Analysis and DNA Metabarcoding. Front. Ecol. Evol. 2019, 7, 395. [Google Scholar] [CrossRef] [Green Version]
- Kaymak, N.; Winemiller, K.O.; Akin, S.; Altuner, Z.; Polat, F.; Dal, T. Stable isotope analysis reveals relative influences of seasonal hydrologic variation and impoundment on assimilation of primary production sources by fish in the Upper Yesilırmak River, Turkey. Hydrobiologia 2015, 753, 131–147. [Google Scholar] [CrossRef]
- Hopkins, J.B.; Ferguson, J.M.; Lyle, K. Estimating the Diets of Animals Using Stable Isotopes and a Comprehensive Bayesian Mixing Model. PLoS ONE 2012, 7, e28478. [Google Scholar] [CrossRef]
- Kadoya, T.; Osada, Y.; Takimoto, G. IsoWeb: A Bayesian Isotope Mixing Model for Diet Analysis of the Whole Food Web. PLoS ONE 2012, 7, e41057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erhardt, E.B.; Bedrick, E.J. A Bayesian framework for stable isotope mixing models. Environ. Ecol. Stat. 2013, 19, 377–397. [Google Scholar] [CrossRef]
- Bao, Y.; Gao, P.; He, X. The water-level fluctuation zone of Three Gorges Reservoir—A unique geomorphological unit. Earth Sci. Rev. 2015, 150, 14–24. [Google Scholar] [CrossRef]
- Zhang, Q.; Lou, Z. The environmental changes and mitigation actions in the Three Gorges Reservoir region, China. Environ. Sci. Policy 2011, 14, 1132–1138. [Google Scholar] [CrossRef]
- Guo, W.X.; Wang, H.X.; Jian-Xin, X.U.; Xia, Z.Q. Ecological operation for Three Gorges Reservoir. Water Sci. Eng. 2011, 4, 143–156. [Google Scholar] [CrossRef]
- Xiang, R.; Wang, L.; Li, H.; Tian, Z.; Zheng, B. Temporal and spatial variation in water quality in the Three Gorges Reservoir from 1998 to 2018. Sci. Total Environ. 2020, 768, 144866. [Google Scholar] [CrossRef]
- Ye, J. Studies in Community Composition and Feeding Ecology of Fish in the Littoral Zone of Mayliang Bay, Taihu Lake. Master Thesis, Huazhong Agricultural University, Wuhan, China, 2006. [Google Scholar]
- Ma, X.; Liu, C.; Liu, L.; Wang, Y.; Wang, J.; Zhang, Y. Study on the food web of fish in Baiyangdian lake based diet analysis. J. Hydroecol. 2011, 32, 85–90. [Google Scholar]
- Ding, R.H. The Fishes of Sichuan, China; Ding, R.H., Ed.; Sichuan Publishing House of Science and Technology: Chengdu, China, 1994. [Google Scholar]
- Eller, G.; Deines, P.; Grey, J.; Richnow, H.-H.; Krüger, M. Methane cycling in lake sediments and its influence on chironomid larval δ13C. FEMS Microbiol. Ecol. 2005, 54, 339–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, S.K.; Lewis, W.M., Jr. Stable carbon and nitrogen isotopes in algae and detritus from the Orinoco River floodplain, Venezuela. Geochim. Cosmochim. Acta 1992, 56, 4237–4246. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Y.; Wang, K.; Zeng, Q.; Zhang, M.; Zhang, H. Protocols for sample collection, pretreatment and preservation of aquatic organisms in stable isotope ecology. Acta Hydrobiol. Sin. 2020, 44, 989–997. [Google Scholar] [CrossRef]
- Kones, J.K.; Soetaert, K.; Oevelen, D.V.; Owino, J.O. Are network indices robust indicators of food web functioning? A Monte Carlo approach. Ecol. Model. 2009, 220, 370–382. [Google Scholar] [CrossRef]
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and Flow Regulation of the World’s Large River Systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Best, J. Anthropogenic stresses on the world’s big rivers. Nat. Geosci. 2018, 12, 7–21. [Google Scholar] [CrossRef]
- Perga, M.-E.; Arfi, R.; Gerdeaux, D. Seasonal variations in fish δ13C and δ15N in two West African reservoirs, Sélingué and Manantali (Mali): Modifications of trophic links in relation to water level. Isot. Environ. Health Stud. 2005, 41, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Winemiller, K.O.; Jepsen, D.B. Effects of seasonality and fish movement on tropical river food webs. J. Fish Biol. 1998, 53, 267–296. [Google Scholar] [CrossRef]
- Jepsen, D.B.; Winemiller, K.O. Structure of tropical river food webs revealed by stable isotope ratios. Oikos 2002, 96, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, J.G.; Kottelat, M.; Smith, G.R.; Gill, S.A.C. So Many Fishes, So Little Time: An Overview of Recent Ichthyological Discovery in Continental Waters. Ann. Mo. Bot. Gard. 2000, 87, 26–62. [Google Scholar] [CrossRef]
- Leira, M.; Cantonati, M. Effects of water-level fluctuations on lakes: An annotated bibliography. Hydrobiologia 2008, 613, 171–184. [Google Scholar] [CrossRef]
- Cremona, F.; Timm, H.; Agasild, H.; Tõnno, I.; Feldmann, T.; Jones, R.I.; Nõges, T. Benthic foodweb structure in a large shallow lake studied by stable isotope analysis. Freshw. Sci. 2014, 33, 885–894. [Google Scholar] [CrossRef] [Green Version]
- Roach, K.A.; Winemiller, K.O.; Layman, C.A.; Zeug, S.C. Consistent trophic patterns among fishes in lagoon and channel habitats of a tropical floodplain river: Evidence from stable isotopes. Acta Oecologica 2009, 35, 513–522. [Google Scholar] [CrossRef]
- Bartels, P.; Cucherousset, J.; Gudasz, C.; Jansson, M.; Karlsson, J.; Persson, L.; Premke, K.; Rubach, A.; Steger, K.; Tranvik, L.J.; et al. Terrestrial subsidies to lake food webs: An experimental approach. Oecologia 2012, 168, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Scharnweber, K.; Syväranta, J.; Hilt, S.; Brauns, M.; Vanni, M.J.; Brothers, S.; Köhler, J.; Knežević-Jarić, J.; Mehner, T. Whole-lake experiments reveal the fate of terrestrial particulate organic carbon in benthic food webs of shallow lakes. Ecology 2014, 95, 1496–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumann, R.; Hammer, A.; Görs, S.; Schubert, H. Winter and spring phytoplankton composition and production in a shallow eutrophic Baltic lagoon. Estuar. Coast. Shelf Sci. 2005, 62, 169–181. [Google Scholar] [CrossRef]
- Paine, R.T. Food Webs: Linkage, Interaction Strength and Community Infrastructure. J. Anim. Ecol. 1980, 49, 666–685. [Google Scholar] [CrossRef]
- Thorp, J.H.; Delong, M.D.; Greenwood, K.S.; Casper, A.F. Isotopic analysis of three food web theories in constricted and floodplain regions of a large river. Oecologia 1998, 117, 551–563. [Google Scholar] [CrossRef]
- Anderson, C.; Cabana, G. Estimating the trophic position of aquatic consumers in river food webs using stable nitrogen isotopes. J. N. Am. Benthol. Soc. 2007, 26, 273–285. [Google Scholar] [CrossRef]
- Lauzanne, L. Régimes alimentaires et relations trophiques des poissons du Lac Tchad. Cahiers ORSTOM Série Hydrobiologie 1976, 10, 267–310. [Google Scholar]
- Paugy, D.; Bénech, V. Les poissons d’eau douce des bassins côtiers du Togo (Afrique de l’Ouest). Revue d’Hydrobiologie Tropicale 1989, 22, 295–316. [Google Scholar]
- McCann, K.; Hastings, A. Re-Evaluating the omnivory-stability relationship in food webs. Proc. Biol. Sci. 1997, 264, 1249–1254. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Nes, E.H.V. Shallow lakes theory revisited: Various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 2007, 584, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Ramstack Hobbs, J.M.; Hobbs, W.O.; Edlund, M.B.; Zimmer, K.D.; Theissen, K.M.; Hoidal, N.; Domine, L.M.; Hanson, M.A.; Herwig, B.R.; Cotner, J.B. The legacy of large regime shifts in shallow lakes. Ecol. Appl. 2016, 26, 2660–2674. [Google Scholar] [CrossRef] [PubMed]
- Lacoul, P.; Freedman, B. Environmental influences on aquatic plants in freshwater ecosystems. Environ. Rev. 2006, 14, 89–136. [Google Scholar] [CrossRef]
- Abrantes, K.G.; Sheaves, M. Importance of freshwater flow in terrestrial–aquatic energetic connectivity in intermittently connected estuaries of tropical Australia. Mar. Biol. 2010, 157, 2071–2086. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Rooney, N.; McCann, K.S. Integrating food web diversity, structure and stability. Trends Ecol. Evol. 2012, 27, 40–46. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Que, Y.; Xie, J.; Xu, J.; Li, W.; Wang, E.; Zhu, B. Ecological Influences of Water-Level Fluctuation on Food Web Network. Water 2021, 13, 2371. https://doi.org/10.3390/w13172371
Que Y, Xie J, Xu J, Li W, Wang E, Zhu B. Ecological Influences of Water-Level Fluctuation on Food Web Network. Water. 2021; 13(17):2371. https://doi.org/10.3390/w13172371
Chicago/Turabian StyleQue, Yanfu, Jiayi Xie, Jun Xu, Weitao Li, Ezhou Wang, and Bin Zhu. 2021. "Ecological Influences of Water-Level Fluctuation on Food Web Network" Water 13, no. 17: 2371. https://doi.org/10.3390/w13172371